
Protective Effects of Fucoxanthin Dampen Pathogen-Associated Molecular Pattern (PAMP) Lipopolysaccharide-Induced Inflammatory Action and Elevated Intraocular Pressure by Activating Nrf2 Signaling and Generating Reactive Oxygen Species

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Induction of Endotoxin-Induced Ocular Disorders and Experimental Design
2.3. Measurement of Nrf2 Protein Levels
2.4. Measurement of IOP
2.5. Determination of SOD and Oxidative Stress-Related Malondialdehyde (MDA)
2.6. Inflammatory Cell Counts in the Aqueous Humor
2.7. Immunohistochemistry of Myeloperoxidase (MPO) and Zonula Occludens-1 (ZO-1) in the Ocular Tissues
2.8. Pupillometry of the Pupillary Light Reflex
2.9. Immunohistochemical Analysis of the Iris Nerve Fibers
2.10. Statistical Analysis
3. Results
3.1. Fucoxanthin Upregulates the Expression of Nrf2 and Enhances the Nuclear Translocation of Nrf2 in PAMP LPS-Induced Uveitis
3.2. Protective Effects of Fucoxanthin on LPS-Induced Elevated IOP
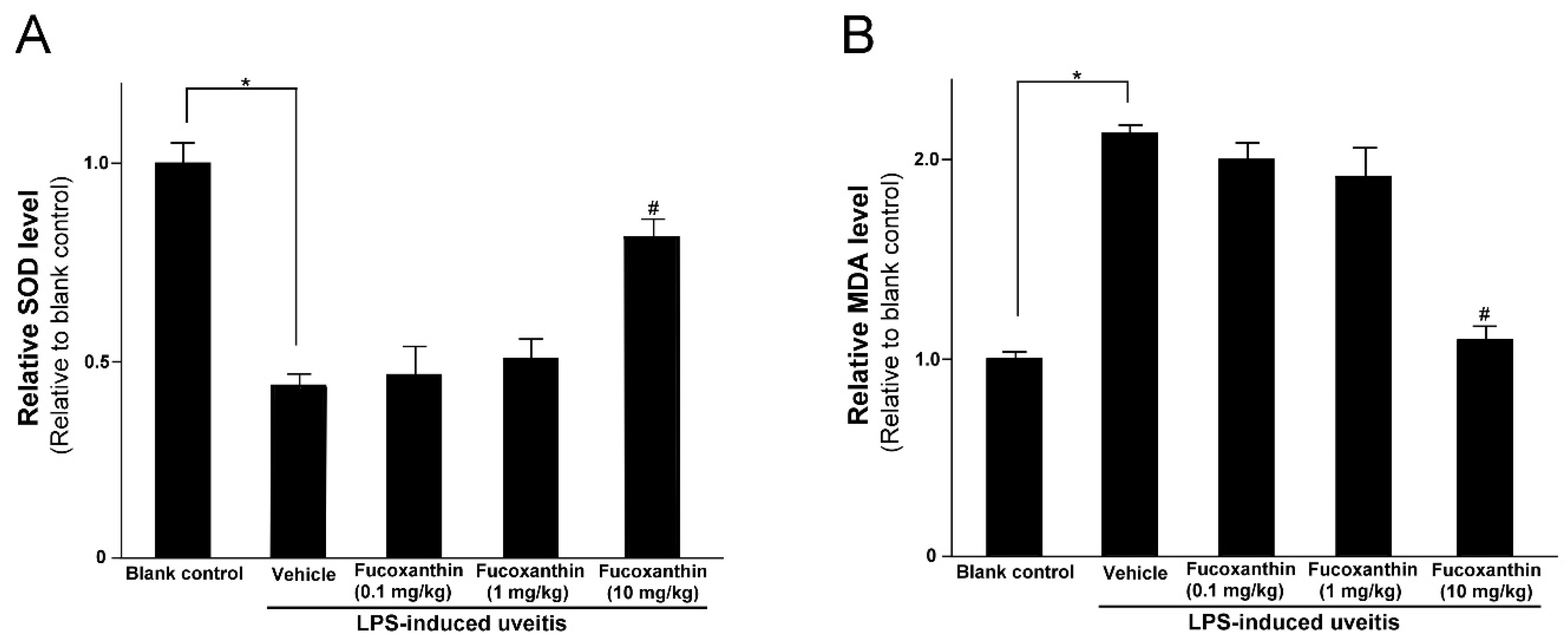
3.3. Protective Effects of Fucoxanthin on SOD Activity and MDA Levels
3.4. Histological Analysis of Ocular Tissues, Inflammatory Cell Counts, and Proinflammatory Cytokine Tumor Necrosis Factor-alpha (TNF-α) Protein Concentration in the Aqueous Humor
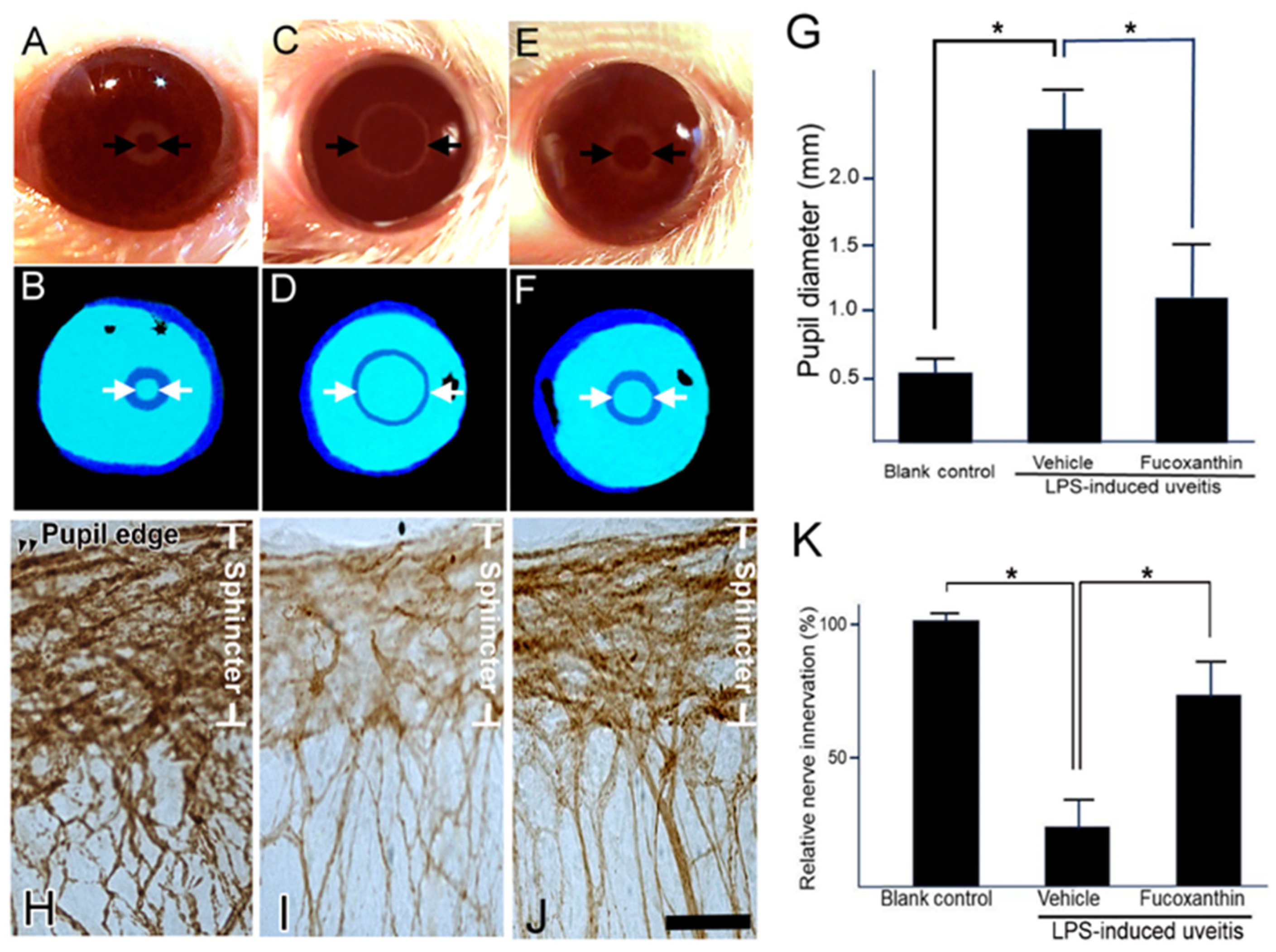
3.5. Effect of Fucoxanthin on the PAMP LPS-Induced Impaired Pupillary Light Reflex and Autonomic Denervation of Iridial Tissues
3.6. Effects of Fucoxanthin on the Cell Infiltration and Endothelial Cell Junctions of Corneal Tissues
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, S.J.; Chang, H.J.; Volin, M.V.; Umar, S.; Van Raemdonck, K.; Chevalier, A.; Palasiewicz, K.; Christman, J.W.; Volkov, S.; Arami, S.; et al. Macrophages are the primary effector cells in IL-7-induced arthritis. Cell Mol. Immunol. 2020, 17, 728–740. [Google Scholar] [CrossRef]
- Liu, Y.L.; Hsu, C.C.; Huang, H.J.; Chang, C.J.; Sun, S.H.; Lin, A.M. Gallic acid attenuated LPS-induced neuroinflammation: Protein aggregation and necroptosis. Mol. Neurobiol. 2020, 57, 96–104. [Google Scholar] [CrossRef]
- Magaña-Guerrero, F.S.; Quiroz-Mercado, J.; Garfias-Zenteno, N.; Garfias, Y. Comparative analysis of inflammatory response in the BALB/c and C57BL/6 mouse strains in an endotoxin-induced uveitis model. J. Immunol. Methods 2020, 476, 112677. [Google Scholar] [CrossRef]
