
Studying the Effect of Straw Returning on the Interspecific Symbiosis of Soil Microbes Based on Carbon Source Utilization


Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials
2.2. Test Design
2.3. Sample Collection and Index Determination
2.3.1. Determination of Soil Microbial Community
2.3.2. Calculation of Straw Decomposition Rate
2.4. Data Analysis
2.4.1. Path Analysis Model (PA)
2.4.2. Grey Relational Analysis Model (GRA)
3. Results and Analysis
3.1. Decomposition Rate of Straw with Different Returning Ways
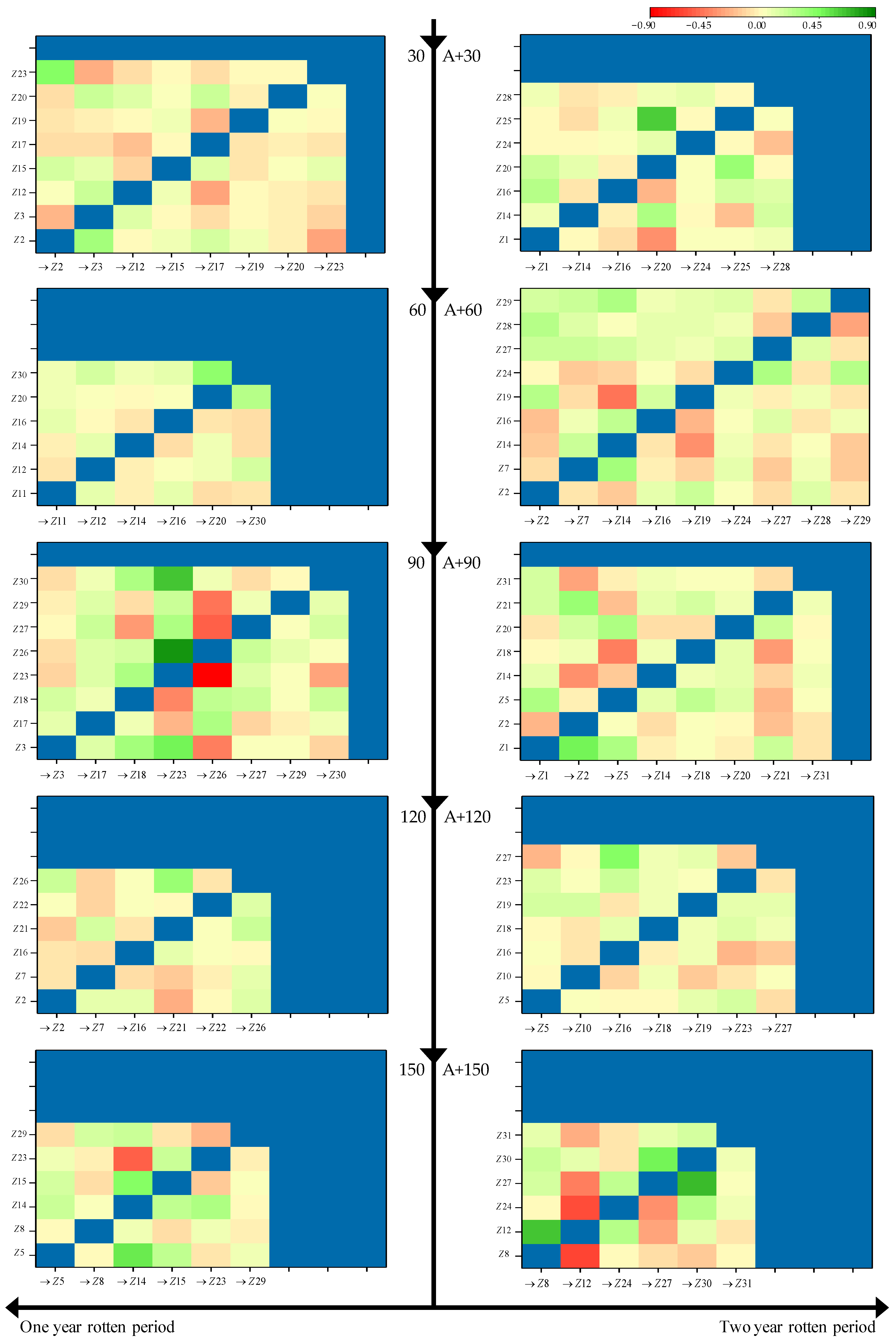
3.2. Relational Analysis between Microbes and Straw Decomposition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gu, Z.J.; Xie, Y.; Gao, Y.; Ren, X.Y.; Cheng, C.C.; Wang, S.C. Quantitative assessment of soil productivity and predicted impacts of water erosion in the black soil region of northeastern China. Sci. Total Environ. 2018, 637, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, G.; Chen, S.L.; Rasmussen, C.; Liu, B.Y. Assessing soil thickness in a black soil watershed in Northeast China using random forest and field observations. Int. Soil Water. Conse. 2021, 9, 49–57. [Google Scholar] [CrossRef]
- Shi, T.T.; Liu, Y.Q.; Zhang, L.B.; Hao, L.; Gao, Z.Q. Burning in agricultural landscapes: An emerging natural and human issue in China. Landsc. Ecol. 2014, 29, 1785–1798. [Google Scholar] [CrossRef]
- Yin, H.J.; Zhao, W.Q.; Li, T.; Cheng, X.Y.; Liu, Q. Balancing straw returning and chemical fertilizers in China: Role of straw nutrient resources. Renew. Sustain. Energy Rev. 2018, 81, 2695–2702. [Google Scholar] [CrossRef]
- Wang, M.Q.; Liu, Y.S.; Huang, Y.L.; Zhao, Y.Y.; Li, Z.X.; Han, Y.H. Research progress on effects of straw incorporation on soil micro-ecological environment. Microbiol. China 2022, 49, 807–816. [Google Scholar] [CrossRef]
- Liu, C.; Lu, M.; Cui, J.; Li, B.; Fang, C.M. Effects of straw carbon input on carbon dynamics in agricultural soils: A meta-analysis. Glob. Chang. Biol. 2014, 20, 1366–1381. [Google Scholar] [CrossRef]
- Nunes, C.; Joao, L.; Nogueirol, R.C.; Santos Menandro, L.M.; Bordonal, R.D.O.; Borges, C.D.; Cantarella, H.; Junqueira Franco, H.C. Agronomic and environmental implications of sugarcane straw removal: A major review. GCB Bioenergy 2017, 9, 1181–1195. [Google Scholar] [CrossRef]
- Sarkar, S.; Skalicky, M.; Hossain, A.; Brestic, M.; Saha, S.; Garai, S.; Ray, K.; Brahmachari, K. Management of crop residues for improving input use efficiency and agricultural sustainability. Sustainability 2020, 12, 9808. [Google Scholar] [CrossRef]
- Mabagala, F.S.; Geng, Y.H.; Cao, G.J.; Wang, L.C.; Wang, M.; Zhang, M.L. Silicon accumulation, partitioning and remobilization in spring maize (Zea mays L.) under silicon supply with straw return in Northeast China. J. Plant Nutr. 2020, 44, 1498–1514. [Google Scholar] [CrossRef]
