
Management Strategies for Ulex europaeus L. Control in a Native Plant Community in Tenerife, Canary Islands

 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. The Species
2.3. Sampling Design
2.4. Statistical Analyses
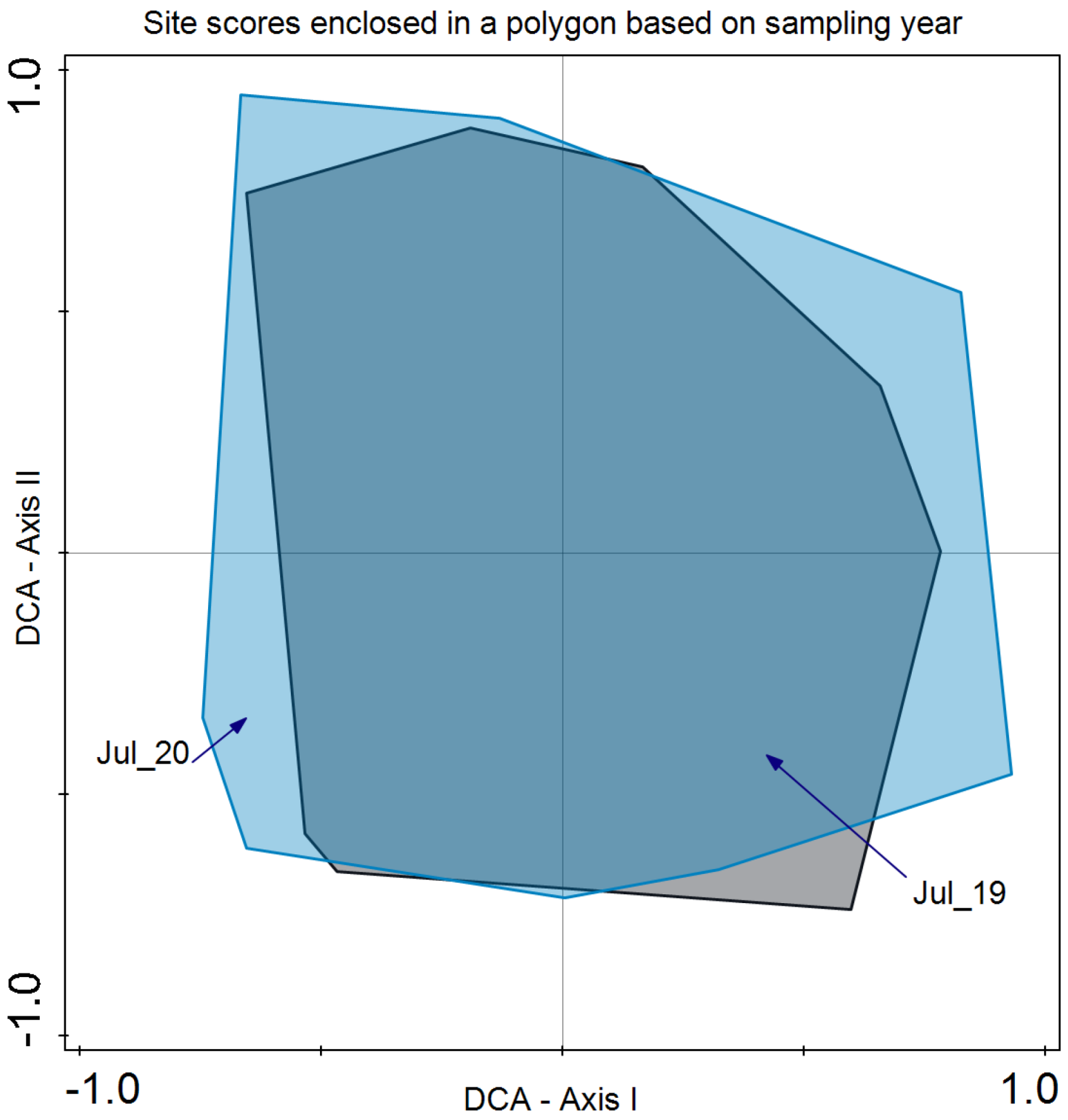
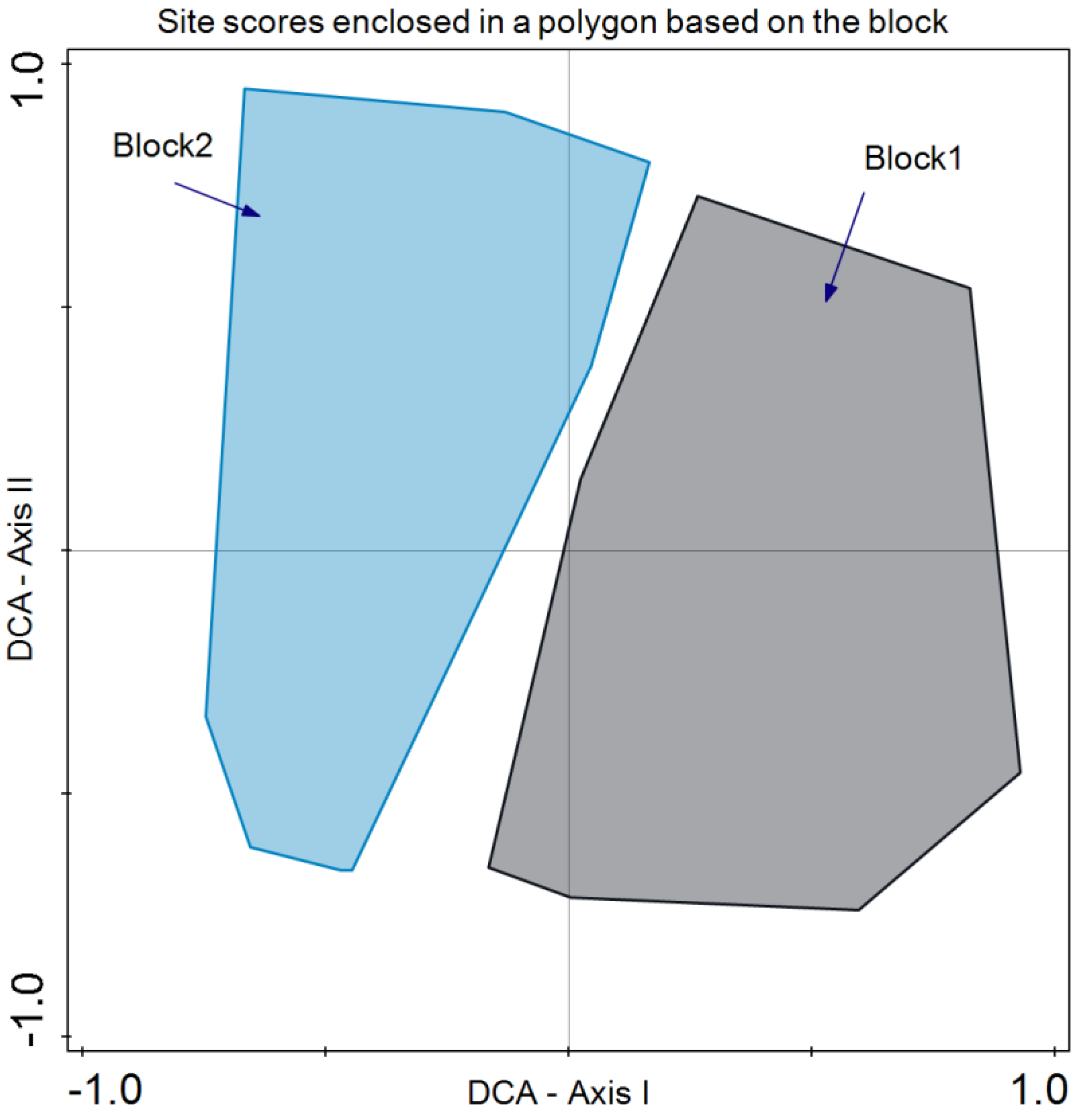
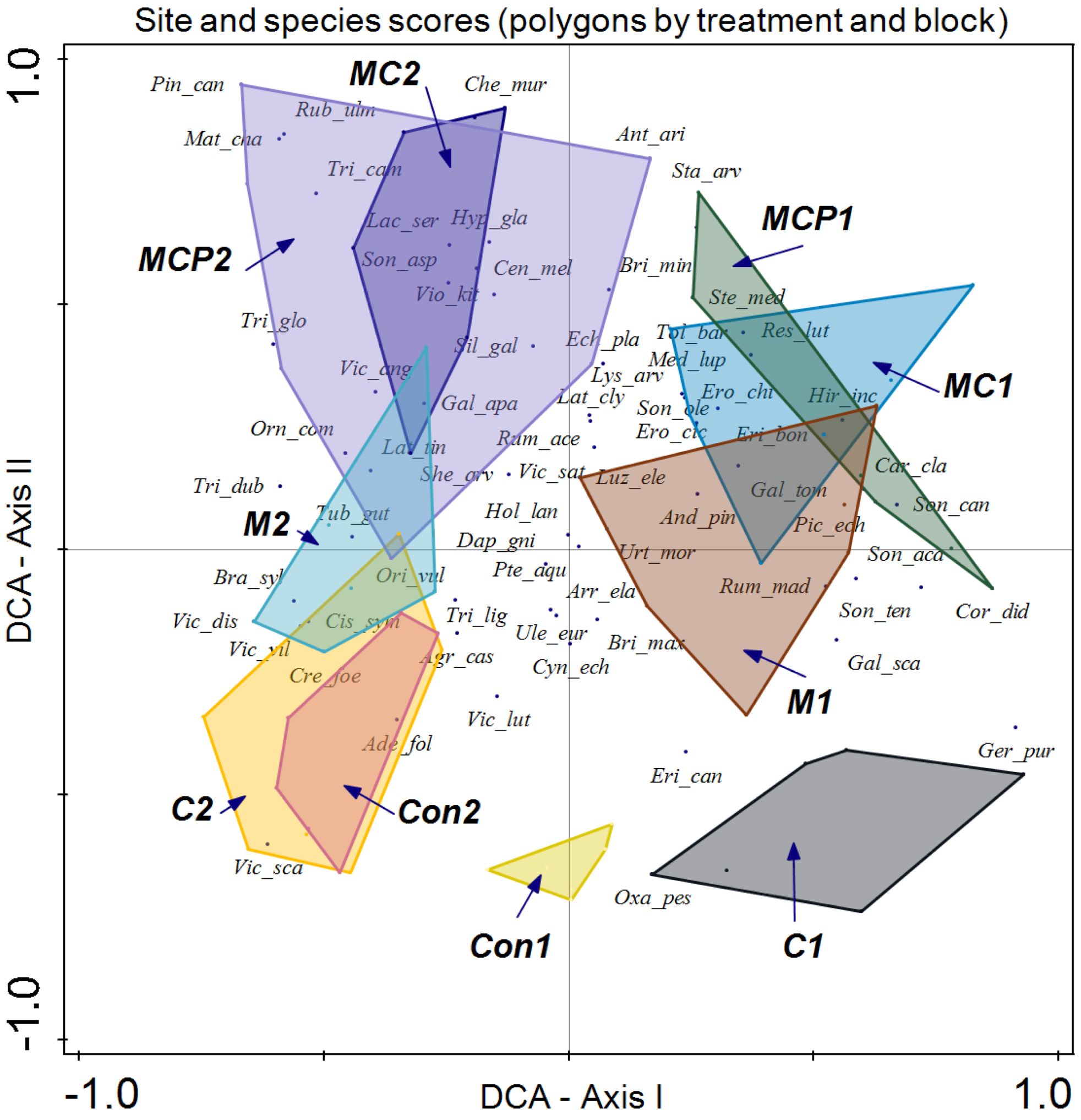
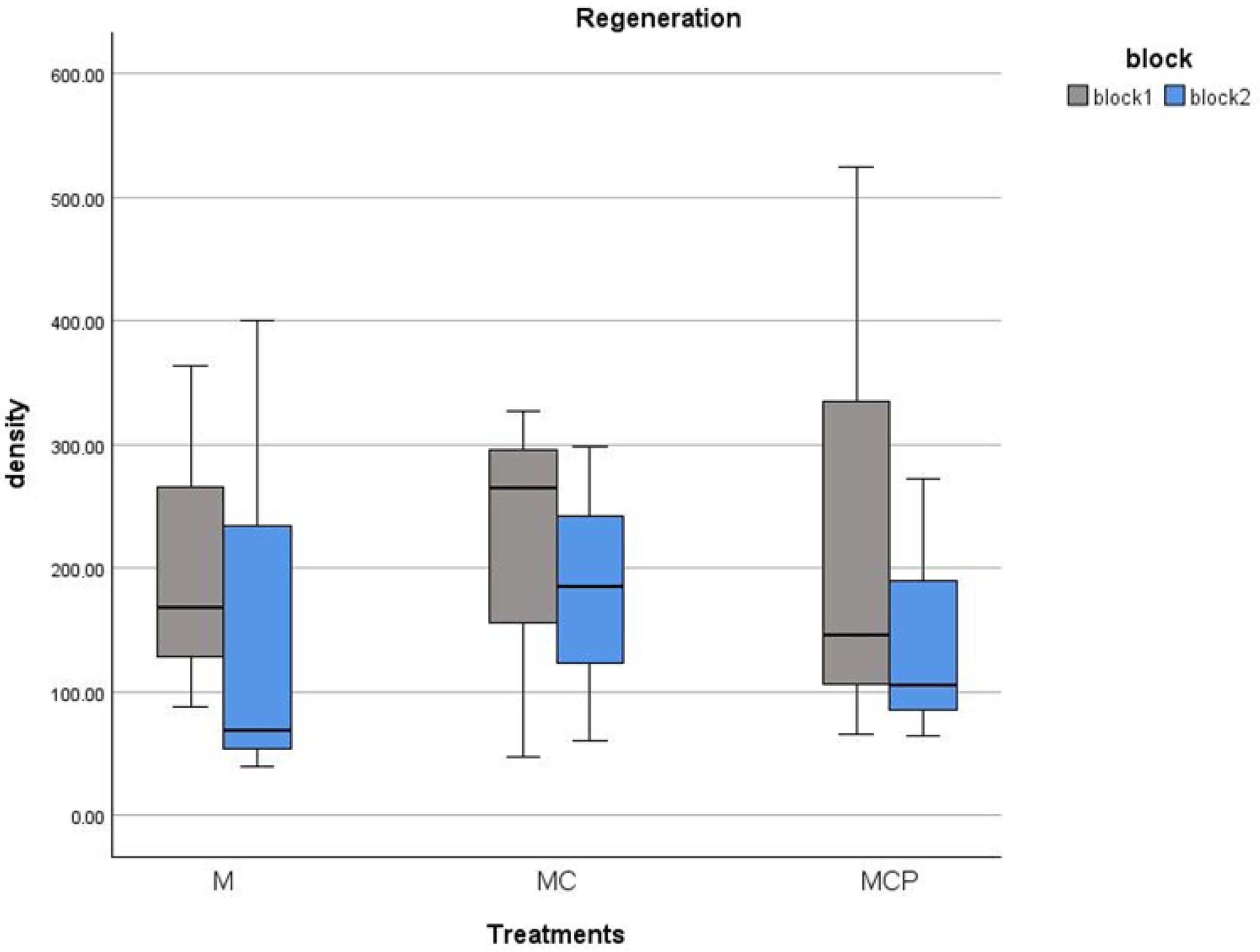
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Family | Endemism | Origin 1 | Raunkiaer Biotypes | Life Form |
|---|---|---|---|---|---|
| Adenocarpus foliolosus (Aiton) DC. | Fabaceae | Yes | SN | Nanophanerophyte | Shrub |
| Agrostis castellana Boiss. & Reut. | Poaceae | No | PN | Hemicryptophyte | Perennial herb |
| Aira caryophyllea L. | Poaceae | No | PN | Therophyte | Herb |
| Andryala pinnatifida Aiton | Asteraceae | No | SN | Therophyte | Herb |
| Anogramma leptophylla (L.) Link | Hemionitidaceae | No | SN | Therophyte | Herb |
| Anthoxanthum aristatum Boiss. | Poaceae | No | PN | Therophyte | Herb |
| Apiaceae sp. | Apiaceae | ||||
| Arrhenatherum elatius (L.) P. Beauv. ex J. Presl & C. Presl subsp. bulbosum (Willd.) Schübl. & Mart. | Poaceae | No | PN | Hemicryptophyte | Perennial herb |
| Brachypodium sylvaticum (Huds.) P. Beauv. | Poaceae | No | PN | Therophyte | Herb |
| Briza maxima L. | Poaceae | No | PN | Therophyte | Herb |
| Briza minor L. | Poaceae | No | PN | Therophyte | Herb |
| Bromus diandrus Roth | Poaceae | No | PN | Therophyte | Herb |
| Bromus hordeaceus L. | Poaceae | No | PN | Therophyte | Herb |
| Calendula arvensis (Vaill.) L. | Asteraceae | No | SIN | Therophyte | Herb |
| Carduus clavulatus Link | Asteraceae | Yes | SN | Therophyte | Herb |
| Carduus tenuiflorus Curtis | Asteraceae | No | PN | Hemicryptophyte | Perennial herb |
| Centaurea melitensis L. | Asteraceae | No | PN | Therophyte | Herb |
| Centranthus calcitrapae (L.) Dufr. | Valerianaceae | No | PN | Therophyte | Herb |
| Cerastium glomeratum Thuill. | Caryophyllaceae | No | PN | Therophyte | Herb |
| Chenopodiastrum murale (L.) S. Fuentes, Uotila & Borsch | Chenopodiaceae | No | PI | Therophyte | Herb |
| Cistus symphytifolius Lam. | Cistaceae | Yes | SN | Nanophanerophyte | Shrub |
| Lepidium didymum L. | Brassicaceae | No | PI | Hemicryptophyte | Perennial herb |
