Recent Development of Advanced Fluorescent Molecular Probes for Organelle-Targeted Cell Imaging


Abstract

1. Introduction
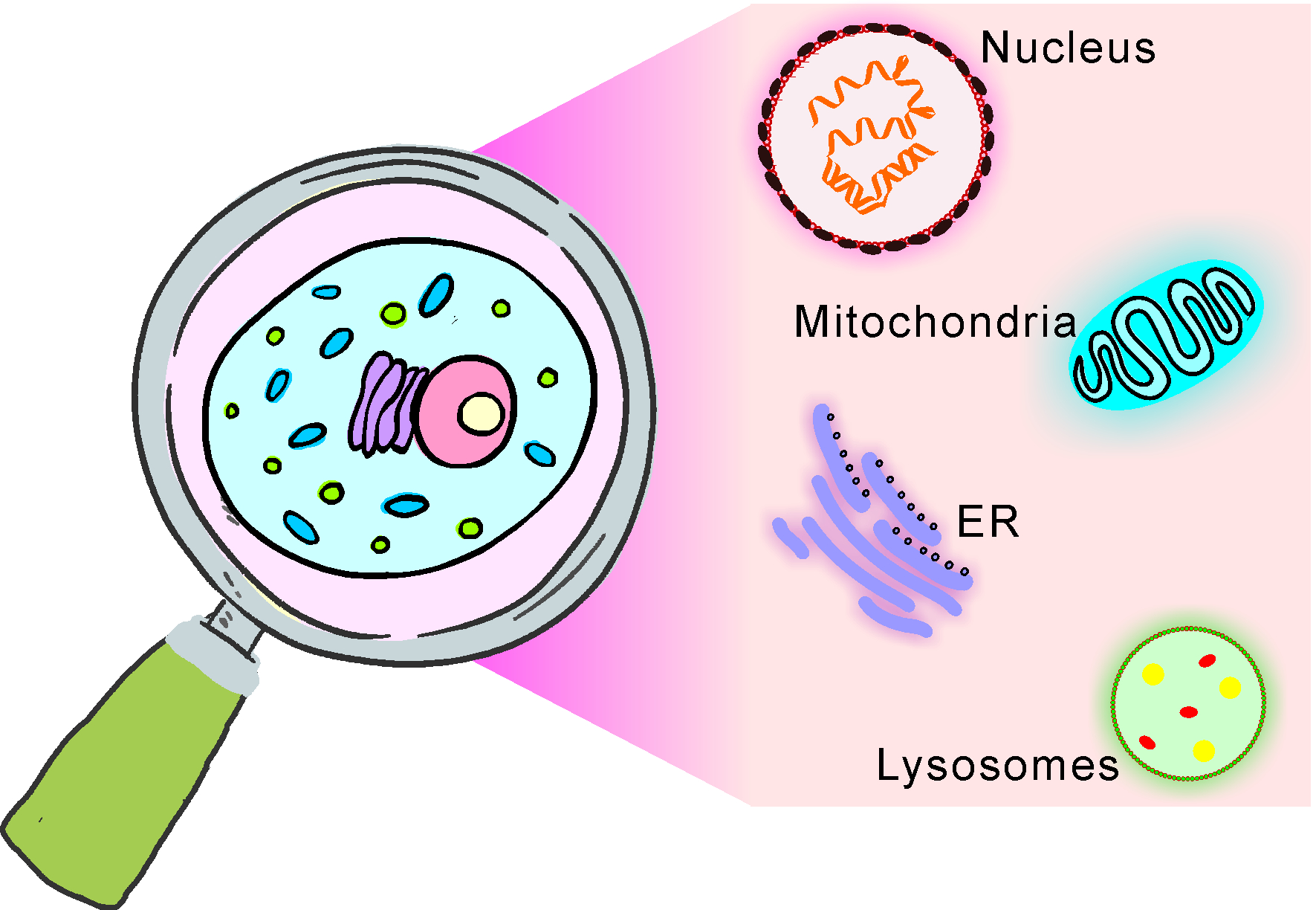
2. Design of Molecular Probes for Organelle-Targeted Cell Imaging
2.1. Nucleus-Targeted Molecular Probes
2.1.1. Properties of the Nucleus
2.1.2. Nucleus-Targeted Probe Design
2.2. Mitochondria-Targeted Molecular Probes
2.2.1. Property of the Mitochondria
2.2.2. Mitochondria-Targeted Probe Design
2.3. Endoplasmic-Reticulum-Targeted Molecular Probes
2.3.1. Properties of the Endoplasmic Reticulum
2.3.2. ER-Targeted Probe Design
2.4. Lysosome-Targeted Molecular Probe
2.4.1. Properties of Lysosomes
2.4.2. Lysosome-Targeted Probe Design
2.5. Dual-Targeted Molecular Probe
3. Summary and Outlook
- (1)
- Low cytotoxicity: The cytotoxicity of the fluorescent molecular probes includes two parts. The first is the phototoxicity generated when the probes were excited by irradiation sources. Under the excitation conditions during cell imaging, the generation of ROS is inevitable. The exceedingly exogenous ROS may break the intracellular redox balance and cause oxidative damage to intracellular biomacromolecules, including DNA, proteins, and lipids, thus influencing the statuses of organelles and living cells. This property of the molecules has been applied to photodynamic therapy for disease treatment; however, it is a limitation of probes in the application of cell imaging, especially in cases of super-resolution and longtime imaging, which need higher emission energy. The modification of the fluorophore is still necessary to obtain a probe that works better inside cells. Chen and co-workers have introduced a triplet-state engineering strategy with which to construct mitochondria-targeted probes with reduced phototoxicity, providing a potential strategy for this direction [182]. The other part of the cytotoxicity of the molecular probes is the interruption of cell activity from the molecules themselves. For example, the membrane potential of the mitochondria will be somehow neutralized by the cationic molecules accumulated around the IMM. The underlying mechanism can introduce interferences with the normal activity of the respiratory chain reaction and thus cause the dysfunction of mitochondria. Therefore, a suitable targeted strategy is needed. The mitochondria-penetrating peptides (MPPs) are promising choices [183,184], which were reported to guide exogenous molecules into the mitochondrial location. Similarly, the sulfonamide group that can react with the K+ channel on the ER surface may influence the bioactivity of the ER. Therefore, several ER localization signal sequences that guide intracellular protein distribution, such as ER retention signal sequences, KDEL (—Lys-Asp-Glu-Leu-COOH), and ER insertion signal sequences, Eriss (ER insertion signal sequence), have been conjugated with molecular probes for ER-targeted imaging [185]. Some special short peptides, named nuclear localization signals (NLSs), or other chemical motifs that may recognize the importins in nuclei were applied to construct the nucleus-targeted probes [186].
- (2)
- The capacity of in vivo imaging: Fluorescent imaging in vivo is always challenging but engaging because it provides the most straightforward biological information. Many of the probes developed at this point possess a relatively short wavelength of excitation/emission located in the UV–Vis wavelength region. This region overlaps with the excitation/emission spectra of biomolecules and biosystems. Therefore, the result might be cluttered by the high background signals. In addition, the absorbance of the short-wavelength light by the biological samples will also limit the application of the probes in deep sample penetration imaging. The satisfying wavelength for in vivo imaging is in the near-infrared range (NIR, 650–950 nm), which can avoid interference form the biological samples. A number of NIR fluorophores have been developed, suggesting the importance of considering the link design with particular moieties for in vivo imaging at the organelle level. Another encouraging method is the design of two-photon-excited fluorophores, which can be excited by two lower-energy NIR photons. Therefore, two-photon fluorescent microscopy can be applied for imaging in vivo by using fluorescent probes with an increased penetration depth and other advantages, such as a prolonged observation time.
- (3)
- Biological guidance: Most molecular probes are developed by chemists, but the utilization of probes is most likely carried out by biologists. The gap between these two fields is obvious, implying that communication between scientists is integral to major breakthroughs and beyond. Our standpoint suggests that the development of biology should guide probe design. The studies on ferroptosis created an urgent demand for the detection of cellular iron and lipid peroxidase, as an example [187]. It was recently found that the calcium transients on the ER surface would trigger the process of autophagy [21], meaning that the ER-targeted Ca2+ probes are promising tools for autophagy monitoring. Moreover, the development of novel probes may also encourage biologists to discover unknown bioprocesses. For example, high-resolution methods can assist in distinguishing the different modes of mitochondrial fission; this may explain how other organelles participate in modulating the fission process in cells [188]. Furthermore, a series of novel probes that can monitor the membrane tension on the plasma membrane and other organelles were developed recently and promote the understanding of mechanobiological processes, thus indicating the importance of mechanoforce as an interesting parameter in the regulation of bioactivities [189,190,191,192]. Therefore, it is critical for scientists in different fields to exchange their cutting-edge knowledge to discover the most suitable molecular probes, thus advancing the growth of science.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kluck, R.; Bossy-Wetzel, E.; Green, D.; Newmeyer, D. The release of cytochrome c from mitochondria: A primary site for Bcl-2 regulation of apoptosis. Science 1997, 275, 1132–1136. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, X.; Bhalla, K.; Kim, C.; Ibrado, A.; Cai, J.; Peng, T.-I.; Jones, D.; Wang, X. Prevention of apoptosis by Bcl-2: Release of cytochrome c from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Yarman, A.; Neumann, B.; Bosserdt, M.; Gajovic-Eichelmann, N.; Scheller, F. Peroxide-dependent analyte conversion by the heme prosthetic group, the heme Peptide “microperoxidase-11” and cytochrome C on chitosan capped gold nanoparticles modified electrodes. Biosensors 2012, 2, 189–204. [Google Scholar] [CrossRef]