- Cai, L.J.; Zhang, J.T.; Liu, J.Q.; Gai, Z.J.; Guo, Z.H.; Zhao, G.F. Effects of long-term no-tillage straw returning on soil organic carbon and soybean yield in cold region. Crops 2021, 6, 189–192. [Google Scholar] [CrossRef]
- Bordonal, R.D.O.; Nunes Carvalho, J.L.; Lal, R.; de Figueiredo, E.B.; de Oliveira, B.G.; Scala, N.L., Jr. Sustainability of sugarcane production in Brazil. A review. Agron. Sustain. Dev. 2018, 38, 13. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Yan, S.S.; Jia, T.Y.; Dong, S.K.; Ma, C.M.; Gong, Z.P. Decomposition characteristics of rice straw returned to the soil in Northeast China. Nutr. Cycl. Agroecosys. 2019, 114, 211–224. [Google Scholar] [CrossRef]
- Chen, A.; Zhang, W.; Sheng, R.; Liu, Y.; Hou, H.; Liu, F.; Ma, G.; Wei, W.; Qin, H. Long-term partial replacement of mineral fertilizer with in situ crop residues ensures continued rice yields and soil fertility: A case study of a 27-year field experiment in subtropical China. Sci. Total Environ. 2021, 787, 147523. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zeng, Y.; Wu, J.; Shi, Q.; Pan, X. Effect of crop residue retention on rice yield in China: A meta-analysis. Field Crop. Res. 2013, 154, 188–194. [Google Scholar] [CrossRef]
- Zhao, J.L.; Wang, X.G.; Zhuang, J.; Cong, Y.J.; Lu, Y.; Guo, M.Z. Fine-crush straw returning enhances dry matter accumulation rate of maize seedlings in Northeast China. Agronomy 2021, 11, 1144. [Google Scholar] [CrossRef]
- Xu, G.M.; Wang, X.C.; He, R.Y.; Ding, Q.S. Performance evaluation of rotary tillage straw returning based on composite indicators and measurement techniques. Trans. Chin. Soc. Agric. Mach. 2022, 53, 58–67. [Google Scholar] [CrossRef]
- Ma, L.J.; Kong, F.X.; Wang, Z.; Luo, Y.; Lv, X.B.; Zhou, Z.G.; Meng, Y.L. Growth and yield of cotton as affected by different straw returning modes with an equivalent carbon input. Field Crop. Res. 2019, 243, 107616. [Google Scholar] [CrossRef]
- Wang, D.; Sun, C.X.; Cui, M.; Shen, X.B.; Zhang, Y.L.; Xiao, J.H.; Liu, P.Y.; Zhang, Y.; Xie, H.T. An integrated analysis of transcriptome and metabolome provides insights into the responses of maize (Zea mays L.) roots to different straw and fertilizer conditions. Environ. Exp. Bot. 2022, 194, 104732. [Google Scholar] [CrossRef]
- Ji, B.Y.; Hu, H.; Zhao, Y.L.; Mu, X.Y.; Liu, K.; Li, C.H. Effects of deep tillage and straw returning on soil microorganism and enzyme activities. Sci. World J. 2014, 2014, 451493. [Google Scholar] [CrossRef]