| Cotula australis (Spreng.) & Hook. f. | Asteraceae | No | SIN | Therophyte | Herb |
| Crepis foetida L. | Asteraceae | No | PN | Hemicryptophyte | Perennial herb |
| Cynosurus echinatus L. | Poaceae | No | PN | Therophyte | Herb |
| Daphne gnidium L. | Thymelaeaceae | No | SN | Chamaephyte | Shrub |
| Echium plantagineum L. | Boraginaceae | No | PN | Hemicryptophyte | Perennial herb |
| Erica canariensis Rivas-Mart., M. Osorio & Wildpret | Ericaceae | No | SN | Microphanerophyte | Large shrub—tree |
| Erigeron bonariensis L. | Asteraceae | No | SIN | Therophyte | Herb |
| Erodium botrys (Cav.) Bertol. | Geraniaceae | No | PN | Therophyte | Herb |
| Erodium chium (L.) Willd. | Geraniaceae | No | PN | Therophyte | Herb |
| Erodium cicutarium (L.) L’Hér. in Aiton | Geraniaceae | No | PN | Therophyte | Herb |
| Euphorbia peplus L. | Euphorbiaceae | No | PN | Therophyte | Herb |
| Filago pyramidata L. | Asteraceae | No | PN | Therophyte | Herb |
| Fumaria cf. bastardii Boreau | Fumariaceae | No | PN | Therophyte | Herb |
| Fumaria montana J.A. Schmidt | Fumariaceae | No | SN | Therophyte | Herb |
| Fumaria sp. | Fumariaceae | Therophyte | Herb | ||
| Galactites tomentosus Moench | Asteraceae | No | PN | Therophyte | Herb |
| Galium aparine L. | Rubiaceae | No | PN | Therophyte | Herb |
| Galium parisiense L. | Rubiaceae | No | PN | Therophyte | Herb |
| Galium scabrum L. | Rubiaceae | No | PN | Chamaephyte | Small shrub |
| Geranium grex molle L. | Geraniaceae | Therophyte | Herb | ||
| Geranium molle L. | Geraniaceae | No | PN | Therophyte | Herb |
| Geranium purpureum Vill. | Geraniaceae | No | PN | Therophyte | Herb |
| Geranium rotundifolium L. | Geraniaceae | No | PN | Therophyte | Herb |
| Geranium sp. | Geraniaceae | ||||
| Hirschfeldia incana (L.) Lagr.-Foss. | Brassicaceae | No | PN | Hemicryptophyte | Perennial herb |
| Holcus lanatus L. | Poaceae | No | PN | Hemicryptophyte | Perennial herb |
| Hypochaeris glabra L. | Asteraceae | No | PN | Therophyte | Herb |
| Juncus sp. | Juncaceae | ||||
| Lactuca serriola L. | Asteraceae | No | PN | Hemicryptophyte | Perennial herb |
| Lathyrus clymenum L. | Fabaceae | No | PI | Therophyte | Herb |
| Lathyrus sp. | Fabaceae | ||||
| Lathyrus sphaericus Retz. | Fabaceae | No | PN | Therophyte | Herb |
| Lathyrus tingitanus L. | Fabaceae | No | PI | Therophyte | Herb |
