
A Molecular Insight into the Synergistic Mechanism of Nigella sativa (Black Cumin) with β-Lactam Antibiotics against Clinical Isolates of Methicillin-Resistant Staphylococcus aureus


Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Considerations
2.2. Bacterial Isolates and Antimicrobial Susceptibility Determination
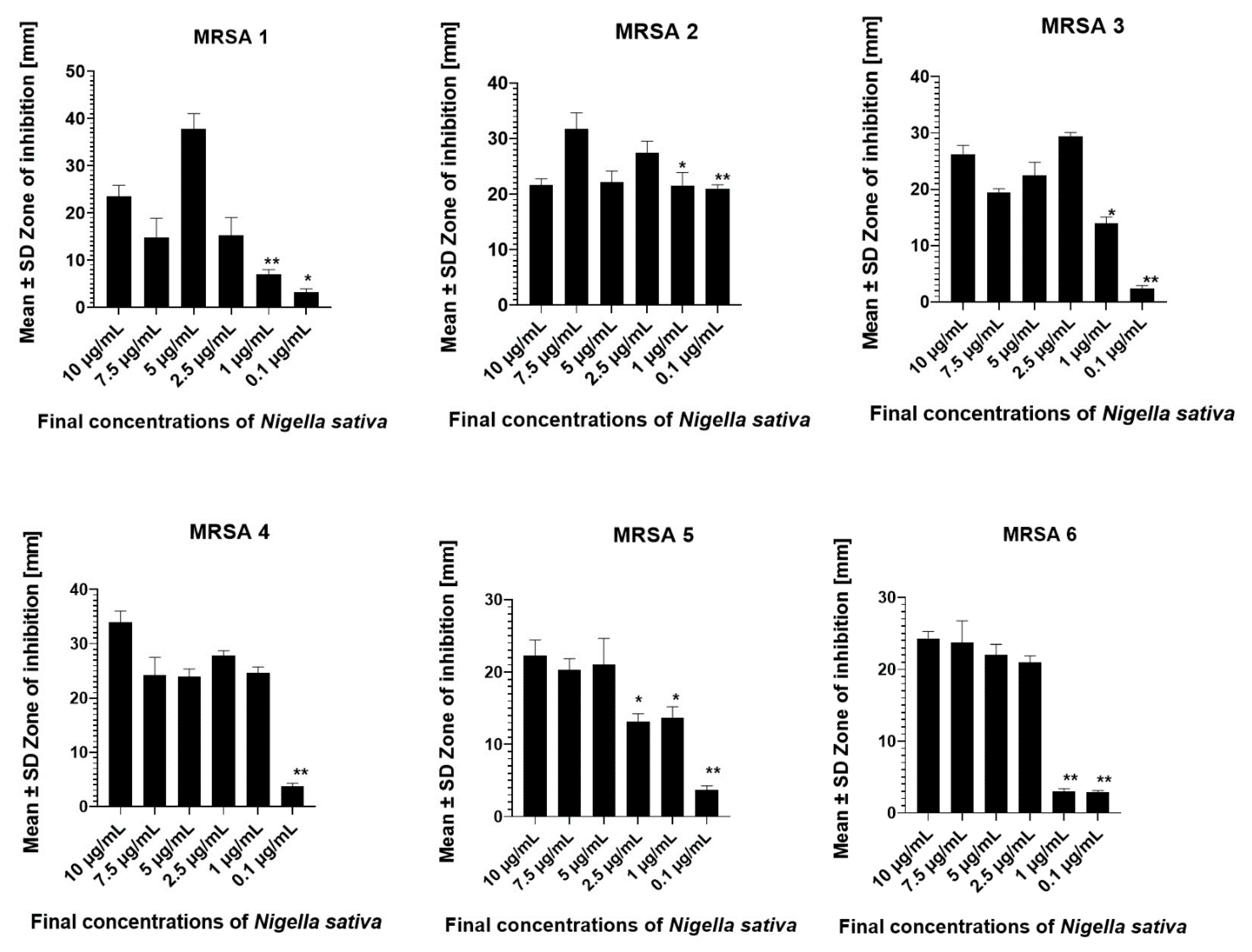
2.3. Nigella sativa and MRSA Susceptibility Test
2.4. Determination of Time–Kill Assay
2.5. Antimicrobial Synergistic NS and β-Lactam Antibiotics Assay
2.6. Molecular Assay by SEM and TEM Microscopy
2.7. Gas Chromatography/Mass Spectrometry (GC/MS) Analysis
2.8. Statistical Analysis
3. Results
3.1. Antimicrobial Susceptibility of the MRSA Isolates
3.2. Time–Kill Kinetics
3.3. Synergistic Effect of NS and β-Lactams with FIC Index Analysis
3.4. SEM and TEM Assay on Effect of Combined NS, Antibiotic Treatment
3.5. Results of GC-MS Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ippolito, G.; Leone, S.; Lauria, F.N.; Nicastri, E.; Wenzel, R.P. Methicillin-resistant Staphylococcus aureus: The superbug. Int. J. Infect. Dis. 2010, 14, S7–S11. [Google Scholar] [CrossRef]
- Papp-Wallace, K.M.; Endimiani, A.; Taracila, M.A.; Bonomo, R.A. Carbapenems: Past, Present, and Future. Antimicrob. Agents Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef] [PubMed]
- Konaté, K.; Mavoungou, J.F.; Lepengué, A.N.; Aworet-Samseny, R.R.; Hilou, A.; Souza, A.; Dicko, M.H.; M’Batchi, B. Antibacterial activity against β- lactamase producing Methicillin and Ampicillin-resistants Staphylococcus aureus: Fractional Inhibitory Concentration Index (FICI) determination. Ann. Clin. Microbiol. Antimicrob. 2012, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Bæk, K.T.; Gründling, A.; Mogensen, R.G.; Thøgersen, L.; Petersen, A.; Paulander, W.; Frees, D. β-Lactam Resistance in Methicillin-Resistant Staphylococcus aureus USA300 Is Increased by Inactivation of the ClpXP Protease. Antimicrob. Agents Chemother. 2014, 58, 4593–4603. [Google Scholar] [CrossRef]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin Resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar] [PubMed]
- Lesho, E.; Yoon, E.-J.; McGann, P.; Snesrud, E.; Kwak, Y.; Milillo, M.; Onmus-Leone, F.; Preston, L.; Clair, K.S.; Nikolich, M.; et al. Emergence of Colistin-Resistance in Extremely Drug-Resistant Acinetobacter baumannii Containing a Novel pmrCAB Operon During Colistin Therapy of Wound Infections. J. Infect. Dis. 2013, 208, 1142–1151. [Google Scholar] [CrossRef]
- Kaur, M.; Rai, J.; Randhawa, G.K. Recent advances in antibacterial drugs. Int. J. Appl. Basic Med. Res. 2013, 3, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef]
- Lee, K.; Yong, D.; Jeong, S.H.; Chong, Y. Multidrug-ResistantAcinetobacterspp.: Increasingly Problematic Nosocomial Pathogens. Yonsei Med. J. 2011, 52, 879–891. [Google Scholar] [CrossRef]
