Abstract
Beekeepers can use a variety of treatments against Varroa destructor, the parasitic mite of Apis mellifera. However, sustainable and easy-to-use solutions are still scarce, considering the complexity of reaching the parasite alone. Current treatments involve soft acaricides, although their mechanism of action is not well understood. We investigated the effects of organic acids such as tartaric, lactic, formic, and citric acids on the attachment abilities of V. destructor under laboratory conditions. Preventing parasites from gripping or holding on to their hosts is a crucial target for mite control strategies. We challenged grip skills through the Rotavar setup after the direct application of acids to mites’ arolia. We also tested the potential for mites to fall off honeybees after bee treatment. We found that tartaric, citric, and lactic acids were good candidates to impair the attachment of V. destructor twenty-four hours post-treatment. However, lactic acid remained the only candidate at a reasonable concentration to destabilise mites after the honey bee’s treatment without reducing their lifespan. While we conducted these experiments under artificial conditions, our results improved our comprehension of the organic acids’ potential impact on V. destructor. They can also help with the development of new methods for hive application for beekeepers worldwide.
1. Introduction
Varroa destructor is one of the major threats against Apis mellifera in Europe [1]. This ectoparasite transmits and replicates viruses such as the Deformed Wing Virus (DWV) [2,3,4]. This chronic virus during varroosis causes wing malformations, one of the key factors in colony weakening [1,5,6]. Varroa destructor females feed on honeybees’ haemolymph and fat bodies [7,8]. They use adult honeybees as feeders and carriers during what is called the dispersal phase [9]. Several methods have been applied to control V. destructor, from hard acaricides (synthetic molecules [9,10]) such as pyrethroids, formamidine, organophosphate, neonicotinoid, or sulfoximines to organic acids (natural molecules) such as oxalic or formic acids. Former treatments are known for their negative impact on honeybees’ health and can create resistance in mites [11,12]. Latters, from natural origins, are considered key alternatives and can be efficient treatments against V. destructor with fewer drawbacks [13,14,15]. Organic acids such as formic, citric, lactic, and tartaric acids are of particular interest due to their natural presence in honey, avoiding the addition of alien molecules to honeybees [16,17]. Formic and oxalic acids are frequently used as alternative treatments against V. destructor [17]; however, little is known about lactic, citric, or tartaric acids. Formic acid is a volatile molecule that reaches dispersal and reproductive mites inside a hive, whereas oxalic acid only defeats dispersive mites [18,19,20]. The specific mode of action of oxalic acid remains unclear. Although it is suspected to create crystals on the arolia of mites, mechanically leading to their death [21,22]. In a similar vein, a new perspective to study and better apply current organic treatments would be to challenge the host attachment ability of mites by exposing their arolia [23]. Indeed, attachment is an essential part of the interaction between a parasite and its host. The study of attachment ability for ticks such as Ixodes ricinus showed a new biological target for tick control strategies: to prevent grip [23,24,25]. For V. destructor, chemical orientation with repellent or attractive molecules was highly explored as a potential control strategy; however, little is known about their attachment abilities [26,27,28]. Arolia are soft pads found at the end of most parasites’ pretarsus and are characterised, for some, as adhesive disc-like structures necessary to walk and grip their hosts, especially for mites and ticks [29]. In ticks, a biphasic fluid with a hydrophilic and hydrophobic phase mediates the stickiness under the arolia [5]; however, the existence of such a fluid in Varroa mites remains unknown as their biology is not fully understood yet [10,30].
In this work, we investigate for the first time if organic acids such as formic, citric, tartaric, and lactic acids can interfere with the host attachment ability of V. destructor. We chose current concentrations of in-the-field organic acids such as formic or lactic acids [31,32] but also less-in-use candidates such as citric and tartaric acids to compare [33]. In order to test their grip skills, we treated V. destructor females’ arolia directly and used a homemade machine with a wooden rotative substrate called Rotavar. Moreover, we investigated host attachment by treating honeybees with organic acids and checking the number of fallen mites (Figure 1).

Figure 1.
Schematic diagram of the experimental design and measured parameters. Grip on wood (Rotavar): This method relies on direct contact between Varroa’s arolia and the organic acids. The Rotavar set-up is a motor-driven rotating toothpick used to assess V. destructor’s grip. Grip on bees: the host attachment experiment applies acids to the backs of honeybees to remove mites. T0 represents the administration time for treatments; T + 1 h 30, 24 h, 48 h, or 72 h stand for the time post administration used to make measurements.
2. Materials and Methods
Experiments were conducted in 2022 and early 2023 during spring, summer, and autumn using bees from three Buckfast colonies and their mites provided by ADA Occitanie, France. Colonies were maintained on the university campus (INU Champollion, Albi, France). They were fed sugar syrup and pollen in February to boost egg production. They were only treated the previous year with oxalic acid for a month, so the V. destructor infestation remained high throughout the time. No treatment was applied either during the experiments or during the previous six months.
2.1. Mites and Honeybees Sampling
Varroa destructor females were collected from adult honeybees with a soft paintbrush [34] (dispersal phase), L5 larvae, or emerging bees (varroas with homogenised body shapes) and taken to the laboratory in the morning. They were randomised to avoid state bias. They were stored for 2 h maximum on pupae to feed ad libitum (34.5 °C, 70% Relative Humidity (RH)) in an incubator before the start of experiments.
Worker honeybees were collected in the morning from food frames and kept for the experiment. They were stored for 2 h maximum in experimental cages (Pain type: 10.5 cm × 7.5 cm × 11.5 cm) in an incubator (28 °C, 60% RH) before the start of the experiment. They were fed ad libitum with a gravity feeder delivering sucrose at 50% (w/v).
2.2. Acid Preparation and Control
Acids were purchased from Sigma-Aldrich, France. All solutions were prepared by M2I Development (Lacq, France). Dilutions of lactic acid (99%) (CAS no. 50-21-5), citric acid anhydre (Omnipur—99% purity) (CAS no. 77-92-9), formic acid (liquid 50–80%) (CAS no. 64-18-6) and tartaric acid (D-tartaric, 99%) (CAS no. 147-71-7) were made with demineralized water (Ciron, France) and sampled as indicated below (see Table 1). The solvent, demineralized water, was used to treat the control group. All stock solutions were kept at 4 °C.

Table 1.
Acid concentrations and sample size according to the type of experiment.