- Ulevitch, R.J.; Tobias, P.S. Receptor-dependent mechanisms of cell stimulation by bacterial endotoxin. Annu. Rev. Immunol. 1995, 13, 437–457. [Google Scholar] [CrossRef]
- Palmieri, B.; Sblendorio, V. Oxidative stress tests: Overview on reliability and use. Part I. Eur. Rev. Med. Pharmacol. Sci. 2007, 11, 309–342. [Google Scholar]
- Klein, J.A.; Ackerman, S.L. Oxidative stress, cell cycle, and neurodegeneration. J. Clin. Investig. 2003, 111, 785–793. [Google Scholar] [CrossRef]
- Kannan, K.; Jain, S.K. Oxidative stress and apoptosis. Pathophysiology 2000, 7, 153–163. [Google Scholar] [CrossRef]
- Gueudry, J.; Muraine, M. Anterior uveitis. J. Fr. Ophtalmol. 2018, 41, e11–e21. [Google Scholar] [CrossRef] [PubMed]
- Qiu, R.; Yang, M.; Wang, W.; Liu, J.; Yang, L.; Lei, B. The protective effects of VVN001 on LPS-induced inflammatory responses in human RPE cells and in a mouse model of EIU. Inflammation 2021, 44, 780–794. [Google Scholar] [CrossRef]
- Liang, W.C.; Ren, J.L.; Yu, Q.X.; Li, J.; Ng, T.K.; Chu, W.K.; Qin, Y.J.; Chu, K.O.; Schally, A.V.; Pang, C.P.; et al. Signaling mechanisms of growth hormone-releasing hormone receptor in LPS-induced acute ocular inflammation. Proc. Natl. Acad. Sci. USA 2020, 117, 6067–6074. [Google Scholar] [CrossRef] [PubMed]
- Shoeb, M.; Zhang, M.; Xiao, T.; Syed, M.F.; Ansari, N.H. Amelioration of endotoxin-induced inflammatory toxic response by a metal chelator in rat eyes. Investig. Ophthalmol. Vis. Sci. 2018, 59, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Hui, B.T.K.; Capewell, N.; Ansari, Y.; Liu, X. Bilateral acute anterior uveitis and iris atrophy caused by moxifloxacin. BMJ Case Rep. 2020, 13, e233528. [Google Scholar] [CrossRef]
- Alaghband, P.; Baneke, A.J.; Galvis, E.; Madekurozwa, M.; Chu, B.; Stanford, M.; Overby, D.; Lim, K.S. Aqueous humor dynamics in uveitic eyes. Am. J. Ophthalmol. 2019, 208, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Kalogeropoulos, D.; Sung, V.C. Pathogenesis of uveitic glaucoma. J. Curr. Glaucoma Pract. 2018, 12, 125–138. [Google Scholar] [CrossRef]