- Li, Y.Z.; Song, D.P.; Dang, P.F.; Wei, L.N.; Qin, X.L.; Siddique, K.H.M. Combined ditch buried straw return technology in a ridge-furrow plastic film mulch system: Implications for crop yield and soil organic matter dynamics. Soil Tillage Res. 2020, 199, 104596. [Google Scholar] [CrossRef]
- Zhang, H.; Liang, S.; Wang, Y.H.; Liu, S.W.; Sun, H.D. Greenhouse gas emissions of rice straw return varies with return depth and soil type in paddy systems of Northeast China. Arch. Agron. Soil Sci. 2021, 67, 1591–1602. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, H.Y.; Yang, Z.H.; Jiang, Z.Z.; Tao, Y.Z.; Liu, C.Z.; Faheem, J.M.; Li, M. Effects of straw returning modes on maize seedling growth under different soil conditions. Chin. J. Ecol. 2022, 41, 479–486. [Google Scholar] [CrossRef]
- Ning, Y.C.; Zhou, H.R.; Zhou, D.X. Study on the microbial community in earthworm and soil under cadmium stress based on contour line analysis. Environ. Sci. Pollut. Res. 2019, 26, 20989–21000. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.X.; Su, Y.; Ning, Y.C.; Rong, G.H.; Wang, G.D.; Liu, D.; Liu, L.Y. Estimation of the effects of maize straw return on soil carbon and nutrients using response surface methodology. Pedosphere 2018, 28, 411–421. [Google Scholar] [CrossRef]
- Rong, G.H.; Ning, Y.C.; Cao, X.; Su, Y.; Li, J.; Li, L.; Liu, L.Y.; Zhou, D.X. Evaluation of optimal straw incorporation characteristics based on quadratic orthogonal rotation combination design. J. Agric. Sci. 2018, 156, 367–377. [Google Scholar] [CrossRef]
- Coffman, D.L.; MacCallum, R.C. Using parcels to convert path analysis models into latent variable models. Multivar. Behav. Res. 2005, 40, 235–259. [Google Scholar] [CrossRef]
- Hu, Y.F.; Zhang, M.K. Composition characteristics of aggregates in red sandstone soil and their responses to amendments. Acta Agric. Jiangxi 2019, 31, 88–93. [Google Scholar] [CrossRef]
- Yang, Z.J.; Guo, X.T.; Sun, J.; Zhang, Y.L. Contextual and organizational factors in sustainable supply chain decision making: Grey relational analysis and interpretative structural modeling. Environ. Dev. Sustain. 2021, 23, 12056–12076. [Google Scholar] [CrossRef]
- Liu, Y.L.; Huang, X.L.; Duan, J.; Zhang, H.M. The assessment of traffic accident risk based on grey relational analysis and fuzzy comprehensive evaluation method. Nat. Hazards 2017, 88, 1409–1422. [Google Scholar] [CrossRef]
- Gao, F.; Jia, Z.K.; Lu, W.T.; Han, Q.F.; Yang, B.P.; Hou, X.Q. Effects of different straw returning treatments on soil water, maize growth and photosyntheticcharacteristics in the semi-arid area of Southern Ninaxia. Acta Ecol. Sin. 2011, 31, 777–783. [Google Scholar]
- Zhang, P.; Wei, T.; Li, Y.L.; Wang, K.; Jia, Z.K.; Han, Q.F.; Ren, X.L. Effects of straw incorporation on the stratification of the soil organic C, total N and C:N ratio in a semiarid region of China. Soil Tillage Res. 2015, 153, 28–35. [Google Scholar] [CrossRef]