2.3. Grip on Wood (Rotavar)
Acid administration—Concentrations of acids and sample sizes of experiments are described in Table 1. We chose a different range of concentrations for formic acid according to the currently available treatment on the market, such as Varromed® (5 mg/mL of formic acid). The highest concentration of citric acid is 580 mg/mL instead of 600 due to the solubilisation limit.
For the treatment procedure, 200 µL of diluted acids or demineralized water (control group) were added in a Petri dish (5 cm diameter) covered with a 5 cm diameter piece of paper (Scott® Slimroll, Roswell, GA, USA). Mites were able to walk freely on the wet, impregnated paper for 3 min. Then, each of them was transferred into a gelatin capsule (LGA, La Seyne sur Mer, France) with a late honeybee pupa to feed on and kept at 34.5 °C (70% RH) in an incubator according to the method developed by Piou et al. (2016) [35].
Set up—An adapted set up, the Rotavar [36], was used to assess the grip skill of V. destructor on wood (Figure 1). Briefly, the Rotavar was inspired by the Rotarod for rodents [37]. The Rotavar machine was composed of a motor-driven rotating toothpick (6 cm long and 2 mm in diameter). It allowed us to standardise the time head-up and head-down of the mite as well as transitions, in our case 3 RPM (revolutions per minute). Indeed, mites were naturally equipped to walk 360°. As the toothpick rotated, V. destructor had to stick to prevent a fall. Mites with deficits affecting their grip were therefore expected to drop during the experiment. One and a half hours after the treatment, each mite was placed on the rotating toothpick (3 cm from both sides) until they fell, or until the end of the test (3 min). The time before falling was recorded. After this first test, V. destructor females were put back in their respective gelatin capsules and kept in the incubator (34.5 °C 70% RH). Dead mites were retrieved from the experiment. The Rotavar experiment was repeated 24 h post-treatment to assess the “long-term” grip deficit. Mites were kept in the incubator until 48 h after treatment to check mortality. The live ones were frozen.
2.4. Grip on Host Experiment (Honeybees)
Acid administration—Two days prior to the experiment, mites were collected in brood cells and put on adult bees to simulate and standardise the dispersal phase in laboratory conditions [35]. Hosts and mites were kept in experimental cages (Pain type: 10.5 cm × 7.5 cm × 11.5 cm) at 28 °C and 60–65% RH. Each bee carrying one or two mites received 5 µL on the back, from the thorax to the abdomen (5 micro drops), of diluted acids or demineralized water (control group) according to the procedure developed by Gashout et al. (2009) [38]. Honeybees were kept for 5 min to let the solutions dry and stick to their backs. Concentrations of acids and sample sizes of experiments were used as described in Table 1.
Set up—Once acids were administered to honeybees, micro-colonies of 5 treated individuals were placed in pierced plastic glasses (7 cm diam, 9.5 cm height) on Petri dishes (90 mm diam) covered with Whatman paper [39,40]. Honeybees were fed sucrose at 50% (w/v) ad libitum. The number of fallen mites (which could be dead or alive but not able to grip a honeybee anymore) and bees’ mortality were checked and removed every 24 h for 72 h (Figure 1). At the end of each experiment, the number of Varroa females still on honeybees was counted to prevent any mistakes due to mite escape.
2.5. Statistical Analyses
The statistical analysis of data was carried out using standard methods on R (version 4.0.5) with RStudio (version 1.3.1093) loaded with packages RVAideMemoire, Car, GGplot2, Survival, and Survminer [41,42,43]. Results for grip assays were presented as percentages with a confidence interval of 95% [CI95]. The proportion of fallen mites was compared using a Binomial GLM (Generalised linear model) with treatments when the proportion was different from 1. It was followed by a Fisher exact test for binary variables with a Bonferroni correction to further analyse the pairwise comparison with the control group. When the Binomial GLM was not possible, the Fisher exact test was conducted directly. For grip skill median time before falling, a Shapiro test was used to check the data normality; thus, the Kruskal-Wallis test followed by a Dunn Test with Bonferroni correction was used when needed. Survival probability was analysed over two days through the Kaplan-Meier method for mites and over three days for honeybees.
3. Results
3.1. Organic Acids Impact on V. destructor Grip Skill on Wood
Control—Mites from the control group were treated with demineralized water. As V. destructor is naturally able to grip and rotate 360°, no fallen mites from the Rotavar were expected. However, one-and-a-half-hours post-treatment, 1.11% [CI95: 4.84–12.73] of control mites fell (represented by the black dashed line and CI95 represented by the grey area around the dashed line (Figure 2)). Twenty-four hours post-treatment, 1.19% [CI95: 6.72–21.91] of control mites fell. For grip duration, the control group is represented by “0” on the boxplot. The median time for each control group is 180 s (the maximum of the experiment), as most of the control mites did not fall (Figure 3).
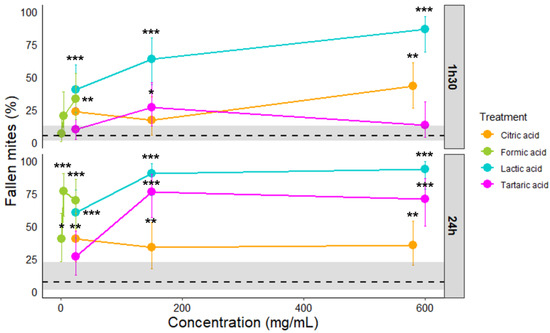
Figure 2.
Effects of citric, formic, lactic, and tartaric acid concentrations on V. destructor females’ grip skill via the Rotavar test one hour and a half as well as twenty-four-hours post-treatment. Dashed line represents the control group. The grey area around dashed line represents the confidence interval of 95%. Color bars represent confidence interval of 95%; *** stands for p < 0.001, ** for p < 0.01, and * for p < 0.05 compared to the control (see Table 2 for detailed statistics).
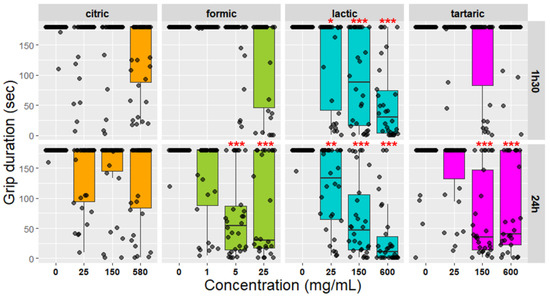
Figure 3.