- Lim, T.-P.; Tan, T.-Y.; Lee, W.; Sasikala, S.; Tan, T.-T.; Hsu, L.-Y.; Kwa, A.L. In-Vitro Activity of Polymyxin B, Rifampicin, Tigecycline Alone and in Combination against Carbapenem-Resistant Acinetobacter baumannii in Singapore. PLoS ONE 2011, 6, e18485. [Google Scholar] [CrossRef]
- Albur, M.; Noel, A.; Bowker, K.; MacGowan, A. Bactericidal Activity of Multiple Combinations of Tigecycline and Colistin against NDM-1-Producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2012, 56, 3441–3443. [Google Scholar] [CrossRef]
- Urban, C.; Mariano, N.; Rahal, J.J. In Vitro Double and Triple Bactericidal Activities of Doripenem, Polymyxin B, and Rifampin against Multidrug-Resistant Acinetobacter baumannii, Pseudomonas aeruginosa, Klebsiella pneumoniae, and Escherichia coli. Antimicrob. Agents Chemother. 2010, 54, 2732–2734. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Chai, D.; Wang, R.; Liang, B.; Bai, N. Colistin resistance of Acinetobacter baumannii: Clinical reports, mechanisms and antimicrobial strategies. J. Antimicrob. Chemother. 2012, 67, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Gharaibeh, M.H.; Shatnawi, S.Q. An overview of colistin resistance, mobilized colistin resistance genes dissemination, global responses, and the alternatives to colistin: A review. Veter. World 2019, 12, 1735–1746. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Beaudry, F.; Letellier, A. Resistance to colistin: What is the fate for this antibiotic in pig production? Int. J. Antimicrob. Agents 2016, 48, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Barbieri, N.L.; Nielsen, D.W.; Wannemuehler, Y.; Cavender, T.; Hussein, A.; Yan, S.-G.; Nolan, L.K.; Logue, C.M. mcr-1 identified in Avian Pathogenic Escherichia coli (APEC). PLoS ONE 2017, 12, e0172997. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, A.; Abdulrahman, A.A.; Aboulmagd, E.; Alsultan, A.A. Synergic bactericidal activity of novel antibiotic combinations against extreme drug resistant Pseudomonas aeruginosa and Acinetobacter baumannii. Afr. J. Microbiol. Res. 2014, 8, 856–861. [Google Scholar] [CrossRef]
- Pena-Miller, R.; Laehnemann, D.; Jansen, G.; Fuentes-Hernandez, A.; Rosenstiel, P.; Schulenburg, H.; Beardmore, R. When the Most Potent Combination of Antibiotics Selects for the Greatest Bacterial Load: The Smile-Frown Transition. PLoS Biol. 2013, 11, e1001540. [Google Scholar] [CrossRef] [PubMed]
- Sibanda, T.; Okoh, A.I. The challenges of overcoming antibiotic resistance: Plant extracts as potential sources of antimicrobial and resistance modifying agents. Afr. J. Biotechnol. 2007, 6, 2886–2896. [Google Scholar]
