
Pectolinarin Inhibits the Bacterial Biofilm Formation and Thereby Reduces Bacterial Pathogenicity


Abstract
:1. Introduction
2. Results
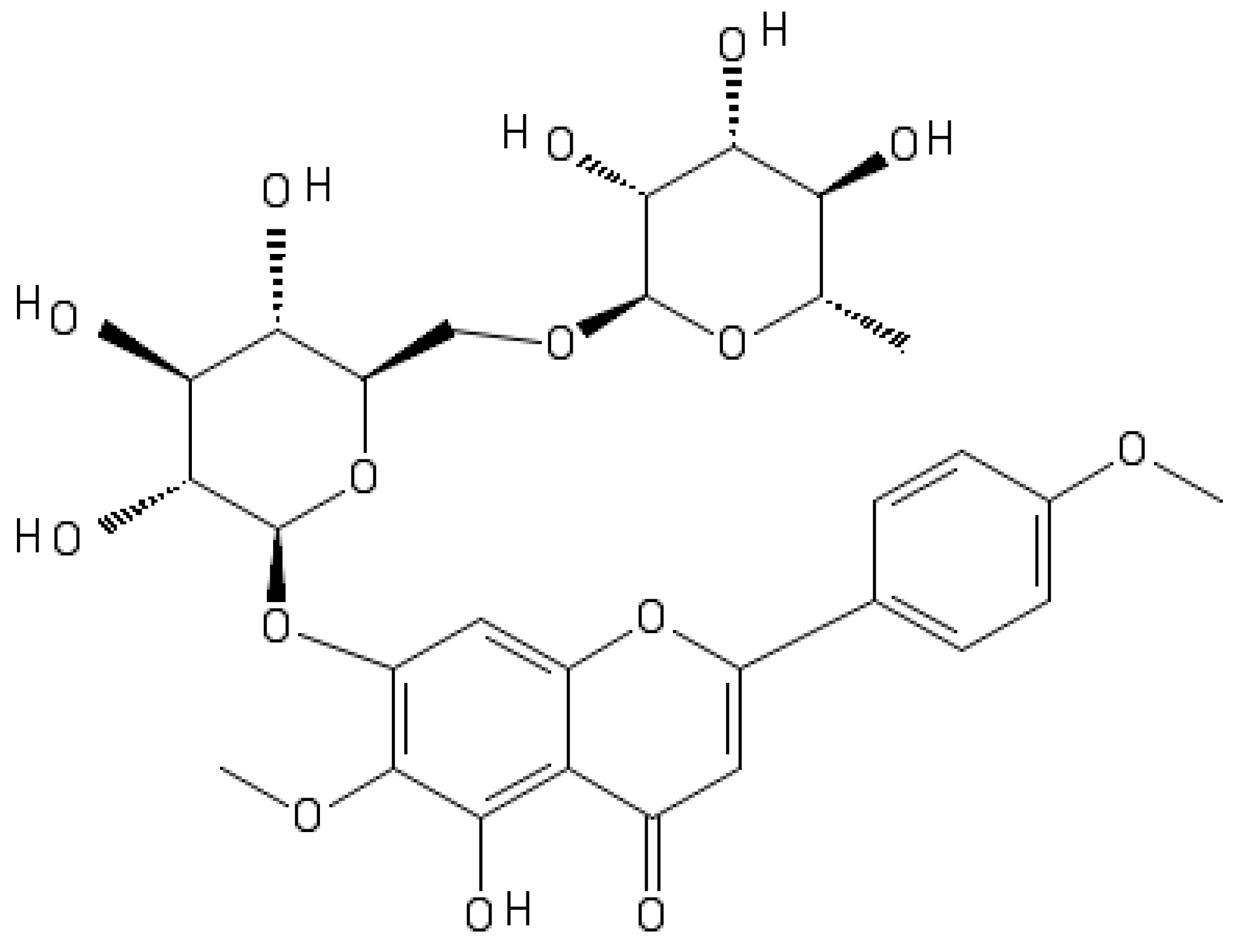
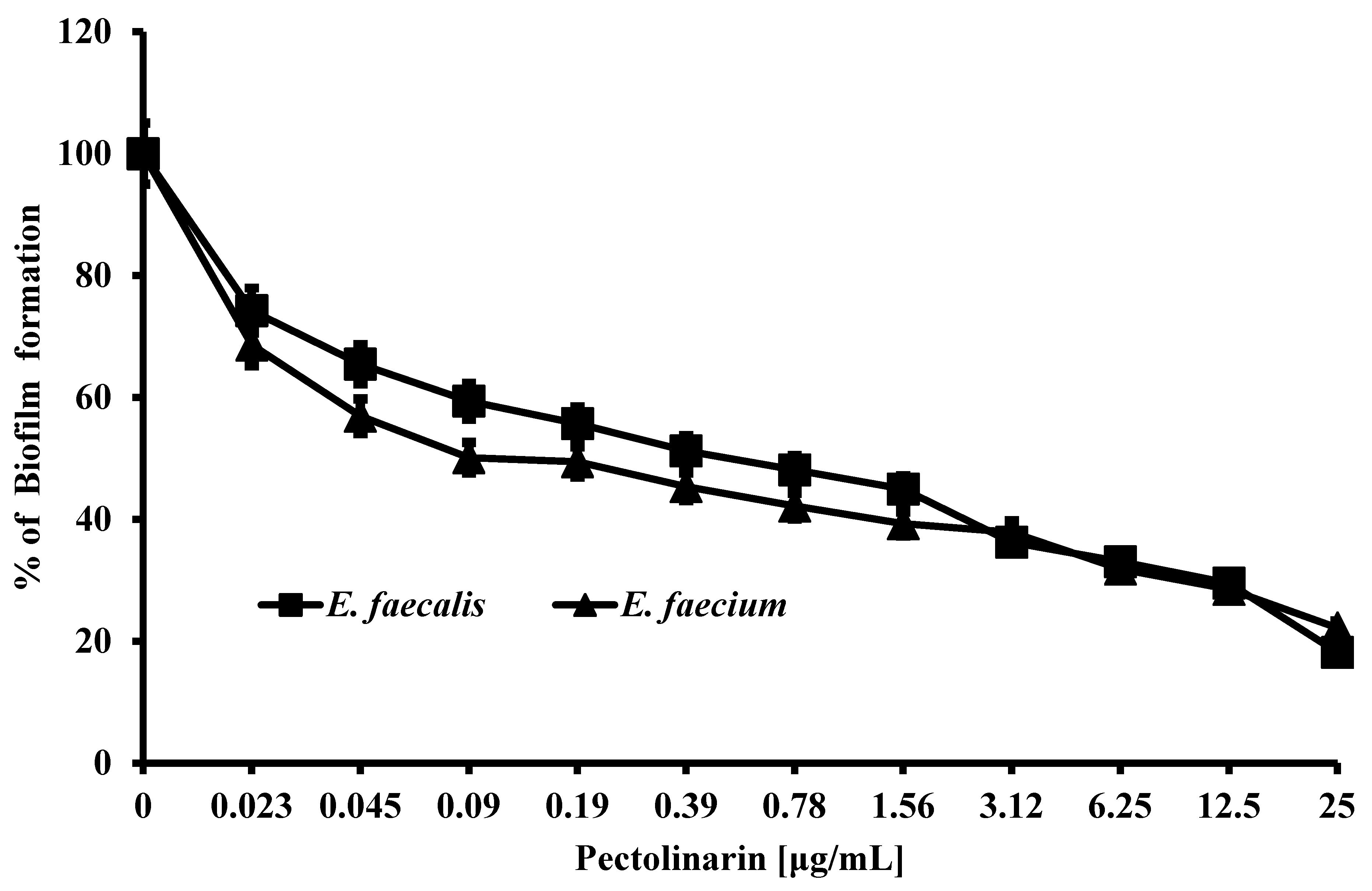
2.1. Pectolinarin Inhibited the Biofilm Formation of Bacteria
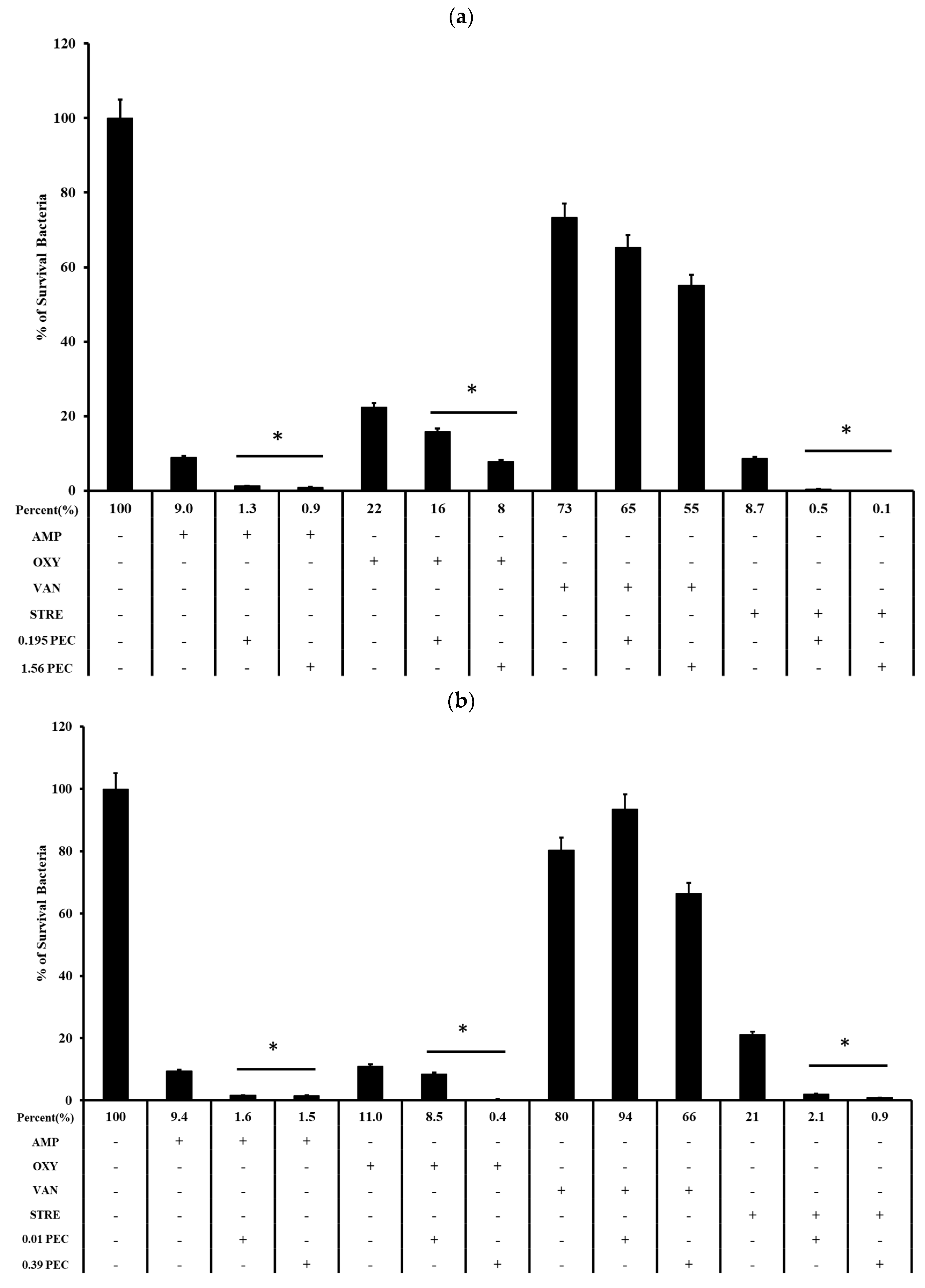
2.2. Pectolinarin Increased the Susceptibility of E. faecalis and E. faecium to Commercialized Antibiotics
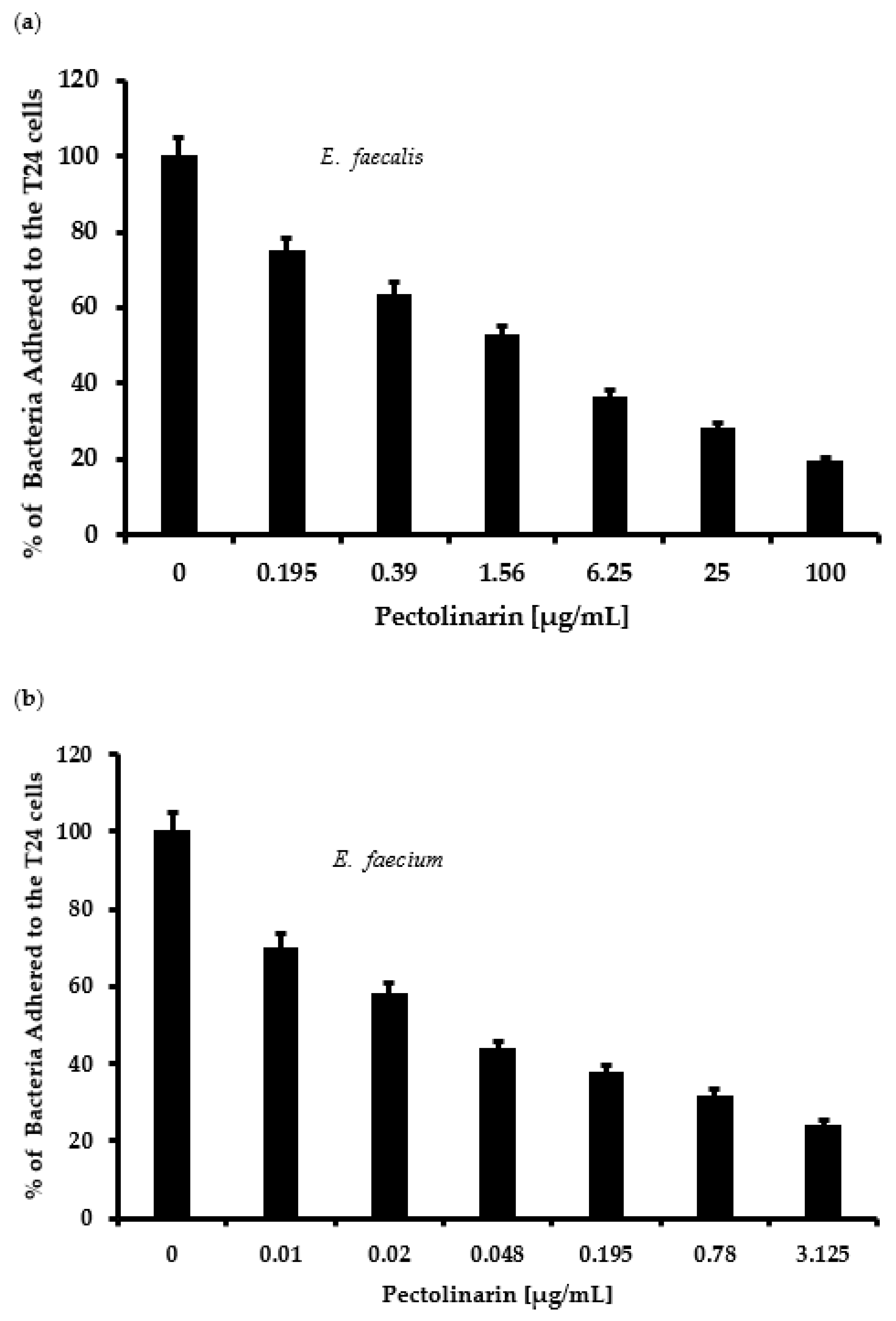
2.3. Pectolinarin Reduced Bacterial Adherence to T24 Cells in a Dose-Dependent Manner
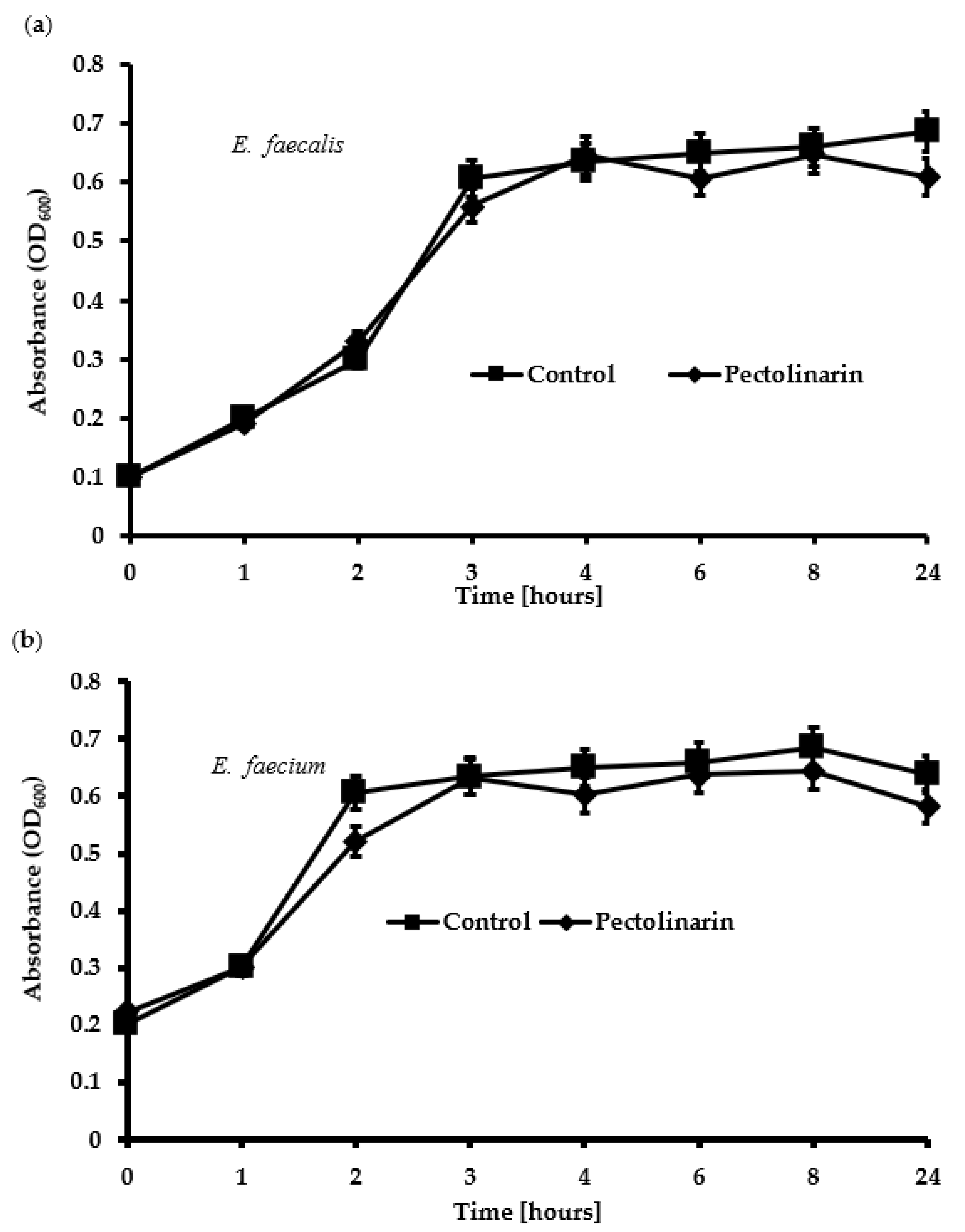
2.4. Pectolinarin Did Not Affect Bacterial Growth
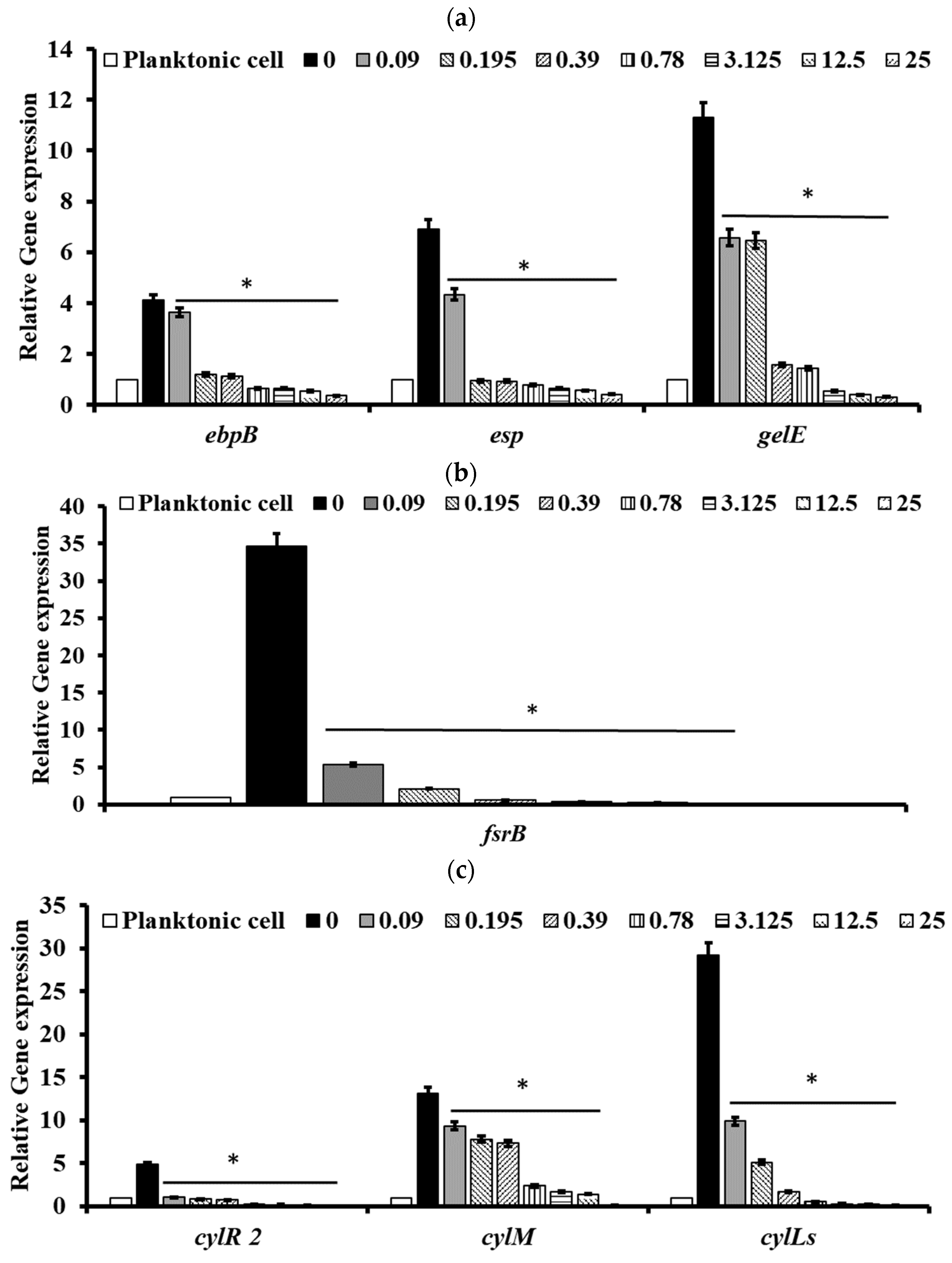
2.5. Pectolinarin Inhibited the Expression of Genes Related to the Biofilm Formation and Virulence of Bacteria
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Biofilm Formation Assay
4.3. The Combinatorial Antibacterial Effects of Pectolinarin with Commercialized Antibiotics
4.4. Bacterial Adherence Assays
4.5. Time Kill Assay
4.6. Quantitative RT-PCR Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Donlan, R.M.; Costerton, J.W. Biofilm: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Mah, T.F.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Tomita, H.; Ike, Y. Tissue-specific Adherent Enterococcus faecalis stains that show highly efficient adhesion to human bladder carcinoma T24 cells also adhere to extracellular matrix proteins. Infect. Immun. 2004, 72, 5877–5885. [Google Scholar] [CrossRef] [Green Version]