Grip duration of mites on the Rotavar (in seconds), 1 h 30, and 24 h post exposition to citric, formic, lactic, or tartaric acid concentrations. “0” represents the control group in each acid experiment. *** stands for p < 0.001, ** for p < 0.01, and * for p < 0.05 compared to the control (see Table 2 for detailed statistics).
Citric acid—Varroa destructor females exposed to 580 mg/mL of citric acid fell significantly more than mites from the control group one hour and a half post-treatment (Figure 2, Binomial GLM p-value = 0.0023; see Table A1 and Table 2 for Fisher test). Twenty-four hours post-treatment, they significantly fell more from the Rotavar regardless of the concentration (Figure 2, Binomial GLM p-value for all <0.05, see Table A1 and Table 2). However, there is an overlap in the 95% confidence intervals between the control group and the citric acid-treated groups. For grip duration, one hour and a half post-treatment, the median duration was not significantly different from the control group. Twenty-four-hours post-treatment, no difference was found between the grip duration of mites treated with citric acid and control mites (Figure 3).
Table 2.
Outcome of statistical analyses for percentages of fallen mites and median grip duration for Rotavar grip skill experiment—(A) Statistical analyses with adjusted p-value using Fisher’s exact test with Bonferroni correction. (B) Statistical analyses with adjusted p-value using Dunn test with Bonferroni correction (vs. = versus; Ctrl = Control).
Formic acid—One hour and a half after formic acid exposition at 25 mg/mL, mites dropped significantly more than the control group (Figure 2, Fisher exact test, p-value = 0.0047, see Table 2). Twenty-four-hours post-treatment with formic acid, they significantly fell more than the control group, whatever the tested concentration (Figure 2, Binomial GLM p-value for all < 0.01, see Table A1 and Table 2). For grip duration, one hour and a half post-treatment, the median duration was not significantly different from the control group. However, the median duration is significantly shorter at twenty-four-hours when V. destructor females were exposed to 5 or 25 mg/mL of formic acid but not 1 mg/mL (Figure 3, Kruskal-Wallis χ2 = 38.353, df = 3, p-value < 0.001, see Table 2).
Lactic acid—In the case of lactic acid, mites fell significantly more one hour and a half post-treatment than the control group, regardless of the concentration used (Figure 2, Fisher exact test with Bonferroni correction p-value for all < 0.001, see Table 2). Twenty-four hours post-treatment with lactic acid (150 mg/mL), 90% [CI95: 73.4–97.8] of V. destructor fell from the Rotavar and 93.3% [CI95: 77.9–99.1] at 600 mg/mL (Figure 2, Binomial GLM, p-value for 150 mg/mL < 0.0001, see Table A1 and Table 2). One hour and a half post-treatment, grip duration was significantly lower while mites were treated with lactic acid (150 mg/mL), compared to the control (Figure 3, Kruskal-Wallis χ2 = 45.404, df = 3, p-value < 0.001, see Table 2). Twenty-four hours post-exposure, mites treated with lactic acid showed a shorter median duration of 46 s at 150 mg/mL compared to 180 s for the control group. It was also the case for mites treated with 25 or 600 mg/mL of lactic acid (Figure 3, Kruskal-Wallis χ2 = 64.744, df = 3, p-value < 0.001, see Table 2).
Tartaric acid—Varroa destructor females exposed to 150 mg/mL of tartaric acid fell significantly more than mites from the control group one hour and a half post-treatment (Figure 2, Fisher exact test p-value = 0.04, see Table 2). Twenty-four hours post-treatment, 75.8% [CI95: 56.4–89.7] of V. destructor fell from the Rotavar at 150 mg/mL and 70.3% [CI95: 49.8–86.2] at 600 mg/mL (Figure 2, Binomial GLM p-value for 150 mg/mL < 0.0001, see Table A1 and Table 2). For grip duration, one hour and a half post-treatment, the median duration was not significantly different from the control group. Lastly, median duration was significantly shorter when V. destructor females were exposed to 150 or 600 mg/mL of tartaric acid twenty-four-hours post-treatment (Figure 3, Kruskal-Wallis χ2 = 37.782, df = 4, p-value < 0.001, see Table 2). However, no difference was found in the grip duration of treated mites with 25 mg/mL of tartaric acid.
Survival—The highest concentrations of lactic acid induced mortality after twenty-four hours (Figure 4, Kaplan-Meier: p-value 600 mg/mL < 0.0001 and p-value 150 mg/mL = 0.0006). Tartaric acid at 150 and 600 mg/mL also led mites to die (Figure 4, Kaplan-Meier: p-value 150 mg/mL = 0.002 and p-value 600 mg/mL < 0.0001). Formic acid, only 25 mg/mL provoked death in mites at 48 h (Figure 4, Kaplan-Meier: p-value < 0.0001). Citric acid was the only one with no significant death during forty-eight hours, regardless of the concentration (Figure 4, Kaplan-Meier: p-value for all > 0.05).
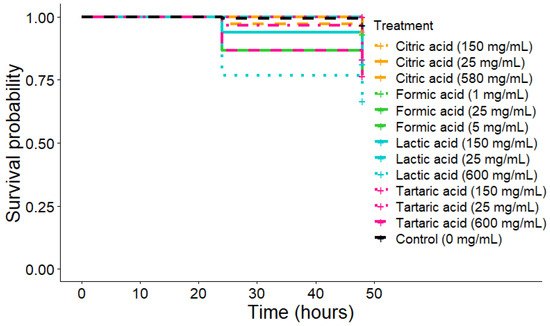
Figure 4.
Survival curves over 48 h for V. destructor after organic acid treatment (walking on impregnated paper).
3.2. Effect of Organic Acids on V. destructor Fall off Honeybees
Honeybees carrying mites received topical treatment on the back and were gathered in a micro-colony of five bees. Seventy-two hours post exposition, the proportion of fallen mites from the control group was 4.74% [CI95: 0.58–18.30], which was expected as mites do not fall naturally from their hosts. They were represented with a dashed line and gray area for CI95 (Figure 5).
