
Phytotoxicity and Accumulation of Copper-Based Nanoparticles in Brassica under Cadmium Stress



Abstract
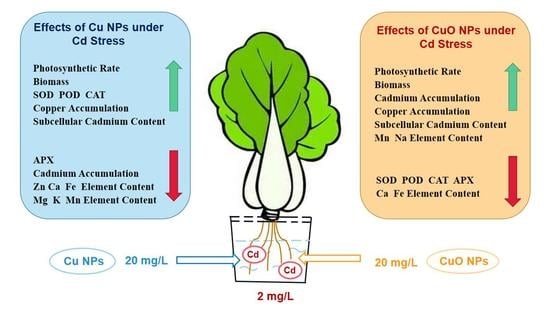
1. Introduction
2. Materials and Methods
2.1. Cu and CuO NPs Characterization
2.2. Hydroponic Experiment Design and Exposure Conditions
2.3. Determination of Physiological Indicators
2.4. Determination of Cu, Cd and Other Nutrient Elements
2.5. Determination of Subcellular Cu, Cd and Nutrient Elements
2.6. Determination of Antioxidant Enzyme Activity
2.7. Statistical Analysis
3. Results
3.1. Photosynthesis and Plant Growth
3.2. Antioxidase Activity
3.3. Uptake of Cu and Cd
3.4. Nutrient Element Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Xie, L.; Hao, P.; Cheng, Y.; Ahmed, I.M.; Cao, F. Effect of combined application of lead, cadmium, chromium and copper on grain, leaf and stem heavy metal contents at different growth stages in rice. Ecotoxicol. Environ. Saf. 2018, 162, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Lee, B.K. Influence of nano-TiO2 particles on the bioaccumulation of Cd in soybean plants (Glycine max): A possible mechanism for the removal of Cd from the contaminated soil. J. Environ. Manag. 2016, 170, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, C.; Tan, L.; Wang, J. Research progress in toxicity of nanomaterials manufactured on microalgae. Mar. Sci. 2017, 41, 134–143. [Google Scholar]
- Yuan, X.; Xue, N.; Han, Z. A meta-analysis of heavy metals pollution in farmland and urban soils in China over the past 20 years. J. Environ. Sci. 2021, 101, 217–226. [Google Scholar] [CrossRef]
- Keller, A.A.; McFerran, S.; Lazareva, A.; Suh, S. Global life cycle releases of engineered nanomaterials. J. Nanopart. Res. 2013, 15, 1692. [Google Scholar] [CrossRef]
- Kah, M.; Navarro, D.; Kookana, R.S.; Kirby, J.K.; Santra, S.; Ozcan, A.; Kabiri, S. Impact of (nano)formulations on the distribution and wash-off of copper pesticides and fertilisers applied on citrus leaves. Environ. Chem. 2019, 16, 401–410. [Google Scholar] [CrossRef]
- Marmiroli, M.; Pagano, L.; Rossi, R.; De La Torre-Roche, R.; Lepore, G.O.; Ruotolo, R.; Gariani, G.; Bonanni, V.; Pollastri, S.; Puri, A.; et al. Copper Oxide Nanomaterial Fate in Plant Tissue: Nanoscale Impacts on Reproductive Tissues. Environ. Sci. Technol. 2021, 55, 10769–10783. [Google Scholar] [CrossRef]
- Adhikari, T.; Sarkar, D.; Mashayekhi, H.; Xing, B. Growth and enzymatic activity of maize (Zea mays L.) plant: Solution culture test for copper dioxide nano particles. J. Plant Nutr. 2016, 39, 102–118. [Google Scholar] [CrossRef]
- Shaw, A.K.; Hossain, Z. Impact of nano-CuO stress on rice (Oryza sativa L.) seedlings. Chemosphere 2013, 93, 906–915. [Google Scholar] [CrossRef]
- Nair, P.M.G.; Chung, I.M. The responses of germinating seedlings of green peas to copper oxide nanoparticles. Biol. Plant. 2015, 59, 591–595. [Google Scholar] [CrossRef]
