Phylogenetic Analysis of Ryegrass (Lolium rigidum) Populations and the Proliferation of ALS Resistance in Saudi Arabia

 ,
,  , , , , and
, , , , and 
Abstract
1. Introduction
2. Materials and Methods
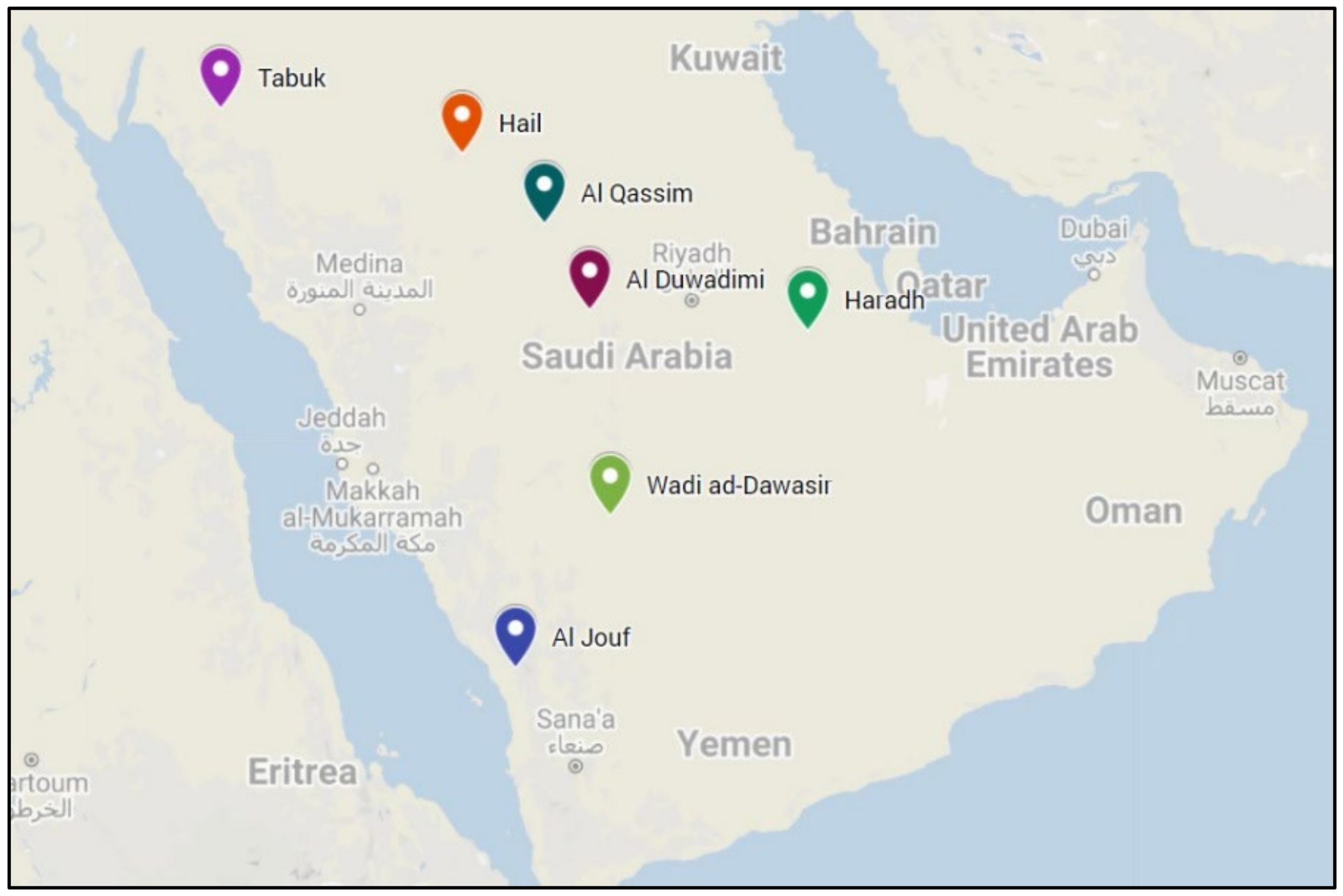
2.1. Field Performance
2.2. Molecular Analysis
3. Results
3.1. Performance of Lolium Populations
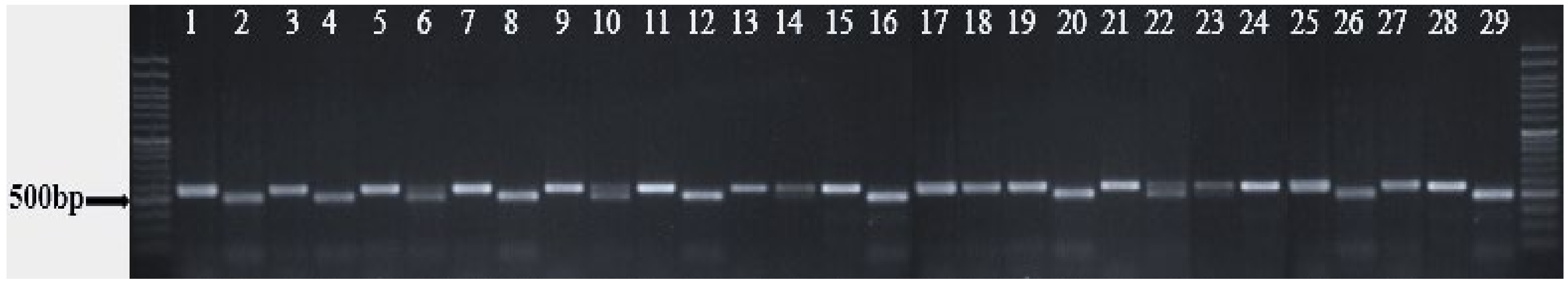
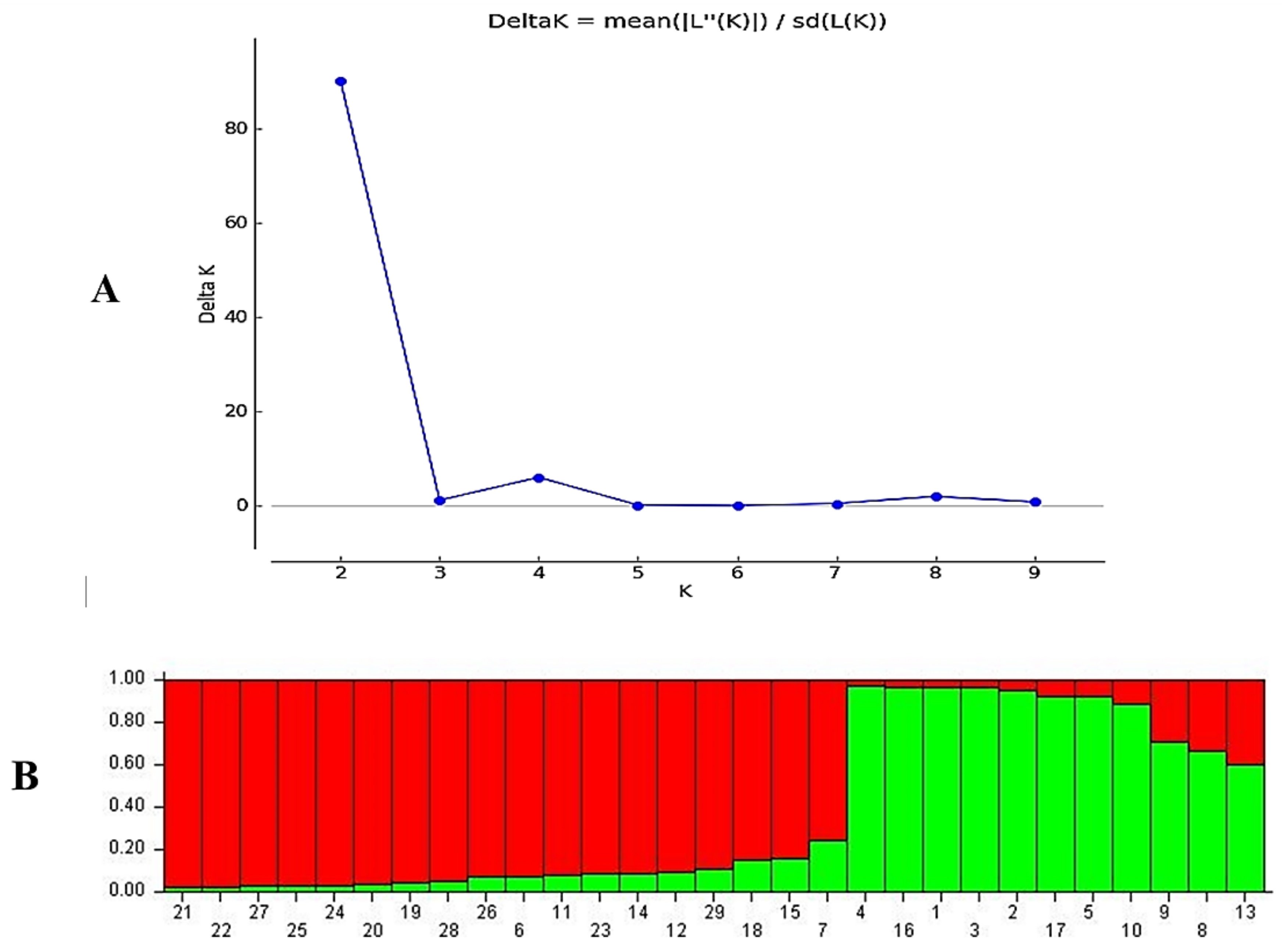
3.2. Molecular Analysis of Lolium Populations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Terrell, E.E. A Taxonomic Revision of the Genus Lolium; Agricultural Research Service, US Department of Agriculture: Washington, DC, USA, 1968. [Google Scholar]
- Spoor, W.; Mccraw, J.M. Self Incompatibility in Lolium—A Reply. Heredity 1984, 53, 239–240. [Google Scholar] [CrossRef]
- Balfourier, F.; Imbert, C.; Charmet, G. Evidence for phylogeographic structure in Lolium species related to the spread of agriculture in Europe. A cpDNA study. Theor. Appl. Genet. 2000, 101, 131–138. [Google Scholar] [CrossRef]
- Kloot, P.M. The Genus Lolium in Australia. Aust. J. Bot. 1983, 31, 421–435. [Google Scholar] [CrossRef]
- Bennett, S.J.J.G.R.; Evolution, C. A phenetic analysis and lateral key of the genus Lolium (Gramineae). Genet. Resour. Crop Evol. 1997, 44, 63–72. [Google Scholar] [CrossRef]
- Hayward, M.D.; Mcadam, N.J.; Jones, J.G.; Evans, C.; Evans, G.M.; Forster, J.W.; Ustin, A.; Hossain, K.G.; Quader, B.; Stammers, M.; et al. Genetic-Markers and the Selection of Quantitative Traits in Forage Grasses. Euphytica 1994, 77, 269–275. [Google Scholar] [CrossRef]
- Hayward, M.D.; Forster, J.W.; Jones, J.G.; Dolstra, O.; Evans, C.; McAdam, N.J.; Hossain, K.G.; Stammers, M.; Will, J.; Humphreys, M.O.; et al. Genetic analysis of Lolium. I. Identification of linkage groups and the establishment of a genetic map. Plant Breed. 1998, 117, 451–455. [Google Scholar] [CrossRef]
- Bert, P.F.; Charmet, G.; Sourdille, P.; Hayward, M.D.; Balfourier, F. A high-density molecular map for ryegrass (Lolium perenne) using AFLP markers. Theor. Appl. Genet. 1999, 99, 445–452. [Google Scholar] [CrossRef]
- Wu, K.S.; Tanksley, S.D. Abundance, polymorphism and genetic mapping of microsatellites in rice. Mol. Gen. Genet. 1993, 241, 225–235. [Google Scholar] [CrossRef]
- Taramino, G.; Tingey, S. Simple sequence repeats for germplasm analysis and mapping in maize. Genome 1996, 39, 277–287. [Google Scholar] [CrossRef]
- Roder, M.S.; Plaschke, J.; Konig, S.U.; Borner, A.; Sorrells, M.E.; Tanksley, S.D.; Ganal, M.W. Abundance, Variability and Chromosomal Location of Microsatellites in Wheat. Molecular and General Genetics 1995, 246, 327–333. [Google Scholar] [CrossRef]