- Ma, F.X.; Wang, Y.Y.; Yan, P.; Wei, F.; Sun, X.Z.; Liu, J.G. Effects of cotton straw incorporation on organic nitrogen fractions in long-term continuous cropping cotton field. Ecol. Environ. Sci. 2018, 27, 1459–1465. [Google Scholar] [CrossRef]
- Long, Z.H.; Wang, J.; Hou, Z.A. Effects of cotton straw biochar returning and N application rate on soil organic nitrogen fractions in cotton field. J. Shihezi Univ. 2019, 37, 154–161. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Zhang, A.Z.; Jiang, N. Effects of mixed fungal fermentation on degradation rate of cellulose and lignin of corn straw. Chin. J. Anim. Nutr. 2019, 31, 1385–1395. [Google Scholar] [CrossRef]
- Zeng, Z.T.; Guo, X.Y.; Xu, P.A.; Xiao, R.; Huang, D.L.; Gong, X.M.; Cheng, M.; Yi, H.; Li, T.; Zeng, G.M. Responses of microbial carbon metabolism and function diversity induced by complex fungal enzymes in lignocellulosic waste composting. Sci. Total Environ. 2018, 643, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.L.; Han, G.M.; Lan, Y.; Liu, S.N.; Gao, J.P.; Yang, X.; Meng, J.; Chen, W.F. Corn cob biochar increases soil culturable bacterial abundance without enhancing their capacities in utilizing carbon sources in Biolog Eco-plates. J. Integr. Agric. 2017, 16, 713–724. [Google Scholar] [CrossRef] [Green Version]
- Ghasemi, A.; Asad, S.; Kabiri, M.; Dabirmanesh, B. Cloning and characterization of Halomonas elongata L-asparaginase, a promising chemotherapeutic agent. Appl. Microbiol. Biotechnol. 2017, 101, 7227–7238. [Google Scholar] [CrossRef]
- Pezzotti, F.; Therisod, H.; Therisod, M. Enzymatic synthesis of D-glucosaminic acid from D-glucosamine. Carbohydr. Res. 2005, 340, 139–141. [Google Scholar] [CrossRef]
- Bhat, U.R.; Forberg, L.S.; Carlson, R.W. Structure of lipid a component of Rhizobium leguminosarum bv. phaseoli lipopolysaccharide. Unique nonphosphorylated lipid A containing 2-amino-2-deoxygluconate, galacturonate, and glucosamine. J. Biol. Chem. 1994, 269, 14402–14410. [Google Scholar] [CrossRef]
- Wu, N.; Li, X.F.; Huang, G.S.; Pan, P.; Wang, C.; Liu, X.Y.; Zeng, M. Adsorption and biodegradation functions of novel microbial embedding polyvinyl alcohol gel beads modified with cyclodextrin: A case study of benzene. Environ. Technol. 2018, 40, 1948–1958. [Google Scholar] [CrossRef]
- Tribak, M.; Ocampo, J.A.; Garcia-Romera, I. Production of xyloglucanolytic enzymes by Trichoderma viride, Paecilomyces farinosus, Wardomyces inflatus, and Pleurotus ostreatus. Mycologia 2002, 94, 404–410. [Google Scholar] [CrossRef]