- Gomez-Pastor, R.; Burchfiel, E.; Thiele, D. Regulation of heat shock transcription factors and their roles in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 4–19. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Matsui, T.; Yamamoto, A.; Okada, T.; Mori, K. XBP1 mRNA is induced by ATF6 and spliced by IRE1 in response to ER stress to produce a highly active transcription factor. Cell 2001, 107, 881–891. [Google Scholar] [CrossRef]
- Han, J.; Backa, S.; Hur, J.; Lin, Y.; Gildersleeve, R.; Shan, J.; Yuan, C.; Krokowski, D.; Wang, S.; Hatzoglou, M.; et al. ER-stress-induced transcriptional regulation increases protein synthesis leading to cell death. Nat. Cell Biol. 2013, 15, 481–490. [Google Scholar] [CrossRef]
- Wu, J.; Kaufman, R. From acute ER stress to physiological roles of the Unfolded Protein Response. Cell Death Differ. 2006, 13, 374–384. [Google Scholar] [CrossRef]
- Ziegler, D.; Vindrieux, D.; Goehrig, D.; Jaber, S.; Collin, G.; Griveau, A.; Wiel, C.; Bendridi, N.; Djebali, S.; Farfariello, V.; et al. Calcium channel ITPR2 and mitochondria-ER contacts promote cellular senescence and aging. Nat. Commun. 2021, 12, 720. [Google Scholar] [CrossRef]
- Simoes, I.; Morciano, G.; Lebiedzinska-Arciszewska, M.; Aguiari, G.; Pinton, P.; Potes, Y.; Wieckowski, M. The mystery of mitochondria-ER contact sites in physiology and pathology: A cancer perspective. Biochim. Et Biophys. Acta Mol. Basis Dis. 2020, 1866, 165834. [Google Scholar] [CrossRef]
- Chen, C.; Peng, J.; Xia, H.-S.; Yang, G.-F.; Wu, Q.-S.; Chen, L.-D.; Zeng, L.-B.; Zhang, Z.-L.; Pang, D.-W.; Li, Y. Quantum dots-based immunofluorescence technology for the quantitative determination of HER2 expression in breast cancer. Biomaterials 2009, 30, 2912–2918. [Google Scholar] [CrossRef]
- Wang, Y.; Shyy, J.; Chien, S. Fluorescence proteins, live-cell imaging, and mechanobiology: Seeing is believing. Annu. Rev. Biomed. Eng. 2008, 10, 1–38. [Google Scholar] [CrossRef]
- Bikoff, E. Formation of complexes between self-peptides and MHC class II molecules in cells defective for presentation of exogenous protein antigens. J. Immunol. 1992, 149, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zeng, Z.; Jiang, J.-H.; Chang, Y.-T.; Yuan, L. Discerning the Chemistry in Individual Organelles with Small-Molecule Fluorescent Probes. Angew. Chem. Int. Ed. 2016, 55, 13658–13699. [Google Scholar] [CrossRef]
- Kapuscinski, J. DAPI: A DNA-specific fluorescent probe. Biotech. Histochem. Off. Publ. Biol. Stain. Comm. 1995, 70, 220–233. [Google Scholar] [CrossRef]
- Pendergrass, W.; Wolf, N.; Poot, M. Efficacy of MitoTracker Green (TM) and CMXRosamine to measure changes in mitochondrial membrane potentials in living cells and tissues. Cytom. Part A 2004, 61A, 162–169. [Google Scholar] [CrossRef]
- Cole, L.; Davies, D.; Hyde, G.; Ashford, A. ER-Tracker dye and BODIPY-brefeldin A differentiate the endoplasmic reticulum and Golgi bodies from the tubular-vacuole system in living hyphae of Pisolithus tinctorius. J. Microsc. 2000, 197, 239–248. [Google Scholar] [CrossRef]
- Chikte, S.; Panchal, N.; Warnes, G. Use of LysoTracker Dyes: A Flow Cytometric Study of Autophagy. Cytom. Part A 2014, 85, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Dutta, T.; Pal, K.; Koner, A. Intracellular Physical Properties with Small Organic Fluorescent Probes: Recent Advances and Future Perspectives. Chem. Rec. 2022, 22, e202200035. [Google Scholar] [CrossRef] [PubMed]
- Pierzynska-Mach, A.; Janowski, P.; Dobrucki, J. Evaluation of Acridine Orange, LysoTracker Red, and Quinacrine as Fluorescent Probes for Long-Term Tracking of Acidic Vesicles. Cytom. Part A 2014, 85A, 729–737. [Google Scholar] [CrossRef]
- Chretien, D.; Benit, P.; Ha, H.-H.; Keipert, S.; El-Khoury, R.; Chang, Y.-T.; Jastroch, M.; Jacobs, H.; Rustin, P.; Rak, M. Mitochondria are physiologically maintained at close to 50 degrees C. PLoS Biol. 2018, 16, e2003992. [Google Scholar] [CrossRef]
- Zheng, Q.; Chen, Y.; Chen, D.; Zhao, H.; Feng, Y.; Meng, Q.; Zhao, Y.; Zhang, H. Calcium transients on the ER surface trigger liquid-liquid phase separation of FIP200 to specify autophagosome initiation sites. Cell 2022, 185, 4082–4098. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, B.; Angeli, J.; Bayir, H.; Bush, A.; Conrad, M.; Dixon, S.; Fulda, S.; Gascon, S.; Hatzios, S.; Kagan, V.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism. Redox Biol. Dis. Cell 2017, 171, 273–285. [Google Scholar]
- Downs, J.; Nussenzweig, M.; Nussenzweig, A. Chromatin dynamics and the preservation of genetic information. Nature 2007, 447, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, S.; Letarte, M. Hereditary haemorrhagic telangiectasia: Current views on genetics and mechanisms of disease. J. Med. Genet. 2006, 43, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Rotthier, A.; Baets, J.; Timmerman, V.; Janssens, K. Mechanisms of disease in hereditary sensory and autonomic neuropathies. Nat. Rev. Neurol. 2012, 8, 73–85. [Google Scholar] [CrossRef]
- Solowska, J.; Baas, P. Hereditary spastic paraplegia SPG4: What is known and not known about the disease. Brain 2015, 138, 2471–2484. [Google Scholar] [CrossRef]
- Prokhorova, E.; Zamaraev, A.; Kopeina, G.; Zhivotovsky, B.; Lavrik, I. Role of the nucleus in apoptosis: Signaling and execution. Cell. Mol. Life Sci. 2015, 72, 4593–4612. [Google Scholar] [CrossRef]
- Kang, Y.; He, R.; Yang, L.; Qin, D.; Guggilam, A.; Elks, C.; Yan, N.; Guo, Z.; Francis, J. Brain tumour necrosis factor-alpha modulates neurotransmitters in hypothalamic paraventricular nucleus in heart failure. Cardiovasc. Res. 2009, 83, 737–746. [Google Scholar] [CrossRef]
- Paine, P.; Moore, L.; Horowitz, S. Nuclear envelope permeability. Nature 1975, 254, 109–114. [Google Scholar] [CrossRef]
- Dervan, P. Molecular recognition of DNA by small molecules. Bioorganic Med. Chem. 2001, 9, 2215–2235. [Google Scholar] [CrossRef]
- Sirajuddin, M.; Ali, S.; Badshah, A. Drug-DNA interactions and their study by UV-Visible, fluorescence spectroscopies and cyclic voltametry. J. Photochem. Photobiol. B Biol. 2013, 124, 1–19. [Google Scholar] [CrossRef]
- Kim, Y.; Sah, R.; Doong, J.; Grodzinsky, A. Fluorometric assay of DNA in cartilage explants using Hoechst 33258. Anal. Biochem. 1988, 174, 168–176. [Google Scholar] [CrossRef]
- Ahn, S.; Costa, J.; Emanuel, J. PicoGreen quantitation of DNA: Effective evaluation of samples pre- or post-PCR. Nucleic Acids Res. 1996, 24, 2623–2625. [Google Scholar] [CrossRef]
- Bucevicius, J.; Lukinavicius, G.; Gerasimaite, R. The Use of Hoechst Dyes for DNA Staining and Beyond. Chemosensors 2018, 6, 18. [Google Scholar] [CrossRef]
- Ishida, M.; Watanabe, H.; Takigawa, K.; Kurishita, Y.; Oki, C.; Nakamura, A.; Hamachi, I.; Tsukiji, S. Synthetic Self-Localizing Ligands That Control the Spatial Location of Proteins in Living Cells. J. Am. Chem. Soc. 2013, 135, 12684–12689. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Takigawa, K.; Kurishita, Y.; Kuwata, K.; Ishida, M.; Shimoda, Y.; Hamachi, I.; Tsukiji, S. Hoechst tagging: A modular strategy to design synthetic fluorescent probes for live-cell nucleus imaging. Chem. Commun. 2014, 50, 6149–6152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ye, Z.; Zhang, X.; Man, H.; Huang, Z.; Li, N.; Xiao, Y. A targetable fluorescent probe for dSTORM super-resolution imaging of live cell nucleus DNA. Chem. Commun. 2019, 55, 1951–1954. [Google Scholar] [CrossRef]