- Guenther, F.; Stroh, P.; Wagner, C.; Obst, U.; Hansch, G.M. Phagocytosis of staphylococci biofilms by polymorph nuclear neutrophils: S. aureus and S. epidermidis differ with regard to their susceptibility towards the host defense. Int. J. Artif. Organs 2009, 32, 565–573. [Google Scholar] [CrossRef]
- Strickertsson, J.A.B.; Desler, C.; Bertelsen, J.M.; Machado, A.M.D.; Wadstrom, T.; Winther, O.; Rasmussen, L.J.; Hansen, L.F. Enterococcus faecalis infection causes inflammation, intracellular oxphos-independent ROS production, and DNA damage in human gastric cancer cells. PLoS ONE 2013, 8, e63147. [Google Scholar] [CrossRef] [Green Version]
- Hidron, A.I.; Edwards, J.R.; Patel, J.; Horan, T.C.; Sievert, D.M.; Pollock, D.A.; Fridkin, S.K. Antimicrobial-resistant pathogens associated with healthcare-associated infection: Annual summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2006–2007. Infect. Control Hosp. Epidemiol. 2008, 29, 996–1011. [Google Scholar] [CrossRef] [Green Version]
- Arias, C.A.; Murray, B.E. The rise of the Enterococcus: Beyond vancomycin resistance. Nat. Rev. Microbiol. 2012, 10, 266–278. [Google Scholar] [CrossRef] [Green Version]