| Leontodon sp. | Asteraceae | ||||
| Luzula elegans Lowe | Juncaceae | No | SN | Therophyte | Herb |
| Lysimachia arvensis (L.) U. Manns & Anderb. | Primulaceae | No | PN | Therophyte | Herb |
| Lysimachia linum-stellatum L. | Primulaceae | No | PN | Therophyte | Herb |
| Matricaria chamomilla L. | Asteraceae | No | SIN | Therophyte | Herb |
| Medicago lupulina L. | Fabaceae | No | PI | Hemicryptophyte | Perennial herb |
| Melilotus sp. | Fabaceae | ||||
| Myosotis ramosissima Rochel | Boraginaceae | No | PN | Therophyte | Herb |
| Neotinea maculata (Desf.) Stearn | Orchidaceae | No | PN | Geophyte | Bulbous perennial herb |
| Origanum vulgare L. | Lamiaceae | No | PN | Chamaephyte | Shrub |
| Ornithopus compressus L. | Fabaceae | No | PN | Therophyte | Herb |
| Ornithopus pinnatus (Mill.) Druce | Fabaceae | No | PI | Therophyte | Herb |
| Oxalis pes-caprae L. | Oxalidaceae | No | SII | Geophyte | Bulbous perennial herb |
| Pallenis spinosa (L.) Cass. | Asteraceae | No | PN | Hemicryptophyte | Perennial herb |
| Papaver pinnatifidum Moris | Papaveraceae | No | PN | Therophyte | Herb |
| Papaver somniferum L. subsp. setigerum (DC.) Arcang. | Papaveraceae | No | PN | Therophyte | Herb |
| Parietaria debilis G. Forst. | Urticaceae | No | PN | Therophyte | Herb |
| Helminthotheca echioides (L.) Holub | Asteraceae | No | PN | Therophyte | Herb |
| Pinus canariensis C. Sm. ex DC. in Buch | Pinaceae | Yes | SN | Macrophanerophyte | Tree |
| Plantago lagopus L. | Plantaginaceae | No | PN | Therophyte | Herb |
| Poaceae sp. | Poaceae | ||||
| Polypogon viridis (Gouan) Breistr. | Poaceae | No | PN | Hemicryptophyte | Perennial herb |
| Prunus sp. | Rosaceae | ||||
| Pteridium aquilinum (L.) Kuhn in Von der Decken | Dennstaedtiaceae | No | PN | Geophyte | Rhizomatous perennial herb |
| Reseda luteola L. | Resedaceae | No | PN | Hemicryptophyte | Perennial herb |
| Rubus ulmifolius Schott | Rosaceae | No | PN | Nanophanerophyte | Liana |
| Rumex acetosella L. | Polygonaceae | No | PI | Hemicryptophyte | Perennial herb |
| Rumex bucephalophorus L. | Polygonaceae | No | SN | Therophyte | Herb |
| Rumex maderensis Lowe | Polygonaceae | No | SN | Chamaephyte | Small shrub |
| Rumex sp. | Polygonaceae | ||||
| Sagina apetala Ard. | Caryophyllaceae | No | PN | Therophyte | Herb |