- Narayanaswamy, N.; Das, S.; Samanta, P.; Banu, K.; Sharma, G.; Mondal, N.; Dhar, S.; Pati, S.; Govindaraju, T. Sequence-specific recognition of DNA minor groove by an NIR-fluorescence switch-on probe and its potential applications. Nucleic Acids Res. 2015, 43, 8651–8663. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, S.; Murugan, N.; Ghosh, D.; Narayanaswamy, N.; Govindaraju, T.; Basu, G. DNA Minor Groove-Induced cis-trans Isomerization of a Near-Infrared Fluorescent Probe. Biochemistry 2021, 60, 2084–2097. [Google Scholar] [CrossRef] [PubMed]
- Abeywickrama, C.; Bertman, K.; Plescia, C.; Stahelin, R.; Pang, Y. Structural Effect on the Cellular Selectivity of an NIR-Emitting Cyanine Probe: From Lysosome to Simultaneous Nucleus and Mitochondria Selectivity with Potential for Monitoring Mitochondria Dysfunction in Cells. ACS Appl. Bio Mater. 2019, 2, 5174–5181. [Google Scholar] [CrossRef]
- Pratihar, S.; Suseela, Y.; Govindaraju, T. Threading Intercalator-Induced Nanocondensates and Role of Endogenous Metal Ions in Decondensation for DNA Delivery. ACS Appl. Bio Mater. 2020, 3, 6979–6991. [Google Scholar] [CrossRef]
- Abeywickrama, C.; Wijesinghe, K.; Plescia, C.; Fisher, L.; Goodson, T.; Stahelin, R.; Pang, Y. A pyrene-based two-photon excitable fluorescent probe to visualize nuclei in live cells. Photochem. Photobiol. Sci. 2020, 19, 1152–1159. [Google Scholar] [CrossRef]
- Sayed, M.; Gubbala, G.; Pal, H. Contrasting interactions of DNA-intercalating dye acridine orange with hydroxypropyl derivatives of -cyclodextrin and -cyclodextrin hosts. New J. Chem. 2019, 43, 724–736. [Google Scholar] [CrossRef]
- Kandinska, M.; Cheshmedzhieva, D.; Kostadinov, A.; Rusinov, K.; Rangelov, M.; Todorova, N.; Ilieva, S.; Ivanov, D.; Videva, V.; Lozanov, V.; et al. Tricationic asymmetric monomeric monomethine cyanine dyes with chlorine and trifluoromethyl functionality—Fluorogenic nucleic acids probes. J. Mol. Liq. 2021, 342, 117501. [Google Scholar] [CrossRef]
- Ali, T.H.; Mandal, A.M.; Alhasan, A.; Dehaen, W. Surface fabrication of magnetic core-shell silica nanoparticles with perylene diimide as a fluorescent dye for nucleic acid visualization. J. Mol. Liq. 2022, 359, 119345. [Google Scholar]
- Zhao, L.; He, X.; Li, D.; Xu, S.; Huang, Y.; Li, X.; Wang, X.; Sun, Y.; Ma, P.; Song, D. A novel fluorescent probe for the localization of nucleoli developed via a chain reaction of endogenous cysteine in cells. J. Mater. Chem. B 2020, 8, 7652–7658. [Google Scholar] [CrossRef]
- Bochman, M.; Paeschke, K.; Zakian, V. DNA secondary structures: Stability and function of G-quadruplex structures. Nat. Rev. Genet. 2012, 13, 770–780. [Google Scholar] [CrossRef]
- Biffi, G.; Tannahill, D.; McCafferty, J.; Balasubramanian, S. Quantitative visualization of DNA G-quadruplex structures in human cells. Nat. Chem. 2013, 5, 182–186. [Google Scholar] [CrossRef]
- Cui, Y.; Kong, D.; Ghimire, C.; Xu, C.; Mao, H. Mutually Exclusive Formation of G-Quadruplex and i-Motif Is a General Phenomenon Governed by Steric Hindrance in Duplex DNA. Biochemistry 2016, 55, 2291–2299. [Google Scholar] [CrossRef]
- Sutherland, C.; Cui, Y.; Mao, H.; Hurley, L. A Mechanosensor Mechanism Controls the G-Quadruplex/i-Motif Molecular Switch in the MYC Promoter NHE III1. J. Am. Chem. Soc. 2016, 138, 14138–14151. [Google Scholar] [CrossRef] [PubMed]
- Kotar, A.; Wang, B.; Shivalingam, A.; Gonzalez-Garcia, J.; Vilar, R.; Plavec, J. NMR Structure of a Triangulenium-Based Long-Lived Fluorescence Probe Bound to a G-Quadruplex. Angew. Chem. Int. Ed. 2016, 55, 12508–12511. [Google Scholar] [CrossRef]
- Summers, P.; Lewis, B.; Gonzalez-Garcia, J.; Porreca, R.; Lim, A.; Cadinu, P.; Martin-Pintado, N.; Mann, D.; Edel, J.; Vannier, J.; et al. Visualising G-quadruplex DNA dynamics in live cells by fluorescence lifetime imaging microscopy. Nat. Commun. 2021, 12, 162. [Google Scholar] [CrossRef]
- Sun, W.; Cui, J.-X.; Ma, L.-L.; Lu, Z.-L.; Gong, B.; He, L.; Wang, R. Imaging nucleus viscosity and G-quadruplex DNA in living cells using a nucleus-targeting two-photon fluorescent probe. Analyst 2018, 143, 5799–5804. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Jia, H.; Zhang, X.; Chen, Y.; Liu, H.; Tan, W. Engineering a subcellular targetable, red-emitting, and ratiometric fluorescent probe for Ca2+ and its bioimaging applications. Anal. Bioanal. Chem. 2010, 397, 1245–1250. [Google Scholar] [CrossRef]
- Exposito-Rodriguez, M.; Laissue, P.; Yvon-Durocher, G.; Smirnoff, N.; Mullineaux, P. Photosynthesis-dependent H2O2 transfer from chloroplasts to nuclei provides a high-light signalling mechanism. Nat. Commun. 2017, 8, 49. [Google Scholar] [CrossRef]
- Maruta, T.; Noshi, M.; Tanouchi, A.; Tamoi, M.; Yabuta, Y.; Yoshimura, K.; Ishikawa, T.; Shigeoka, S. H2O2-triggered Retrograde Signaling from Chloroplasts to Nucleus Plays Specific Role in Response to Stress. J. Biol. Chem. 2012, 287, 11717–11729. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.; Xie, Q. Nitric oxide synthases: Roles, tolls, and controls. Cell 1994, 78, 915–918. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, B.; Tang, Y.; Chang, Z.; Chang, C. A Nuclear-Localized Fluorescent Hydrogen Peroxide Probe for Monitoring Sirtuin-Mediated Oxidative Stress Responses In Vivo. Chem. Biol. 2011, 18, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Huang, Z.; Ma, D.; Yan, Y.; Zhang, X.; Xiao, Y. A nucleus targetable fluorescent probe for ratiometric imaging of endogenous NO in living cells and zebrafishes. Analyst 2021, 146, 4130–4134. [Google Scholar] [CrossRef] [PubMed]
- McBride, H.; Neuspiel, M.; Wasiak, S. Mitochondria: More than just a powerhouse. Curr. Biol. 2006, 16, R551–R560. [Google Scholar] [CrossRef] [PubMed]
- Shadel, G.; Horvath, T. Mitochondrial ROS Signaling in Organismal Homeostasis. Cell 2015, 163, 560–569. [Google Scholar] [CrossRef]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of cell death: The calcium-apoptosis link. Nat. Rev. Mol. Cell Biol. 2003, 4, 552–565. [Google Scholar] [CrossRef]
- Narendra, D.; Tanaka, A.; Suen, D.-F.; Youle, R. Parkin is recruited selectively to impaired mitochondria and promotes their autophagy. J. Cell Biol. 2008, 183, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xue, Y.; Xu, X.; Wang, G.; Liu, Y.; Wu, H.; Li, W.; Wang, Y.; Chen, Z.; Zhang, W.; et al. A mitochondrial FUNDC1/HSC70 interaction organizes the proteostatic stress response at the risk of cell morbidity. EMBO J. 2019, 38, e98786. [Google Scholar] [CrossRef]
- Green, D.; Reed, J. Mitochondria and apoptosis. Science 1998, 281, 1309–1312. [Google Scholar] [CrossRef] [PubMed]
