
Feedforward Loops: Evolutionary Conserved Network Motifs Redesigned for Synthetic Biology Applications




Abstract
1. Introduction
2. FFLs: A General Description
2.1. Structure and Working of FFLs
2.2. FFL Is Evolutionary Conserved
2.3. FFL Abundance
2.4. Logic Behavior
3. Coherent FFLs
3.1. C1-FFL: A Sign-Sensitive Delay Device
3.1.1. AND Logic C1-FFL
3.1.2. OR Logic C1-FFL
3.1.3. Filtering Out Spurious Signals
3.2. Coherent FFLs in Eukaryotic Cells
4. Incoherent FFLs
4.1. I1-FFL as a Pulse Generator and a Response-Time Accelerator
4.2. Synthetic I1-FFLs in E. coli
4.3. Incoherent FFLs in Eukaryotes
4.4. Incoherent FFLs in Cell-Free System
4.5. Alternative Designs for Incoherent FFLs
5. Interlocked FFLs
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Endy, D. Foundations for engineering biology. Nature 2005, 438, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Costello, A.; Badran, A.H. Synthetic Biological Circuits within an Orthogonal Central Dogma. Trends Biotechnol. 2021, 39, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Müller-Hill, B. The Lac Operon: A Short History of a Genetic Paradigm; De Gruyter: Berlin, Germany, 1996. [Google Scholar]
- Balleza, E.; López-Bojorquez, L.N.; Martínez-Antonio, A.; Resendis-Antonio, O.; Lozada-Chávez, I.; Balderas-Martínez, Y.I.; Encarnación, S.; Collado-Vides, J. Regulation by transcription factors in bacteria: Beyond description. FEMS Microbiol. Rev. 2008, 33, 133–151. [Google Scholar] [CrossRef]
- Okano, H.; Hermsen, R.; Kochanowski, K.; Hwa, T. Regulation underlying hierarchical and simultaneous utilization of carbon substrates by flux sensors in Escherichia coli. Nat. Microbiol. 2020, 5, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Alon, U. An Introduction to Systems Biology: Design Principles of Biological Circuits, 2nd ed.; CRC: Boca Raton, FL, USA, 2020. [Google Scholar]
- Erdős, P.; Rényi, A. On the evolution of random graphs. Publ. Math. Inst. Hung. Acad. Sci. 1960, 5, 17–61. [Google Scholar]
- Marchisio, M.A. Polynomial Observables in the Graph Partitioning Problem. Int. J. Mod. Phys. C 2001, 12, 13–18. [Google Scholar] [CrossRef]
- Alon, U. Network motifs: Theory and experimental approaches. Nat. Rev. Genet. 2007, 8, 450–461. [Google Scholar] [CrossRef]
- Adler, M.; Alon, U. Fold-change detection in biological systems. Curr. Opin. Syst. Biol. 2018, 8, 81–89. [Google Scholar] [CrossRef]
- Song, S.; Sjöström, P.J.; Reigl, M.P.; Nelson, S.M.; Chklovskii, D.B.S. Highly Nonrandom Features of Synaptic Connectivity in Local Cortical Circuits. PLoS Biol. 2005, 3, e68. [Google Scholar] [CrossRef]