- Ramsay, L.; Macaulay, M.; degli Ivanissevich, S.; MacLean, K.; Cardle, L.; Fuller, J.; Edwards, K.J.; Tuvesson, S.; Morgante, M.; Massari, A.; et al. A simple sequence repeat-based linkage map of barley. Genetics 2000, 156, 1997–2005. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Schertz, K.F.; Cartinhour, S.; Hart, G.E. Comparative genome mapping of Sorghum bicolor (L.) Moench using an RFLP map constructed in a population of recombinant inbred lines. Plant Breed. 1999, 118, 225–235. [Google Scholar] [CrossRef]
- Jones, E.S.; Dupal, M.P.; Kolliker, R.; Drayton, M.C.; Forster, J.W. Development and characterisation of simple sequence repeat (SSR) markers for perennial ryegrass (Lolium perenne L.). Theor. Appl. Genet. 2001, 102, 405–415. [Google Scholar] [CrossRef]
- Saha, M.C.; Mian, R.; Zwonitzer, J.C.; Chekhovskiy, K.; Hopkins, A.A. An SSR- and AFLP-based genetic linkage map of tall fescue (Festuca arundinacea Schreb.). Theor. Appl. Genet. 2005, 110, 323–336. [Google Scholar] [CrossRef]
- Cai, H.W.; Yuyama, N.; Tamaki, H.; Yoshizawa, A. Isolation and characterization of simple sequence repeat markers in the hexaploid forage grass timothy (Phleum pratense L.). Theor. Appl. Genet. 2003, 107, 1337–1349. [Google Scholar] [CrossRef]
- Chen, X.; Cho, Y.G.; McCouch, S.R. Sequence divergence of rice microsatellites in Oryza and other plant species. Mol. Genet. Genom. 2002, 268, 331–343. [Google Scholar] [CrossRef]
- Jones, S.; Dupal, P.; Dumsday, L.; Hughes, J.; Forster, W. An SSR-based genetic linkage map for perennial ryegrass (Lolium perenne L.). Theor. Appl. Genet. 2002, 105, 577–584. [Google Scholar] [CrossRef]
- Jones, E.S.; Mahoney, N.L.; Hayward, M.D.; Armstead, I.P.; Jones, J.G.; Humphreys, M.O.; King, I.P.; Kishida, T.; Yamada, T.; Balfourier, F.; et al. An enhanced molecular marker based genetic map of perennial ryegrass (Lolium perenne) reveals comparative relationships with other Poaceae genomes. Genome 2002, 45, 282–295. [Google Scholar] [CrossRef]
- Cardle, L.; Ramsay, L.; Milbourne, D.; Macaulay, M.; Marshall, D.; Waugh, R. Computational and experimental characterization of physically clustered simple sequence repeats in plants. Genetics 2000, 156, 847–854. [Google Scholar] [CrossRef]