- Alejandro, T.A.; Valle, J.; Solano, C.; Arrizubieta, M.J.; Cucarella, C.; Lamata, M.; Amorena, B.; Leiva, J.; Penades, J.R.; Lasa, I. The entrococcal surface protein, Esp, is involved in Enterococcus faecalis biofilm formation. Appl. Environ. Microbiol. 2001, 67, 4538–4545. [Google Scholar]
- Lim, H.; Soo, K.H.; Chang, H.W.; Bae, K.; Kang, S.S.; Kim, H.P. Anti-inflammatory activity of pentolinarigenin and pectolinarin isolated form Cirsium chanroenicum. Biol. Pharm. Bull. 2008, 31, 2063–2067. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Jing, B.H.; King, S.M.; Chen, Y.C. Inhibition of cell growth and VEGF expression in ovarian cancer cells by flavonoids. Nutr. Cancer 2008, 60, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Top, J.; Paganelli, F.L.; Zhang, X.; Schaik, W.V.; Leavis, H.L.; Asbroek, M.V.L.; Poll, T.V.D.; Leendertse, M.; Bonten, M.J.M.; Willems, R.J.L. The Enterococcus faecium Enterococcal biofilm regulator, EbrB, regulates the esp operon and is implicated in biofilm formation and intestinal colonization. PLoS ONE 2013, 8, e65224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, C.; Tangjuan, L.; Ke, W.; Hou, C.; Cai, S.; Huang, Y.; Du, Z.; Huang, H.; Kong, J.; Chen, Y. Baicalein inhibits Staphylococcus aureus biofilm formation and the quorum sensing system in vitro. PLoS ONE 2016, 11, e0153469. [Google Scholar]