- Hübsch, Z.; Van Zyl, R.; Cock, I.; Van Vuuren, S. Interactive antimicrobial and toxicity profiles of conventional antimicrobials with Southern African medicinal plants. S. Afr. J. Bot. 2014, 93, 185–197. [Google Scholar] [CrossRef]
- Aiyegoro, O.A.; Okoh, A.I. Use of bioactive plant products in combination with standard antibiotics: Implications in antimicrobial chemotherapy. J. Med. Plant Res. 2009, 3, 1147–1152. [Google Scholar]
- Haroun, M.F.; Al-Kayali, R.S. Synergistic effect of Thymbra spicata L. extracts with antibiotics against multidrug- resistant Staphylococcus aureus and Klebsiella pneumoniae strains. Iran. J. Basic Med. Sci. 2016, 19, 1193–1200. [Google Scholar] [PubMed]
- Teow, S.-Y.; Liew, K.; Ali, S.A.; Khoo, A.S.-B.; Peh, S.-C. Antibacterial Action of Curcumin against Staphylococcus aureus: A Brief Review. J. Trop. Med. 2016, 2016, 1–10. [Google Scholar] [CrossRef]
- Emeka, L.B.; Emeka, P.M.; Khan, T.M. Antimicrobial activity of Nigella sativa L. seed oil against multi-drug resistant Staphylococcus aureus isolated from diabetic wounds. Pak. J. Pharm. Sci. 2015, 28, 1985–1990. [Google Scholar] [PubMed]
- Ugur, A.R.; Dagi, H.T.; Ozturk, B.; Tekin, G.; Findik, D. Assessment of In vitro antibacterial activity and cytotoxicity effect of Nigella sativa oil. Pharmacogn. Mag. 2016, 12, 471–S474. [Google Scholar] [CrossRef] [PubMed]
- Utami, A.T.; Pratomo, B.; Noorhamdani. Study of Antimicrobial Activity of Black Cumin Seeds (Nigella sativa L.) Against Salmonella typhi In Vitro. J. Med. Surg. Pathol. 2016, 1, 3. [Google Scholar] [CrossRef]
- Emeka, P.M.; Badger-Emeka, L.I.; Eneh, C.M.; Khan, T.M. Dietary supplementation of chloroquine with nigella sativa seed and oil extracts in the treatment of malaria induced in mice with plasmodium berghei. Pharmacogn. Mag. 2014, 10, S357–S362. [Google Scholar] [CrossRef] [PubMed]
- Magaldi, S.; Mata-Essayag, S.; de Capriles, C.H.; Perez, C.; Colella, M.; Olaizola, C.; Ontiveros, Y. Well diffusion for antifungal susceptibility testing. Int. J. Infect. Dis. 2004, 8, 39–45. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Methods for Determining Bactericidal Activity of Antimicrobial Agents; Approved Guideline, CLSI document M26-A; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 1998. [Google Scholar]
- Uzair, B.; Hameed, A.; Nazir, S.; Khan, B.A.; Fasim, F.; Khan, S.; Menaa, F. Synergism Between Nigella sativa Seeds Extract and Synthetic Antibiotics Against Mec A Gene Positive Human Strains of Staphylococcus aureus. Int. J. Pharmacol. 2017, 13, 958–968. [Google Scholar] [CrossRef]
- Sopirala, M.M.; Mangino, J.E.; Gebreyes, W.A.; Biller, B.; Bannerman, T.; Balada-Llasat, J.-M.; Pancholi, P. Synergy Testing by Etest, Microdilution Checkerboard, and Time-Kill Methods for Pan-Drug-Resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010, 54, 4678–4683. [Google Scholar] [CrossRef] [PubMed]