- Tait, S.; Green, D. Mitochondria and cell death: Outer membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Angelova, P.; Esteras, N.; Abramov, A. Mitochondria and lipid peroxidation in the mechanism of neurodegeneration: Finding ways for prevention. Med. Res. Rev. 2021, 41, 770–784. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Calvani, R.; Coelho-Junior, H.; Marzetti, E. Cell Death and Inflammation: The Role of Mitochondria in Health and Disease. Cells 2021, 10, 537. [Google Scholar] [CrossRef]
- Carinci, M.; Vezzani, B.; Patergnani, S.; Ludewig, P.; Lessmann, K.; Magnus, T.; Casetta, I.; Pugliatti, M.; Pinton, P.; Giorgi, C. Different Roles of Mitochondria in Cell Death and Inflammation: Focusing on Mitochondrial Quality Control in Ischemic Stroke and Reperfusion. Biomedicines 2021, 9, 169. [Google Scholar] [CrossRef]
- Obrador, E.; Salvador-Palmer, R.; Lopez-Blanch, R.; Jihad-Jebbar, A.; Valles, S.; Estrela, J. The Link between Oxidative Stress, Redox Status, Bioenergetics and Mitochondria in the Pathophysiology of ALS. Int. J. Mol. Sci. 2021, 22, 6352. [Google Scholar] [CrossRef]
- Huang, M.; Myers, C.; Wang, Y.; You, M. Mitochondria as a Novel Target for Cancer Chemoprevention: Emergence of Mitochondrial-targeting Agents. Cancer Prev. Res. 2021, 14, 285–306. [Google Scholar] [CrossRef]
- Zielonka, J.; Joseph, J.; Sikora, A.; Hardy, M.; Ouari, O.; Vasquez-Vivar, J.; Cheng, G.; Lopez, M.; Kalyanaraman, B. Mitochondria-Targeted Triphenylphosphonium-Based Compounds: Syntheses, Mechanisms of Action, and Therapeutic and Diagnostic Applications. Chem. Rev. 2017, 117, 10043–10120. [Google Scholar] [CrossRef]
- Frey, T.; Mannella, C. The internal structure of mitochondria. Trends Biochem. Sci. 2000, 25, 319–324. [Google Scholar] [CrossRef]
- Henze, K.; Martin, W. Evolutionary biology: Essence of mitochondria. Nature 2003, 426, 127–128. [Google Scholar] [CrossRef]
- Liu, Z.; Bushnell, W.; Brambl, R. Pontentiometric cyanine dyes are sensitive probes for mitochondria in intact plant cells: Kinetin enhances mitochondrial fluorescence. Plant Physiol. 1987, 84, 1385–1390. [Google Scholar] [CrossRef] [PubMed]
- Zhai, R.; Fang, B.; Lai, Y.; Peng, B.; Bai, H.; Liu, X.; Li, L.; Huang, W. Small-molecule fluorogenic probes for mitochondrial nanoscale imaging. Chem. Soc. Rev. 2022, 52, 942–972. [Google Scholar] [CrossRef]
- Chazotte, B. Labeling mitochondria with MitoTracker dyes. Cold Spring Harb. Protoc. 2011, 2011, 990–992. [Google Scholar] [CrossRef]
- Kasparov, S. Feasibility of Photodynamic Therapy for Glioblastoma with the Mitochondria-Targeted Photosensitizer Tetramethylrhodamine Methyl Ester (TMRM). Biomedicines 2021, 9, 1453. [Google Scholar]
- Sivandzade, F.; Bhalerao, A.; Cucullo, L. Analysis of the Mitochondrial Membrane Potential Using the Cationic JC-1 Dye as a Sensitive Fluorescent Probe. Bio-Protocol 2019, 9, e3128. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C. Reconciling the chemistry and biology of reactive oxygen species. Nat. Chem. Biol. 2008, 4, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Chouchani, E.; Pell, V.; Gaude, E.; Aksentijevic, D.; Sundier, S.; Robb, E.; Logan, A.; Nadtochiy, S.; Ord, E.; Smith, A.; et al. Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS. Nature 2014, 515, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.; Janes, M.; Pehar, M.; Monette, J.; Ross, M.; Hagen, T.; Murphy, M.; Beckman, J. Selective fluorescent imaging of superoxide in vivo using ethidium-based probes. Proc. Natl. Acad. Sci. USA 2006, 103, 15038–15043. [Google Scholar] [CrossRef]
- Li, Z.; Ren, T.-B.; Zhang, X.-X.; Xu, S.; Gong, X.-Y.; Yang, Y.; Ke, G.; Yuan, L.; Zhang, X.-B. Precipitated Fluorophore-Based Probe for Accurate Detection of Mitochondrial Analytes. Anal. Chem. 2021, 93, 2235–2243. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, M.; Liu, Z.; Tian, Y. Real-Time Imaging and Simultaneous Quantification of Mitochondrial H2O2 and ATP in Neurons with a Single Two-Photon Fluorescence-Lifetime-Based Probe. J. Am. Chem. Soc. 2020, 142, 7532–7541. [Google Scholar] [CrossRef]
- Niu, H.; Tang, J.; Zhu, X.; Li, Z.; Zhang, Y.; Ye, Y.; Zhao, Y. A three-channel fluorescent probe to image mitochondrial stress. Chem. Commun. 2020, 56, 7710–7713. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Fan, J.; Sun, W.; Sui, K.; Jin, X.; Wang, J.; Peng, X. A highly specific BODIPY-based probe localized in mitochondria for HClO imaging. Analyst 2013, 138, 6091–6096. [Google Scholar] [CrossRef] [PubMed]
- Lacza, Z.; Pankotai, E.; Csordas, A.; Geroo, D.; Kiss, L.; Horvath, E.; Kollai, M.; Busija, D.; Szabo, C. Mitochondrial NO and reactive nitrogen species production: Does mtNOS exist? Nitric Oxide Biol. Chem. 2006, 14, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, C.; Carroll, K. Cysteine-Mediated Redox Signaling: Chemistry, Biology, and Tools for Discovery. Chem. Rev. 2013, 113, 4633–4679. [Google Scholar] [CrossRef]
- Trollinger, D.; Cascio, W.; Lemasters, J. Selective loading of Rhod 2 into mitochondria shows mitochondrial Ca-2+ transients during the contractile cycle in adult rabbit cardiac myocytes. Biochem. Biophys. Res. Commun. 1997, 236, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Sensi, S.; Ton-That, D.; Weiss, J.; Rothe, A.; Gee, K. A new mitochondrial fluorescent zinc sensor. Cell Calcium 2003, 34, 281–284. [Google Scholar] [CrossRef]
- Taki, M.; Akaoka, K.; Iyoshi, S.; Yamamoto, Y. Rosamine-Based Fluorescent Sensor with Femtomolar Affinity for the Reversible Detection of a Mercury Ion. Inorg. Chem. 2012, 51, 13075–13077. [Google Scholar] [CrossRef]
- Glickstein, H.; El, R.B.; Shvartsman, M.; Cabantchik, Z. Intracellular labile iron pools as direct targets of iron chelators: A fluorescence study of chelator action in living cells. Blood 2005, 106, 3242–3250. [Google Scholar] [CrossRef]
- Gao, P.; Pan, W.; Li, N.; Tang, B. Fluorescent probes for organelle-targeted bioactive species imaging. Chem. Sci. 2019, 10, 6035–6071. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, C.; Zheng, Y.; Huang, Z.; Zhang, X.; Xiao, Y. Bio-orthogonal Toolbox for Monitoring Nitric Oxide in Targeted Organelles of Live Cells and Zebrafishes. Anal. Chem. 2022, 94, 15678–15685. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Huo, Y.; Lv, X.; Li, Z.; Cao, H.; Shi, H.; Shi, Y.; Guo, W. Fast-response and highly selective fluorescent probes for biological signaling molecule NO based on N-nitrosation of electron-rich aromatic secondary amines. Biomaterials 2016, 78, 11–19. [Google Scholar] [CrossRef]
- Zhang, J.; Bao, X.; Zhou, J.; Peng, F.; Ren, H.; Dong, X.; Zhao, W. A mitochondria-targeted turn-on fluorescent probe for the detection of glutathione in living cells. Biosens. Bioelectron. 2016, 85, 164–170. [Google Scholar] [CrossRef]