| Senecio teneriffae Sch. Bip. ex Bolle | Asteraceae | No | SN | Therophyte | Herb |
| Sherardia arvensis L. | Rubiaceae | No | PN | Therophyte | Herb |
| Silene gallica L. | Caryophyllaceae | No | PN | Therophyte | Herb |
| Silene vulgaris (Moench) Garcke | Caryophyllaceae | No | PN | Therophyte | Herb |
| Sonchus acaulis Dum. Cours. | Asteraceae | Yes | SN | Hemicryptophyte | Perennial herb |
| Sonchus asper (L.) A. W. Hill | Asteraceae | No | PN | Therophyte | Herb |
| Sonchus canariensis (Sch. Bip.) Boulos | Asteraceae | Yes | SN | Nanophanerophyte | Shrub |
| Sonchus congestus Willd. | Asteraceae | Yes | SN | Nanophanerophyte | Shrub |
| Sonchus oleraceus L. | Asteraceae | No | PN | Therophyte | Herb |
| Sonchus sp. | Asteraceae | ||||
| Sonchus tenerrimus L. | Asteraceae | No | PN | Therophyte | Herb |
| Spergula arvensis L. | Caryophyllaceae | No | PN | Therophyte | Herb |
| Stachys arvensis (L.) L. | Lamiaceae | No | PN | Therophyte | Herb |
| Stellaria media (L.) Vill. | Caryophyllaceae | No | PI | Therophyte | Herb |
| Tolpis barbata (L.) Gaertn. | Asteraceae | No | PN | Therophyte | Herb |
| Trifolium campestre Schreb. in Sturm | Fabaceae | No | PN | Therophyte | Herb |
| Trifolium dubium Sibth. | Fabaceae | No | PN | Therophyte | Herb |
| Trifolium glomeratum L. | Fabaceae | No | PN | Therophyte | Herb |
| Trifolium ligusticum Balbis ex Loisel. | Fabaceae | No | PN | Therophyte | Herb |
| Trifolium sp. | Fabaceae | ||||
| Trifolium striatum L. | Fabaceae | No | PN | Therophyte | Herb |
| Trifolium subterraneum L. | Fabaceae | No | PN | Therophyte | Herb |
| Tuberaria guttata (L.) Fourr. | Cistaceae | No | PN | Therophyte | Herb |
| Ulex europaeus L. | Fabaceae | No | SII | Nanophanerophyte | Shrub |
| Urospermum picroides (L.) Scop. ex F. W. Schmidt | Asteraceae | No | PN | Therophyte | Herb |
| Urtica morifolia Poir. in Lam. | Urticaceae | No | SN | Nanophanerophyte | Liana |
| Vicia sativa L. subsp. nigra (L.) Ehrh. | Fabaceae | No | PN | Therophyte | Herb |
| Vicia disperma DC. | Fabaceae | No | PN | Therophyte | Herb |
| Vicia lutea L. | Fabaceae | No | PN | Therophyte | Herb |
| Vicia sativa L. | Fabaceae | No | PN | Therophyte | Herb |
| Vicia sativa L. subsp. sativa | Fabaceae | No | PN | Therophyte | Herb |
| Vicia scandens R. P. Murray | Fabaceae | Yes | SN | Therophyte | Herb |
| Vicia sp. | Fabaceae | Therophyte | Herb | ||