- Al-Doss, A.A.; Ghazy, A.I.; Salem, A.E.-A.K.; Migdadi, H.M.; Al-Faifi, S.A. Agriculture; Environment. Identification and Distribution of ALS resistant Lolium rigidum populations in Saudi Arabia. J. Food Agric. Environ. 2013, 11, 1311–1314. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons: New York, NY, USA, 1984. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- You, F.M.; Huo, N.; Gu, Y.Q.; Luo, M.C.; Ma, Y.; Hane, D.; Lazo, G.R.; Dvorak, J.; Anderson, O.D. BatchPrimer3: A high throughput web application for PCR and sequencing primer design. BMC Bioinform. 2008, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- Jaccard, P. Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Anderson, J.A.; Churchill, G.A.; Autrique, J.E.; Tanksley, S.D.; Sorrells, M.E. Optimizing parental selection for genetic linkage maps. Genome 1993, 36, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Ozkose, A.; Tamkoc, A. Morphological and Agronomic Characteristic of Perennial Ryegrass (Lolium perenne L.) Genotypes. Turk. J. Field Crop. 2014, 19, 231–237. [Google Scholar] [CrossRef][Green Version]
- Acar, Z.; Ayan, I.; Tongel, O.; Mut, H.; Basaran, U. Morphological traits of perennial ryegrass accessions in Black Sea Region of Turkey. In The Contributions of Grasslands to Conservation of Mediterranean Biodiversity; CIHEAM/CIBIO/FAO/SEEP: Zaragoza, Spain, 2010; pp. 117–120. [Google Scholar]
- Barre, P.; Turner, L.B.; Escobar-Gutierrez, A.J. Leaf Length Variation in Perennial Forage Grasses. Agriculture 2015, 5, 682–696. [Google Scholar] [CrossRef]
- Andersen, J.R.; Lubberstedt, T. Functional markers in plants. Trends Plant Sci. 2003, 8, 554–560. [Google Scholar] [CrossRef]
- Saha, M.C.; Mian, M.A.R.; Eujayl, I.; Zwonitzer, J.C.; Wang, L.J.; May, G.D. Tall fescue EST-SSR markers with transferability across several grass species. Theor. Appl. Genet. 2004, 109, 783–791. [Google Scholar] [CrossRef]
- Jensen, A.M.D.; Mikkelsen, L.; Roulund, N. Variation in genetic markers and ergovaline production in endophyte (Neotyphodium)-infected fescue species collected in Italy, Spain, and Denmark. Crop Sci. 2007, 47, 139–147. [Google Scholar] [CrossRef]
- Kuester, A.; Chang, S.M.; Baucom, R.S. The geographic mosaic of herbicide resistance evolution in the common morning glory, Ipomoea purpurea: Evidence for resistance hotspots and low genetic differentiation across the landscape. Evol. Appl. 2015, 8, 821–833. [Google Scholar] [CrossRef] [PubMed]
- Karn, E.; Jasieniuk, M. Genetic diversity and structure of Lolium perenne ssp. multiflorum in California vineyards and orchards indicate potential for spread of herbicide resistance via gene flow. Evol. Appl. 2017, 10, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Al-Doss, A.A.; Al-Faifi, S.A.; Ghazy, A.I.; Salem, A.E.A.; Khan, M.A.; Althamrah, M.; Ammar, M.H.; Farooq, M.; Migdadi, H.M. Resistance of Annual Ryegrass against Acetolactate Synthase-Inhibiting Herbicides in Wheat Fields: Field Evaluation and Molecular Analysis. Int. J. Agric. Biol. 2017, 19, 1401–1408. [Google Scholar] [CrossRef]