- Ghita, A.C.; Ilie, L.; Ghita, A.M. The effects of inflammation and anti-inflammatory treatment on corneal endothelium in acute anterior uveitis. Rom. J. Ophthalmol. 2019, 63, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Trinh, L.; Brignole-Baudouin, F.; Labbé, A.; Raphaël, M.; Bourges, J.L.; Baudouin, C. The corneal endothelium in an endotoxin-induced uveitis model: Correlation between in vivo confocal microscopy and immunohistochemistry. Mol. Vis. 2008, 14, 1149–1156. [Google Scholar] [CrossRef]
- Zemba, M.; Camburu, G. Uveitis-glaucoma-hyphaema syndrome. Rom. J. Ophthalmol. 2017, 61, 11–17. [Google Scholar] [CrossRef]
- Hejazian, S.M.; Hosseiniyan Khatibi, S.M.; Barzegari, A.; Pavon-Djavid, G.; Razi Soofiyani, S.; Hassannejhad, S.; Ahmadian, E.; Ardalan, M.; Zununi Vahed, S. Nrf-2 as a therapeutic target in acute kidney injury. Life Sci. 2021, 264, 118581. [Google Scholar] [CrossRef]
- Ikram, M.; Park, T.J.; Ali, T.; Kim, M.O. Antioxidant and neuroprotective effects of caffeine against Alzheimer’s and Parkinson’s disease: Insight into the role of Nrf-2 and A2AR signaling. Antioxidants 2020, 9, 902. [Google Scholar] [CrossRef]
- Yang, P.M.; Cheng, K.C.; Yuan, S.H.; Wung, B.S. Carbon monoxide-releasing molecules protect against blue light exposure and inflammation in retinal pigment epithelial cells. Int. J. Mol. Med. 2020, 46, 1096–1106. [Google Scholar] [CrossRef]
- Choi, J.I.; Kim, J.; Choung, S.Y. Polyphenol-enriched fraction of Vaccinium uliginosum L. protects selenite-induced cataract formation in the lens of Sprague-Dawley rat pups. Mol. Vis. 2019, 25, 118–128. [Google Scholar]
- Chen, S.J.; Lee, C.J.; Lin, T.B.; Peng, H.Y.; Liu, H.J.; Chen, Y.S.; Tseng, K.W. Protective Effects of fucoxanthin on ultraviolet B-induced corneal denervation and inflammatory pain in a rat model. Mar. Drugs 2019, 17, 152. [Google Scholar] [CrossRef]
- Thounaojam, M.C.; Powell, F.L.; Patel, S.; Gutsaeva, D.R.; Tawfik, A.; Smith, S.B.; Nussbaum, J.; Block, N.L.; Martin, P.M.; Schally, A.V.; et al. Protective effects of agonists of growth hormone-releasing hormone (GHRH) in early experimental diabetic retinopathy. Proc. Natl. Acad. Sci. USA 2017, 114, 13248–13253. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Kikuchi, M.; Kubodera, A.; Kawakami, Y. Proton-donative antioxidant activity of fucoxanthin with 1,1-diphenyl-2-picrylhydrazyl (DPPH). Biochem. Mol. Biol. Int. 1997, 42, 361–370. [Google Scholar] [CrossRef]
- Chen, S.J.; Lee, C.J.; Lin, T.B.; Liu, H.J.; Huang, S.Y.; Chen, J.Z.; Tseng, K.W. Inhibition of ultraviolet B-induced expression of the proinflammatory cytokines TNF-α and VEGF in the cornea by fucoxanthin treatment in a rat model. Mar. Drugs 2016, 14, 13. [Google Scholar] [CrossRef]