- Du, L.L.; Li, G.X.; Yuan, J.; Yuan, J.B. Effect of additives on NH3 and H2S emissions during kitchen waste composting. Trans. Chin. Soc. Agric. Eng. 2015, 31, 195–200. [Google Scholar] [CrossRef]
- Yan, J.L.; Zhang, M.K.; Wang, D.Z. Different amendments: Effect on soil physical properties of newly reclaimed land in low hilly region. Chin. Agric. Sci. Bull. 2021, 37, 67–73. [Google Scholar] [CrossRef]
- Zhang, G.W.; Yang, C.Q.; Liu, R.X.; Ni, W.C. Effects of p-hydroxybenzoic acid and phloroglucinol on mitochondria function and root growth in cotton (Gossypium hirsutum L.) seedling roots. Chin. J. Appl. Ecol. 2018, 29, 231–237. [Google Scholar] [CrossRef]
- Sun, S.S.; Gong, B.; Wen, D.; Wang, X.F.; Wei, M.; Yang, F.J.; Li, Y.; Shi, Q.H. Effect of exogenous melatonin on physiological and biochemical characteristics of cucumber radicles under p-hydroxybenzoic acid. Chin. J. Appl. Ecol. 2016, 27, 897–903. [Google Scholar] [CrossRef]
- Zhao, X.S.; Qi, Y.Z.; Yan, C.M.; Zhen, W.C. Allelopathy of six organic acids on wheat sheath blight in the soil of winter wheat-summer maize double cropping straw returning system. Sci. Agric. Sin. 2020, 53, 3095–3107. [Google Scholar] [CrossRef]
- Tang, L.L.; Nie, S.R.; Li, W.H.; Fan, C.; Wang, S.Q.; Wu, F.Z.; Pan, K. Wheat straw increases the defense response and resistance of watermelon monoculture to Fusarium wilt. BMC Plant Biol. 2019, 19, 551. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.Z.; Zhen, W.C.; Li, H.Y. Allelopathy of decomposed maize straw products on three soil-born diseases of wheat and the analysis by GC-MS. J. Integr. Agric. 2015, 14, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, Y.B.; Teng, L.P.; Yue, J. Allelopathy of different decomposed liquids of cotton stalk on fusarium oxysporum and verticillium dahliae. J. Agro-Environ. Sci. 2012, 31, 1696–1701. [Google Scholar]
- Han, Z.; Di, C.Q.; Rahman, M.; Gao, D.M.; Wu, F.Z.; Pan, K. Repeated application of rice straw stabilizes soil bacterial community composition and inhibits clubroot disease. Agriculture 2021, 11, 108. [Google Scholar] [CrossRef]
- Chen, L.J.; Zhou, J.H.; Chen, G.; Liu, L.; Tan, J. Research progress of influence and mechanism of field straw residue incorporation on soil-borne diseases in crops. Crop Res. 2018, 32, 535–540. [Google Scholar] [CrossRef]
- Yang, L.P.; Yao, X.Y.; Li, Q.; Jiang, Q.P. An analysis of function diversity of soil microbes in relation to the outbreak of tobacco root rot. Plant Dr. 2020, 33, 36–41. [Google Scholar] [CrossRef]
- Zhang, S.T.; Liu, X.J.; Zhou, L.H.; Deng, L.Y.; Zhao, W.A.; Liu, Y.; Ding, W. Alleviating soil acidification could increase disease suppression of bacterial wilt by recruiting potentially beneficial rhizobacteria. Microbiol. Spectr. 2022, 10, e0233321. [Google Scholar] [CrossRef]