- Noli, E.; Conti, S.; Maccaferri, M.; Sanguineti, M.C. Molecular characterization of tomato cultivars. Seed Sci. Technol. 1999, 27, 1–10. [Google Scholar]
- Lee, S.H.; Bailey, M.A.; Mian, M.A.; Carter, T.E., Jr.; Shipe, E.R.; Ashley, D.A.; Parrott, W.A.; Hussey, R.S.; Boerma, H.R. RFLP loci associated with soybean seed protein and oil content across populations and locations. Theor. Appl. Genet. 1996, 93, 649–657. [Google Scholar] [CrossRef]
- Lombard, V.; Baril, C.P.; Dubreuil, P.; Blouet, F.; Zhang, D. Genetic relationships and fingerprinting of rapeseed cultivars by AFLP: Consequences for varietal registration. Crop Sci. 2000, 40, 1417–1425. [Google Scholar] [CrossRef]
- Pejic, I.; Ajmone-Marsan, P.; Morgante, M.; Kozumplick, V.; Castiglioni, P.; Taramino, G.; Motto, M. Comparative analysis of genetic similarity among maize inbred lines detected by RFLPs, RAPDs, SSRs, and AFLPs. Theor. Appl. Genet. 1998, 97, 1248–1255. [Google Scholar] [CrossRef]
- De Riek, J.; Dendauw, J.; Mertens, M.; De Loose, M.; Heursel, J.; Van Bockstaele, E. Validation of criteria for the selection of AFLP markers to assess the genetic variation of a breeders’ collection of evergreen azaleas. Theor. Appl. Genet. 1999, 99, 1155–1165. [Google Scholar] [CrossRef]
- Busi, R.; Yu, Q.; Barrett-Lennard, R.; Powles, S. Long distance pollen-mediated flow of herbicide resistance genes in Lolium rigidum. Theor. Appl. Genet. 2008, 117, 1281–1290. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Genotypes | Resistance index (Resistance Risk) |
|---|---|---|
| 1 | Syngenta | Susceptible |
| 2 | Harad1 | Susceptible |
| 3 | Harad2 | Susceptible |
| 4 | Harad3 | Susceptible |
| 5 | Harad4 | Susceptible |
| 6 | Quaseim1 | Resistance |
| 7 | Quaseim2 | Resistance |
| 8 | Quaseim3 | Resistance |
| 9 | Quaseim4 | Resistance |
| 10 | Haiel1 | Moderate resistance |
| 11 | Haiel2 | Moderate resistance |
| 12 | Haiel3 | Moderate resistance |
| 13 | Haiel4 | Moderate resistance |
| 14 | Gouf1 | Moderate resistance |
| 15 | Gouf2 | Moderate resistance |
| 16 | Gouf3 | Moderate resistance |
| 17 | Gouf4 | Moderate resistance |
| 18 | W. Dawaser1 | Low resistance |
| 19 | W. Dawaser2 | Low resistance |
| 20 | W. Dawaser3 | Low resistance |
| 21 | W. Dawaser4 | Low resistance |
| 22 | Tabouk1 | High resistance |
| 23 | Tabouk2 | High resistance |
| 24 | Tabouk3 | High resistance |
| 25 | Tabouk4 | High resistance |
| 26 | Dwadmi1 | Moderate resistance |
| 27 | Dwadmi2 | Moderate resistance |