| Vicia villosa Roth | Fabaceae | No | PI | Therophyte | Herb |
| Viola kitaibeliana Schult. in Roem. & Schult. | Violaceae | No | SN | Therophyte | Herb |
| Vulpia bromoides (L.) S. F. Gray | Poaceae | No | PN | Therophyte | Herb |
| Vulpia myuros (L.) C. C. Gmel. | Poaceae | No | PN | Therophyte | Herb |
| Wahlenbergia lobelioides (L. f.) Link | Campanulaceae | No | SN | Therophyte | Herb |
References
- Geeson, N.A.; Brandt, C.J.; Thornes, J.B. Mediterranean Desertification: A Mosaic of Processes and Responses; Wiley: Chichester, UK, 2002; 459p. [Google Scholar]
- Rackham, O. Holocene history of Mediterranean island landscapes. In Mediterranean Island Landscapes: Natural and Cultural Approaches; Vogiatzakis, I., Pungetti, G., Mannion, A.M., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 36–60. [Google Scholar]
- Palmer, C.; Colledge, S.; Bevan, S.; Conolly, J. Vegetation recolonisation of abandoned agricultural terraces on Antikythera, Greece. Environ. Archaeol. 2010, 15, 64–80. [Google Scholar] [CrossRef]
- Rounsevell, M.D.A.; Reginster, I.; Araujo, M.B.; Carter, T.R.; Dendoncker, N.; Ewert, F.; House, J.I.; Kankaanpää, S.; Leemans, R.; Metzger, M.J.; et al. A coherent set of future land use change scenarios for Europe. Agric. Ecosyst. Environt. 2006, 114, 57–68. [Google Scholar] [CrossRef]
- Hernández, M. Paisajes Agrarios y Medio Ambiente en Alicante: Evolución e Impactos Ambientales de Los Paisajes Agrarios Alicantinos, 1950–1995; Universidad de Alicante: Alicante, Spain, 1997; 290p. [Google Scholar]
- Ruecker, G.; Schad, P.; Alcubilla, M.M.; Ferrer, C. Natural regeneration of degraded soils and site changes on abandoned agricultural terraces in Mediterranean Spain. Land Degrad. Dev. 1998, 9, 179–188. [Google Scholar] [CrossRef]
- Lasanta, T.; García-Ruiz, J.M.; Pérez Rontomé, M.C.; Sancho, C. Runoff and sediment yield in a semi-arid environment: The effect of land management after farmland abandonment. Catena 2000, 38, 265–278. [Google Scholar] [CrossRef]
- Aide, T.M.; Grau, H.R. Globalization, migration, and Latin American ecosystems. Science 2004, 305, 1915–1916. [Google Scholar] [CrossRef]
- Cramer, V.A.; Hobbs, R.J.; Standish, R.J. What’s new about old fields? Land abandonment and ecosystem assembly. Trends Ecol. Evol. 2008, 23, 104–112. [Google Scholar] [CrossRef]