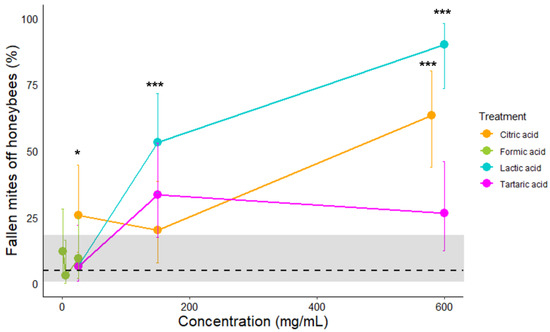
Figure 5.
Effects of citric, formic, lactic, and tartaric acid concentrations on V. destructor female removal from honeybees cumulated over seventy-two-hours post-treatment. Dashed line represents the control group. The grey area around dashed line represents the confidence interval of 95%. Color bars represent confidence interval of 95%; *** stands for p < 0.001, and * for p < 0.05 compared to the control (see Table 3 for detailed statistics).
The proportion of fallen mites from the bees was significantly higher for citric acid at 580 mg/mL than the control group with 63.3% [CI95: 43.8–80] (Figure 5, Binomial GLM p-value = 0.0003, see Table A2 and Table 3). Caution must be taken with citric acid at 25 mg/mL. Indeed, it was significantly higher than the control group. However, their CI95s were overlapping. No difference was detected between 25 and 150 mg/mL for citric acid or any concentrations of formic and tartaric acids in the control group. The proportion of fallen mites from the bees was significantly higher, with 53.3% [CI95: 34.3–71.6] for lactic acid at 150 mg/mL and 90% [CI95: 73.4–97.8] at 600 mg/mL, seventy-two hours post-treatment (Figure 5, Binomial GLM p-value for 150 mg/mL = 0.0005, see Table A2 and Table 3).
Table 3.
Outcome of statistical analyses for percentages of fallen mites off honeybees cumulated over seventy-two-hours post-treatment—Statistical analyses with adjusted p-value using Fisher exact test with Bonferroni correction (vs. = versus).
The highest number of fallen mites occurred during the first twenty-four hours of lactic acid. The highest concentration of lactic acid was toxic for bees, inducing high mortality after forty-eight hours (Figure 6; Kaplan-Meier: p-value = 0.022), as well as tartaric acid 600 mg/mL (Kaplan-Meier: p-value = 0.036) and citric acid 25 mg/mL (Kaplan-Meier: p-value < 0.0001). Therefore, none of the tested concentrations were considered toxic for bees except 600 mg/mL of lactic and tartaric acids as well as 25 mg/mL of citric acid.
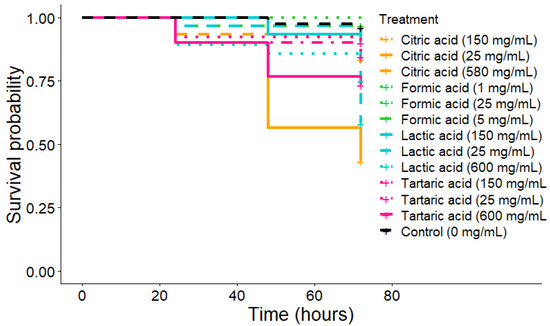
Figure 6.
Survival curves over 72 h for honeybees after organic acid treatment (drops on bees’ backs).
4. Discussion
This study is the first attempt to compare the effects of citric, formic, lactic, and tartaric acids on V. destructor host attachment abilities. The miticide impact of formic and lactic acids was demonstrated in anterior field studies [15,31,44]; however, little is known about their effects on adherence abilities. We investigated mites’ grip skills when their arolia were directly exposed to different organic acids.
We chose concentrations of acids according to their common applications in the field. Indeed, formic acid is used in commercial products such as Varromed® at 5 mg/mL for trickling [32]. Colonies are treated in Switzerland with lactic acid at 150 mg/mL by spraying [31,45]. In our conditions, twenty-four-hours post-treatment, we observed that citric, formic, lactic, and tartaric acids impaired the attachment abilities of mites. From a chemical perspective, we could suggest that the acidity of the tested molecules explained the results, regardless of the specific properties of each acid. However, in our preliminary experiment, we tested acetic acid (Figure A1), which did not significantly impact grip skill, preventing the idea that the acidity (pH) of the molecules was sufficient to disrupt attachment abilities in the conditions tested. In addition, it is known that citric, formic, lactic, and tartaric acids have different molar sizes; thus, the quantities of organic acids compared are very different from one another. Yet, even in molar concentration, the difference observed in the rate of fallen mites between acids was not adjusted.
Several hypotheses arise from the fall of Varroa females after topical treatment of bees with acids. First, organic acids could interfere with the nervous system of mites and paralyse them, inducing the fall [46,47]. Although it is unlikely for lactic acid, according to our previous findings [36] and the survival curve associated with it (Figure 4). Second, a toxic effect may lead to their fall [18]. It could be the case for tartaric acid, as it reduced V. destructor lifespan; however, it seems not to be true for lactic or citric acid. Third, females’ falls could be the result of an interaction between acids and the fluid under their arolia as well as a burn of their arolia, impairing their attachment abilities [36].
Many arachnids and insects leave a footprint under their arolia [48,49,50,51]. Among them, the nature of the fluid may vary from proteins and polypeptides to polysaccharides or lipids. For ticks, the nature of the fluid seemed to be a biphasic oil-water emulsion with solid particles [24,29,52]. We do not know the nature of V. destructor footprint fluid; however, we can hypothesise a similar nature to Ixodides due to their close phylogeny [29]. It is therefore interesting to compare grip abilities between a hydrophilic substrate, here the wood, and a hydrophobic one, in our case the cuticle of the honeybee. We found out that for citric acid, mites fell significantly more from honeybees than from wood (when considering 580 mg/mL). On the other hand, tartaric acid at 150 and 600 mg/mL induced a significant fall in wood but not in honeybees. It is even more interesting when considering formic acid. This volatile acid is known for dysregulating cellular respiration in mites, leading to their death [53]. Here we described impaired grip skill on wood but not on honeybees, suggesting that adherence is not challenged in hive treatment. The physical and chemical properties of each acid are crucial when investigating their effects on mite attachment to a host. As a matter of fact, citric and lactic acids are known as biodemulsifiers [54,55,56], whereas tartaric acid is a well-known emulsifier [57,58]. Accordingly, the nature of the fluid cannot be overlooked, as it may be destabilised through the interaction with acids, becoming slippery or sticky [59,60].