- Hong, J.; Rico, C.M.; Zhao, L.; Adeleye, A.S.; Keller, A.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Toxic effects of copper-based nanoparticles or compounds to lettuce (Lactuca sativa) and alfalfa (Medicago sativa). Environ. Sci.-Process Impacts 2015, 17, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Zuverza-Mena, N.; Medina-Velo, I.A.; Barrios, A.C.; Tan, W.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Copper nanoparticles/compounds impact agronomic and physiological parameters in cilantro (Coriandrum sativum). Environ. Sci.-Process Impacts 2015, 17, 1783–1793. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhao, L.; Huang, Y.; Qian, W.; Zhu, X.; Wang, Z.; Cai, Z. Combined toxicity of nano-TiO2 and Cd2+ to Scenedesmus obliquus: Effects at different concentration ratios. J. Hazard. Mater. 2021, 418, 126354. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, P.; Jayaraj, M.; Manikandan, R.; Geetha, N.; Rene, E.R.; Sharma, N.C.; Sahi, S.V. Zinc oxide nanoparticles (ZnONPs) alleviate heavy metal-induced toxicity in Leucaena leucocephala seedlings: A physiochemical analysis. Plant Physiol. Biochem. 2017, 110, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Ali, B.; Adrees, M.; Arshad, M.; Hussain, A.; Rehman, M.Z.U.; Waris, A.A. Zinc and iron oxide nanoparticles improved the plant growth and reduced the oxidative stress and cadmium concentration in wheat. Chemosphere 2019, 214, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Xu, L.N.; Zhao, J.; Wang, X.K.; White, J.C.; Xing, B.S. CuO Nanoparticle Interaction with Arabidopsis thaliana: Toxicity, Parent-Progeny Transfer, and Gene Expression. Environ. Sci. Technol. 2016, 50, 6008–6016. [Google Scholar] [CrossRef]
- Deng, F.; Wang, S.L.; Xin, H. Toxicity of CuO Nanoparticles to Structure and Metabolic Activity of Allium cepa Root Tips. Bull. Environ. Contam. Toxicol. 2016, 97, 702–708. [Google Scholar] [CrossRef]
- Zhao, L.J.; Huang, Y.X.; Zhou, H.J.; Adeleye, A.S.; Wang, H.T.; Ortiz, C.; Mazer, S.J.; Keller, A.A. GC-TOF-MS based metabolomics and ICP-MS based metallomics of cucumber (Cucumis sativus) fruits reveal alteration of metabolites profile and biological pathway disruption induced by nano copper. Environ.-Sci. Nano 2016, 3, 1114–1123. [Google Scholar] [CrossRef]
- Li, H.; Luo, N.; Zhang, L.J.; Zhao, H.M.; Li, Y.W.; Cai, Q.Y.; Wong, M.H.; Mo, C.H. Do arbuscular mycorrhizal fungi affect cadmium uptake kinetics, subcellular distribution and chemical forms in rice? Sci. Total Environ. 2016, 571, 1183–1190. [Google Scholar] [CrossRef]
- Tandy, N.E.; Giulio, R.T.; Richardson, C.J. Assay and Electrophoresis of Superoxide Dismutase from Red Spruce (Picea rubens Sarg.), Loblolly Pine (Pinus taeda L.), and Scotch Pine (Pinus sylvestris L.) A Method for Biomonitoring. Plant Physiol. 1989, 90, 742–748. [Google Scholar] [CrossRef]
- Jurkow, R.; Skara, A.; Pokluda, R.; Smoleń, S.; Kalisz, A. Biochemical response of oakleaf lettuce seedlings to different concentrations of some metal(oid) oxide nanoparticles. Agronomy 2020, 10, 997. [Google Scholar] [CrossRef]
- Shen, S.L.; Xia, B.C.; Xue, F. Cadmium effects on photosynthetic system of sauropus androgynus as a potential cd-accumulator. Fresenius Environ. Bull. 2012, 21, 3551–3557. [Google Scholar]
- Li, P.H.; Lin, Q.; Xu, G.Z. Effects of pb and cd stress on the photosynthetic physiological characters of potato in heavy metal pollution of soil. Appl. Ecol. Environ. Res. 2019, 17, 12287–12295. [Google Scholar] [CrossRef]