- Peng, J.; Yuan, J.P.; Wu, C.F.; Wang, J.H. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Shiratori, K.; Ohgami, K.; Ilieva, I.; Jin, X.H.; Koyama, Y.; Miyashita, K.; Yoshida, K.; Kase, S.; Ohno, S. Effects of fucoxanthin on lipopolysaccharide-induced inflammation in vitro and in vivo. Exp. Eye Res. 2005, 81, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Horng, C.T.; Chen, Y.H.; Chen, P.L.; Chen, C.L.; Liang, C.M.; Chien, M.W.; Chen, J.T. Inhibitory effects of glucosamine on endotoxin-induced uveitis in Lewis rats. Investig. Ophthalmol. Vis. Sci. 2008, 49, 5441–5449. [Google Scholar] [CrossRef]
- Tseng, K.W.; Peng, M.L.; Wen, Y.C.; Liu, K.J.; Chien, C.L. Neuronal degeneration in autonomic nervous system of Dystonia musculorum mice. J. Biomed. Sci. 2011, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.Q.; Zhang, M.; Matis, K.I.; Kim, C.; Rosenblatt, M.I. Vascular endothelial growth factor mediates corneal nerve repair. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3870–3878. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta. Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Nagai, N.; Thimmulappa, R.K.; Cano, M.; Fujihara, M.; Izumi-Nagai, K.; Kong, X.; Sporn, M.B.; Kensler, T.W.; Biswal, S.; Handa, J.T. Nrf2 is a critical modulator of the innate immune response in a model of uveitis. Free Radic. Biol. Med. 2009, 47, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Han, H.; Liu, J.; Tang, M.; Wu, X.; Cao, X.; Zhao, T.; Lu, Y.; Niu, T.; Chen, J.; et al. Fucoxanthin prevents 6-OHDA-induced neurotoxicity by targeting Keap1. Oxid. Med. Cell Longev. 2021, 2021, 6688708. [Google Scholar] [CrossRef]
- Hu, R.; Saw, C.L.; Yu, R.; Kong, A.N. Regulation of NF-E2-related factor 2 signaling for cancer chemoprevention: Antioxidant coupled with antiinflammatory. Antioxid. Redox Signal. 2010, 13, 1679–1698. [Google Scholar] [CrossRef] [PubMed]
- Jang, E.J.; Kim, S.C.; Lee, J.H.; Lee, J.R.; Kim, I.K.; Baek, S.Y.; Kim, Y.W. Fucoxanthin, the constituent of Laminaria japonica, triggers AMPK-mediated cytoprotection and autophagy in hepatocytes under oxidative stress. BMC Complement. Altern. Med. 2018, 8, 97. [Google Scholar] [CrossRef]