- Michikawa, Y.; Mazzucchelli, F.; Bresolin, N.; Scarlato, G.; Attardi, G. Aging-dependent large accumulation of point mutations in the human mtDNA control region for replication. Science 1999, 286, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.-G.; Kajigaya, S.; Feng, X.; Samsel, L.; McCoy, J.; Torelli, G., Jr.; Young, N. Accumulation of mtDNA variations in human single CD34(+) cells from maternally related individuals: Effects of aging and family genetic background. Stem Cell Res. 2013, 10, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Uno, K.; Sugimoto, N.; Sato, Y. N-aryl pyrido cyanine derivatives are nuclear and organelle DNA markers for two-photon and super-resolution imaging. Nat. Commun. 2021, 12, 2650. [Google Scholar] [CrossRef]
- Chen, X.-C.; Tang, G.-X.; Luo, W.-H.; Shao, W.; Dai, J.; Zeng, S.-T.; Huang, Z.-S.; Chen, S.-B.; Tan, J.-H. Monitoring and Modulating mtDNA G-Quadruplex Dynamics Reveal Its Close Relationship to Cell Glycolysis. J. Am. Chem. Soc. 2021, 143, 20779–20791. [Google Scholar] [CrossRef]
- Burch, S.; Lopez, C. Effects of Cell Density and Microenvironment on Stem Cell Mitochondria Transfer among Human Adipose-Derived Stem Cells and HEK293 Tumorigenic Cells. Int. J. Mol. Sci. 2022, 23, 2003. [Google Scholar] [CrossRef]
- Lee, M.; Park, N.; Yi, C.; Han, J.; Hong, J.; Kim, K.; Kang, D.; Sessler, J.; Kang, C.; Kim, J. Mitochondria-Immobilized pH-Sensitive Off-On Fluorescent Probe. J. Am. Chem. Soc. 2014, 136, 14136–14142. [Google Scholar] [CrossRef]
- Arai, S.; Suzuki, M.; Park, S.-J.; Yoo, J.; Wang, L.; Kang, N.-Y.; Ha, H.-H.; Chang, Y.-T. Mitochondria-targeted fluorescent thermometer monitors intracellular temperature gradient. Chem. Commun. 2015, 51, 8044–8047. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Li, N.; Zhang, X.; Wang, C.; Xiao, Y. Fixable Molecular Thermometer for Real-Time Visualization and Quantification of Mitochondrial Temperature. Anal. Chem. 2018, 90, 13953–13959. [Google Scholar] [CrossRef]
- Huang, Z.; Li, N.; Zhang, X.; Xiao, Y. Mitochondria-Anchored Molecular Thermometer Quantitatively Monitoring Cellular Inflammations. Anal. Chem. 2021, 93, 5081–5088. [Google Scholar] [CrossRef]
- Chen, B.; Mao, S.; Sun, Y.; Sun, L.; Ding, N.; Li, C.; Zhou, J. A mitochondria-targeted near-infrared fluorescent probe for imaging viscosity in living cells and a diabetic mice model. Chem. Commun. 2021, 57, 4376–4379. [Google Scholar] [CrossRef]
- Zhang, G.; Quan, W.; Li, Y.; Song, W.; Lin, W. Near-Infrared Mitochondria-Targetable Single-Molecule probe for Dual-Response of viscosity and sulfur dioxide in vivo. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2022, 270, 120796. [Google Scholar] [CrossRef]
- Nicchitta, C. A platform for compartmentalized protein synthesis: Protein translation and translocation in the ER. Curr. Opin. Cell Biol. 2002, 14, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.; LaFerla, F.; Chan, S.; Leissring, M.; Shepel, P.; Geiger, J. Calcium signaling in the ER: Its role in neuronal plasticity and neurodegenerative disorders. Trends Neurosci. 2000, 23, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Walter, P.; Ron, D. The Unfolded Protein Response: From Stress Pathway to Homeostatic Regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef]
- Thaxton, J.; Wallace, C.; Riesenberg, B.; Zhang, Y.; Paulos, C.; Beeson, C.; Liu, B.; Li, Z. Modulation of Endoplasmic Reticulum Stress Controls CD4(+) T-cell Activation and Antitumor Function. Cancer Immunol. Res. 2017, 5, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, W.; Bone, R.; Sohn, P.; Syed, F.; Reissaus, C.; Mosley, A.; Wijeratne, A.; True, J.; Tong, X.; Kono, T.; et al. Endoplasmic reticulum stress alters ryanodine receptor function in the murine pancreatic cell. J. Biol. Chem. 2019, 294, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Samidurai, A.; Thompson, J.; Hu, Y.; Das, A.; Willard, B.; Lesnefsky, E. Endoplasmic reticulum stress-mediated mitochondrial dysfunction in aged hearts. Biochim. Et Biophys. Acta Mol. Basis Dis. 2020, 1866, 165899. [Google Scholar] [CrossRef]
- Yang, M.; Luo, S.; Wang, X.; Li, C.; Yang, J.; Zhu, X.; Xiao, L.; Sun, L. ER-Phagy: A New Regulator of ER Homeostasis. Front. Cell Dev. Biol. 2021, 9, 684526. [Google Scholar] [CrossRef]
- Wilkinson, S. ER-phagy: Shaping up and destressing the endoplasmic reticulum. FEBS J. 2019, 286, 2645–2663. [Google Scholar] [CrossRef]
- Markgraf, D.; Klemm, R.; Junker, M.; Hannibal-Bach, H.; Ejsing, C.; Rapoport, T. An ER Protein Functionally Couples Neutral Lipid Metabolism on Lipid Droplets to Membrane Lipid Synthesis in the ER. Cell Rep. 2014, 6, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Colston, J.; Horobin, R.; Rashid-Doubell, F.; Pediani, J.; Johal, K. Why fluorescent probes for endoplasmic reticulum are selective: An experimental and QSAR-modelling study. Biotech. Histochem. 2003, 78, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; He, H.-J.; Suzuki, R.; Liu, K.-X.; Tanaka, S.; Sekiguchi, M.; Itoh, H.; Kawahara, K.; Abe, H. Localization of sulfonylurea receptor subunits, SUR2A and SUR2B, in rat heart. J. Histochem. Cytochem. 2007, 55, 795–804. [Google Scholar] [CrossRef]
- Yuriev, E.; Kong, D.; Iskander, M. Investigation of structure-activity relationships in a series of glibenclamide analogues. Eur. J. Med. Chem. 2004, 39, 835–847. [Google Scholar] [CrossRef]
- Singh, D.; Rajput, D.; Kanvah, S. Fluorescent probes for targeting endoplasmic reticulum: Design strategies and their applications. Chem. Commun. 2022, 58, 2413–2429. [Google Scholar] [CrossRef]
- Phaniraj, S.; Gao, Z.; Rane, D.; Peterson, B. Hydrophobic resorufamine derivatives: Potent and selective red fluorescent probes of the endoplasmic reticulum of mammalian cells. Dye. Pigment. 2016, 135, 127–133. [Google Scholar] [CrossRef]
- Banerjee, A.; Banerjee, V.; Czinn, S.; Blanchard, T. Increased reactive oxygen species levels cause ER stress and cytotoxicity in andrographolide treated colon cancer cells. Oncotarget 2017, 8, 26142–26153. [Google Scholar] [CrossRef]
- Khandelwal, A.; Hebert, V.; Dugas, T. Essential role of ER-alpha-dependent NO production in resveratrol-mediated inhibition of restenosis. Am. J. Physiol. -Heart Circ. Physiol. 2010, 299, H1451–H1458. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, S.; Tan, W.; Tang, F.; Long, J.; Li, Z.; Liang, B.; Chu, C.; Yang, J. Gaseous signalling molecule SO2 via Hippo-MST pathway to improve myocardial fibrosis of diabetic rats. Mol. Med. Rep. 2017, 16, 8953–8963. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Huang, H.; Song, W.; Lin, W. A near-infrared endoplasmic reticulum-targeted fluorescent probe to visualize the fluctuation of SO2 during endoplasmic reticulum stress. Chem. Eng. J. 2022, 431, 133468. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, J.; Xiong, H.; Zhang, Y.; Chen, W.; Sheng, J.; Song, X. An endoplasmic reticulum-targetable fluorescent probe for highly selective detection of hydrogen sulfide. Org. Biomol. Chem. 2019, 17, 1436–1441. [Google Scholar] [CrossRef]