- Li, D.; Zhu, Z.; Liu, Y.; Wang, Y. Influence of cadmium on photosynthesis of Brassica campestris subsp. chinensis L. J. Zhejiang Univ. 2005, 31, 459–464. [Google Scholar]
- Servin, A.; Elmer, W.; Mukherjee, A.; De la Torre-Roche, R.; Hamdi, H.; White, J.C.; Bindraban, P.; Dimkpa, C. A review of the use of engineered nanomaterials to suppress plant disease and enhance crop yield. J. Nanopart. Res. 2015, 17, 1–21. [Google Scholar] [CrossRef]
- Govorov, A.O.; Carmeli, I. Hybrid structures composed of photosynthetic system and metal nanoparticles: Plasmon enhancement effect. Nano Lett. 2007, 7, 620–625. [Google Scholar] [CrossRef]
- Pradhan, S.; Patra, P.; Mitra, S.; Dey, K.K.; Basu, S.; Chandra, S.; Palit, P.; Goswami, A. Copper Nanoparticle (CuNP) Nanochain Arrays with a Reduced Toxicity Response: A Biophysical and Biochemical Outlook on Vigna radiata. J. Agric. Food Chem. 2015, 63, 2606–2617. [Google Scholar] [CrossRef]
- Nuzahat, H.; Zhang, Y.; Asiya, K.; Abdurexit, A. Influence of Different Zinc and Copper Levels on Cadmium Accumulation and Physiological Characterization in Vetiveria zizanioides L. J. Inn. Mong. Univ. 2012, 43, 543–550. [Google Scholar]
- Aibibu, N.; Liu, Y.; Zeng, G.; Wang, X.; Chen, B.; Song, H.; Xu, L. Cadmium accumulation in vetiveria zizanioides and its effects on growth, physiological and biochemical characters. Bioresour. Technol. 2010, 101, 6297–6303. [Google Scholar] [CrossRef]
- Mir, A.R.; Pichtel, J.; Hayat, S. Copper: Uptake, toxicity and tolerance in plants and management of Cu-contaminated soil. Biometals 2021, 34, 737–759. [Google Scholar] [CrossRef]
- Zhou, J.; Cheng, K.; Zheng, J.; Liu, Z.; Shen, W.; Fan, H.; Jin, Z. Physiological and Biochemical Characteristics of Cinnamomum camphora in Response to Cu- and Cd-Contaminated Soil. Water Air Soil Pollut. 2019, 230, 1–11. [Google Scholar] [CrossRef]
- Karlsson, H.L.; Gustafsson, J.; Cronholm, P.; Moller, L. Size-dependent toxicity of metal oxide particles-A comparison between nano- and micrometer size. Toxicol. Lett. 2009, 188, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicka, M.; Obidzińska, J. The effect of lead on seed imbibition and germination in different plant species. Plant Sci. 1998, 137, 155–171. [Google Scholar] [CrossRef]
- Van, L.; Ma, C.; Shang, J.; Rui, Y.; Liu, S.; Xing, B. Effects of CuO nanoparticles on insecticidal activity and phytotoxicity in conventional and transgenic cotton. Chemosphere 2016, 144, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Santala, K.R.; Ryser, P. Influence of heavy-metal contamination on plant response to water availability in white birch, Betula papyrifera. Environ. Exp. Bot. 2009, 66, 334–340. [Google Scholar] [CrossRef]
- Ahmed, B.; Rizvi, A.; Zaidi, A.; Khan, M.S.; Musarrat, J. Understanding the phyto-interaction of heavy metal oxide bulk and nanoparticles: Evaluation of seed germination, growth, bioaccumulation, and metallothionein production. RSC Adv. 2019, 9, 4210–4225. [Google Scholar] [CrossRef]
- Dietz, K.J.; Herth, S. Plant nanotoxicology (vol 16, pg 582, 2011). Trends Plant Sci. 2012, 17, 180. [Google Scholar] [CrossRef]