| 28 | Dwadmi3 | Moderate resistance |
| 29 | Dwadmi4 | Moderate resistance |
| No. | Primer Name | Forward Sequence (5′→3′) | Reverse Sequence (5′→3′) | Motif l | Reference | EST Putative Function |
|---|---|---|---|---|---|---|
| 1 | LT EST-SSR2 | CTTCATCTTC GGTAGCATCG | CAAAACTTCGGTGGAGCAAT | (TAAA)3 | gi|13661745|gb|AF321856.1| | Lolium rigidum clone FHH-t putative cytochrome P450 mRNA, complete cds |
| 2 | LT EST-SSR4 | TCGTGTTCGGTAGCATCGTA | GCTACAGGAAAACTAGTGCCAAA | (TAAA)4 | gi|13661743|gb|AF321855.1| | Lolium rigidum clone FHH-v putative cytochrome P450 mRNA, complete cds |
| 3 | LT EST-SSR5 | TCTAGACAAATCCGCGTCAA | GAAGCACAGTTTTGGCAAGG | (ATA)4 | gi|28172922|gb|AF343456.1| | Lolium rigidum clone cyt2 cytosolic acetyl-CoA carboxylase (Acc-2) gene, partial cds |
| 4 | LT EST-SSR6 | AAAGAAATGCGAGAGCTGGA | AGGTCATTGTGGGTAGTGTGC | (GCA)4 | gi|257721363|emb|HB811281.1 | Sequence 160495 from Patent EP2090662 Genomic DNA |
| 5 | LT EST-SSR7 | TTACAACCGCTGATGAAGCA | GCCAGGAACTTCACCCTGTA | (GGT)4| | gi|20975573|emb|AJ310767.1 | gi|20975573|emb|AJ310767.1| mRNA |
| 6 | LT EST- SSR8 | GATATGGCTCGGACGAATAA | TCGAATGAAAATGCCAATAA | (at)5 | gi|148340742|gb|EF379036.1| | Lolium rigidum trnT-trnL intergenic spacer, partial sequence; chloroplast |
| 7 | LT EST- SSR9 | AAGGGAATTGGATTTCAGAT | TCGAATGAAAATGCCAATAA | (at)5 | gi|148340740|gb|EF379034.1| | Lolium rigidum trnT-trnL intergenic spacer, partial sequence; chloroplast |
| 8 | LT EST- SSR10 | GGCTCGGACGAATAATCTAA | CGATAAGGATCGAAGGAAAA | (at)5 | gi|148340741|gb|EF379035.1| | Lolium rigidum trnT-trnL intergenic spacer, partial sequence; chloroplast |
| 9 | LT EST-C SSR1 | AGGAGACCATGAGGCTACAC | GCTAGTGCAAAGCCATCAC | (GGA) 3, (TCA)3 (AG)3 | gi|13661761|gb|AF321864.1| | Lolium rigidum clone Lol-5-v putative cytochrome P450 mRNA, complete cds |
| 10 | LT EST-C SSR5 | CAGGGAACAATCATCCAGAC | CCCAAACGGCAGATACAT | (TGC)3 | gi|13661755|gb|AF321861.1| | Lolium rigidum clone Lol-31-b putative cytochrome P450 mRNA, complete cds |
| 11 | LT EST-C SSR6 | CTACCTGCAGTGCATCGTC | TGCCAAAACTTCGGTAGAGT | (TAAA)4 (GCT)3, (CG)4 | gi|13661747|gb|AF321857.1| | Lolium rigidum clone FHH-y putative cytochrome P450 mRNA, complete cds |
| 12 | LT EST-C SSR8 | CACGGACACGATCTACAAGA | GGTGTCCTCGATTGATTCAC | (TGC)3, (GAG)3 | gi|148596801|dbj|AB097496.1| | Lolium rigidum CYP71R4 mRNA for cytochrome P450, complete cds |
| 13 | LT EST- C SSR9 | AGGCTTTTGGTTTACACGAC | CCCCAAAGGACGTACTAAAG | (TTA)3, (CAA)3, (AT)3 | gi|110915824|gb|DQ786854.1| | Lolium rigidum NADH dehydrogenase subunit F (ndhF) gene, partial cds; chloroplast |