- Meiners, S.J.; Pickett, S.T.A.; Cadenasso, M.L. Exotic plant invasions over 40 years of old field succession: Community patterns and associations. Ecography 2002, 25, 215–223. [Google Scholar] [CrossRef]
- Corbin, J.D.; D’Antonio, C.M. Competition between native perennial and exotic annual grasses: Implications for an historical invasion. Ecology 2004, 85, 1273–1283. [Google Scholar] [CrossRef]
- Kulmatiski, A. Exotic plants establish persistent communities. Plant Ecol. 2006, 187, 261–275. [Google Scholar] [CrossRef]
- Arévalo, J.R.; Fernández-Lugo, S.; Mellado, M.; de la Concepción, T. Experimental management control of Opuntia dillenii Haw. and Agave americana L. in the Teno Rural Park, Canary Islands. Plant Spec. Biol. 2015, 30, 137–146. [Google Scholar] [CrossRef]
- Mellado, M.; Encina-Domínguez, J.A.; García, J.E.; Estrada-Castillón, E.A.; Arévalo, J.R. Vegetation response to removal of plant functional groups and grass seeding in a microphyllous desert shrubland: A 4-year field experimente. Agriculture 2021, 11, 322. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.G.; Schmitz, O.J. Rapid recovery of damaged ecosystems. PLoS ONE 2009, 4, e5653. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, R.J.; Harris, J.A.; Pärtel, M. Anthropogenic ecosystem disruption and the pathways of recovery. Ecol. Lett. 2006, 9, 773–782. [Google Scholar]
- García-Gallo, A.; Wildpret, W.; del Arco, M.J.; Pérez de Paz, P.L. Sobre la presencia de Ulex europaeus L. en la isla de Tenerife. Bol. Soc. Brot. 1989, 62, 221–225. [Google Scholar]
- García-Gallo, A.; Wildpret, W.; Martín, V. Especies vegetales consideradas invasoras de hábitats, en la Historia Natural de Canarias. Lazaroa 2008, 29, 49–67. [Google Scholar]
- Roberts, J.; Florentine, S. Biology, distribution, and control of the invasive species Ulex europaeus (Gorse): A global synthesis of current and future management challenges and research gaps. Weed Res. 2021, 61, 272–281. [Google Scholar] [CrossRef]
- von Buch, L. Physicalische Beschreibung der Canarischen Inseln; Gedruckt in der Druckerei der Koeninglichen Academie der Wuseebschaften: Berlin, Germany, 1825; 429p. [Google Scholar]
- Sanz, M.; Dana, E.D.; Sobrino, E. Atlas de las Plantas Alóctonas Invasoras en España; Ministerio del Medio Ambiente: Madrid, Spain, 2004; 384p.