- Khatun, S.; Ali, M.B.; Hahn, E.J.; Paek, K.Y.; Botany, E. Copper toxicity in Withania somnifera: Growth and antioxidant enzymes responses of in vitro grown plants. Environ. Exp. Bot. 2008, 64, 279–285. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; VanMontagu, M.; Inze, D.; VanCamp, W. Catalase is a sink for H2O2 and is indispensable for stress defence in C-3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef]
- Karimi, P.; Khavari-Nejad, R.A.; Niknam, V.; Ghahremaninejad, F.; Najafi, F. The Effects of Excess Copper on Antioxidative Enzymes, Lipid Peroxidation, Proline, Chlorophyll, and Concentration of Mn, Fe, and Cu in Astragalus neo-mobayenii. Sci. World J. 2012, 2012, 615670. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, S.; Lee, S.; Lee, I. Alteration of Phytotoxicity and Oxidant Stress Potential by Metal Oxide Nanoparticles in Cucumis sativus. Water Air Soil Pollut. 2012, 223, 2799–2806. [Google Scholar] [CrossRef]
- Li, Z.; Li, J.; Zhang, G. Expression regulation of plant ascorbate peroxidase and its tolerance to abiotic stresses. Hereditas 2013, 35, 45–54. [Google Scholar] [CrossRef]
- Trujillo-Reyes, J.; Majumdar, S.; Botez, C.E.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Exposure studies of core-shell Fe/Fe3O4 and Cu/CuO NPs to lettuce (Lactuca sativa) plants: Are they a potential physiological and nutritional hazard? J. Hazard. Mater. 2014, 267, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Franklin, N.M.; Rogers, N.J.; Apte, S.C.; Batley, G.E.; Gadd, G.E.; Casey, P.S. Comparative toxicity of nanoparticulate ZnO, bulk ZnO, and ZnCl2 to a freshwater microalga (Pseudokirchneriella subcapitata): The importance of particle solubility. Environ. Sci. Technol. 2007, 41, 8484–8490. [Google Scholar] [CrossRef]
- Shi, J.; Peng, C.; Yang, Y.; Yang, J.; Zhang, H.; Yuan, X.; Chen, Y.; Hu, T. Phytotoxicity and accumulation of copper oxide nanoparticles to the Cu-tolerant plant Elsholtzia splendens. Nanotoxicology 2014, 8, 179–188. [Google Scholar] [CrossRef]
- Miralles, P.; Church, T.L.; Harris, A.T. Toxicity, Uptake, and Translocation of Engineered Nanomaterials in Vascular plants. Environ. Sci. Technol. 2012, 46, 9224–9239. [Google Scholar] [CrossRef]
- Kadri, O.; Karmous, I.; Kharbech, O.; Arfaoui, H.; Chaoui, A. Cu and CuO Nanoparticles Affected the Germination and the Growth of Barley (Hordeum vulgare L.) Seedling. Bull. Environ. Contam. Toxicol. 2022, 108, 585–593. [Google Scholar] [CrossRef]
- Verkleij, J.A.C.; Golan-Goldhirsh, A.; Antosiewisz, D.M.; Schwitzguébel, J.; Schröder, P. Dualities in plant tolerance to pollutants and their uptake and translocation to the upper plant parts. Environ. Exp. Bot. 2009, 67, 10–22. [Google Scholar] [CrossRef]
- Mishra, V.K.; Tripathi, B.D. Concurrent removal and accumulation of heavy metals by the three aquatic macrophytes. Bioresour. Technol. 2008, 99, 7091–7097. [Google Scholar] [CrossRef]
- Cvjetko, P.; Tolic, S.; Sikic, S.; Balen, B.; Tkale, M.; Vidakovic-Cifrek, Z.; Pavlica, M. Effect of copper on the toxicity and genotoxicity of cadmium in duckweed (Lemna minor L.). Arh. Hig. Rada. Toksikol. 2010, 61, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, Y.; Kim, E.J.; Gu, S.; Sohn, E.J.; Seo, Y.S.; An, H.J.; Chang, Y.S. Exposure of Iron Nanoparticles to Arabidopsis thaliana Enhances Root Elongation by Triggering Cell Wall Loosening. Environ. Sci. Technol. 2014, 48, 3477–3485. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; White, J.C.; Dhankher, O.P.; Xing, B. Metal-Based Nanotoxicity and Detoxification Pathways in Higher Plants. Environ. Sci. Technol. 2015, 49, 7109–7122. [Google Scholar] [CrossRef] [PubMed]