- Sreedhar, R.N.; Kavindra, V.S.; Jouko, S.; Danielle, A.G.; Magnus, H.; Stanley, L.E.; Barbara, E.M. Endocarditis and biofilm-associated pili of Enterococcus faecalis. J. Clin. Investig. 2006, 116, 2799–2807. [Google Scholar]
- Ken-ichi, A.M.; Kunitoshi, Y.; Yoshimitsu, M.; Tanaka, Y.; Ogura, T.; Sugimoto, S. Inhibitory effects of Myricetin derivatives on curli-dependent biofilm formation in Escherichia coli. Sci. Rep. 2018, 8, 8452. [Google Scholar]
- Ong, T.H.; Chitra, E.; Ramamurthy, S.; Siddalingam, R.P.; Yen, K.H.; Ambu, S.P.; Davamani, F. Chitosan-propolis nanoparticle formulation demonstrates anti-bacterial activity against Enterococcus faecalis biofilm. PLoS ONE 2017, 12, e0174888. [Google Scholar]
- Chien, Y.C. Surface sensing for biofilm formation in Pseudomonas aeruginosa. Front. Microbiol. 2017, 8, 2671. [Google Scholar]
- Hahnel, S.; Muhlbauer, G.; Hoffmann, J.; Ionescu, A.; Burgers, R.; Rosentritt, M.; Handel, G.; Haberlein, I. Streptococcus mutans and Streptococcus sobrinus biofilm formation and metabolic activity on dental materials. Acta Odontol. Scand. 2012, 70, 114–121. [Google Scholar] [CrossRef]
- He, L.; Wang, H.; Zhang, R.; Li, H. The regulation of Porphyromonas gingivalis biofilm formation by ClpP. Biochem. Biophys. Res. Commun. 2019, 509, 335–340. [Google Scholar] [CrossRef]