Altogether, we showed that lactic acid was the only candidate impairing mite grip skill both on wood and on bees at a reasonable concentration (150 mg/mL) and able to dislodge V. destructor without killing bees seventy-two hours post-treatment (Figure 5). During the experiment, for the lactic acid condition, we noticed a detached V. destructor on the floor, stuck on the back, or unable to walk normally in between daily checkpoints. Later, we would find them dead, presumably from starvation and exhaustion (Figure 4). This was consistent with our previous experiments showing attachment deficits on wood. However, indirect exposure is always challenging [61,62]. The mite indeed had to move on the bee to get in contact with the molecule. Several hypotheses could lead to such contact. First, mites could be dislodged by the grooming behaviour of bees and come into contact with lactic acid. Regardless of the condition, we observed two grooming behaviours in honeybees: auto-grooming and allo-grooming [63]. Second, lactic acid could attract or repel mites, inducing a movement of Varroa females from their initial feeding site, thus touching the acid. Third, mites could naturally move without being attracted or repulsed and randomly enter in contact with the lactic acid coating on honeybees’ backs. Yet, few V. destructors fell off honeybees. All of these hypotheses seem consistent with the lower percentage of fallen mites where arolia are the contact point.
5. Conclusions
To conclude, this study provides a new perspective on the mode of action of organic acids on V. destructor as a control strategy. Among tartaric, citric, formic, and lactic acids, we identified potential candidates able to interfere with host attachment under laboratory conditions. In particular, for lactic acid already in use in the field, we showed that through an application of the acid on bees’ backs or when V. destructor females walked directly on impregnated paper, we obtained the same impaired grip skill. It may indicate that V. destructor collected at the bottom of treated hives several weeks after the application died from their fall and not from toxicity [31]. Further investigations would help to get a deeper understanding of the level of bees’ behaviour and its implications for the fall of mites after lactic acid administration. For example, we could quantify honeybee auto- and allo-grooming to determine if the action and contact of the acid on the mite are dependent on bees’ behaviour. Lastly, attachment skills for parasites are crucial to complete their lifecycle, so they represent an alternative target for mite control strategies [64]. This work supports evidence for the use of a reduced amount of organic acids in the hive, as it is not necessary to kill mites with a high quantity of acids but to adequately make them fall. Our results can help in the development of new methods for in-hive applications for beekeepers worldwide.
6. Patents
Results from this article may be part of a patent.
Author Contributions
Experiments and analysis: C.V.; writing—original draft preparation, C.V.; writing—review and editing, C.V., V.P., S.B., F.V. and A.V.; supervision, A.V.; funding acquisition, F.V. and A.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by M2i Biocontrol and the Région Occitanie (ADEME grant No. 2082C0061).
Institutional Review Board Statement
Invertebrate welfare was considered in all of our studies.
Informed Consent Statement
Not applicable.
Data Availability Statement
Datasets generated and analysed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Armengaud Catherine for her support of the idea of the Rotavar. We also thank Yannick Escudie from M2I development for the acid solutions. We thank Olivier Guerret from M2I Biocontrol for his support in the conception of the experiments.
Conflicts of Interest
The authors declare no conflict of interest. M2I Biocontrol did not influence the analysis and publication.
Appendix A
Table A1.
Grip on wood (Rotavar)—Estimate parameters from the binomial GLM for the proportion of fallen mites treated compared to control. Significant p-values are shown in bold. SE: standard errors.
Table A1.
Grip on wood (Rotavar)—Estimate parameters from the binomial GLM for the proportion of fallen mites treated compared to control. Significant p-values are shown in bold. SE: standard errors.
| Parameters | Estimate | SE | p-Value |
|---|---|---|---|
| Lactic acid—24 h | |||
| Intercept | 3.367 | 1.017 | 0.0009 |
| 25 mg/mL | 3.367 | 1.083 | 0.0004 |
| 150 mg/mL | 5.565 | 1.185 | <0.0001 |
| 600 mg/mL | 6.006 | 1.253 | <0.0001 |
| Formic acid—24 h | |||
| Intercept | −3.367 | 1.017 | 0.0009 |
| 1 mg/mL | 2.962 | 1.083 | 0.0062 |
| 5 mg/mL | 4.557 | 1.105 | <0.0001 |
| 25 mg/mL | 4.178 | 1.102 | 0.0001 |
| Tartaric acid—24 h | |||
| Intercept | −1.8718 | 0.5371 | 0.0004 |
| 25 mg/mL | 0.8602 | 0.6774 | 0.2041 |
| 150 mg/mL | 3.0169 | 0.6905 | <0.0001 |
| 600 mg/mL | 2.7368 | 0.6827 | <0.0001 |
| Citric acid—1 h 30 | |||
| Intercept | −2.7408 | 0.7296 | 0.0001 |
| 25 mg/mL | 1.5513 | 0.8477 | 0.0672 |
| 150 mg/mL | 1.1314 | 0.8788 | 0.1979 |
| 580 mg/mL | 2.4532 | 0.8056 | 0.0023 |
| Citric acid—24 h | |||
| Intercept | −3.434 | 1.016 | 0.0007 |
| 25 mg/mL | 3.029 | 1.082 | 0.0051 |
| 150 mg/mL | 2.741 | 1.087 | 0.0117 |
| 580 mg/mL | 2.828 | 1.078 | 0.0086 |
Table A2.
Grip on bees—Estimate parameters from the binomial GLM for the proportion of fallen mites treated compared to control. Significant p-values are shown in bold. SE: standard errors.
Table A2.
Grip on bees—Estimate parameters from the binomial GLM for the proportion of fallen mites treated compared to control. Significant p-values are shown in bold. SE: standard errors.
| Parameters | Estimate | SE | p-Value |
|---|---|---|---|
| Lactic acid | |||
| Intercept | −3.5835 | 1.0138 | 0.0004 |
| 25 mg/mL | 0.9445 | 1.2504 | 0.45 |
| 150 mg/mL | 3.7171 | 1.0778 | 0.0005 |
| 600 mg/mL | 5.7807 | 1.1824 | <0.0001 |
| Formic acid | |||
| Intercept | −3.4965 | 1.015 | 0.0005 |
| 1 mg/mL | 1.5155 | 1.1466 | 0.1862 |
| 5 mg/mL | 0.0625 | 1.4361 | 0.9652 |
| 25 mg/mL | 1.2278 | 1.1824 | 0.299 |
| Tartaric acid | |||
| Intercept | −2.1972 | 0.6086 | 0.0003 |
| 25 mg/mL | −0.4418 | 0.9519 | 0.6425 |
| 150 mg/mL | 1.5041 | 0.7214 | 0.037 |
| 600 mg/mL | 1.1856 | 0.7354 | 0.1069 |
| Citric acid | |||
| Intercept | −3.367 | 1.017 | 0.0009 |
| 25 mg/mL | 2.311 | 1.097 | 0.035 |
| 150 mg/mL | 1.981 | 1.115 | 0.0755 |
| 580 mg/mL | 3.914 | 1.085 | 0.0003 |
Appendix B

Figure A1.