- Arévalo, J.R.; Delgado, J.D.; Otto, R.; Naranjo-Cigala, A.; Salas, M.; Fernández-Palacios, J.M. Distribution of alien vs. native plant species in roadside communities along an altitudinal gradient in Tenerife and Gran Canaria (Canary Islands). Perspect. PlantEcol. Evol. Syst. 2005, 7, 185–202. [Google Scholar] [CrossRef]
- Pauchard, A.; Kueffer, C.; Dietz, H.; Daehler, C.C.; Alexander, J.; Edwards, P.; Arévalo, J.R.; Cavieres, L.A.; Guisan, A.; Haider, S.; et al. Ain’t not mountain high enough: Plant invasions reaching high elevations? Front. Ecol. Environ. 2009, 9, 479–486. [Google Scholar] [CrossRef]
- Iseli, E.; Chisholm, C.; Lenoir, J.; Haider, S.; Seipel, T.; Barros, A.; Hargreaves, A.L.; Kardol, P.; Lembrechts, J.J.; McDougall, K.; et al. Rapid upwards spread of non-native plants in mountains across continents. Nat. Ecol. Evol. 2023, 7, 405–413. [Google Scholar] [CrossRef]
- GRAFCAN. Sistema de Información Territorial de Canarias. 2024. Available online: https://visor.grafcan.es/visorweb/ (accessed on 2 May 2024).
- Lorenzo, M.J. Los Cochineros de Icod el Alto; Comisión de Cultura de Ayuntamiento de Los Realejos: Los Realejos, Spain, 1983; 143p. [Google Scholar]
- Patiño, J.; Collart, F.; Vanderpoorten, A.; Martin-Esquivel, J.L.; Naranjo-Cigala, A.; Mirolo, S.; Karger, D.N. Spatial resolution impacts projected plant responses to climate change on topographically complex islands. Divers. Distrib. 2023, 29, 1245–1262. [Google Scholar] [CrossRef]
- Del Arco Aguilar, M.; Rodríguez-Delgado, O. Vegetation of the Canary Islands. Plant and Vegetation; Werger, M.J.A., Ed.; Springer: Utrecht, The Netherlands, 2016; Volume 16, 429p. [Google Scholar]
- Altamirano, A.; Cely, J.P.; Etter, A.; Miranda, A.; Fuentes-Ramírez, A.; Acevedo, P.; Salas, C.; Vargas, R. The invasive species Ulex europaeus (Fabaceae) shows high dynamism in a fragmented landscape of south-central Chile. Environ. Monit. Assess. 2016, 188, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Broadfield, N.; McHenry, M.T. A world of gorse: Persistence of Ulex europaeus in managed landscapes. Plants 2019, 8, 523. [Google Scholar] [CrossRef] [PubMed]
- POWO. Plants of the World Online; Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet. 2023. Available online: http://www.plantsoftheworldonline.org/ (accessed on 14 March 2024).
- Clements, D.R.; Peterson, D.J.; Prasad, R. The biology of Canadian weeds. 112. Ulex europaeus L. Can. J. Plant Sci. 2001, 81, 325–337. [Google Scholar] [CrossRef]
- Ángel-Vallejo, M.C.; Aguirre-Acosta, N.; Rodríguez-Rey, G.T.; García-Marín, E.J.; Álvarez-Mejía, L.M.; Feuillet-Hurtado, C. Distribution models in invasive plants with climatic niche expansion: A case study of Ulex europaeus L. in Colombian Andes. Biol. Invasions 2024, 26, 1919–1930. [Google Scholar] [CrossRef]
- Christina, M.; Gire, C.; Bakker, M.R.; Leckie, A.; Xue, J.; Clinton, P.W.; Negrin-Perez, Z.; Sierra, J.R.A.; Domec, J.-C.; Gonzalez, M. Native and invasive seedling drought-resistance under elevated temperature in common gorse populations. J. Plant Ecol. 2023, 16, rtac097. [Google Scholar] [CrossRef]
- Kariyawasam, C.S.; Ratnayake, S.S. Reproductive biology of gorse, Ulex europaeus (Fabaceae) in the Mount Lofty Ranges of South Australia and Sri Lanka. Int. J. Plant Reprod. Biol. 2019, 11, 145–152. [Google Scholar]
- Udo, N.; Tarayre, M.; Atlan, A. Evolution of germination strategy in the invasive species Ulex Europaeus. J. Plant Ecol. 2017, 10, 375–385. [Google Scholar]
- Bateman, J.B.; Vitousek, P.M. Soil fertility response to Ulex europaeus invasion and restoration efforts. Biol. Invasions 2018, 20, 2777–2791. [Google Scholar] [CrossRef]
- Atlan, A.; Limbada, F. World Distribution of Gorse Ulex europaeus in Introduced Areas. Geonetwork, OSURIS. 2019. Available online: https://www.osuris.fr/geonetwork/srv/fre/catalog.search#/search (accessed on 25 September 2024).