- Holmberg, A.; Lood, R.; Morgelin, M.; Soderquist, B.; Holst, E.; Collin, M.; Christensson, B.; Rasmussen, M. Biofilm formation by Propionibacterium acnes is a characteristic of invasive isolates. Clin. Microbiol. Infect. 2009, 15, 787–795. [Google Scholar] [CrossRef] [Green Version]
- Mok, J.Y.; Kang, H.J.; Cho, J.K.; Jeon, I.H.; Kim, H.S.; Park, J.M.; Jeong, S.I.; Shim, J.S.; Jang, S.I. Antioxidative and anti-inflammatory effects of extracts from different organs of Cirsium japonicum var. ussuriense. Korea J. Herbol. 2011, 26, 39–47. [Google Scholar]
- Kim, J.G.; Ha, Q.B.T.; Shin, Y.K.; Kim, K.Y. Antifungal activity of magnoflorine against Candida strains. World J. Microbiol. Biotechnol. 2018, 34, 167. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S. Mechanisms of antibiotic resistance in bacterial biofilms. Int. J. Med. Microbiol. 2002, 292, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Thomas, V.C.; Thurlow, L.R.; Boyle, D.; Hancock, L. Regulation of autolysis-dependent extracellular DNA release by Enterococcus faecalis extracellular proteases influences biofilm development. J. Bacteriol. 2008, 190, 5690–5698. [Google Scholar] [CrossRef] [Green Version]
- Becerra, S.C.; Roy, D.C.; Sanchez, C.J.; Christy, R.J.; Burmeister, D.M. An optimized staining technique for the detection of Gram positive and Gram-negative bacteria within tissue. BMC Res. Notes 2016, 9, 216. [Google Scholar] [CrossRef] [Green Version]