Effects of acetic acid concentrations on V. destructor females’ grip skill via the Rotavar test after one and a half hours as well as twenty-four-hours post-treatment. Dashed line represents the control group. The grey area around the dashed line represents the confidence interval of 95%. Colour bars represent confidence interval of 95%.
References
- Brodschneider, R.; Schlagbauer, J.; Arakelyan, I.; Ballis, A.; Brus, J.; Brusbardis, V.; Cadahía, L.; Charrière, J.-D.; Chlebo, R.; Coffey, M.F.; et al. Spatial Clusters of Varroa destructor Control Strategies in Europe. J. Pest Sci. 2023, 96, 759–783. [Google Scholar] [CrossRef]
- Gisder, S.; Aumeier, P.; Genersch, E. Deformed Wing Virus: Replication and Viral Load in Mites (Varroa destructor). J. Gen. Virol. 2009, 90, 463–467. [Google Scholar] [CrossRef]
- Wilfert, L.; Long, G.; Leggett, H.C.; Schmid-Hempel, P.; Butlin, R.; Martin, S.J.M.; Boots, M. Deformed Wing Virus Is a Recent Global Epidemic in Honeybees Driven by Varroa Mites. Science 2016, 351, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Brettell, L.E. Deformed Wing Virus in Honeybees and Other Insects. Annu. Rev. Virol. 2019, 6, 49–69. [Google Scholar] [CrossRef] [PubMed]
- Boecking, O.; Genersch, E. Varroosis—The Ongoing Crisis in Bee Keeping. J. Verbr. Lebensm. 2008, 3, 221–228. [Google Scholar] [CrossRef]
- Dainat, B.; Evans, J.D.; Chen, Y.P.; Gauthier, L.; Neumann, P. Dead or Alive: Deformed Wing Virus and Varroa Destructor Reduce the Life Span of Winter Honeybees. Appl. Environ. Microbiol. 2012, 78, 981–987. [Google Scholar] [CrossRef]
- Ramsey, S.D.; Ochoa, R.; Bauchan, G.; Gulbronson, C.; Mowery, J.D.; Cohen, A.; Lim, D.; Joklik, J.; Cicero, J.M.; Ellis, J.D.; et al. Varroa destructor Feeds Primarily on Honey Bee Fat Body Tissue and Not Hemolymph. Proc. Natl. Acad. Sci. USA 2019, 116, 1792–1801. [Google Scholar] [CrossRef]
- Piou, V.; Vilarem, C.; Blanchard, S.; Strub, J.; Bertile, F.; Bocquet, M.; Arafah, K.; Bulet, P.; Vétillard, A. Honey Bee Larval Hemolymph as a Source of Key Nutrients and Proteins Offers a Promising Medium for Varroa destructor Artificial Rearing. Int. J. Mol. Sci. 2023, 24, 12443. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and Control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef]
- Vilarem, C.; Piou, V.; Vogelweith, F.; Vétillard, A. Varroa destructor from the Laboratory to the Field: Control, Biocontrol and IPM Perspectives—A Review. Insects 2021, 12, 800. [Google Scholar] [CrossRef]
- Sanchez-Bayo, F.; Goka, K. Pesticide Residues and Bees—A Risk Assessment. PLoS ONE 2014, 9, e94482. [Google Scholar] [CrossRef]
- Rinkevich, F.D. Detection of Amitraz Resistance and Reduced Treatment Efficacy in the Varroa Mite, Varroa destructor, within Commercial Beekeeping Operations. PLoS ONE 2020, 15, e0227264. [Google Scholar] [CrossRef] [PubMed]
- Maggi, M.; Tourn, E.; Negri, P.; Szawarski, N.; Marconi, A.; Gallez, L.; Medici, S.; Ruffinengo, S.; Brasesco, C.; De Feudis, L.; et al. A New Formulation of Oxalic Acid for Varroa destructor Control Applied in Apis mellifera Colonies in the Presence of Brood. Apidologie 2016, 47, 596–605. [Google Scholar] [CrossRef]
- Steube, X.; Beinert, P.; Kirchner, W.H. Efficacy and Temperature Dependence of 60% and 85% Formic Acid Treatment against Varroa destructor. Apidologie 2021, 52, 720–729. [Google Scholar] [CrossRef]
- Căuia, E.; Căuia, D. Improving the Varroa (Varroa destructor) Control Strategy by Brood Treatment with Formic Acid—A Pilot Study on Spring Applications. Insects 2022, 13, 149. [Google Scholar] [CrossRef] [PubMed]
- Nozal, M.J.; Bernal, J.L.; Gomez, L.A.; Higes, M.; Meana, A. Determination of Oxalic Acid and Other Organic Acids in Honey and in Some Anatomic Structures of Bees. Apidologie 2003, 34, 181–188. [Google Scholar] [CrossRef]
- Mato, I.; Huidobro, J.F.; Simal-Lozano, J.; Sancho, M.T. Analytical Methods for the Determination of Organic Acids in Honey. Crit. Rev. Anal. Chem. 2006, 36, 3–11. [Google Scholar] [CrossRef]
- Gregorc, A.; Planinc, I. Acaricidal Effect of Oxalic Acid in Honeybee (Apis mellifera) Colonies. Apidologie 2001, 32, 333–340. [Google Scholar] [CrossRef][Green Version]
- Charriére, J.-D.; Imdorf, A. Oxalic Acid Treatment by Trickling against Varroa Destructor: Recommendations for Use in Central Europe and under Temperate Climate Conditions. Bee World 2002, 83, 51–60. [Google Scholar] [CrossRef]
- Jack, C.J.; van Santen, E.; Ellis, J.D. Evaluating the Efficacy of Oxalic Acid Vaporization and Brood Interruption in Controlling the Honey Bee Pest Varroa destructor (Acari: Varroidae). J. Econ. Entomol. 2020, 113, 582–588. [Google Scholar] [CrossRef]