- Levine, M.; Davidson, E.H. Gene Regulatory Networks for Development. Proc. Natl. Acad. Sci. USA 2005, 102, 4936–4942. [Google Scholar] [CrossRef]
- Shin, Y.-J.; Nourani, M. Statecharts for Gene Network Modeling. PLoS ONE 2010, 5, e9376. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.T.; Sano, M. Regulatory Dynamics of Synthetic Gene Networks with Positive Feedback. J. Mol. Biol. 2006, 359, 1107–1124. [Google Scholar] [CrossRef] [PubMed]
- Milo, R.; Itzkovitz, S.R.; Kashtan, N.S.; Levitt, R.N.; Alon, U.R. Response to Comment on “Network Motifs: Simple Building Blocks of Complex Networks” and “Superfamilies of Evolved and Designed Networks”. Science 2004, 305, 1107. [Google Scholar] [CrossRef]
- Shen-Orr, S.S.; Milo, R.; Mangan, S.R.; Alon, U. Network motifs in the transcriptional regulation network of Escherichia coli. Nat. Genet. 2002, 31, 64–68. [Google Scholar] [CrossRef]
- Mangan, S.; Alon, U. Structure and function of the feed-forward loop network motif. Proc. Natl. Acad. Sci. USA 2003, 100, 11980–11985. [Google Scholar] [CrossRef]
- Milo, R.; Itzkovitz, S.; Kashtan, N.; Levitt, R.; Shen-Orr, S.; Ayzenshtat, I.; Sheffer, M.; Alon, U. Superfamilies of Evolved and Designed Networks. Science 2004, 303, 1538–1542. [Google Scholar] [CrossRef]
- Eichenberger, P.; Fujita, M.; Jensen, S.T.; Conlon, E.M.; Rudner, D.Z.; Wang, S.T.; Ferguson, C.; Haga, K.; Sato, T.; Liu, J.S.; et al. The Program of Gene Transcription for a Single Differentiating Cell Type during Sporulation in Bacillus subtilis. PLOS Biol. 2004, 2, e328. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.I.; Rinaldi, N.J.; Robert, F.; Odom, D.T.; Bar-Joseph, Z.; Gerber, G.K.; Hannett, N.M.; Harbison, C.T.; Thompson, C.M.; Simon, I.; et al. Transcriptional Regulatory Networks in Saccharomyces cerevisiae. Science 2002, 298, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Odom, D.T.; Zizlsperger, N.; Gordon, D.B.; Bell, G.W.; Rinaldi, N.J.; Murray, H.L.; Volkert, T.L.; Schreiber, J.; Rolfe, P.A.; Gifford, D.K.; et al. Control of Pancreas and Liver Gene Expression by HNF Transcription Factors. Science 2004, 303, 1378–1381. [Google Scholar] [CrossRef]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core Transcriptional Regulatory Circuitry in Human Embryonic Stem Cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef]
- Saddic, L.A.; Huvermann, B.; Bezhani, S.; Su, Y.; Winter, C.M.; Kwon, C.S.; Collum, R.P.; Wagner, D. The leafy target LMI1 is a meristem identity regulator and acts together with LEAFY to regulate expression of cauliflower. Development 2006, 133, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Swiers, G.; Patient, R.; Loose, M. Genetic regulatory networks programming hematopoietic stem cells and erythroid lineage specification. Dev. Biol. 2006, 294, 525–540. [Google Scholar] [CrossRef]