- Daw, K.; Baghdayan, A.S.; Awasthi, S.; Shankar, N. Biofilm and planktonic Enterococcus faecalis elicit different responses from host phagocytes in vitro. FEMS Immunol. Med. Microbiol. 2012, 65, 270–282. [Google Scholar] [CrossRef] [Green Version]
- Sillanpää, J.; Nallapareddy, S.R.; Singh, K.V.; Prakash, V.P.; Fothergill, T.; Ton-That, H.; Murray, B.E. Characterization of the ebpfm pilus-encoding operon of Enterococcus faecium and its role in biofilm formation and virulence in a murine model of urinary tract infection. Virulence 2010, 1, 236. [Google Scholar] [CrossRef] [Green Version]
- Hendrickx, A.P.A.; Luit-Asbroek, M.V.; Schapendonk, C.M.E.; Wamel, W.J.B.; Braat, J.C.; Wijnands, L.M.; Bonten, M.J.M.; Willems, R.J.L. SgrA, a Nidogen-Binding LPXTG Surface Adhesin Implicated in Biofilm Formation, and EcbA, a Collagen Binding MSCRAMM, Are Two Novel Adhesins of Hospital-Acquired Enterococcus faecium. Infect Immun. 2009, 77, 5097. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Strain Number | IC50 (Half Maximal Inhibitory Concentration; μg/mL) | Source |
|---|---|---|---|
| Enterococcus faecalis | CCARM 5511 | 0.39 | Purchased from KACC (Korean Agricultural Culture Collection, Wanju, Korea), CCARM (Culture Collection of Antimicrobial Resistant Microbes, Seoul, Korea), or KCTC (Korean Collection for Type Cultures, Daejon, Korea) |