- Eming, S.A.; Wynn, T.A.; Martin, P. Inflammation and metabolism in tissue repair and regeneration. Science 2017, 356, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ji, Y. A literature review on Fuchs uveitis syndrome: An update. Surv. Ophthalmol. 2020, 65, 133–143. [Google Scholar] [CrossRef]
- Pleyer, U.; Sherif, Z. Corticosteroids in ophthalmology. Ernst Scher. Res. 2002, 40, 65–81. [Google Scholar]
- Tuft, S.J.; Coster, D.J. The corneal endothelium. Eye 1990, 4, 389–424. [Google Scholar] [CrossRef] [PubMed]
- Nejsum, L.N.; Piec, A.; Fijak, M.; Ernstsen, C.V.; Fischer, D.; Maier, T.J.; Kinscherf, R.; Hofmann, R.; Urbschat, A. Systemic LPS induces toll-like receptor 3 (TLR3) expression and apoptosis in testicular mouse tissue. Cell Tissue Res. 2019, 378, 143–154. [Google Scholar] [CrossRef]
- Savran, M.; Ozmen, O.; Erzurumlu, Y.; Savas, H.B.; Asci, S.; Kaynak, M. The Impact of Prophylactic Lacosamide on LPS-Induced Neuroinflammation in Aged Rats. Inflammation 2019, 42, 1913–1924. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.C.; Wang, M.L.; Wu, D.B.; Luo, C.; Tang, H.; Chen, E.Q. Curcumin alleviates lipopolysaccharide induced sepsis and liver failure by suppression of oxidative stress-related inflammation via PI3K/AKT and NF-κB related signaling. J. Transl. Med. 2019, 17, 151. [Google Scholar] [CrossRef]
- Uchida, T.; Honjo, M.; Yamagishi, R.; Aihara, M. The Anti-Inflammatory Effect of ripasudil (K-115), a rho kinase (ROCK) inhibitor, on endotoxin-induced uveitis in rats. Investig. Ophthalmol. Vis. Sci. 2017, 58, 5584–5593. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Gambino, F., Jr.; Algenio, C.S.; Wu, C.; Gao, Y.; Bouchard, C.S.; Qiao, L.; Bu, P.; Zhao, S. Inflammation and oxidative stress induced by lipid peroxidation metabolite 4-hydroxynonenal in human corneal epithelial cells. Graefes. Arch. Clin. Exp. Ophthalmol. 2020, 258, 1717–1725. [Google Scholar] [CrossRef]
- Perez-Garmendia, R.; Lopez de Eguileta Rodriguez, A.; Ramos-Martinez, I.; Zuñiga, N.M.; Gonzalez-Salinas, R.; Quiroz-Mercado, H.; Zenteno, E.; Hernández, E.R.; Hernández-Zimbrón, L.F. Interplay between oxidative stress, inflammation, and amyloidosis in the anterior segment of the eye; its pathological implications. Oxid. Med. Cell Longev. 2020, 2020, 6286105. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, J.T.; Lee, S.J.; Kim, J.C. The anti-inflammatory effects of angiogenin in an endotoxin induced uveitis in rats. Int. J. Mol. Sci. 2020, 21, 413. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, W.; Lu, H.; Hu, X.; Li, S.; Wang, J.; Zhao, L. The expression of cytokines in the aqueous humor and serum during endotoxin-induced uveitis in C3H/HeN mice. Mol. Vis. 2010, 16, 1689–1695. [Google Scholar]
- Donmez, O.; Yaman, A.; Ozturk, T.; Aktas, S.; Altun, Z.S.; Yılmaz, O. The efficacy of systemic and intravitreal infliximab treatments in an endotoxin-induced uveitis model. Cutan. Ocul. Toxicol. 2019, 38, 360–369. [Google Scholar] [CrossRef]
- Ge, Q.; Wang, S.; Zheng, Y. Systemic administration of an anti-tumor necrosis factor-alpha monoclonal antibody protects against endotoxin-induced uveitis in rats. Indian J. Ophthalmol. 2016, 64, 873–877. [Google Scholar] [CrossRef]
- Song, H.B.; Park, S.Y.; Ko, J.H.; Park, J.W.; Yoon, C.H.; Kim, D.H.; Kim, J.H.; Kim, M.K.; Lee, R.H.; Prockop, D.J.; et al. Mesenchymal stromal cells inhibit inflammatory lymphangiogenesis in the cornea by suppressing macrophage in a TSG-6-dependent manner. Mol. Ther. 2018, 26, 162–172. [Google Scholar] [CrossRef]
- Semkova, I.; Muether, P.S.; Kuebbeler, M.; Meyer, K.L.; Kociok, N.; Joussen, A.M. Contribution of TNF-alpha to leukocyte adhesion, vascular leakage, and apoptotic cell death in endotoxin-induced uveitis in vivo. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6101–6108. [Google Scholar] [CrossRef]
- Daniel, E.; Ebenezer, G.J.; Job, C.K. Pathology of iris in leprosy. Br. J. Ophthalmol. 1997, 81, 490–492. [Google Scholar] [CrossRef][Green Version]
- Tan, E.T.; Lambie, D.G.; Johnson, R.H.; Whiteside, E.A. Parasympathetic denervation of the iris in alcoholics with vagal neuropathy. J. Neurol. Neurosurg Psychiatry 1984, 47, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Whitcup, S.M.; Wakefield, D.; Li, Q.; Nussenblatt, R.B.; Chan, C.C. Endothelial leukocyte adhesion molecule-1 in endotoxin-induced uveitis. Investig. Ophthalmol. Vis. Sci. 1992, 33, 2626–2630. [Google Scholar]
- Wang, Y.; Zhang, Z.; Zhang, L.; Li, X.; Lu, R.; Xu, P.; Zhang, X.; Dai, M.; Dai, X.; Qu, J.; et al. S100A8 promotes migration and infiltration of inflammatory cells in acute anterior uveitis. Sci. Rep. 2016, 6, 36140. [Google Scholar] [CrossRef]