- Iranfar, N.; Fuller, D.; Loomis, W.F. Transcriptional regulation of post-aggregation genes in Dictyostelium by a feed-forward loop involving GBF and LagC. Dev. Biol. 2006, 290, 460–469. [Google Scholar] [CrossRef]
- Reeves, G.; Bandodkar, P.; Al Asafen, H. Spatiotemporal control of gene expression boundaries using a feedforward loop. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Goldbeter, A.; Koshland, D.E. Ultrasensitivity in biochemical systems controlled by covalent modification. Interplay between zero-order and multistep effects. J. Biol. Chem. 1984, 259, 14441–14447. [Google Scholar] [CrossRef]
- Gui, R.; Liu, Q.; Yao, Y.; Deng, H.; Ma, C.; Jia, Y.; Yi, M. Noise Decomposition Principle in a Coherent Feed-Forward Transcriptional Regulatory Loop. Front. Physiol. 2016, 7, 600. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Khetarpal, I.; Sen, S.; Murray, R. Synthetic circuit for exact adaptation and fold-change detection. Nucleic Acids Res. 2014, 42, 6078–6089. [Google Scholar] [CrossRef]
- Kuttykrishnan, S.; Sabina, J.; Langton, L.L.; Johnston, M.; Brent, M.R. A quantitative model of glucose signaling in yeast reveals an incoherent feed forward loop leading to a specific, transient pulse of transcription. Proc. Natl. Acad. Sci. USA 2010, 107, 16743–16748. [Google Scholar] [CrossRef]
- Mangan, S.; Itzkovitz, S.; Zaslaver, A.; Alon, U. The Incoherent Feed-forward Loop Accelerates the Response-time of the gal System of Escherichia coli. J. Mol. Biol. 2006, 356, 1073–1081. [Google Scholar] [CrossRef]
- Pérez-Morales, D.; Nava-Galeana, J.; Rosales-Reyes, R.; Teehan, P.; Yakhnin, H.; Melchy-Pérez, E.I.; Rosenstein, Y.; De la Cruz, M.A.; Babitzke, P.; Bustamante, V.H. An incoherent feedforward loop formed by SirA/BarA, HilE and HilD is involved in controlling the growth cost of virulence factor expression by Salmonella Typhimurium. PLOS Pathog. 2021, 17, e1009630. [Google Scholar] [CrossRef]
- Dekel, E.; Mangan, S.; Alon, U. Environmental selection of the feed-forward loop circuit in gene-regulation networks. Phys. Biol. 2005, 2, 81–88. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lynch, M. Feedforward loop for diversity. Nature 2015, 523, 414–416. [Google Scholar] [CrossRef] [PubMed]
- Agorio, A.; Durand, S.; Fiume, E.; Brousse, C.; Gy, I.; Simon, M.; Anava, S.; Rechavi, O.; Loudet, O.; Camilleri, C.; et al. An Arabidopsis Natural Epiallele Maintained by a Feed-Forward Silencing Loop between Histone and DNA. PLoS Genet. 2017, 13, e1006551. [Google Scholar] [CrossRef] [PubMed]