- Meletiadis, J.; Pournaras, S.; Roilides, E.; Walsh, T.J. Defining fractional inhibitory concentration index cut offs for additive interactions based on self-drug additive combinations, Monte Carlo simulation analysis, and in vitro-in vivo correlation data for antifungal drug combinations against Aspergillus fumigatus. Antimicrob. Agents Chemother. 2010, 54, 602–609. [Google Scholar] [CrossRef]
- Meletiadis, J.; Mouton, J.W.; Meis, J.F.G.M.; Bouman, B.A.; Donnelly, P.J.; Verweij, P.E.; Eurofung Network. Comparison of Spectrophotometric and Visual Readings of NCCLS Method and Evaluation of a Colorimetric Method Based on Reduction of a Soluble Tetrazolium Salt, 2,3-Bis {2-Methoxy-4-Nitro-5-[(Sulfenylamino) Carbonyl]-2H- Tetrazolium-Hydroxide}, for Antifungal Susceptibility Testing of Aspergillus Species. J. Clin. Microbiol. 2001, 39, 4256–4263. [Google Scholar] [CrossRef] [PubMed]
- Thammawat, S.; Sangdee, K. Time-kill profiles and cell-surface morphological effects of crude Polycephalomyces nipponicus Cod-MK1201 mycelial extract against antibiotic-sensitive and -resistant Staphylococcus aureus. Trop. J. Pharm. Res. 2017, 16, 407. [Google Scholar] [CrossRef]
- Thomas, P.; Reddy, K.M. Microscopic elucidation of abundant endophytic bacteria colonizing the cell wall–plasma membrane peri-space in the shoot-tip tissue of banana. AoB Plants 2013, 5, 11. [Google Scholar] [CrossRef]
- Ran, Y.; Hu, W.; Zhuang, K.; Lu, M.; Huang, J.; Xu, F.; Xu, X.; Hua, X.; Lama, J.; Ran, X.; et al. Observation of Viruses, Bacteria, and Fungi in Clinical Skin Samples under Transmission Electron Microscopy. In The Transmission Electron Microscope—Theory and Applications; Khan Maaz; IntechOpen: Rijeka, Croatia, 2015. [Google Scholar] [CrossRef]
- Falagas, M.E.; Karageorgopoulos, D.E. Pandrug Resistance (PDR), Extensive Drug Resistance (XDR), and Multidrug Resistance (MDR) among Gram-Negative Bacilli: Need for International Harmonization in Terminology. Clin. Infect. Dis. 2008, 46, 1121–1122. [Google Scholar] [CrossRef] [PubMed]
- Basak, S.; Singh, P.; Rajurkar, M. Multidrug Resistant and Extensively Drug Resistant Bacteria: A Study. J. Pathog. 2016, 2016, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.T.; Lubbers, B.V.; Schwarz, S.; Watts, J.L. Applying definitions for multidrug resistance, extensive drug resistance and pandrug resistance to clinically significant livestock and companion animal bacterial pathogens. J. Antimicrob. Chemother. 2018, 73, 1460–1463. [Google Scholar] [CrossRef] [PubMed]
- Naz, H. Nigella sativa: The miraculous herb. Pak. J. Biochem. Mol. Biol. 2011, 44, 44–48. Available online: http://pjbmb.org.pk/images/PJBMBArchive/2011/PJBMB_44_1_Mar_2011/10.pdf (accessed on 26 March 2021).