| Enterococcus faecium | KACC11954 | 0.19 | |
| Escherihia coli | KACC11598 | 0.25 | |
| Streptococcus mutans | KACC16833 | 1.2 | |
| Streptococcus sobrinus | CCARM3506 | 1.4 | |
| Staphylococcus aureus | KCTC5809 | 0.39 | |
| Pseudomonas aeruginosa | KACC14021 | 0.9 | |
| Cutibacterium acnes | CCARM9009 | 12.5 | |
| Porphyromonas gingivalis | KCTC5352 | 0.9 |
| Genes | Primer Sequence: 5′ to 3′ | Function | Reference |
|---|---|---|---|
| For E. faecium | |||
| esp | F: CCACGAGTTAGAGGGAACAG R: TTGGAGCCCCATCTTTTTCA | Biofilm formation | [13] |
| bps | F: TATCAGCAACAAGCGGTCAA R: AATCCTGCCCTTTTTCGATT | Biofilm formation | [27] |
| fsrC | F: GCTTATTTGGAAGAACAACGTATCAA R:CGAAACATCGCTAGCTCTTCGT | Efae regulator | [12] |
| gelE | F: CGGAACATACTGCCGGTTTAGA R: TGGATTAGATGCCACCCGAAAT | Gelatinase | [12] |
| fsrB | F: TGCTCAAAAAGCAAAGCCTTATAA R: GATGACGAGACCGTAGAGTATTACTGAA | Efae regulator | [12] |
| ebpA | F: ACCAAGCCAGACGAAATAGAAGAAG R: ATTGTTTTGGTCAGGTGCATCATAGA | Biofilm-associated pili | [27] |
| acm | F: TCAGCAGTAATGTCACTTCGTTG R: GAATAGGCTGTTCATCTGCTCG | Gelatinase | [28] |
| scm | F: CTAACTGGTAACTATGGCTTGT R: GTCCGTGCTGTCACTTGT | Gelatinase | [28] |
| tufA | F: TACACGCCACTACGCTCAC R: AGCTCCGTCCATTTGAGCAG | Housekeeping gene | [12] |
| For E. faecalis | |||