- Gotoh, T.; Mori, M. Nitric oxide and endoplasmic reticulum stress. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1439–1446. [Google Scholar] [CrossRef]
- Li, S.-J.; Zhou, D.-Y.; Li, Y.; Liu, H.-W.; Wu, P.; Ou-Yang, J.; Jiang, W.-L.; Li, C.-Y. Efficient Two-Photon Fluorescent Probe for Imaging of Nitric Oxide during Endoplasmic Reticulum Stress. ACS Sens. 2018, 3, 2311–2319. [Google Scholar] [CrossRef]
- Peng, S.; Li, Z.; Zhang, Y.; Cao, W.; Liu, J.; Zhu, W.; Ye, Y. A two-photon fluorescent probe for HNO rapid visualization in endoplasmic reticulum. Sens. Actuator B Chem. 2020, 317, 128211. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, R.; Sun, Y.; Tian, M.; Dong, B. Endoplasmic reticulum-specific fluorescent probe for the two-photon imaging of endogenous superoxide anion (O-2(center dot-)) in live cells and zebrafishes. Talanta 2021, 225, 122020. [Google Scholar] [CrossRef]
- Pak, Y.; Park, S.; Song, G.; Yim, Y.; Kang, H.; Kim, H.; Boufard, J.; Yoon, J. Endoplasmic Reticulum-Targeted Ratiometric N-Heterocyclic Carbene Borane Probe for Two-Photon Microscopic Imaging of Hypochlorous Acid. Anal. Chem. 2018, 90, 12937–12943. [Google Scholar] [CrossRef]
- Ellis, C.; Wang, F.; MacDiarmid, C.; Clark, S.; Lyons, T.; Eide, D. Zinc and the Msc2 zinc transporter protein are required for endoplasmic reticulum function. J. Cell Biol. 2004, 166, 325–335. [Google Scholar] [CrossRef]
- Fang, L.; Trigiante, G.; Crespo-Otero, R.; Hawes, C.; Philpott, M.; Jones, C.; Watkinson, M. Endoplasmic reticulum targeting fluorescent probes to image mobile Zn2+. Chem. Sci. 2019, 10, 10881–10887. [Google Scholar] [CrossRef]
- Lee, M.; Lee, H.; Chang, M.; Kim, H.; Kang, C.; Kim, J. A fluorescent probe for the Fe3+ ion pool in endoplasmic reticulum in liver cells. Dye. Pigment. 2016, 130, 245–250. [Google Scholar] [CrossRef]
- Schneider, L.; Zhang, J. Lysosomal function in macromolecular homeostasis and bioenergetics in Parkinson’s disease. Mol. Neurodegener. 2010, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Platt, F.; d’Azzo, A.; Davidson, B.; Neufeld, E.; Tifft, C. Lysosomal storage diseases. Nat. Rev. Dis. Prim. 2018, 4, 27. [Google Scholar] [CrossRef]
- Platt, F.; Boland, B.; van der Spoel, A. Lysosomal storage disorders: The cellular impact of lysosomal dysfunction. J. Cell Biol. 2012, 199, 723–734. [Google Scholar] [CrossRef]
- Banik, S.; Pedram, K.; Wisnovsky, S.; Ahn, G.; Riley, N.; Bertozzi, C. Lysosome-targeting chimaeras for degradation of extracellular proteins. Nature 2020, 584, 291–297. [Google Scholar] [CrossRef]
- Zhao, L.; Zhao, J.; Zhong, K.; Tong, A.; Jia, D. Targeted protein degradation: Mechanisms, strategies and application. Signal Transduct. Target. Ther. 2022, 7, 113. [Google Scholar] [CrossRef] [PubMed]
- Mindell, J. Lysosomal Acidification Mechanisms. In Annual Review of Physiology; Julius, D., Clapham, D., Eds.; Annual Reviews: Palo Alto, CA, USA, 2012; Volume 74, pp. 69–86. [Google Scholar]
- Kaufmann, A.; Krise, J. Lysosomal sequestration of amine-containing drugs: Analysis and therapeutic implications. J. Pharm. Sci. 2007, 96, 729–746. [Google Scholar] [CrossRef] [PubMed]
- Stark, M.; Silva, T.; Levin, G.; Machuqueiro, M.; Assaraf, Y. The Lysosomotropic Activity of Hydrophobic Weak Base Drugs is Mediated via Their Intercalation into the Lysosomal Membrane. Cells 2020, 9, 1082. [Google Scholar] [CrossRef] [PubMed]
- Tekpli, X.; Rivedal, E.; Gorria, M.; Landvik, N.; Rissel, M.; Dimanche-Boitrel, M.; Baffet, G.; Holme, J.; Lagadic-Gossmann, D. The B a P-increased intercellular communication via translocation of connexin-93 into gap junctions reduces apoptosis. Toxicol. Appl. Pharmacol. 2010, 242, 231–240. [Google Scholar] [CrossRef]
- DeVorkin, L.; Gorski, S. LysoTracker staining to aid in monitoring autophagy in Drosophila. Cold Spring Harb. Protoc. 2014, 2014, 951–958. [Google Scholar] [CrossRef]
- Fogel, J.; Thein, T.; Mariani, F. Use of LysoTracker to Detect Programmed Cell Death in Embryos and Differentiating Embryonic Stem Cells. JoVE J. Vis. Exp. 2012, 68, e4254. [Google Scholar]
- Kurz, T.; Terman, A.; Gustafsson, B.; Brunk, U. Lysosomes in iron metabolism, ageing and apoptosis. Histochem. Cell Biol. 2008, 129, 389–406. [Google Scholar] [CrossRef]
- Rizzollo, F.; More, S.; Vangheluwe, P.; Agostinis, P. The lysosome as a master regulator of iron metabolism. Trends Biochem. Sci. 2021, 46, 960–975. [Google Scholar] [CrossRef] [PubMed]
- Kazan, H.; Urfali-Mamatoglu, C.; Gunduz, U. Iron metabolism and drug resistance in cancer. Biometals 2017, 30, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, F.; Pu, C.; Tong, Z.; Wang, M.; Wang, J. Galactose-imidazole mediated dual-targeting fluorescent probe for detecting Fe3+ in the lysosomes of hepatocytes: Design, synthesis and evaluation. Biosens. Bioelectron. 2022, 204, 114083. [Google Scholar] [CrossRef] [PubMed]
- Giuffrida, M.; Sfrazzetto, G.; Satriano, C.; Zimbone, S.; Tomaselli, G.; Copani, A.; Rizzarelli, E. A New Ratiometric Lysosomal Copper(II) Fluorescent Probe To Map a Dynamic Metallome in Live Cells. Inorg. Chem. 2018, 57, 2365–2368. [Google Scholar] [CrossRef]
- Chen, Y.; Long, Z.; Wang, C.; Zhu, J.; Wang, S.; Liu, Y.; Wei, P.; Yi, T. A lysosome-targeted near-infrared fluorescent probe for cell imaging of Cu2+. Dye. Pigment. 2022, 204, 110472. [Google Scholar] [CrossRef]
- Zhang, S.; Ong, C.; Shen, H. Critical roles of intracellular thiols and calcium in parthenolide-induced apoptosis in human colorectal cancer cells. Cancer Lett. 2004, 208, 143–153. [Google Scholar] [CrossRef]
- Jung, H.; Chen, X.; Kim, J.; Yoon, J. Recent progress in luminescent and colorimetric chemosensors for detection of thiols. Chem. Soc. Rev. 2013, 42, 6019–6031. [Google Scholar] [CrossRef]
- Mao, G.-J.; Liang, Z.-Z.; Bi, J.; Zhang, H.; Meng, H.-M.; Su, L.; Gong, Y.-J.; Feng, S.; Zhang, G. A near-infrared fluorescent probe based on photostable Si-rhodamine for imaging hypochlorous acid during lysosome-involved inflammatory response. Anal. Chim. Acta 2019, 1048, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ren, J.; Bao, X.; Gao, W.; Wu, C.; Zhao, Y. pH-Switchable Fluorescent Probe for Spatially-Confined Visualization of Intracellular Hydrogen Peroxide. Anal. Chem. 2016, 88, 5865–5870. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Wang, J.; Xu, J.; Zheng, C.; Niu, Y.; Wang, C.; Xu, F.; Yuan, L.; Zhao, X.; Liang, L.; et al. A Smart Fluorescent Probe for NO Detection and Application in Myocardial Fibrosis Imaging. Anal. Chem. 2020, 92, 5064–5072. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Xiao, Y.; Jin, L. A Lysosome-Targetable and Two-Photon Fluorescent Probe for Monitoring Endogenous and Exogenous Nitric Oxide in Living Cells. J. Am. Chem. Soc. 2012, 134, 17486–17489. [Google Scholar] [CrossRef]