- Murugan, R. Theory on the Dynamics of Feedforward Loops in the Transcription Factor Networks. PLoS ONE 2012, 7, e41027. [Google Scholar] [CrossRef]
- Mangan, S.; Zaslaver, A.; Alon, U. The Coherent Feedforward Loop Serves as a Sign-sensitive Delay Element in Transcription Networks. J. Mol. Biol. 2003, 334, 197–204. [Google Scholar] [CrossRef]
- Pieters, P.A.; Nathalia, B.L.; van der Linden, A.J.; Yin, P.; Kim, J.; Huck, W.T.S.; de Greef, T.F.A. Cell-Free Characterization of Coherent Feed-Forward Loop-Based Synthetic Genetic Circuits. ACS Synth. Biol. 2021, 10, 1406–1416. [Google Scholar] [CrossRef]
- Kalir, S.; Mangan, S.; Alon, U. A coherent feed-forward loop with a SUM input function prolongs flagella expression in Escherichia coli. Mol. Syst. Biol. 2005, 1, 2005.0006. [Google Scholar] [CrossRef]
- Xiong, K.; Lancaster, A.K.; Siegal, M.L.; Masel, J. Feed-forward regulation adaptively evolves via dynamics rather than topology when there is intrinsic noise. Nat. Commun. 2019, 10, 2418. [Google Scholar] [CrossRef]
- Lluís, F.; Perdiguero, E.; Nebreda, A.R.; Muñoz-Cánoves, P. Regulation of skeletal muscle gene expression by p38 MAP kinases. Trends Cell Biol. 2006, 16, 36–44. [Google Scholar] [CrossRef]
- Penn, B.H.; Bergstrom, D.A.; Dilworth, F.J.; Bengal, E.; Tapscott, S.J. A MyoD-generated feed-forward circuit temporally patterns gene expression during skeletal muscle differentiation. Genes Dev. 2004, 18, 2348–2353. [Google Scholar] [CrossRef]
- Gimeno, C.J.; Ljungdahl, P.O.; Styles, C.A.; Fink, G.R. Unipolar cell divisions in the yeast S. cerevisiae lead to filamentous growth: Regulation by starvation and RAS. Cell 1992, 68, 1077–1090. [Google Scholar] [CrossRef]
- Wolf, J.J.; Dowell, R.D.; Mahony, S.; Rabani, M.; Gifford, D.K.; Fink, G.R. Feed-Forward Regulation of a Cell Fate Determinant by an RNA-Binding Protein Generates Asymmetry in Yeast. Genetics 2010, 185, 513–522. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Levine, J.H.; Lin, Y.; Elowitz, M.B. Functional Roles of Pulsing in Genetic Circuits. Science 2013, 342, 1193–1200. [Google Scholar] [CrossRef]
- Hao, N.; O’Shea, E.K. Signal-dependent dynamics of transcription factor translocation controls gene expression. Nat. Struct. Mol. Biol. 2012, 19, 31–39. [Google Scholar] [CrossRef]