- Chaieb, K.; Kouidhi, B.; Jrah, H.; Mahdouani, K.; Bakhrouf, A. Antibacterial activity of Thymoquinone, an active principle of Nigella sativa and its potency to prevent bacterial biofilm formation. BMC Complement. Altern. Med. 2011, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H. Synergy research: Approaching a new generation of phytopharmaceuticals. Fitoterapia 2011, 82, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25859123 (accessed on 26 March 2021).
- Thakur, P.; Chawla, R.; Goel, R.; Narula, A.; Arora, R.; Sharma, R.K. Augmenting the potency of third-line antibiotics with Berberis aristata: In vitro synergistic activity against carbapenem-resistant Escherichia coli. J. Glob. Antimicrob. Resist. 2016, 6, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Rosato, A.; Vitali, C.; De Laurentis, N.; Armenise, D.; Milillo, M.A. Antibacterial effect of some essential oils administered alone or in combination with Norfloxacin. Phytomedicine 2007, 14, 727–732. [Google Scholar] [CrossRef]
- Ozkaya, G.U.; Durak, M.Z.; Akyar, I.; Karatuna, O. Antimicrobial Susceptibility Test for the Determination of Resistant and Susceptible S. aureus and Enterococcus spp. Using a Multi-Channel Surface Plasmon Resonance Device. Diagnostics 2019, 9, 191. [Google Scholar] [CrossRef]
- Park, H.; Jang, C.-H.; Cho, Y.B.; Choi, C.-H. Antibacterial effect of tea-tree oil on methicillin-resistant Staphylococcus aureus biofilm formation of the tympanostomy tube: An in vitro study. In Vivo 2008, 21, 1027–1030. [Google Scholar]
- Pinho, M.G.; Filipe, S.R.; De Lencastre, H.; Tomasz, A. Complementation of the Essential Peptidoglycan Transpeptidase Function of Penicillin-Binding Protein 2 (PBP2) by the Drug Resistance Protein PBP2A in Staphylococcus aureus. J. Bacteriol. 2001, 183, 6525–6531. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, M. Phytochemical Composition, Antioxidant, Anti-inflammatory and Antimicrobial Activity ofNigella sativaL. Essential Oil. J. Essent. Oil Bear. Plants 2014, 17, 1002–1011. [Google Scholar] [CrossRef]
- Herman, A.; Tambor, K.; Herman, A. Linalool Affects the Antimicrobial Efficacy of Essential Oils. Curr. Microbiol. 2015, 72, 165–172. [Google Scholar] [CrossRef]
- Forouzanfar, F.; Bazzaz, B.S.F.; Hosseinzadeh, H. Black cumin (Nigella sativa) and its constituent (thymoquinone): A review on antimicrobial effects. Iran. J. Basic Med. Sci. 2014, 17, 929–938. [Google Scholar] [PubMed]
- Kwiatkowski, P.; Pruss, A.; Masiuk, H.; Mnichowska-Polanowska, M.; Kaczmarek, M.; Giedrys-Kalemba, S.; Dołęgowska, B.; Zielińska-Bliźniewska, H.; Olszewski, J.; Sienkiewicz, M. The effect of fennel essential oil and trans-anethole on antibacterial activity of mupirocin against Staphylococcus aureus isolated from asymptomatic carriers. Adv. Dermatol. Allergol. 2019, 36, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Halawani, E. Antibacterial activity of thymoquinone and thymohydroquinone of Nigella sativa L. and their interaction with some antibiotics. Adv. Biol. Res. 2009, 3, 148–152. [Google Scholar]
- Elnour, S.A.; Abdelsalam, E.B. Some Biological and Pharmacological Effects of the Black Cumin (Nigella sativa): A Concise Review. Am. J. Res. Commun. 2018, 6, 10–35. Available online: http://www.usa-journals.com/wp-content/uploads/2018/02/Elnour_Vol63.pdf (accessed on 26 March 2021).