- Eom, Y.; Lee, D.Y.; Kang, B.R.; Heo, J.H.; Shin, K.H.; Kim, H.M.; Song, J.S. Comparison of aqueous levels of inflammatory mediators between toxic anterior segment syndrome and endotoxin-induced uveitis animal models. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6704–6710. [Google Scholar] [CrossRef] [PubMed]
- Rajashekhar, G.; Shivanna, M.; Kompella, U.B.; Wang, Y.; Srinivas, S.P. Role of MMP-9 in the breakdown of barrier integrity of the corneal endothelium in response to TNF-α. Exp. Eye Res. 2014, 122, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, S.P. Cell signaling in regulation of the barrier integrity of the corneal endothelium. Exp. Eye Res. 2012, 95, 8–15. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-J.; Lin, T.-B.; Peng, H.-Y.; Lin, C.-H.; Lee, A.-S.; Liu, H.-J.; Li, C.-C.; Tseng, K.-W. Protective Effects of Fucoxanthin Dampen Pathogen-Associated Molecular Pattern (PAMP) Lipopolysaccharide-Induced Inflammatory Action and Elevated Intraocular Pressure by Activating Nrf2 Signaling and Generating Reactive Oxygen Species. Antioxidants 2021, 10, 1092. https://doi.org/10.3390/antiox10071092
Chen S-J, Lin T-B, Peng H-Y, Lin C-H, Lee A-S, Liu H-J, Li C-C, Tseng K-W. Protective Effects of Fucoxanthin Dampen Pathogen-Associated Molecular Pattern (PAMP) Lipopolysaccharide-Induced Inflammatory Action and Elevated Intraocular Pressure by Activating Nrf2 Signaling and Generating Reactive Oxygen Species. Antioxidants. 2021; 10(7):1092. https://doi.org/10.3390/antiox10071092
Chicago/Turabian StyleChen, Shiu-Jau, Tzer-Bin Lin, Hsien-Yu Peng, Cheng-Hsien Lin, An-Sheng Lee, Hsiang-Jui Liu, Chun-Chieh Li, and Kuang-Wen Tseng. 2021. "Protective Effects of Fucoxanthin Dampen Pathogen-Associated Molecular Pattern (PAMP) Lipopolysaccharide-Induced Inflammatory Action and Elevated Intraocular Pressure by Activating Nrf2 Signaling and Generating Reactive Oxygen Species" Antioxidants 10, no. 7: 1092. https://doi.org/10.3390/antiox10071092
APA StyleChen, S.-J., Lin, T.-B., Peng, H.-Y., Lin, C.-H., Lee, A.-S., Liu, H.-J., Li, C.-C., & Tseng, K.-W. (2021). Protective Effects of Fucoxanthin Dampen Pathogen-Associated Molecular Pattern (PAMP) Lipopolysaccharide-Induced Inflammatory Action and Elevated Intraocular Pressure by Activating Nrf2 Signaling and Generating Reactive Oxygen Species. Antioxidants, 10(7), 1092. https://doi.org/10.3390/antiox10071092