- Wang, K.; Guo, R.; Chen, X.-Y.; Yang, Y.-S.; Qiao, L.-Q.; Wang, M.-L. Multifunctional lysosome-targetable fluorescent probe for imaging peroxynitrite in acute liver injury model. Chem. Eng. J. 2022, 455, 140491. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, L.; Chen, W.; Huang, J.; Huang, C.; Sheng, J.; Song, X. A Lysosome-Targetable Fluorescent Probe for Simultaneously Sensing Cys/Hcy, GSH, and H2S from Different Signal Patterns. ACS Sens. 2018, 3, 2513–2517. [Google Scholar] [CrossRef]
- Wang, L.; Xiao, Y.; Tian, W. Deng, Activatable Rotor for Quantifying Lysosomal Viscosity in Living Cells. J. Am. Chem. Soc. 2013, 135, 2903–2906. [Google Scholar] [CrossRef]
- Shi, D.; Hu, L.; Li, X.; Liu, W.; Gao, Y.; Li, X.; Jiang, B.; Xia, C.; Guo, Y.; Li, J. Lysosomal polarity increases with aging as revealed by a lysosome- targetable near -infrared fluorescent probe. Sens. Actuator B Chem. 2020, 319, 128302. [Google Scholar] [CrossRef]
- Futerman, A.; van Meer, G. The cell biology of lysosomal storage disorders. Nat. Rev. Mol. Cell Biol. 2004, 5, 554–565. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, T.; He, S.; Zhao, L.; Zeng, X. A lysosome-targeting viscosity-sensitive fluorescent probe based on a novel functionalised near-infrared xanthene-indolium dye and its application in living cells. J. Mater. Chem. B 2020, 8, 8838–8844. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, Z.; Liu, X.; Jiang, Y.; Shen, J. Lysosome-targeting benzothiazole-based fluorescent probe for imaging viscosity and hypochlorite levels in living cells and zebrafish. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2022, 275, 121141. [Google Scholar] [CrossRef]
- Shi, W.-J.; Chen, R.; Yang, J.; Wei, Y.-F.; Guo, Y.; Wang, Z.-Z.; Yan, J.-w.; Niu, L. Novel Meso-Benzothiazole-Substituted BODIPY-Based AIE Fluorescent Rotor for Imaging Lysosomal Viscosity and Monitoring Autophagy. Anal. Chem. 2022, 94, 14707–14715. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yan, H.; Wu, J.; Wang, Z.; He, S.; Zhao, L.; Zeng, X. Lysosomes-targeting near-infrared fluorescent probe for the detection of pH in living cells. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2022, 278, 121368. [Google Scholar] [CrossRef]
- Rutkowski, D.; Kaufman, R. A trip to the ER: Coping with stress. Trends Cell Biol. 2004, 14, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Nargund, A.; Pellegrino, M.; Fiorese, C.; Baker, B.; Haynes, C. Mitochondrial Import Efficiency of ATFS-1 Regulates Mitochondrial UPR Activation. Science 2012, 337, 587–590. [Google Scholar] [CrossRef]
- Rowland, A.; Voeltz, G. Endoplasmic reticulum-mitochondria contacts: Function of the junction. Nat. Rev. Mol. Cell Biol. 2012, 13, 607–615. [Google Scholar] [CrossRef]
- Phillips, M.; Voeltz, G. Structure and function of ER membrane contact sites with other organelles. Nat. Rev. Mol. Cell Biol. 2016, 17, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Giordano, S.; Zhang, J. Autophagy, mitochondria and oxidative stress: Cross-talk and redox signalling. Biochem. J. 2012, 441, 523–540. [Google Scholar] [CrossRef]
- Tian, M.; Sun, J.; Dong, B.; Lin, W. Dynamically Monitoring Cell Viability in a Dual-Color Mode: Construction of an Aggregation/Monomer-Based Probe Capable of Reversible Mitochondria-Nucleus Migration. Angew. Chem. Int. Ed. 2018, 57, 16506–16510. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Yin, J.; Li, M.; Zhu, L.; Dong, B.; Ma, Y.; Lin, W. Simultaneously imaging of SO2 in lysosomes and mitochondria based on a dual organelle-targeted fluorescent probe. Sens. Actuators B Chem. 2019, 292, 80–87. [Google Scholar] [CrossRef]
- Ho, P.-Y.; Chou, T.; Kam, C.; Huang, W.; He, Z.; Ngan, A.; Chen, S. A dual organelle-targeting mechanosensitive probe. Sci. Adv. 2023, 9, eabn5390. [Google Scholar] [CrossRef]
- Shan, Y.-M.; Yu, K.-K.; Wang, N.; Yu, F.-Y.; Li, K.; Liu, Y.-H.; Yu, X.-Q. Assessing ClO- level during ER stress and cellular senescence through a ratio fluorescent probe with dual organelle targeting ability. Sens. Actuator B Chem. 2022, 358, 131383. [Google Scholar] [CrossRef]
- Dai, F.; Zhang, S.; Zhou, B.; Duan, D.-C.; Liu, J.; Zheng, Y.-L.; Chen, H.; Zhang, X.; Zhang, Y. Cellular and Intravital Imaging of NAD(P)H by a Red-Emitting Quinolinium-Based Fluorescent Probe that Features a Shift of Its Product from Mitochondria to the Nucleus. Anal. Chem. 2022, 95, 1335–1342. [Google Scholar] [CrossRef]
- Wang, H.; Sun, Y.; Lin, X.; Feng, W.; Li, Z.; Yu, M. Multi-organelle-targeting pH-dependent NIR fluorescent probe for lysosomal viscosity. Chin. Chem. Lett. 2023, 34, 107626. [Google Scholar] [CrossRef]
- Zhuang, J.; Li, N.; Zhang, Y.; Li, B.; Tang, B. Esterase-Activated Theranostic Prodrug for Dual Organelles Targeted Imaging and Synergetic Chemo-Photodynamic Cancer Therapy. CCS Chem. 2022, 3, 1028–1043. [Google Scholar] [CrossRef]
- Ge, E.; Tian, M.; Lin, W. A unique fluorescent probe for visualization of cell death via its subcellular immigration from lysosomes to nucleus. Sens. Actuators B Chem. 2021, 347, 130656. [Google Scholar] [CrossRef]
- Yapici, N.; Gao, X.; Yan, X.; Hou, S.; Jockusch, S.; Lesniak, L.; Gibson, K.; Bi, L. Novel Dual-Organelle-Targeting Probe (RCPP) for Simultaneous Measurement of Organellar Acidity and Alkalinity in Living Cells. ACS Omega 2021, 6, 31447–31456. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, Y.; Huo, F. A dual-targeted organelles SO2 specific probe for bioimaging in related diseases and food analysis. Chem. Eng. J. 2022, 433, 133750. [Google Scholar] [CrossRef]
- Yang, Z.; Li, L.; Ling, J.; Liu, T.; Huang, X.; Ying, Y.; Zhao, Y.; Lei, K.; Chen, L.; Chen, Z. Cyclooctatetraene-conjugated cyanine mitochondrial probes minimize phototoxicity in fluorescence and nanoscopic imaging. Chem. Sci. 2020, 11, 8506–8516. [Google Scholar] [CrossRef]
- Horton, K.; Stewart, K.; Fonseca, S.; Guo, Q.; Kelley, S. Mitochondria-penetrating peptides. Chem. Biol. 2008, 15, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.; Ahmed, M.; Lei, E.; Wisnovsky, S.; Kelley, S. Peptide-Mediated Delivery of Chemical Probes and Therapeutics to Mitochondria. Acc. Chem. Res. 2016, 49, 1893–1902. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gao, H.; Liu, R.; Chen, C.; Zeng, S.; Liu, Q.; Ding, D. Endoplasmic reticulum targeted AIE bioprobe as a highly efficient inducer of immunogenic cell death. Sci. China Chem. 2020, 63, 1428–1434. [Google Scholar] [CrossRef]
- Lange, A.; Mills, R.; Lange, C.; Stewart, M.; Devine, S.; Corbett, A. Classical nuclear localization signals: Definition, function, and interaction with importin alpha. J. Biol. Chem. 2007, 282, 5101–5105. [Google Scholar] [CrossRef]
- Wang, Y.; Li, C.; Zhuo, J.; Hui, H.; Zhou, B.; Tian, J. The Detection of Divalent Iron and Reactive Oxygen Species During Ferroptosis with the Use of a Dual-Reaction Turn-On Fluorescent Probe. Mol. Imaging Biol. 2022, 1–12. [Google Scholar] [CrossRef]
- Kleele, T.; Rey, T.; Winter, J.; Zaganelli, S.; Mahecic, D.; Lambert, H.; Ruberto, F.; Nemir, M.; Wai, T.; Pedrazzini, T.; et al. Distinct fission signatures predict mitochondrial degradation or biogenesis. Nature 2021, 593, 435–439. [Google Scholar] [CrossRef]