- Raj, A.; Reddy, M.M.K.; Chandra, R.; Purohit, H.J.; Kapley, A. Biodegradation of kraft-lignin by Bacillus sp isolated from sludge of pulp and paper mill. Biodegradatioin 2007, 18, 783–792. [Google Scholar] [CrossRef]
- Liu, X.C.; Wang, J.; Guo, S.H.; Chen, C.M. Effects of co-substrates and inorganic salts on degradation of crude oil by oil degrading bacteria. Environ. Prot. Chem. Ind. 2008, 3, 218–221. [Google Scholar] [CrossRef]
- Cheng, J.; Wu, D.; Chen, S.; Wu, J.; Chen, J. Effect of complex and synthetic medium on extracellular production of α-cyclodextrin glycosyltransferase in E. coli. China Biotechnol. 2010, 30, 36–42. [Google Scholar] [CrossRef]
- Hayward, M.R.; AbuOun, M.; Woodward, M.J.; Jansen, V.A.A. Temperature and oxygen dependent metabolite utilization by salmonella enterica serovars derby and mbandaka. PLoS ONE 2015, 10, e0120450. [Google Scholar] [CrossRef] [Green Version]
- Makonde, H.M.; Mwirichia, R.; Osiemo, Z.; Boga, H.I.; Klenk, H.P. 454 Pyrosequencing-based assessment of bacterial diversity and community structure in termite guts, mounds and surrounding soils. SpringerPlus 2015, 4, 471. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.X.; Wei, J.H.; Li, J.J.; Ni, J.F. Whole-genome analysis of the dominant bacterium Dysgonomonas macrotermitis in the hindgut of Macrotermes barneyi. Acta Microbiol. Sin. 2018, 58, 995–1003. [Google Scholar] [CrossRef]
- Gong, Z.P.; Deng, N.Z.; Song, Q.L.; Li, Z.T. Decomposing characteristics of maize straw returning in Songnen Plain in long-time located experiment. Trans. Chin. Soc. Agric. Eng. 2018, 34, 139–145. [Google Scholar] [CrossRef]
- Chen, L.J.; Chen, R.; Zhou, J.H.; Yan, C.B.; Liu, L.; Li, Q.; Zhang, Y. Decomposition characteristics of three crop straws and the effects of their decomposed liquids on phytophthora nicotianae. Chin. Tob. Sci. 2021, 42, 33–39. [Google Scholar] [CrossRef]
- Schmid, C.A.O.; Schroder, P.; Armbruster, M.; Schloter, M. Organic amendments in a long-term field trial-consequences for the bulk soil bacterial community as revealed by network analysis. Microb. Ecol. 2018, 76, 226–239. [Google Scholar] [CrossRef]
- Tang, L.L.; Xia, Y.; Kou, J.M.; Wu, F.Z.; Li, W.H.; Pan, K. Control of fusarium wilt by wheat straw is associated with microbial network changes in watermelon rhizosphere. Sci. Rep. 2020, 10, 12736. [Google Scholar] [CrossRef]
- Ma, L.; Li, Y.; Wei, J.L.; Li, Z.S.; Zhou, X.L.; Zheng, F.L.; Wu, X.B.; Wang, L.; Liu, Z.H.; Tan, D.S. Effects of long-term straw returning on fungal community, enzyme activity and wheat yield in a fluvo-aquic soil. Environ. Sci. 2022. [CrossRef]
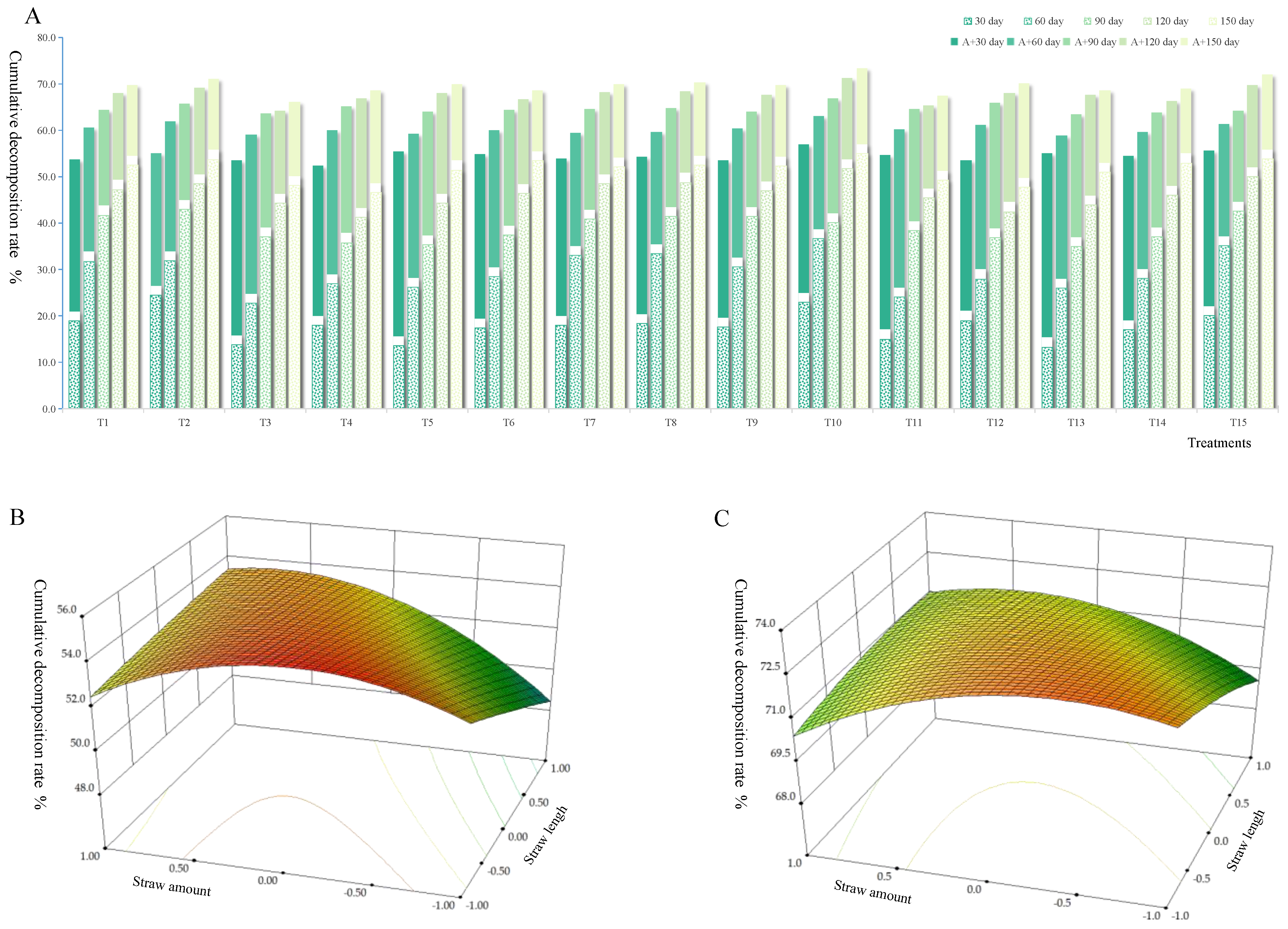


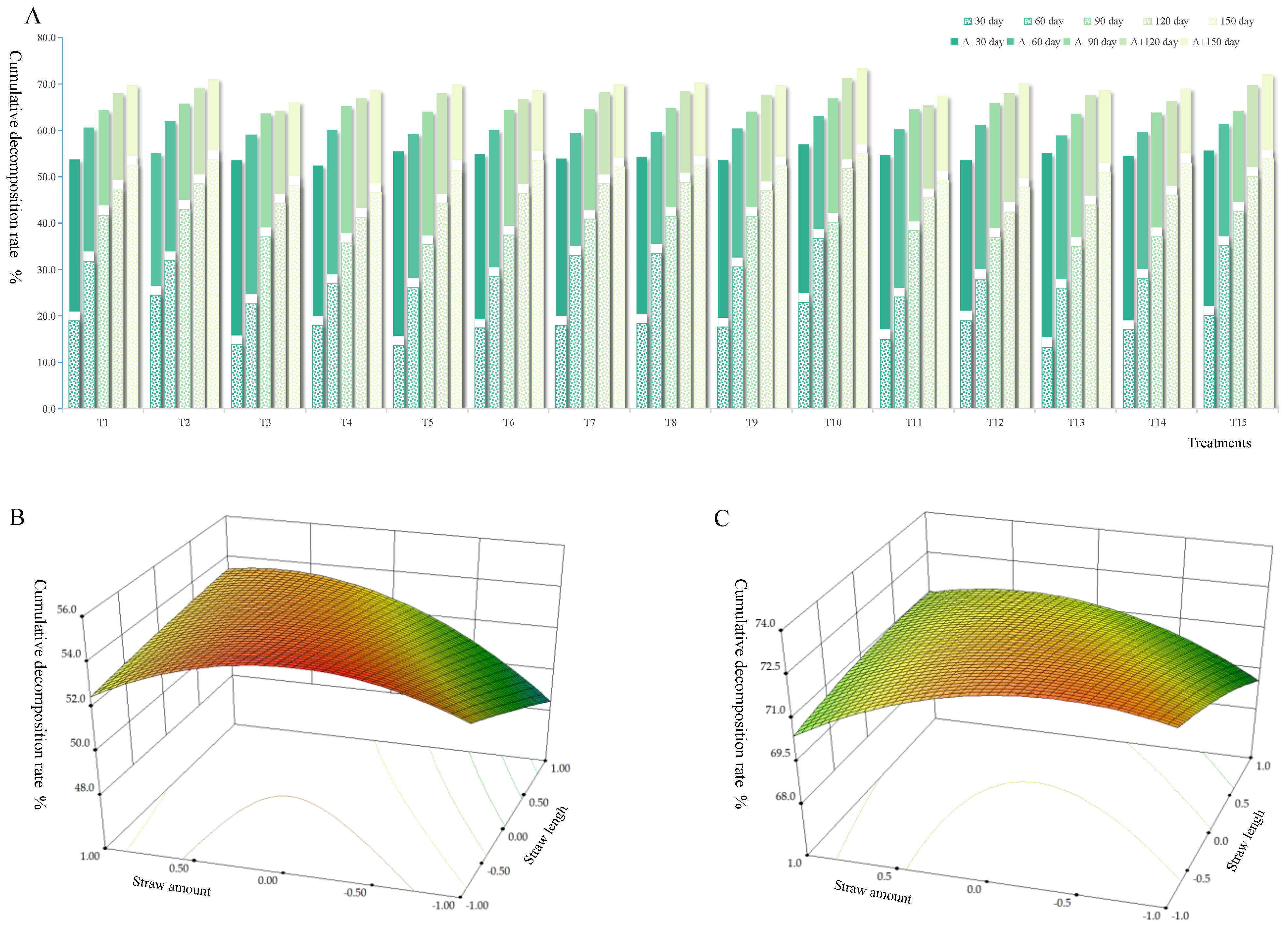{kind=link}
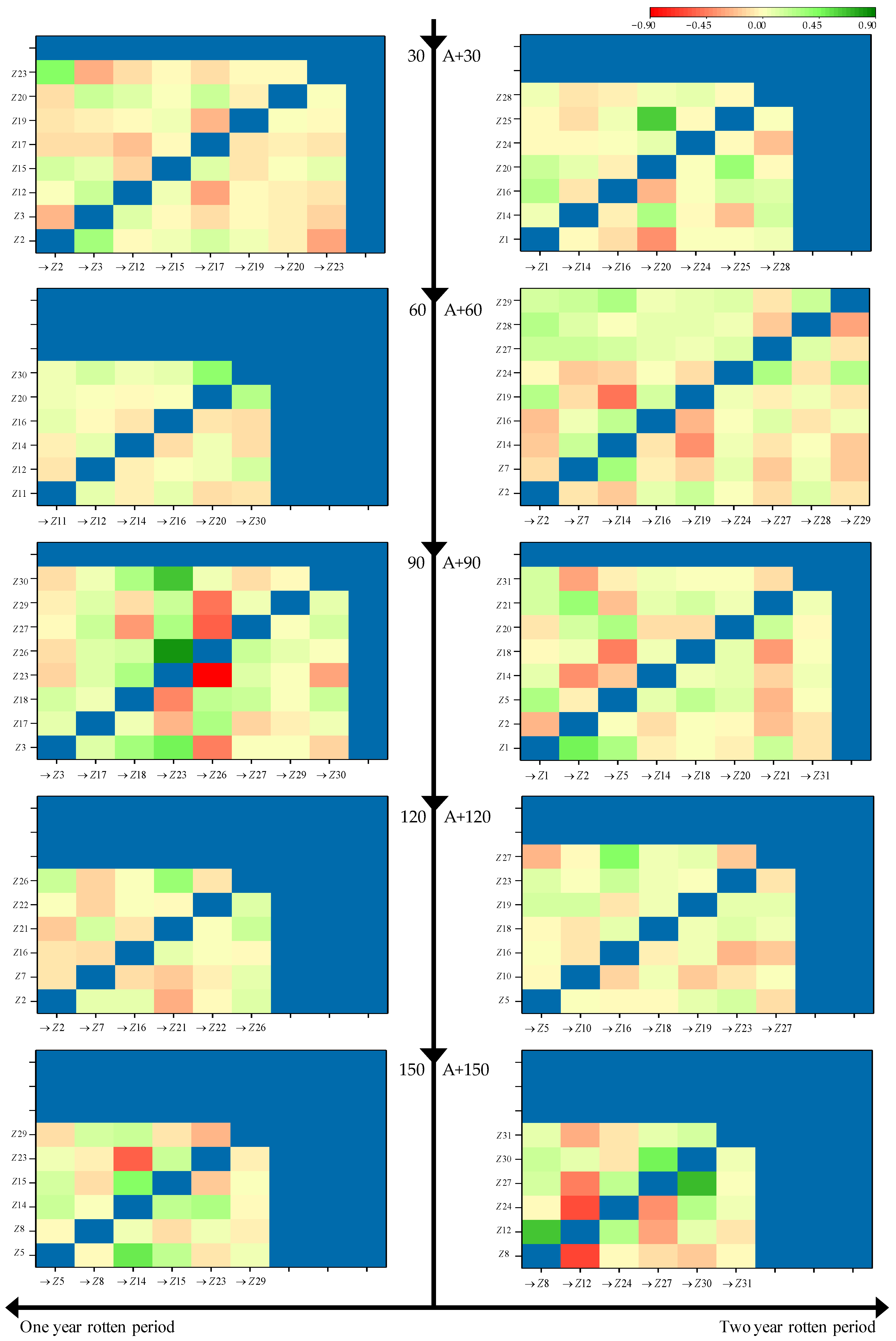{kind=link}