- Yissachar, N.; Fischler, T.S.; Cohen, A.A.; Reich-Zeliger, S.; Russ, D.; Shifrut, E.; Porat, Z.; Friedman, N. Dynamic Response Diversity of NFAT Isoforms in Individual Living Cells. Mol. Cell 2013, 49, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, S.; Bren, A.; Dekel, E.; Alon, U. The incoherent feed-forward loop can generate non-monotonic input functions for genes. Mol. Syst. Biol. 2008, 4, 203. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Mehreja, R.; Thiberge, S.; Chen, M.-T.; Weiss, R. Spatiotemporal control of gene expression with pulse-generating networks. Proc. Natl. Acad. Sci. USA 2004, 101, 6355–6360. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Gerchman, Y.; Collins, C.; Arnold, F.H.; Weiss, R. A synthetic multicellular system for programmed pattern formation. Nature 2005, 434, 1130–1134. [Google Scholar] [CrossRef]
- Barone, F.; Dorr, F.; E. Marasco, L.; Mildiner, S.; Patop, I.L.; Sosa, S.; Vattino, L.G.; A. Vignale, F.; Altszyler, E.; Basanta, B.; et al. Design and evaluation of an incoherent feed-forward loop for an arsenic biosensor based on standard iGEM parts. Synth. Biol. 2017, 2, ysx006. [Google Scholar] [CrossRef]
- Entus, R.; Aufderheide, B.; Sauro, H.M. Design and implementation of three incoherent feed-forward motif based biological concentration sensors. Syst. Synth. Biol. 2007, 1, 119–128. [Google Scholar] [CrossRef]
- Xiong, H.; Veedu, R.; Diermeier, S. Recent Advances in Oligonucleotide Therapeutics in Oncology. Int. J. Mol. Sci. 2021, 22, 3295. [Google Scholar] [CrossRef] [PubMed]
- Reeves, G.T. The engineering principles of combining a transcriptional incoherent feedforward loop with negative feedback. J. Biol. Eng. 2019, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Lou, S.; Pan, X.; Huang, T.; Duan, B.; Yang, F.-C.; Yang, J.; Xiong, M.; Liu, Y.; Ma, Q. Incoherent Feed-Forward Regulatory Loops Control Segregation of C-Mechanoreceptors, Nociceptors, and Pruriceptors. J. Neurosci. 2015, 35, 5317–5329. [Google Scholar] [CrossRef] [PubMed]
- Lou, S.; Duan, B.; Vong, L.; Lowell, B.B.; Ma, Q. Runx1 Controls Terminal Morphology and Mechanosensitivity of VGLUT3-expressing C-Mechanoreceptors. J. Neurosci. 2013, 33, 870–882. [Google Scholar] [CrossRef] [PubMed]
- Abdusselamoglu, M.D.; Eroglu, E.; Burkard, T.R.; A. Knoblich, J. The transcription factor odd-paired regulates temporal identity in transit-amplifying neural progenitors via an incoherent feed-forward loop. eLife 2019, 8, e46566. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Shao, D.; Adler, M.; Charest, P.G.; Loomis, W.F.; Levine, H.; Groisman, A.; Rappel, W.-J.; Firtel, R.A. Incoherent Feedforward Control Governs Adaptation of Activated Ras in a Eukaryotic Chemotaxis Pathway. Sci. Signal. 2012, 5, ra2. [Google Scholar] [CrossRef]
- Janetopoulos, C.; Jin, T.; Devreotes, P. Receptor-Mediated Activation of Heterotrimeric G-Proteins in Living Cells. Science 2001, 291, 2408–2411. [Google Scholar] [CrossRef]