| 14 | LT EST- C SSR10 | AGTTGAACCTTGAACGTTATGT | TCGTTTCTGGAGAAGCAACT | (ATA)4 (ATT)3, (AC)3, (AT) 3 | gi|28172922|gb|AF343456.1| | Lolium rigidum clone cyt2 cytosolic acetyl-CoA carboxylase (Acc-2) gene, partial cds |
| 15 | LP EST SSR1 | GTAGTCCAGCGGAGGTCAAT | TACCTGGGCAAATCTTG | (AG(23) | LpSSR006 | Lolium perenne clone LpSSR006 SSR marker sequence |
| 16 | LP EST SSR3 | GGGGAAATACAGTTCTGC | GATGCTCCTGCCTACTTTA | (AG)16, (GA)5, (ATGA)3 | LpSSR020 | Lolium perenne clone LpSSR020 SSR marker sequence |
| 17 | LP EST SSR4 | ATGCACGGGTTTTATTCATT | CGCGAGGCTTAAGGTGT | (TG)20, (GA)23 | LpSSR023 | Lolium perenne clone LpSSR023 SSR marker sequence |
| 18 | LP EST SSR8 | CTAAACTAAATGTTCATCGT | CCTGCTTACTCCTGTT | (AC)26 | LpSSR082 | Lolium perenne clone LpSSR082 SSR marker sequence |
| 19 | LP EST SSR9 | GACCCCGAGACAGCCTA | ACGCATATGGTCTTCAGAA | (TG)9, (GT)10 | LpSSR112 | Lolium perenne clone LpSSR112 SSR marker sequence |
| 20 | LP EST SSR10 | CCGTTGCTTGATACTTGGAC | GAACGAGCATTCTTCCTTTCT | (CTA)7 | DLF008 | library Lolium perenne cDNA clone DLF008, mRNA sequence |
| 21 | LP EST SSR7 | CCCATACTTCGAGGCATAAA | AAATTCCCCCATCAGAGAAC | (AC)29 | LpSSR076 | Lolium perenne clone LpSSR076 SSR marker sequence |
| 22 | LP EST SSR12 | GTGCAGCAGTTTGAATTGGA | AGCATCGGGAGCTATGAATG | (GA)14 | B1A2 (AJ872206.1) | Festuca glaucescens x Lolium multiflorum microsatellite DNA, clone B1-A2 |
| 23 | LP EST SSR13 | AGGTGTCCTGTTGCTTTGGA | TTTACCCCCAGGGATCAAAT | (TG)7 | B1B3 ( AJ872214.1) | Festuca glaucescens x Lolium multiflorum microsatellite DNA, clone B1-B3 |
| 24 | LP EST SSR14 | CCAACTAGACAAAGGGGATTG | GGAGAGCACCATTCATCCAT | (TGA)8 | B2G6a (AJ872228.1) | Festuca glaucescens x Lolium multiflorum microsatellite DNA, clone B2-G6 |
| 25 | TF EST SSR1 | GCACGAGGCTCTTTCCTCTA | GGTGCTTGGCCTTCTTCC | NFA020 | No Hits, PCR amplified Saha, 2004 | |
| 26 | TF EST SSR3 | TCTGCAAGGTCACTGGATCA | GGAGCAAGAAGGACGGAGAC | NFA098 | No Hits, PCR amplified Saha, 2004 | |
| 27 | LP SSR4 | CAAGTGCCACCATAGATACAA | CGTGAAGATCACTATAAACACGA | (AG)8 | LPSSRH01D09 | Kindiger, 2006 |
| 28 | LP SSR6 | TCTGTGGGTCCTTCTGGAT | TCGGGTGATGATGTTGACTT | (TCGC)6 | LPSSRH01F02 | Kindiger, 2006 |
| 29 | LP SSR7 | ATTGACTGGCTTCCGTGTT | CGCGATTGCAGATTCTTG | (CA)9 | LPSSRH01H06 | Kindiger, 2006 |
| 30 | LP SSR8 | TGGAATAACGATGAAAAG | CATCACGAATTAACAAGAG | LPSSRH02D11 | Kindiger, 2006 | |
| 31 | LP SSR9 | GGACGAACTGCCGAGACA | CGGGCATGGTGAGAAGGA | LPSSRK01A03 | Kindiger, 2006 |
| Sov | df | Plant Height | Days to Heading | Panicle Length | No. of Tillers | Fresh Weight | Dry Weight |
|---|---|---|---|---|---|---|---|
| Replecation | 2 | 4.135 | 3.167 | 1.839 | 26,329.6 | 14,163.0 | 558.977 |