- Colom, A.; Derivery, E.; Soleimanpour, S.; Tomba, C.; Molin, M.D.; Sakai, N.; Gonzalez-Gaitan, M.; Matile, S.; Roux, A. A fluorescent membrane tension probe. Nat. Chem. 2018, 10, 1118–1125. [Google Scholar] [CrossRef]
- Piazzolla, F.; Mercier, V.; Assies, L.; Sakai, N.; Roux, A.; Matile, S. Fluorescent Membrane Tension Probes for Early Endosomes. Angew. Chem. Int. Ed. 2021, 60, 12258–12263. [Google Scholar] [CrossRef]
- Garcia-Calvo, J.; Lopez-Andarias, J.; Maillard, J.; Mercier, V.; Roffay, C.; Roux, A.; Fuerstenberg, A.; Sakai, N.; Matile, S. HydroFlipper membrane tension probes: Imaging membrane hydration and mechanical compression simultaneously in living cells. Chem. Sci. 2022, 13, 2086–2093. [Google Scholar] [CrossRef]
- Chen, X.-X.; Bayard, F.; Gonzalez-Sanchis, N.; Pamungkas, K.; Sakai, N.; Matile, S. Fluorescent Flippers: Small-Molecule Probes to Image Membrane Tension in Living Systems. Angew. Chem. 2023, e202217868. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Probe | Analyte | λex/nm | λem/nm | Φ | Probe Concentration | LOD | Ref. |
|---|---|---|---|---|---|---|---|---|
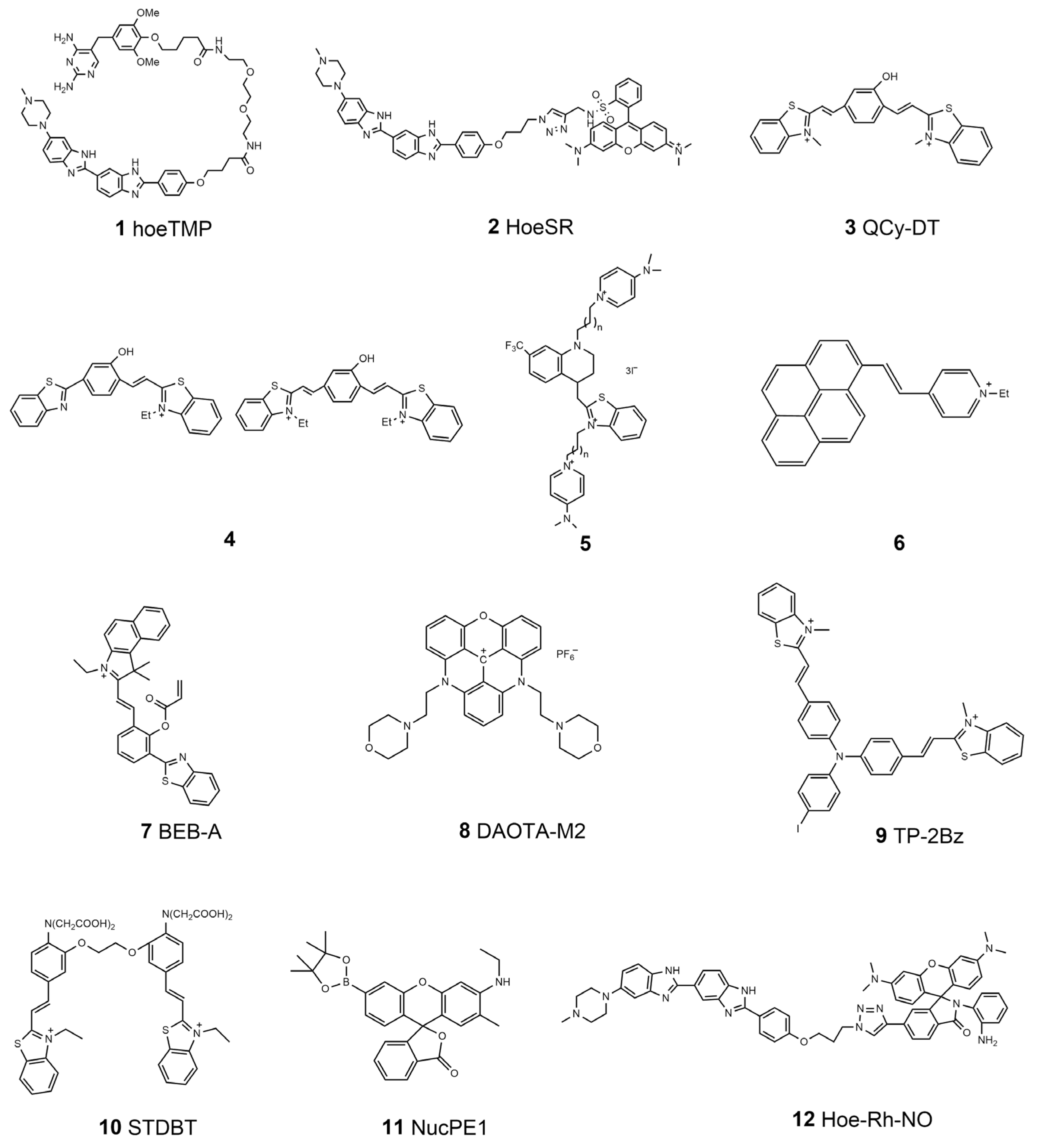
| 1 | hoeTMP | DNA | \ | \ | \ | \ | \ | [35] |
| 2 | HoeSR | DNA | 532 | 590 | 0.09 | \ | \ | [37] |
| 3 | \ | DNA | 530 | 680 | 0.32 | \ | \ | [38] |
| 4 | \ | DNA | 455, 492 | 612, 716 | 0.31, 0.41 | \ | \ | [40] |
| 5 | \ | dsDNA | 480 | 577, 567 | 0.022, 0.015 | \ | \ | [44] |
| 6 | \ | DNA | 477 | 610 | 0.42 | \ | \ | [42] |
| 7 | BEB-A | Cys | 543 | 616 | \ | 5 μM | 27 nM | [46] |
| 8 | DAOTA-M2 | G-quadruplexes | \ | \ | \ | \ | \ | [51] |
| 9 | TP-2Bz | G-quadruplexes | 488 | 600–670 | \ | \ | \ | [53] |
| 10 | STDBT | Ca2+ | 488 | 524–540 | \ | \ | \ | [54] |
| 11 | NucPE1 | H2O2 | 514 | 530 | 0.626 | \ | \ | [58] |
| 12 | Hoe-Rh-NO | NO | 405 | 463\603 | \ | 5 μM | 58 nM | [59] |
| Number | Probe Name | Analyte | λex/nm | λem/nm | Φ | Probe Concentration | LOD | Ref. |
|---|---|---|---|---|---|---|---|---|
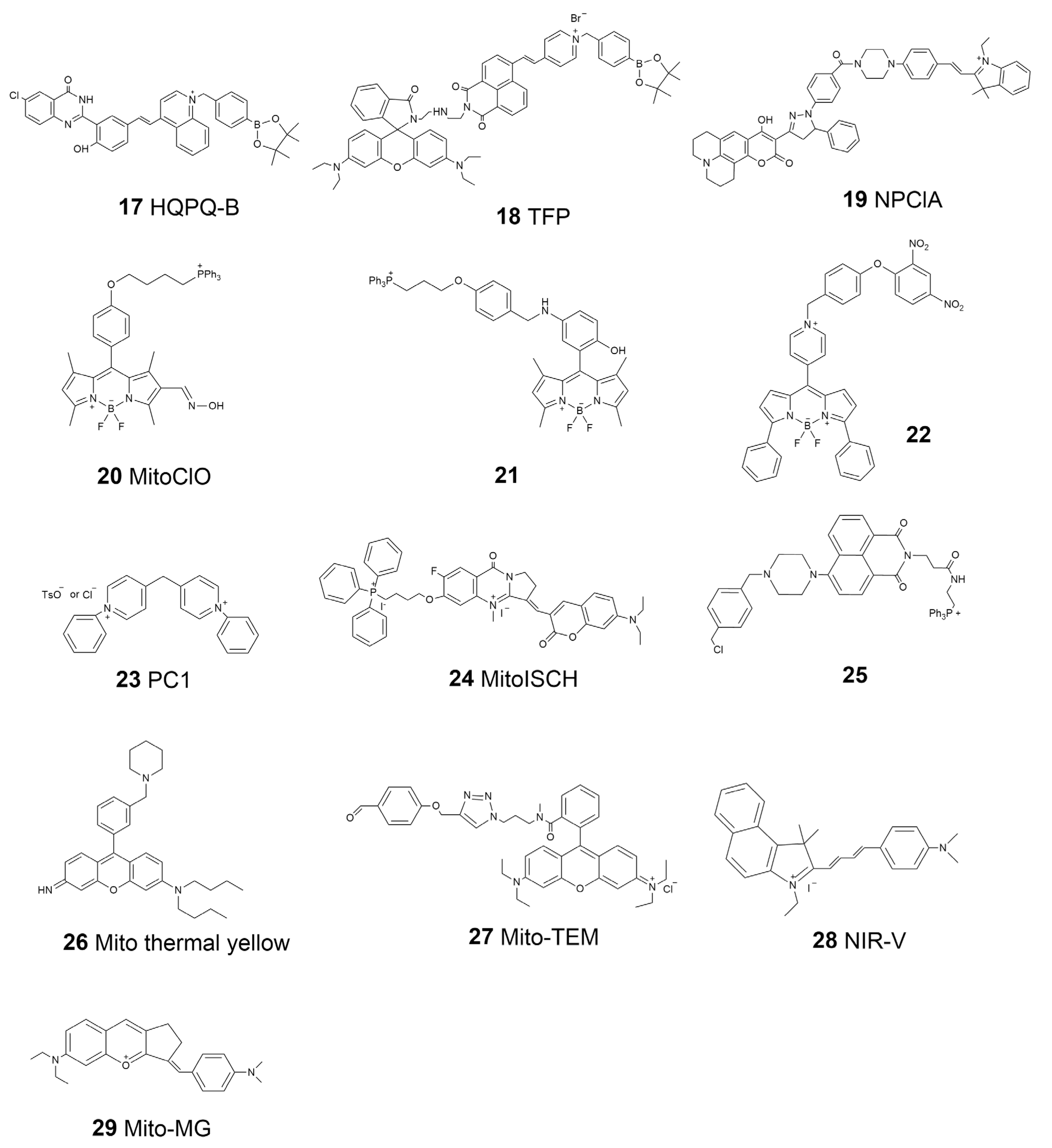
| 17 | HQPQ-B | H2O2 | 450 | 575 | \ | 10 μM | 1.76 μM | [83] |
| 18 | TFP | H2O2 ATP | 710 | 430–530 550–650 | 0.31 0.72 | 10 μM | 68 ± 5 nM 33 ± 2 μM | [84] |
| 19 | NPClA | SO2 ClO− | 395 | 482 425 | \ | 10 μM | 250 nM 16.6 nM | [85] |
| 20 | MitoClO | HClO | 480 | 529 | 0.44 | 1 μM | 0.52 μM | [86] |
| 21 | \ | NO | 488 | 500–600 | 0.11 | 4 μM | 4.8 nM | [95] |
| 22 | \ | GSH | 550 | 599 | 0.238 | 10 μM | 109 nM | [96] |
| 23 | PC1 | mtDNA | 532 | 546 | 0.09 | \ | \ | [99] |
| 24 | MitoISCH | mtDNA G4 | 560 | 580–800 | \ | \ | \ | [100] |
| 25 | \ | pH | 407 | 525 | \ | \ | \ | [102] |
| 26 | Mito thermal yellow | Temperature | \ | \ | \ | \ | \ | [103] |
| 27 | Mito-TEM | Temperature | 559 | 575–620 | \ | \ | \ | [104] |
| 28 | NIR-V | Viscosity | 580 | 700 | \ | \ | \ | [106] |
| 29 | Mito-MG | SO2 Viscosity | 675 | 675 770–781 | \ | 10 μM | 0.24 μM \ | [107] |
| Number | Probe Name | Analyte | λex/nm | λem/nm | Φ | Probe Concentration | LOD | Ref. |
|---|---|---|---|---|---|---|---|---|
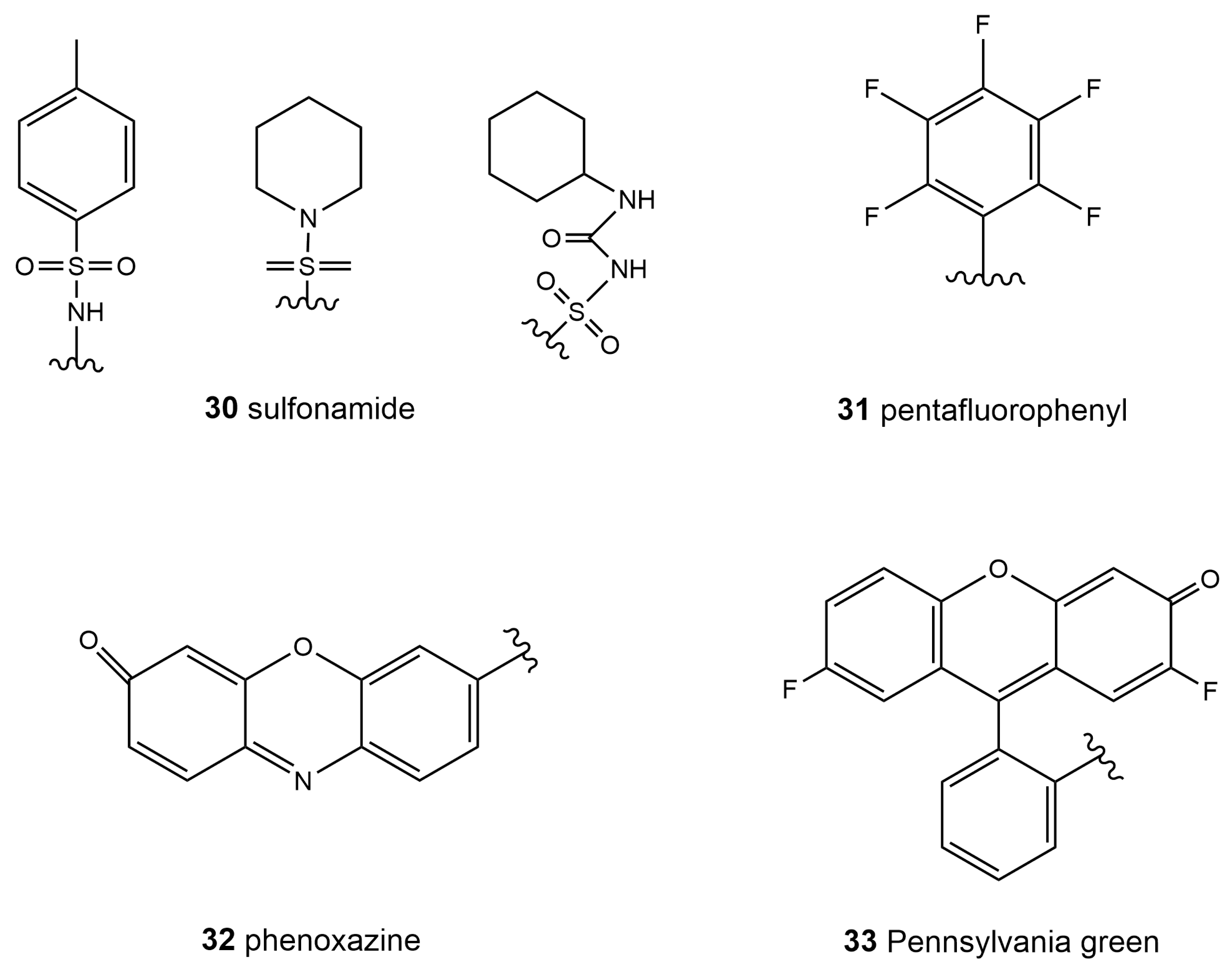
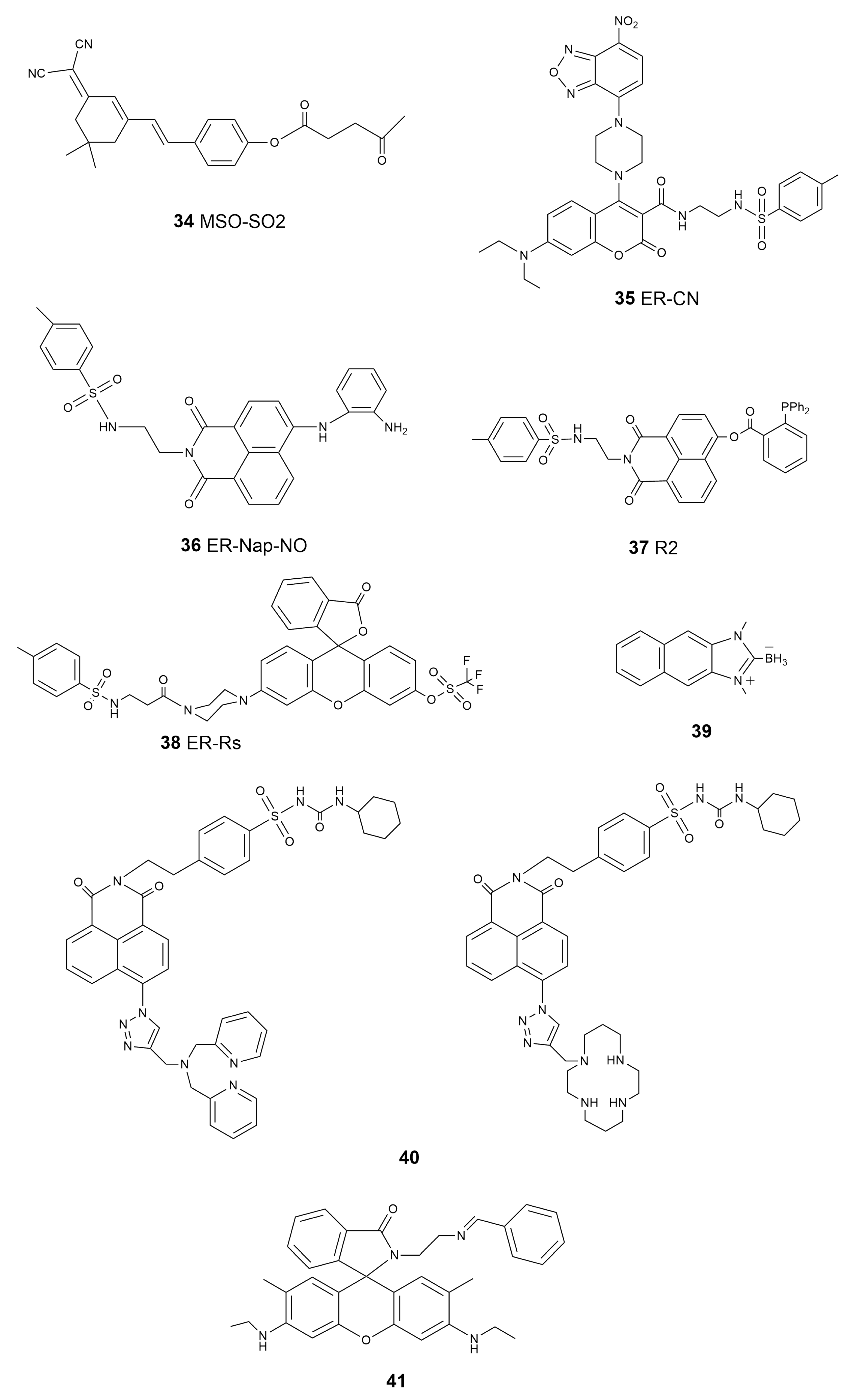
| 34 | MSO-SO2 | SO2 | 560 | 670–720 | \ | 10 μM | 12.3 nM | [126] |
| 35 | ER-CN | H2S | 405 | 470–510 | \ | 10 μM | 4.9 μM | [127] |
| 36 | ER-Nap-NO | NO | 440 | 538 | 0.214 | 10 μM | 3.3 nM | [129] |
| 37 | R2 | HNO | 450 | 555 | \ | 10 μM | 32 nM | [130] |
| 38 | ER-Rs | O2•− | 500 | 558 | 0.41 | 5 μM | 0.12 μM | [131] |
| 39 | \ | HClO | 326 | 450 | 0.29 | 10 μM | 3.6 μM | [132] |
| 40 | \ | Zn2+ | 346 | 414, 406 | 0.25 0.041 | 1 μM 1 μM | 47 pM 0.71 nM | [134] |
| 41 | \ | Fe3+ | 528 | 552 | \ | \ | \ | [135] |
| Number | Probe Name | Analyte | λex/nm | λem/nm | Φ | Probe Concentration | LOD | Ref. |
|---|---|---|---|---|---|---|---|---|
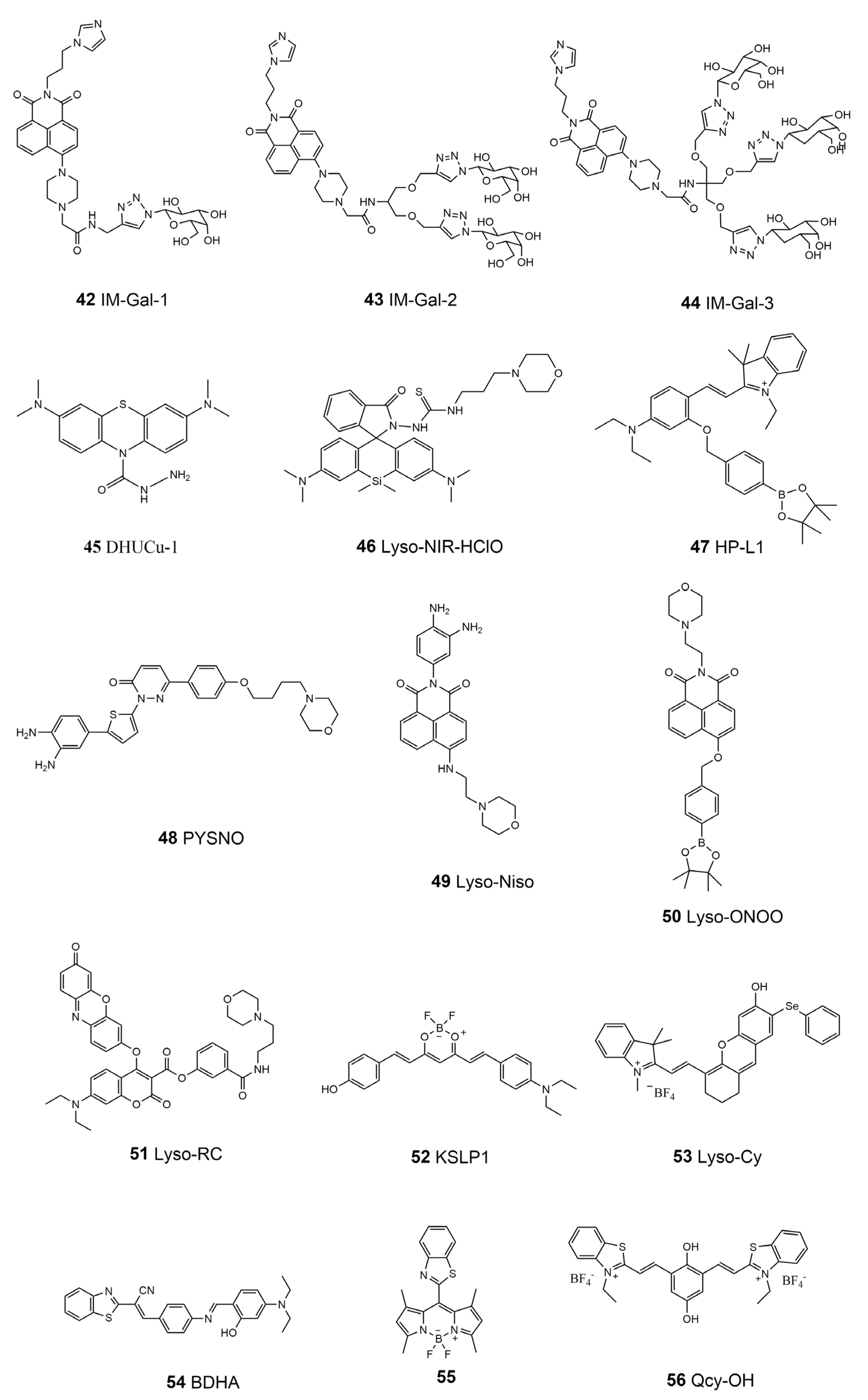
| 42 | IM-Gal-1 | Fe3+ | 405 | 510–550 | \ | \ | (129.3 ± 0.5) nM | |
| 43 | IM-Gal-2 | \ | (77.35 ± 0.05) nM | [150] | ||||
| 44 | IM-Gal-3 | \ | (132.5 ± 2.3) nM | |||||
| 45 | DHUCu-1 | Cu2+ | 620 | 686 | \ | 10 μM | 19.1 nM | [152] |
| 46 | Lyso-NIR-HClO | HClO | 635 | 680 | \ | 5 μM | 20 nM | [155] |
| 47 | HP-L1 | H2O2 | 520 | 584 | \ | 5 μM | 0.23 μM | [156] |
| 48 | PYSNO | NO | 405 | 515–565 | 0.28 | 5 μM | 242 nM | [157] |
| 49 | Lyso-Nino | NO | 440 | 520–560 | 0.3 | 5 μM | 5 nM | [158] |
| 50 | Lyso-ONOO | ONOO− | 450 | 555 | \ | 10 μM | 0.13 μM | [159] |
| 51 | Lyso-RC | Cys/HCy | 376 | 480 | \ | 10 μM | 27 nM/33 nM | |
| GSH | 438 | 542 | \ | 16 nM | [160] | |||
| H2S | 580 | 602 | \ | 0.38 μM | ||||
| 52 | KSLP1 | Polarity | 560 | 650–750 | \ | 10 μM | \ | [162] |
| 53 | Lyso-Cy | Viscosity | 635 | 685–785 | 0.41 | 5 μM | \ | [164] |
| 54 | BDHA | Viscosity | 457 | 626 | \ | 10 μM | \ | |
| ClO− | 405 | 570–620 | \ | 2.8 μM | [165] | |||
| 55 | \ | Viscosity | 488 | 520–600 | 0.329 | 10 μM | 0.38 cP | [166] |
| 56 | Qcy-OH | pH | 635 | 650–750 | 0.187 | 10 μM | \ | [167] |
| Number | Probe Name | Targeting Site | Analyte | λex/nm | λem/nm | Φ | Probe Concentration | LOD | Ref. |
|---|---|---|---|---|---|---|---|---|---|
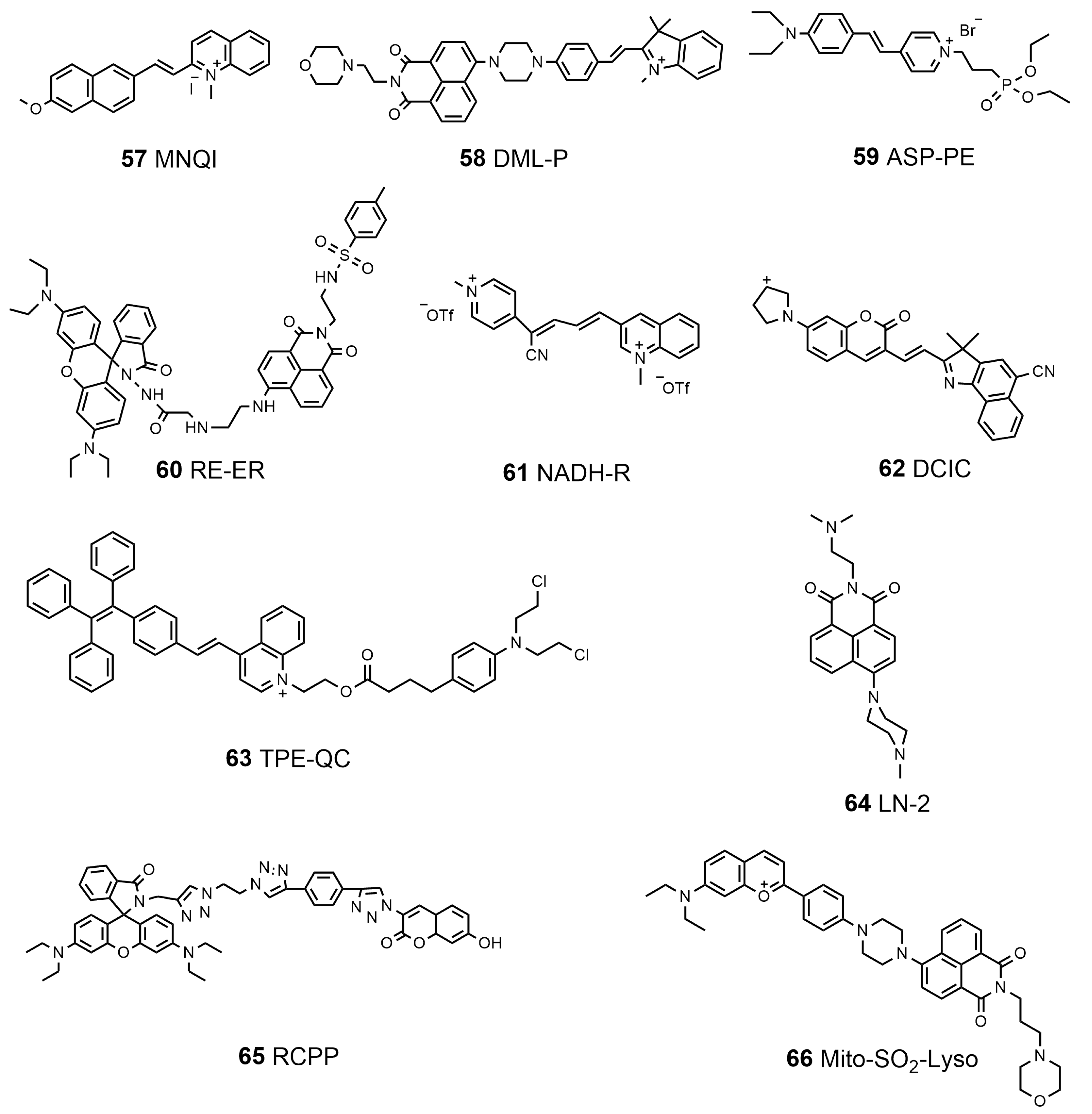
| 57 | MNQI | Mitochondria Nucleus | DNA | 488 | 570 (aq) 690 (s) | \ | \ | \ | [173] |
| 58 | DML-P | Mitochondria Lysosome | SO2 | 405 | 530 | 0.89 | 10 μM | 0.82 μM | [174] |
| 59 | ASP-PE | Mitochondria Plasma membrane | Membrane tension | 515 | 610 | \ | \ | \ | [175] |
| 60 | RT-ER | ER Lysosome | ClO− | 456 | 576 | 0.1245 | 10 μM | 3.37 μM | [176] |
| 61 | NADH-R | Mitochondria ER | NAD (P)H | 610 | 657 | \ | 5 μM | 12 nM | [177] |
| 62 | DCIC | Lysosome, mitochondria, ER Golgi apparatus | Viscosity | 500 | 630 | 0.1969 | 10 μM | 1.0 cp–438.4 cp | [178] |
| 63 | TEP-QC | Lysosome, mitochondria | Esterase | 488 | 520–620 | 0.126 | 10 μM | \ | [179] |
| 64 | LN-2 | Lysosome Nucleus | Cell death | 488 | 500–550 | \ | 1 μM | \ | [180] |
| 65 | RCPP | Mitochondria Lysosome | pH | 592.5 405 | 608–648 419–465 | \ | 10 μM | \ | [181] |
| 66 | Mito-SO2-Lyso | Mitochondria Lysosome | SO32− HSO3− | 470 | 570 650 | \ | 10 μM | 0.017 μM 0.3 μM | [182] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, S.; Dai, Z.; Cui, Y.; Kong, D.-M. Recent Development of Advanced Fluorescent Molecular Probes for Organelle-Targeted Cell Imaging. Biosensors 2023, 13, 360. https://doi.org/10.3390/bios13030360
Lu S, Dai Z, Cui Y, Kong D-M. Recent Development of Advanced Fluorescent Molecular Probes for Organelle-Targeted Cell Imaging. Biosensors. 2023; 13(3):360. https://doi.org/10.3390/bios13030360
Chicago/Turabian StyleLu, Sha, Zhiqi Dai, Yunxi Cui, and De-Ming Kong. 2023. "Recent Development of Advanced Fluorescent Molecular Probes for Organelle-Targeted Cell Imaging" Biosensors 13, no. 3: 360. https://doi.org/10.3390/bios13030360
APA StyleLu, S., Dai, Z., Cui, Y., & Kong, D.-M. (2023). Recent Development of Advanced Fluorescent Molecular Probes for Organelle-Targeted Cell Imaging. Biosensors, 13(3), 360. https://doi.org/10.3390/bios13030360