- Ma, W.; Trusina, A.; El-Samad, H.; Lim, W.A.; Tang, C. Defining Network Topologies that Can Achieve Biochemical Adaptation. Cell 2009, 138, 760–773. [Google Scholar] [CrossRef]
- Marshall, R.; Noireaux, V. Quantitative modeling of transcription and translation of an all-E. coli cell-free system. Sci. Rep. 2019, 9, 11980. [Google Scholar] [CrossRef]
- Shin, J.; Noireaux, V. An E. coli Cell-Free Expression Toolbox: Application to Synthetic Gene Circuits and Artificial Cells. ACS Synth. Biol. 2012, 1, 29–41. [Google Scholar] [CrossRef]
- Guo, S.; Murray, R.M. Construction of Incoherent Feedforward Loop Circuits in a Cell-Free System and in Cells. ACS Synth. Biol. 2019, 8, 606–610. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Anderson, J.; Contreras, L.M. Regulatory RNAs: Charming gene management styles for synthetic biology applications. RNA Biol. 2013, 10, 1778–1797. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Oertner, T.G.; Hegemann, P.; Larkum, M.E. Active cortical dendrites modulate perception. Science 2016, 354, 1587–1590. [Google Scholar] [CrossRef]
- Beisel, C.; Storz, G. Base pairing small RNAs and their roles in global regulatory networks. FEMS Microbiol. Rev. 2010, 34, 866–882. [Google Scholar] [CrossRef]
- Chappell, J.; Takahashi, M.; Lucks, J.B. Creating small transcription activating RNAs. Nat. Chem. Biol. 2015, 11, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Lucks, J.B.; Qi, L.; Mutalik, V.K.; Wang, D.; Arkin, A.P. Versatile RNA-sensing transcriptional regulators for engineering genetic networks. Proc. Natl. Acad. Sci. USA 2011, 108, 8617–8622. [Google Scholar] [CrossRef]
- Westbrook, A.; Tang, X.; Marshall, R.; Maxwell, C.S.; Chappell, J.; Agrawal, D.K.; Dunlop, M.J.; Noireaux, V.; Beisel, C.L.; Lucks, J.; et al. Distinct timescales of RNA regulators enable the construction of a genetic pulse generator. Biotechnol. Bioeng. 2019, 116, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Nakamura, M.; Srinivasan, P.; Chavez, M.; Carter, M.; Dominguez, A.A.; La Russa, M.; Lau, M.B.; Abbott, T.R.; Xu, X.; Zhao, D.; et al. Anti-CRISPR-mediated control of gene editing and synthetic circuits in eukaryotic cells. Nat. Commun. 2019, 10, 194. [Google Scholar] [CrossRef]
- Del Vecchio, D.; Ninfa, A.J.; Sontag, E.D. Modular cell biology: Retroactivity and insulation. Mol. Syst. Biol. 2008, 4, 161. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Belta, C.; Isaacson, S.A. How Retroactivity Affects the Behavior of Incoherent Feedforward Loops. iScience 2020, 23, 101779. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