- Luan, Z.Q.; Cao, H.C.; Yan, B.X. Individual and combined phytotoxic effects of cadmium, lead and arsenic on soybean in Phaeozem. Plant Soil Environ. 2008, 54, 403–411. [Google Scholar] [CrossRef]
- Majsec, K.; Cvjetko, P.; Tolic, S.; Tkalec, M.; Balen, B.; Pavlica, M. Integrative approach gives new insights into combined Cd/Cu exposure in tobacco. Acta Physiol. Plant. 2016, 38, 1–14. [Google Scholar] [CrossRef]
- Weng, B.S.; Xie, X.Y.; Weiss, D.J.; Liu, J.C.; Lu, H.L.; Yan, C.L. Kandelia obovata (S., L.) Yong tolerance mechanisms to Cadmium: Subcellular distribution, chemical forms and thiol pools. Mar. Pollut. Bull. 2012, 64, 2453–2460. [Google Scholar] [CrossRef]
- Rondeau-Mouro, C.; Defer, D.; Leboeuf, E.; Lahaye, M. Assessment of cell wall porosity in Arabidopsis thaliana by NMR spectroscopy. Int. J. Biol. Macromol. 2008, 42, 83–92. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, X.; Wang, M.; Wang, J.; Yan, X. Cadmium tolerance, accumulation and relationship with Cd subcellular distribution in Ricinus communis L. Acta Sci. Circumstantiae 2014, 34, 2440–2446. [Google Scholar]
- Printz, B.; Lutts, S.; Hausman, J.F.; Sergeant, K. Copper Trafficking in Plants and Its Implication on Cell Wall Dynamics. Front. Plant Sci. 2016, 7, 601. [Google Scholar] [CrossRef]
- Roy, S.K.; Cho, S.W.; Kwon, S.J.; Kamal, A.H.M.; Lee, D.G.; Sarker, K.; Lee, M.S.; Xin, Z.; Woo, S.H. Proteome characterization of copper stress responses in the roots of sorghum. Biometals 2017, 30, 765–785. [Google Scholar] [CrossRef]
- Cambrolle, J.; Garcia, J.L.; Figueroa, M.E.; Cantos, M. Evaluating wild grapevine tolerance to copper toxicity. Chemosphere 2015, 120, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, Z.A.; Khan, M.R.; Abd_Allah, E.F.; Aiman, P.; Parveen, A. Titanium dioxide and zinc oxide nanoparticles affect some bacterial diseases, and growth and physiological changes of beetroot. J. Veg. Sci. 2019, 25, 409–430. [Google Scholar] [CrossRef]
- Zhang, K.; Tong, H.; Huang, S.; Yuan, H. Effect of Cu stress on Cu accumulation and other nutrient element absorption of Iris pseudacaorus and I. lactea var. chinensis. J. Plant Resour. Environ. 2007, 16, 18–22. [Google Scholar]
- DalCorso, G.; Manara, A.; Piasentin, S.; Furini, A. Nutrient metal elements in plants. Metallomics 2014, 6, 1770–1788. [Google Scholar] [CrossRef] [PubMed]
- Moral, R.; Gomez, I.; Navarro-Pedreno, J.; Mataix, J. Effects of cadmium on nutrient distribution, yield, and growth of tomato grown in soilless culture. J. Plant Nutr. 1994, 17, 953–962. [Google Scholar] [CrossRef]
- Gajewska, E.; Glowacki, R.; Mazur, J.; Sklodowska, M. Differential response of wheat roots to Cu, Ni and Cd treatment: Oxidative stress and defense reactions. Plant Growth Regul. 2013, 71, 13–20. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| D1 | D2 | D3 | D4 | D5 | D6 | D7 | |