- Papežíková, I.; Palíková, M.; Kremserová, S.; Zachová, A.; Peterová, H.; Babák, V.; Navrátil, S. Effect of Oxalic Acid on the Mite Varroa destructor and Its Host the Honey Bee Apis mellifera. J. Apic. Res. 2017, 56, 400–408. [Google Scholar] [CrossRef]
- Evans, J.D.; Cook, S.C. Genetics and Physiology of Varroa Mites. Curr. Opin. Insect Sci. 2018, 26, 130–135. [Google Scholar] [CrossRef]
- Schimpf, D.J.; Ewert, M.M.; Lai, V.K.; Clarke, B.L. Responses of Ticks to Immersion in Hot Bathing Water: Effect of Surface Type, Water Temperature, and Soap on Tick Motor Control. PLoS ONE 2021, 16, e0261592. [Google Scholar] [CrossRef] [PubMed]
- Voigt, D.; Gorb, S. Functional Morphology of Tarsal Adhesive Pads and Attachment Ability in Ticks Ixodes ricinus (Arachnida, Acari, Ixodidae). J. Exp. Biol. 2017, 220, 1984–1996. [Google Scholar] [CrossRef]
- van den Boogaart, L.M.; Langowski, J.K.A.; Amador, G.J. Studying Stickiness: Methods, Trade-Offs, and Perspectives in Measuring Reversible Biological Adhesion and Friction. Biomimetics 2022, 7, 134. [Google Scholar] [CrossRef] [PubMed]
- Dietemann, V.; Pflugfelder, J.; Anderson, D.; Charrière, J.; Chejanovsky, N.; Dainat, B.; de Miranda, J.; Delaplane, K.; Dillier, F.; Fuch, S.; et al. Varroa destructor: Research Avenues towards Sustainable Control. J. Apic. Res. 2012, 51, 125–132. [Google Scholar] [CrossRef]
- Dhooria, M.S. Morphology and Anatomy of Acari. In Fundamentals of Applied Acarology; Springer: Singapore, 2016; pp. 41–61. ISBN 978-981-10-1592-2. [Google Scholar]
- Plettner, E.; Eliash, N.; Singh, N.K.; Pinnelli, G.R.; Soroker, V. The Chemical Ecology of Host-Parasite Interaction as a Target of Varroa destructor Control Agents. Apidologie 2017, 48, 78–92. [Google Scholar] [CrossRef]
- Wolff, J.O.; Gorb, S.N. Attachment Structures and Adhesive Secretions in Arachnids; Biologically-Inspired Systems; Springer International Publishing: Cham, Switzerland, 2016; Volume 7, ISBN 978-3-319-45712-3. [Google Scholar]
- Mitton, G.A.; Meroi Arcerito, F.; Cooley, H.; Fernández de Landa, G.; Eguaras, M.J.; Ruffinengo, S.R.; Maggi, M.D. More than Sixty Years Living with Varroa destructor: A Review of Acaricide Resistance. Int. J. Pest Manag. 2022, 68, 1–18. [Google Scholar] [CrossRef]
- Kraus, B.; Berg, S. Effect of a Lactic Acid Treatment during Winter in Temperate Climate upon Varroa jacobsoni Oud. and the Bee (Apis mellifera L.) Colony. Exp. Appl. Acarol. 1994, 18, 459–468. [Google Scholar] [CrossRef]
- Smodiš Škerl, M.I.; Rivera-Gomis, J.; Tlak Gajger, I.; Bubnič, J.; Talakić, G.; Formato, G.; Baggio, A.; Mutinelli, F.; Tollenaers, W.; Laget, D.; et al. Efficacy and Toxicity of VarroMed® Used for Controlling Varroa Destructor Infestation in Different Seasons and Geographical Areas. Appl. Sci. 2021, 11, 8564. [Google Scholar] [CrossRef]
- Milani, N. Activity of Oxalic and Citric Acids on the Mite Varroa destructor in Laboratory Assays. Apidologie 2001, 32, 127–138. [Google Scholar] [CrossRef]
- Dietemann, V.; Nazzi, F.; Martin, S.J.; Anderson, D.L.; Locke, B.; Delaplane, K.S.; Wauquiez, Q.; Tannahill, C.; Frey, E.; Ziegelmann, B.; et al. Standard Methods for Varroa Research. J. Apic. Res. 2013, 52, 1–54. [Google Scholar] [CrossRef]
- Piou, V.; Tabart, J.; Urrutia, V.; Hemptinne, J.; Vétillard, A. Impact of the Phoretic Phase on Reproduction and Damage Caused by Varroa destructor (Anderson and Trueman) to Its Host, the European Honey Bee (Apis mellifera L.). PLoS ONE 2016, 11, e0153482. [Google Scholar] [CrossRef]
- Vilarem, C.; Piou, V.; Blanchard, S.; Armengaud, C.; Vogelweith, F.; Vétillard, A. Lactic Acid Impairs Varroa destructor Grip Skill: Effects on Its Biology and Behaviour under Artificial Conditions. 2023; under review. [Google Scholar]
- Dunham, N.W.; Miya, T.S. A Note on a Simple Apparatus for Detecting Neurological Deficit in Rats and Mice** College of Pharmacy, University of Nebraska, Lincoln 8. J. Am. Pharm. Assoc. 1957, 46, 208–209. [Google Scholar] [CrossRef] [PubMed]
- Gashout, H.A.; Guzmán-Novoa, E. Acute Toxicity of Essential Oils and Other Natural Compounds to the Parasitic Mite, Varroa destructor, and to Larval and Adult Worker Honey Bees (Apis mellifera L.). J. Apic. Res. 2009, 48, 263–269. [Google Scholar] [CrossRef]
- Evans, J.; Chen, Y.; di Prisco, G.; Pettis, J.; Williams, V. Bee Cups: Single-Use Cages for Honey Bee Experiments. J. Apic. Res. 2009, 48, 300–302. [Google Scholar] [CrossRef]
- Huang, S.K.; Csaki, T.; Doublet, V.; Dussaubat, C.; Evans, J.D.; Gajda, A.M.; Gregorc, A.; Hamilton, M.C.; Kamler, M.; Lecocq, A.; et al. Evaluation of Cage Designs and Feeding Regimes for Honey Bee (Hymenoptera: Apidae) Laboratory Experiments. J. Econ. Entom. 2014, 107, 54–62. [Google Scholar] [CrossRef]
- Hervé, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. 2020. Available online: https://rdrr.io/cran/RVAideMemoire/ (accessed on 5 July 2023).