| genotypes | 28 | 235.059** | 367.271** | 18.290** | 30,039.1** | 74,686.42** | 3120.700** |
| Error | 56 | 6.604 | 7.208 | 2.042 | 11,978.66 | 20,538.24 | 707.568 |
| Population | Plant Height | Days to Heading | Panicle Length | No. of Tillers | Fresh Weight | Dry Weight |
|---|---|---|---|---|---|---|
| Syngenta | 66.00 | 103.17 | 25.15 | 1240.67 | 1256.50 | 278.35 |
| Harad1 | 82.33 | 77.67 | 27.10 | 1211.33 | 1320.33 | 283.80 |
| Harad2 | 79.67 | 77.33 | 25.33 | 1285.33 | 1407.67 | 305.23 |
| Harad3 | 83.67 | 77.00 | 23.10 | 1346.00 | 1499.67 | 324.87 |
| Harad4 | 86.00 | 74.67 | 26.27 | 1510.33 | 1616.00 | 352.53 |
| Quaseim1 | 79.67 | 77.00 | 28.27 | 1173.00 | 1355.33 | 294.90 |
| Quaseim2 | 76.00 | 76.33 | 23.77 | 1158.67 | 1216.67 | 272.63 |
| Quaseim3 | 79.67 | 78.33 | 23.27 | 1196.00 | 1367.00 | 303.60 |
| Quaseim4 | 84.67 | 77.33 | 25.27 | 1088.00 | 1218.00 | 279.33 |
| Haiel1 | 66.33 | 82.67 | 20.70 | 1103.67 | 1291.33 | 288.30 |
| Haiel2 | 67.67 | 82.00 | 20.97 | 1200.33 | 1488.67 | 312.50 |
| Haiel3 | 64.67 | 83.33 | 19.10 | 1268.67 | 1634.00 | 363.10 |
| Haiel4 | 64.33 | 81.00 | 19.70 | 1378.00 | 1497.00 | 329.40 |
| Gouf1 | 65.67 | 82.67 | 23.83 | 1135.33 | 1384.33 | 300.97 |
| Gouf2 | 62.00 | 81.33 | 22.70 | 1154.33 | 1549.67 | 329.87 |
| Gouf3 | 62.33 | 83.00 | 23.23 | 1377.67 | 1671.33 | 362.23 |
| Gouf4 | 59.33 | 82.67 | 21.23 | 1294.33 | 1337.67 | 285.53 |
| WDawaser1 | 81.67 | 65.33 | 24.27 | 1313.00 | 1723.67 | 374.60 |
| WDawaser2 | 82.67 | 64.67 | 21.87 | 1175.33 | 1327.00 | 301.67 |
| WDawaser3 | 77.67 | 70.67 | 20.87 | 1164.00 | 1319.33 | 299.90 |
| WDawaser4 | 84.33 | 64.33 | 21.47 | 1286.00 | 1524.67 | 330.60 |
| Tabouk1 | 61.67 | 92.67 | 19.93 | 1140.33 | 1735.00 | 377.03 |
| Tabouk2 | 63.33 | 93.00 | 22.17 | 1138.67 | 1411.00 | 300.17 |
| Tabouk3 | 61.00 | 93.00 | 19.20 | 1353.67 | 1385.33 | 309.23 |
| Tabouk4 | 63.67 | 94.67 | 20.87 | 1282.00 | 1661.67 | 365.80 |
| Dwadmi1 | 65.00 | 74.00 | 25.43 | 1150.00 | 1275.00 | 277.70 |
| Dwadmi2 | 66.67 | 76.33 | 24.37 | 1209.67 | 1483.00 | 324.93 |
| Dwadmi3 | 78.00 | 73.33 | 27.00 | 1166.00 | 1286.67 | 292.40 |
| Dwadmi4 | 66.00 | 73.33 | 25.13 | 1139.67 | 1406.67 | 308.60 |
| LSD (0.05) | 4.20 | 4.38 | 2.33 | 178.73 | 234.00 | 43.44 |
| No. | Primer ID | Total Number of Bands | Number of Alleles | Polymorphic Alleles | % Polymorphism | PIC Value |
|---|---|---|---|---|---|---|
| 1 | LT EST-SSR2 | 49 | 2 | 2 | 100 | 0.50 |
| 2 | LT EST-SSR4 | 53 | 3 | 3 | 100 | 0.61 |
| 3 | LT EST-SSR5 | 34 | 2 | 2 | 100 | 0.11 |
| 4 | LT EST-SSR6 | 57 | 4 | 4 | 100 | 0.61 |
| 5 | LT EST-SSR7 | 32 | 1 | 0 | 0 | 0 |