{kind=link}
| Treatments | Code Value | Actual Value | ||||
|---|---|---|---|---|---|---|
| Test Factor A | Test Factor B | Test Factor C | Straw Length cm | Straw Amount kg/hm2 | Straw Buried Depth cm | |
| 1 | 1 | 1 | 1 | 20 | 6800 | 20 |
| 2 | 1 | 1 | −1 | 20 | 6800 | 10 |
| 3 | 1 | −1 | 1 | 20 | 2800 | 20 |
| 4 | 1 | −1 | −1 | 20 | 2800 | 10 |
| 5 | −1 | 1 | 1 | 10 | 6800 | 20 |
| 6 | −1 | 1 | −1 | 10 | 6800 | 10 |
| 7 | −1 | −1 | 1 | 10 | 2800 | 20 |
| 8 | −1 | −1 | −1 | 10 | 2800 | 10 |
| 9 | 1.682 | 0 | 0 | 25 | 4800 | 15 |
| 10 | −1.682 | 0 | 0 | 5 | 4800 | 15 |
| 11 | 0 | 1.682 | 0 | 15 | 8000 | 15 |
| 12 | 0 | −1.682 | 0 | 15 | 1600 | 15 |
| 13 | 0 | 0 | 1.682 | 15 | 4800 | 25 |
| 14 | 0 | 0 | −1.682 | 15 | 4800 | 5 |
| 15 | 0 | 0 | 0 | 15 | 4800 | 15 |
| Test Design | Straw Returning Time (Day) | ||||
|---|---|---|---|---|---|
| One-year decomposition period | 30 | 60 | 90 | 120 | 150 |
| Two-year decomposition period | A + 30 | A + 60 | A + 90 | A + 120 | A + 150 |
| Time (Day) | One-Year Decomposition Period | Two-Year Decomposition Period | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Factors | Path Coefficient | GCD | Factors | Path Coefficient | GCD | |||||
| Directly | Indirectly | Total | Directly | Indirectly | Total | |||||
| 30 A + 30 | Z2 | −0.646 | 0.182 | −0.464 | 0.646 | Z1 | 0.500 | −0.439 | 0.062 | 0.659 |
| Z3 | 0.846 | −0.531 | 0.315 | 0.758 | Z14 | −0.299 | 0.166 | −0.133 | 0.685 | |
| Z12 | 0.560 | −0.255 | 0.305 | 0.702 | Z16 | −0.241 | 0.195 | −0.047 | 0.678 | |
| Z15 | −0.192 | 0.206 | 0.014 | 0.662 | Z20 | −1.030 | 0.552 | −0.478 | 0.678 | |
| Z17 | 0.837 | −0.736 | 0.101 | 0.681 | Z24 | −0.217 | −0.215 | −0.432 | 0.677 | |
| Z19 | 0.354 | −0.359 | −0.005 | 0.666 | Z25 | −0.578 | 0.535 | −0.043 | 0.667 | |
| Z20 | −0.282 | 0.346 | 0.064 | 0.709 | Z28 | 0.506 | 0.052 | 0.558 | 0.760 | |
| Z23 | 0.507 | −0.135 | 0.371 | 0.710 | ||||||
| 60 A + 60 | Z11 | 0.437 | −0.093 | 0.344 | 0.677 | Z2 | 0.787 | −0.073 | 0.714 | 0.805 |
| Z12 | −0.511 | 0.093 | −0.418 | 0.645 | Z7 | 0.506 | −0.205 | 0.301 | 0.757 | |
| Z14 | 0.282 | −0.138 | 0.144 | 0.674 | Z14 | 0.765 | −0.707 | 0.058 | 0.659 | |
| Z16 | 0.359 | −0.223 | 0.136 | 0.695 | Z16 | −0.339 | −0.064 | −0.403 | 0.664 | |
| Z20 | −0.962 | 0.331 | −0.631 | 0.586 | Z19 | 0.594 | −0.211 | 0.383 | 0.712 | |
| Z30 | −0.581 | 0.741 | 0.161 | 0.679 | Z24 | −0.222 | −0.034 | −0.256 | 0.674 | |
| Z27 | −0.499 | 0.776 | 0.277 | 0.708 | ||||||
| Z28 | 0.316 | 0.124 | 0.440 | 0.715 | ||||||
| Z29 | −0.538 | 0.919 | 0.380 | 0.740 | ||||||
| 90 A + 90 | Z3 | −0.374 | 0.374 | 0.000 | 0.690 | Z1 | −0.701 | 0.798 | 0.097 | 0.722 |
| Z17 | −0.555 | 0.043 | −0.512 | 0.651 | Z2 | 1.296 | −0.648 | 0.648 | 0.818 | |