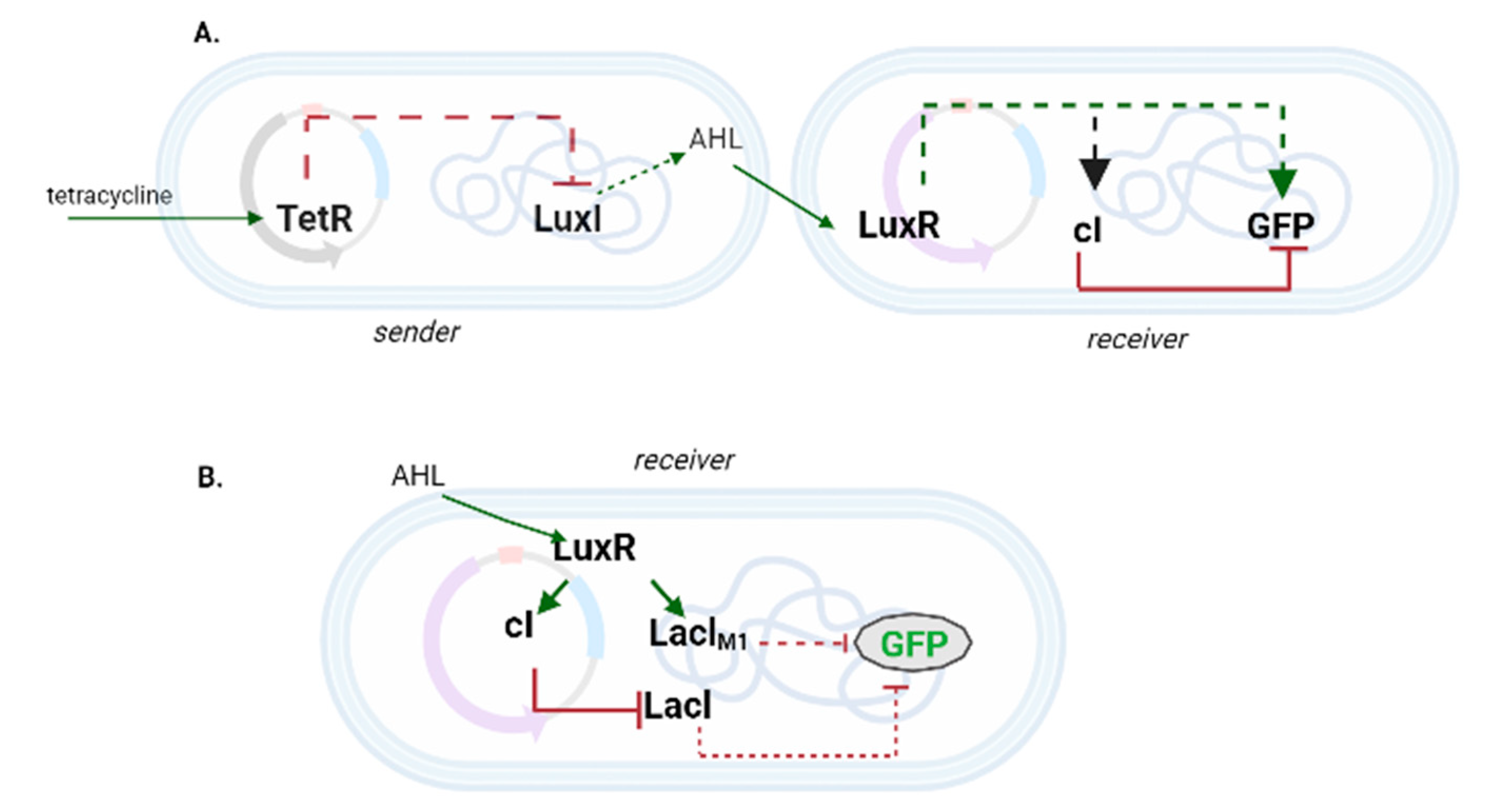{kind=link}
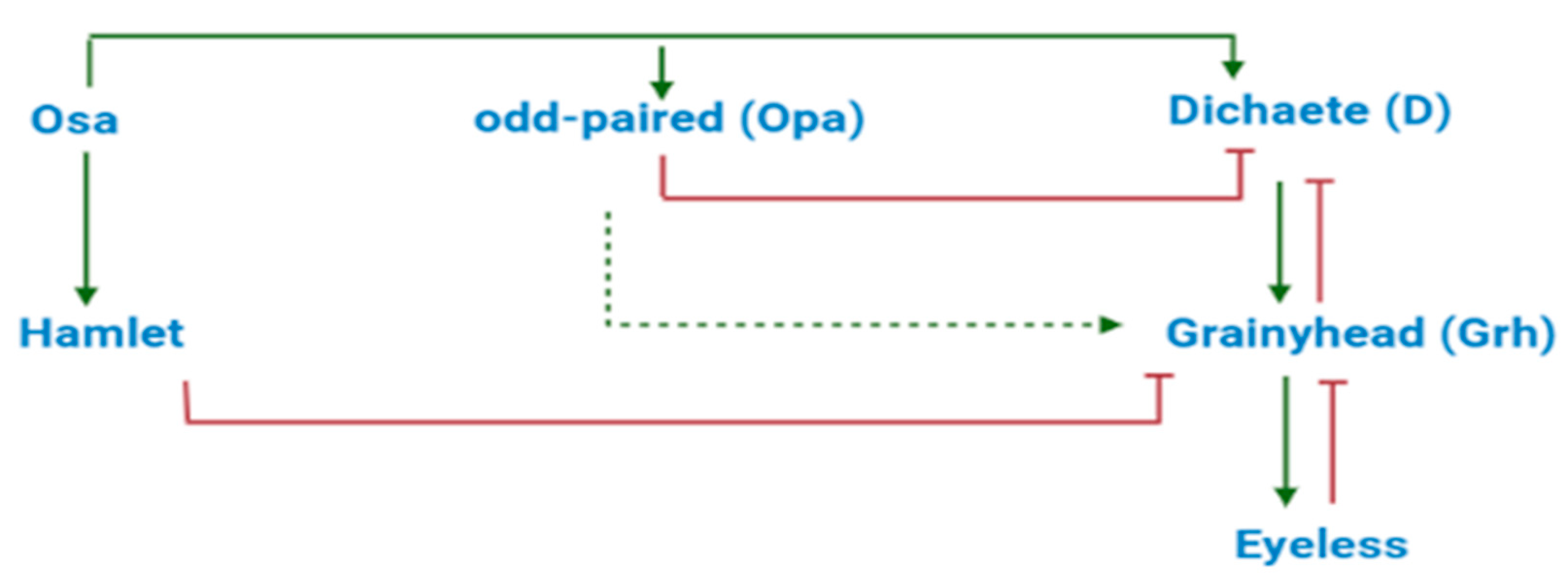{kind=link}
{kind=link}
{kind=link}
| Rare FFL | Abundance in E. coli | Abundance in S. cerevisiae |
|---|---|---|
| C2 | 2 | 5 |
| C3 | 4 | - |
| C4 | 1 | - |
| I2 | - | 3 |
| I3 | 1 | 1 |
| I4 | 1 | - |
| Type | Unique Features [Steady-State (Ss) with AND/OR Gates, Response Delay or Acceleration] | |
|---|---|---|
| Coherent | C1 | Ss is sensitive to both Sx and Sy, and shows delay from Sx on step with AND gate. Whereas it is sensitive to Sx only and delay occurs from Sx off step with OR gate. |
| C2 | Ss is sensitive to both Sx and Sy and shows delay from Sx off step with AND gate, whereas it is not sensitive to Sx only and delay occurs from Sx on step with OR gate. | |
| C3 | Ss is not sensitive to Sx with AND gate and not sensitive with Sx and Sy with OR gate. Delay occurs from Sx off step in both logic gates. | |
| C4 | Ss is sensitive to only Sx with AND gate and not sensitive with Sx or Sy with OR gate. Delay occurs from Sx on step in both logic gates. | |
| Incoherent | I1 | Ss is sensitive to both Sx but not to Sy. Weak pulsers from Sx on step and Sy effect destroys pulse, accelerate response only from Sx on step. |
| I2 | Ss is not sensitive to both Sx and Sy. Weak pulsers from Sx off step and Sy effect destroys pulse, accelerate response from Sx off step. | |
| I3 | Strong pulsers from Sx off step and Sy effect enables pulse generation, accelerate response from Sx off step. | |
| I4 | Strong pulsers from Sx on step and Sy effect enables pulse generation, accelerate response from Sx on step. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weldemichael, T.; Asemoloye, M.D.; Marchisio, M.A. Feedforward Loops: Evolutionary Conserved Network Motifs Redesigned for Synthetic Biology Applications. Appl. Sci. 2022, 12, 8292. https://doi.org/10.3390/app12168292
Weldemichael T, Asemoloye MD, Marchisio MA. Feedforward Loops: Evolutionary Conserved Network Motifs Redesigned for Synthetic Biology Applications. Applied Sciences. 2022; 12(16):8292. https://doi.org/10.3390/app12168292
Chicago/Turabian StyleWeldemichael, Tsigereda, Michael Dare Asemoloye, and Mario Andrea Marchisio. 2022. "Feedforward Loops: Evolutionary Conserved Network Motifs Redesigned for Synthetic Biology Applications" Applied Sciences 12, no. 16: 8292. https://doi.org/10.3390/app12168292
APA StyleWeldemichael, T., Asemoloye, M. D., & Marchisio, M. A. (2022). Feedforward Loops: Evolutionary Conserved Network Motifs Redesigned for Synthetic Biology Applications. Applied Sciences, 12(16), 8292. https://doi.org/10.3390/app12168292