| 6 | LT EST- SSR8 | 32 | 1 | 0 | 0 | 0 |
| 7 | LT EST- SSR9 | 32 | 1 | 0 | 0 | 0 |
| 8 | LT EST- SSR10 | 32 | 1 | 0 | 0 | 0 |
| 9 | LT C SSR1 | 107 | 5 | 5 | 100 | 0.76 |
| 10 | LT C SSR5 | 64 | 3 | 3 | 100 | 0.60 |
| 11 | LT C SSR6 | 61 | 3 | 3 | 100 | 0.50 |
| 12 | LT C SSR8 | 39 | 2 | 2 | 100 | 0.30 |
| 13 | LT C SSR9 | 32 | 1 | 0 | 0 | 0 |
| 14 | LT C SSR10 | 32 | 1 | 0 | 0 | 0 |
| 15 | LP EST SSR1 | 59 | 2 | 2 | 100 | 0.50 |
| 16 | LP EST SSR3 | 34 | 2 | 2 | 100 | 0.46 |
| 17 | LP EST SSR4 | 35 | 2 | 2 | 100 | 0.38 |
| 18 | LP EST SSR8 | 38 | 3 | 3 | 100 | 0.48 |
| 19 | LP EST SSR9 | 57 | 2 | 2 | 100 | 0.50 |
| 20 | LP EST SSR10 | 59 | 3 | 3 | 100 | 0.56 |
| 21 | LP EST SSR7 | 46 | 3 | 3 | 100 | 0.65 |
| 22 | LP EST SSR12 | 42 | 3 | 3 | 100 | 0.65 |
| 23 | LP EST SSR13 | 32 | 3 | 3 | 100 | 0.65 |
| 24 | LP EST SSR14 | 44 | 4 | 4 | 100 | 0.70 |
| 25 | TF EST SSR1 | 53 | 4 | 4 | 100 | 0.69 |
| 26 | TF EST SSR3 | 40 | 3 | 3 | 100 | 0.64 |
| 27 | LP SSR4 | 32 | 1 | 0 | 0 | 0 |
| 28 | LP SSR6 | 32 | 1 | 0 | 0 | 0 |
| 29 | LP SSR7 | 47 | 3 | 3 | 100 | 0.52 |
| 30 | LP SSR8 | 32 | 2 | 2 | 100 | 0.50 |
| 31 | LP SSR9 | 96 | 4 | 4 | 100 | 0.72 |
| Total | 1434 | 75 | 67 | |||
| Average | 46.26 | 2.42 | 2.16 | 74.19 | 0.41 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghazy, A.I.; Al-Ateeq, T.K.; Ibrahim, E.I.; Migdadi, H.M.; Attia, K.A.; Javed, M.; Khan, M.A.; Al-Ashkar, I.; Al-Doss, A. Phylogenetic Analysis of Ryegrass (Lolium rigidum) Populations and the Proliferation of ALS Resistance in Saudi Arabia. Agriculture 2022, 12, 290. https://doi.org/10.3390/agriculture12020290
Ghazy AI, Al-Ateeq TK, Ibrahim EI, Migdadi HM, Attia KA, Javed M, Khan MA, Al-Ashkar I, Al-Doss A. Phylogenetic Analysis of Ryegrass (Lolium rigidum) Populations and the Proliferation of ALS Resistance in Saudi Arabia. Agriculture. 2022; 12(2):290. https://doi.org/10.3390/agriculture12020290
Chicago/Turabian StyleGhazy, Abdelhalim I., Talal K. Al-Ateeq, Eid I. Ibrahim, Hussein M. Migdadi, Kotb A. Attia, Muhammad Javed, Muhammad Altaf Khan, Ibrahim Al-Ashkar, and Abdullah Al-Doss. 2022. "Phylogenetic Analysis of Ryegrass (Lolium rigidum) Populations and the Proliferation of ALS Resistance in Saudi Arabia" Agriculture 12, no. 2: 290. https://doi.org/10.3390/agriculture12020290
APA StyleGhazy, A. I., Al-Ateeq, T. K., Ibrahim, E. I., Migdadi, H. M., Attia, K. A., Javed, M., Khan, M. A., Al-Ashkar, I., & Al-Doss, A. (2022). Phylogenetic Analysis of Ryegrass (Lolium rigidum) Populations and the Proliferation of ALS Resistance in Saudi Arabia. Agriculture, 12(2), 290. https://doi.org/10.3390/agriculture12020290