| Z18 | −0.807 | 0.406 | −0.401 | 0.705 | Z5 | −0.679 | 0.440 | −0.239 | 0.672 | |
| Z23 | 1.160 | −0.780 | 0.379 | 0.751 | Z14 | 0.367 | −0.457 | −0.091 | 0.724 | |
| Z26 | −1.239 | 1.253 | 0.014 | 0.722 | Z18 | 0.348 | −0.640 | −0.292 | 0.699 | |
| Z27 | 0.427 | −0.237 | 0.190 | 0.727 | Z20 | −0.285 | 0.263 | −0.022 | 0.735 | |
| Z29 | 0.193 | −0.203 | −0.011 | 0.692 | Z21 | −0.818 | 0.602 | −0.216 | 0.699 | |
| Z30 | −0.543 | 0.774 | 0.231 | 0.739 | Z31 | 0.284 | −0.270 | 0.014 | 0.674 | |
| 120 A + 120 | Z2 | −0.437 | 0.012 | −0.425 | 0.727 | Z5 | −0.663 | 0.102 | −0.561 | 0.682 |
| Z7 | 0.549 | −0.337 | 0.212 | 0.758 | Z10 | 0.311 | −0.373 | −0.062 | 0.668 | |
| Z16 | 0.536 | −0.097 | 0.439 | 0.782 | Z16 | 0.677 | −0.475 | 0.202 | 0.696 | |
| Z21 | −0.677 | 0.087 | −0.591 | 0.690 | Z18 | −0.280 | 0.262 | −0.018 | 0.702 | |
| Z22 | 0.198 | −0.012 | 0.186 | 0.773 | Z19 | −0.362 | 0.429 | 0.067 | 0.681 | |
| Z26 | −0.323 | 0.338 | 0.015 | 0.732 | Z23 | −0.860 | 0.346 | −0.514 | 0.624 | |
| Z27 | −0.294 | 0.101 | −0.193 | 0.664 | ||||||
| 150 A + 150 | Z5 | −0.389 | 0.635 | 0.246 | 0.726 | Z8 | 1.038 | −0.979 | 0.059 | 0.716 |
| Z8 | −0.522 | −0.077 | −0.599 | 0.701 | Z12 | −1.035 | 0.603 | −0.432 | 0.690 | |
| Z14 | −1.135 | 0.666 | −0.469 | 0.694 | Z24 | 0.442 | −0.729 | −0.287 | 0.727 | |
| Z15 | −0.538 | 0.299 | −0.239 | 0.726 | Z27 | −0.793 | 0.654 | −0.139 | 0.735 | |
| Z23 | 0.577 | −0.418 | 0.159 | 0.724 | Z30 | −1.085 | 0.689 | −0.397 | 0.738 | |
| Z29 | 0.130 | −0.104 | 0.026 | 0.708 | Z31 | −0.259 | 0.012 | −0.246 | 0.701 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ning, Y.; Wang, X.; Yang, Y.; Cao, X.; Wu, Y.; Zou, D.; Zhou, D. Studying the Effect of Straw Returning on the Interspecific Symbiosis of Soil Microbes Based on Carbon Source Utilization. Agriculture 2022, 12, 1053. https://doi.org/10.3390/agriculture12071053
Ning Y, Wang X, Yang Y, Cao X, Wu Y, Zou D, Zhou D. Studying the Effect of Straw Returning on the Interspecific Symbiosis of Soil Microbes Based on Carbon Source Utilization. Agriculture. 2022; 12(7):1053. https://doi.org/10.3390/agriculture12071053
Chicago/Turabian StyleNing, Yucui, Xu Wang, Yanna Yang, Xu Cao, Yulong Wu, Detang Zou, and Dongxing Zhou. 2022. "Studying the Effect of Straw Returning on the Interspecific Symbiosis of Soil Microbes Based on Carbon Source Utilization" Agriculture 12, no. 7: 1053. https://doi.org/10.3390/agriculture12071053
APA StyleNing, Y., Wang, X., Yang, Y., Cao, X., Wu, Y., Zou, D., & Zhou, D. (2022). Studying the Effect of Straw Returning on the Interspecific Symbiosis of Soil Microbes Based on Carbon Source Utilization. Agriculture, 12(7), 1053. https://doi.org/10.3390/agriculture12071053