|---|---|---|---|---|---|---|---|
| Cd | 39.68 ± 1.83 b | 38.79 ± 0.65 c | 38.80 ± 3.22 c | 38.76 ± 3.16 b | 37.91 ± 1.48 c | 38.59 ± 1.82 c | 38.46 ± 3.23 c |
| Cu NPs | 41.37 ± 4.19 b | 43.76 ± 2.11 ab | 44.26 ± 1.61 ab | 44.53 ± 4.84 ab | 44.15 ± 2.74 b | 44.53 ± 1.28 b | 44.60 ± 2.28 ab |
| CuO NPs | 47.30 ± 2.74 a | 47.06 ± 3.13 a | 47.57 ± 1.42 a | 48.14 ± 5.41 a | 48.42 ± 1.67 a | 49.31 ± 2.61 a | 49.86 ± 3.77 a |
| Cu NPs + Cd | 40.87 ± 1.14 b | 41.23 ± 1.22 bc | 41.54 ± 1.40 bc | 42.44 ± 1.88 ab | 42.54 ± 0.70 b | 42.90 ± 1.80 b | 42.76 ± 1.47 bc |
| CuO NPs + Cd | 43.96 ± 3.51 ab | 43.48 ± 1.24 ab | 43.44 ± 2.56 ab | 43.51 ± 2.04 ab | 43.42 ± 1.53 b | 43.45 ± 2.25 b | 45.98 ± 3.34 ab |
| Nutrition Elements | Cd | Cu NPs | CuO NPs | Cu NPs + Cd | CuO NPs + Cd | |
|---|---|---|---|---|---|---|
| Zn | Leaves | 90.54 ± 20.71 a | 79.28 ± 14.20 ab | 71.15 ± 7.23 ab | 62.87 ± 6.01 b | 69.45 ± 6.52 ab |
| Roots | 80.30 ± 11.96 b | 82.60 ± 3.43 b | 98.59 ± 8.42 ab | 91.28 ± 8.32 ab | 121.82 ± 37.70 a | |
| Ca | Leaves | 82.83 ± 0.45 a | 74.54 ± 1.86 b | 73.57 ± 4.32 b | 63.90 ± 4.52 c | 68.35 ± 6.98 bc |
| Roots | 77.28 ± 5.16 b | 87.23 ± 2.53 ab | 90.09 ± 4.50 a | 91.03 ± 8.78 a | 89.30 ± 5.32 a | |
| Fe | Leaves | 5.77 ± 0.03 a | 2.76 ± 0.21 c | 2.99 ± 0.18 bc | 3.08 ± 0.07 bc | 3.27 ± 0.22 b |
| Roots | 98.04 ± 7.98 b | 99.66 ± 5.10 b | 147.11 ± 28.01 a | 130.31 ± 19.82 ab | 114.49 ± 18.89 ab | |
| Mg | Leaves | 108.24 ± 4.42 a | 88.25 ± 3.10 b | 84.02 ± 3.23 b | 83.87 ± 1.62 b | 110.48 ± 3.55 a |
| Roots | 55.16 ± 3.31 c | 53.72 ± 2.17 c | 85.38 ± 10.34 a | 80.46 ± 8.76 ab | 72.38 ± 1.34 b | |
| K | Leaves | 945.79 ± 51.23 ab | 911.33 ± 42.53 b | 949.09 ± 26.97 ab | 745.87 ± 22.82 c | 1001.68 ± 46.02 a |
| Roots | 474.67 ± 43.25 c | 493.99 ± 10.67 c | 774.41 ± 79.53 a | 602.24 ± 67.05 b | 697.33 ± 43.84 ab | |
| Mn | Leaves | 118.64 ± 1.80 b | 116.79 ± 7.25 b | 97.33 ± 4.48 d | 109.15 ± 2.02 c | 147.43 ± 2.11 a |
| Roots | 40.12 ± 4.71 cd | 34.86 ± 0.79 d | 64.93 ± 14.16 a | 49.12 ± 5.01 bc | 54.35 ± 4.94 ab | |
| Na | Leaves | 21.22 ± 0.80 b | 21.45 ± 0.91 b | 18.07 ± 0.56 c | 22.13 ± 0.28 b | 27.25 ± 1.06 a |
| Roots | 93.52 ± 6.34 b | 101.82 ± 5.99 ab | 103.50 ± 12.14 ab | 92.38 ± 7.96 b | 108.56 ± 1.90 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Fu, Y.; Zheng, S.; Xu, Y.; Sun, Y. Phytotoxicity and Accumulation of Copper-Based Nanoparticles in Brassica under Cadmium Stress. Nanomaterials 2022, 12, 1497. https://doi.org/10.3390/nano12091497
Wang S, Fu Y, Zheng S, Xu Y, Sun Y. Phytotoxicity and Accumulation of Copper-Based Nanoparticles in Brassica under Cadmium Stress. Nanomaterials. 2022; 12(9):1497. https://doi.org/10.3390/nano12091497
Chicago/Turabian StyleWang, Shiqi, Yutong Fu, Shunan Zheng, Yingming Xu, and Yuebing Sun. 2022. "Phytotoxicity and Accumulation of Copper-Based Nanoparticles in Brassica under Cadmium Stress" Nanomaterials 12, no. 9: 1497. https://doi.org/10.3390/nano12091497
APA StyleWang, S., Fu, Y., Zheng, S., Xu, Y., & Sun, Y. (2022). Phytotoxicity and Accumulation of Copper-Based Nanoparticles in Brassica under Cadmium Stress. Nanomaterials, 12(9), 1497. https://doi.org/10.3390/nano12091497