| gelE | F: CGFAACATACTCAACGTTTGAC R: TGGATTAGATGCADDDGAAAT | Gelatinase | [21] |
| esp | F: GCATCAGTATTAGTTGGT R: TTCCTTGTAACACATCAC | Biofilm formation | [21] |
| fsrB | F: TGCYCAAAAAGCAAAGCCTTATAA R: GATGACGAGACCGTAGAGTATTACTGAA | Efae regulator | [21] |
| ebpB | F: CGTACAGGAGGCAAGTCTTT R: AGGTATTCCCCGCTTGATTT | Biofilm-associated pili | [21] |
| cylLS | F: CTGTTGCGGCGACAGCT R: CCACCAACCCAGCCACAA | Cytolysin toxin | [21] |
| cylR2 | F: TTTATTTTTATTGGATATCATTCTGTAGTC R: TTCGCTCATCTTTTTTGAATACAG | Cytolysin regulatory | [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.; Kim, K.-Y. Pectolinarin Inhibits the Bacterial Biofilm Formation and Thereby Reduces Bacterial Pathogenicity. Antibiotics 2022, 11, 598. https://doi.org/10.3390/antibiotics11050598
Kim D, Kim K-Y. Pectolinarin Inhibits the Bacterial Biofilm Formation and Thereby Reduces Bacterial Pathogenicity. Antibiotics. 2022; 11(5):598. https://doi.org/10.3390/antibiotics11050598
Chicago/Turabian StyleKim, Daseul, and Ki-Young Kim. 2022. "Pectolinarin Inhibits the Bacterial Biofilm Formation and Thereby Reduces Bacterial Pathogenicity" Antibiotics 11, no. 5: 598. https://doi.org/10.3390/antibiotics11050598
APA StyleKim, D., & Kim, K.-Y. (2022). Pectolinarin Inhibits the Bacterial Biofilm Formation and Thereby Reduces Bacterial Pathogenicity. Antibiotics, 11(5), 598. https://doi.org/10.3390/antibiotics11050598