- Alboukadel, K.; Marcin, K.; Przemyslaw, B.; Scheipl, F. Survminer: Drawing Survival Curves Using “Ggplot2”. 2022. Available online: https://rdrr.io/cran/survminer/ (accessed on 5 July 2023).
- Therneau, T. Survival: Survival Analysis 2023; Agroscope: Bern, Switzerland, 2004. [Google Scholar]
- Lupo, A.; Gerling, D. A Comparison between the Efficiency of Summer Treatments Using Formic Acid and Taktic® against Varroa jacobsoni in Beehives. Apidologie 1990, 21, 261–267. [Google Scholar] [CrossRef]
- Charrière, J.; Imdorf, A.; Kuhn, R. Bienenverträglichkeit von Varroabehandlungen Im Winter. Schweiz. Bienen-Ztg. 2004, 14, 19–23. [Google Scholar]
- Evans, P.D.; Gee, J.D. Action of Formamidine Pesticides on Octopamine Receptors. Nature 1980, 287, 60–62. [Google Scholar] [CrossRef]
- Blenau, W.; Rademacher, E.; Baumann, A. Plant Essential Oils and Formamidines as Insecticides/Acaricides: What Are the Molecular Targets? Apidologie 2012, 43, 334–347. [Google Scholar] [CrossRef]
- Federle, W. An Integrative Study of Insect Adhesion: Mechanics and Wet Adhesion of Pretarsal Pads in Ants. Integr. Comp. Biol. 2002, 42, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Gladun, D.; Gorb, S.N.; Frantsevich, L.I. Alternative Tasks of the Insect Arolium with Special Reference to Hymenoptera. In Functional Surfaces in Biology; Springer: Dordrecht, The Netherlands, 2009; pp. 67–103. ISBN 978-1-4020-6694-8. [Google Scholar]
- Peattie, A.M.; Dirks, J.; Henriques, S.; Federle, W. Arachnids Secrete a Fluid over Their Adhesive Pads. PLoS ONE 2011, 6, e20485. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wolff, J.O.; Huber, S.J.; Gorb, S.N. How to Stay on Mummy’s Back: Morphological and Functional Changes of the Pretarsus in Arachnid Postembryonic Stages. Arthropod Struct. Dev. 2015, 44, 301–312. [Google Scholar] [CrossRef]
- Vötsch, W.; Nicholson, G.; Müller, R.; Stierhof, Y.-D.; Gorb, S.; Schwarz, U. Chemical Composition of the Attachment Pad Secretion of the Locust Locusta Migratoria. Insect Biochem. Mol. Biol. 2002, 32, 1605–1613. [Google Scholar] [CrossRef]
- Genath, A.; Petruschke, H.; von Bergen, M.; Einspanier, R. Influence of Formic Acid Treatment on the Proteome of the Ectoparasite Varroa destructor. PLoS ONE 2021, 16, e0258845. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Suo, Y.; Zhao, J.; Zhu, P.; Tan, J.; Wang, B.; Lu, H. Effect of Demulsification for Crude Oil-in-Water Emulsion: Comparing CO2 and Organic Acids. Energy Fuels 2018, 32, 757–764. [Google Scholar] [CrossRef]
- Zolfaghari, R.; Fakhru’l-Razi, A.; Abdullah, L.C.; Elnashaie, S.S.E.H.; Pendashteh, A. Demulsification Techniques of Water-in-Oil and Oil-in-Water Emulsions in Petroleum Industry. Sep. Purif. Technol. 2016, 170, 377–407. [Google Scholar] [CrossRef]
- Yao, L.; Selmi, A.; Esmaeili, H. A Review Study on New Aspects of Biodemulsifiers: Production, Features and Their Application in Wastewater Treatment. Chemosphere 2021, 284, 131364. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ge, L.; Friberg, S.E.; Guo, R. Equilibration in a Tartaric Acid Emulsion System. Colloids Surf. A Physicochem. Eng. Asp. 2010, 358, 135–141. [Google Scholar] [CrossRef]
- Yesiltas, B.; García-Moreno, P.J.; Sørensen, A.D.M.; Anankanbil, S.; Guo, Z.; Jacobsen, C. Effects of Modified DATEMs with Different Alkyl Chain Lengths on Improving Oxidative and Physical Stability of 70% Fish Oil-in-Water Emulsions. J. Agric. Food Chem. 2018, 66, 12512–12520. [Google Scholar] [CrossRef]
- Bohn, H.F.; Federle, W. Insect Aquaplaning: Nepenthes Pitcher Plants Capture Prey with the Peristome, a Fully Wettable Water-Lubricated Anisotropic Surface. Proc. Natl. Acad. Sci. USA 2004, 101, 14138–14143. [Google Scholar] [CrossRef]
- Dirks, J.; Clemente, C.J.; Federle, W. Insect Tricks: Two-Phasic Foot Pad Secretion Prevents Slipping. J. R. Soc. Interface. 2010, 7, 587–593. [Google Scholar] [CrossRef]
- Aliano, N.P.; Ellis, M.D. Bee-to-Bee Contact Drives Oxalic Acid Distribution in Honey Bee Colonies. Apidologie 2008, 39, 481–487. [Google Scholar] [CrossRef]
- Rademacher, E.; Harz, M.; Schneider, S. Effects of Oxalic Acid on Apis mellifera (Hymenoptera: Apidae). Insects 2017, 8, 84. [Google Scholar] [CrossRef]
- Pritchard, D.J. Grooming by Honey Bees as a Component of Varroa Resistant Behavior. J. Apic. Res. 2016, 55, 38–48. [Google Scholar] [CrossRef]
- Büscher, T.H.; Petersen, D.S.; Bijma, N.N.; Bäumler, F.; Pirk, C.W.W.; Büsse, S.; Heepe, L.; Gorb, S.N. The Exceptional Attachment Ability of the Ectoparasitic Bee Louse Braula coeca (Diptera, Braulidae) on the Honeybee. Physiol. Entomol. 2022, 47, 83–95. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).