- Smith, B.; Wilson, J.B. A consumer’s guide to evenness indices. Oikos 1996, 76, 70–82. [Google Scholar] [CrossRef]
- Gauch, H.G., Jr. Multivariate Analysis in Community Ecology; Cambridge University Press: Cambridge, UK, 1982; 312p. [Google Scholar]
- Franklin, S.B.; Robertson, P.A.; Fralish, J.S.; Kettler, S.M. Overstory vegetation and successional trends of land between the Lakes, USA. J. Veg. Sci. 1993, 4, 509–520. [Google Scholar] [CrossRef]
- Arévalo, J.R.; Fernández-Palacios, J.M.; Palmer, M. Tree regeneration and predicted future dynamics in a laurel forest (Tenerife, Canary Islands). J. Veg. Sci. 1999, 10, 861–868. [Google Scholar] [CrossRef]
- Hill, M.O.; Gauch, H.J., Jr. Detrended Correspondence Analysis: An improved ordination technique. Vegetatio 1980, 42, 47–58. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows Users Guide: Software for Canonical Community Ordination; Version 5.1; Microcomputer Power: Ithaca, NY, USA, 2018. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 1998; 853p. [Google Scholar]
- Pallant, J. SPSS Survival Manual: A Step-by-Step Guide to Data Analysis Using IBM SPSS; Open University Press: London, UK, 2021; 378p. [Google Scholar]
- Richardson, D.M.; Pyšek, P. Plant invasions: Merging the concepts of species invasiveness and community invasibility. Prog. Phys. Geogr. 2006, 30, 409–431. [Google Scholar] [CrossRef]
- Dietz, H.; Edwards, P.J. Recognition of changing processes during plant invasions may help reconcile conflicting evidence of the causes. Ecology 2006, 87, 1359–1367. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: Cambridge, MA, USA, 2014; 1586p. [Google Scholar]
- Fenner, M. The effects of the parent environment on seed germinability. Seed Sci. Res. 1991, 1, 75–84. [Google Scholar] [CrossRef]
- Baird, D.D.; Upchurch, R.P.; Homesley, W.B.; Franz, J.E. Introduction of a new broad spectrum postemergence herbicide class with utility for herbaceous perennial weed control. Proc. North Cent. Weed Control. Conf. 1971, 26, 64–68. [Google Scholar]
- Diatloff, G. New uses of glyphosate. In Proceedings of the 12th Australian Weeds Conference Proceedings, Hobart, Austria, 12–16 September 1999; pp. 299–301. [Google Scholar]
- Mokotjomela, T.M.; Nelufule, T.; Scott, Z.; Vukeya, L.R.; Xivuri, T.; Matsokane, K.; Mweli, N.; Magqabi, F.L.; Thulisile, J. The invasion threat of the emerging alien cactus Cylindropuntia pallida (Rosa), F.M. Knuth in South Africa and the potential for control using herbicides. Environ. Monit. Assess. 2024, 196, 673. [Google Scholar] [CrossRef]
- Arévalo, J.R.; Encina-Domínguez, J.A.; Juanes-Márquez, S.; Álvarez-Vázquez, P.; Nuñez-Colima, J.A.; Mellado, M. Restoration of rangelands invaded by Amelichloa clandestina (Hack.) Arriaga & Barkworth after 12 years of agriculture abandonment (Coahuila, Mexico). Agriculture 2021, 11, 886. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: New York, NY, USA, 1995; 436p. [Google Scholar]
- Stace, C.A. New Flora of the British Isles; Cambridge University Press: New York, NY, USA, 2010; 1165p. [Google Scholar]




| Block 1 | Block 2 | |
|---|---|---|
| Altitude (m) | 1151.7 | 1079.0 |
| Aspect (°) | 297.5 | 309.7 |
| Slope (°sex) | 10.0 | 8.9 |
| Cover in % | ||
| Grasses | 22.8 | 70.3 |
| Forbs | 49.6 | 47.1 |
| Woody | 37.6 | 70.3 |
| Rock | 9.9 | 0.6 |
| Soil | 1.3 | 0.1 |
| Litter | 88.9 | 93.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Montelongo, C.; Padrón-Mederos, M.A.; Negrín-Pérez, Z.; González, M.; Arévalo, J.R. Management Strategies for Ulex europaeus L. Control in a Native Plant Community in Tenerife, Canary Islands. Agriculture 2024, 14, 1683. https://doi.org/10.3390/agriculture14101683
González-Montelongo C, Padrón-Mederos MA, Negrín-Pérez Z, González M, Arévalo JR. Management Strategies for Ulex europaeus L. Control in a Native Plant Community in Tenerife, Canary Islands. Agriculture. 2024; 14(10):1683. https://doi.org/10.3390/agriculture14101683
Chicago/Turabian StyleGonzález-Montelongo, Cristina, Miguel A. Padrón-Mederos, Zaira Negrín-Pérez, Maya González, and José Ramón Arévalo. 2024. "Management Strategies for Ulex europaeus L. Control in a Native Plant Community in Tenerife, Canary Islands" Agriculture 14, no. 10: 1683. https://doi.org/10.3390/agriculture14101683
APA StyleGonzález-Montelongo, C., Padrón-Mederos, M. A., Negrín-Pérez, Z., González, M., & Arévalo, J. R. (2024). Management Strategies for Ulex europaeus L. Control in a Native Plant Community in Tenerife, Canary Islands. Agriculture, 14(10), 1683. https://doi.org/10.3390/agriculture14101683