- Sahak, M.K.A.; Kabir, N.; Abbas, G.; Draman, S.; Hashim, N.H.; Adli, D.S.H. The Role ofNigella sativaand Its Active Constituents in Learning and Memory. Evid. Based Complement. Altern. Med. 2016, 2016, 1–6. [Google Scholar] [CrossRef]
- Dera, A.A.; Ahmad, I.; Rajagopalan, P.; Al Shahrani, M.; Saif, A.; Alshahrani, M.Y.; Alraey, Y.; Alamri, A.M.; Alasmari, S.; Makkawi, M.; et al. Synergistic efficacies of thymoquinone and standard antibiotics against multi-drug resistant isolates. Saudi. Med. J. 2021, 42, 196–204. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic | MRSA 1 | MRSA 2 | MRSA 3 | MRSA 4 | MRSA 5 | MRSA 6 |
|---|---|---|---|---|---|---|
| Augmentin | ≤2 | ≤2 | ≤2 | ≤2 | ≤2 | ≤2 |
| Benzyl penicillin | ≤0.25 | ≤0.25 | ≤0.25 | ≤0.25 | ≤0.25 | ≤0.25 |
| Oxacillin | ≥4 | ≥8 | ≥8 | ≥4 | ≥8 | ≥4 |
| Cefuroxime | ≤1 | 8 | 2 | 8 | ≤1 | ≤1 |
| Cefuroxime/Axetil | 8 | ≤1 | ≥64 | 8 | ≥64 | 2 |
| Clindamycin | ≤0.5 | 2 | ≤0.5 | ≥4 | ≥4 | ≥4 |
| Amikacin | ≥16 | 8 | ≤4 | ≥16 | ≥16 | ≥16 |
| Imipenem | ≤1 | ≤1 | ≤1 | ≤1 | ≤1 | ≤1 |
| Ciprofloxacin | ≥8 | ≥8 | ≥8 | ≥8 | ≥8 | ≥8 |
| Levofloxacin | ≥8 | ≥8 | 4 | ≥4 | ≥8 | ≥8 |
| Erythromycin | ≤0.25 | ≥8 | ≥8 | ≥8 | 1 | ≥8 |
| Sulfamethoxazole/trimethoprim | 8 | 16 | 16 | 8 | 16 | 16 |
| Tigecycline | ≤0.12 | ≤0.12 | ≤0.12 | ≤0.12 | ≤0.12 | 256 |
| Tetracycline | ≥32 | ≥32 | ≥32 | ≥32 | ≥32 | ≥320 |
| Rifampicin | ≤1 | ≤0.5 | ≥4 | 2 | ≥4 | ≥4 |
| Gentamicin | ≥16 | ≤4 | ≤4 | ≤4 | ≤4 | 8 |
| Bacterial Isolate/MRSA Type | Antibiotic Combination | Fractional Inhibitory Concentration Index | Outcome | |||
|---|---|---|---|---|---|---|
| Time (h) | ||||||
| 2 | 4 | 8 | 12 | |||
| MRSA 1 | NS + OXA | 0.4 | 0.5 | 0.7 | 0.7 | Synergism/Additive Effect |
| NS + AUG | 0.3 | 0.5 | 0.6 | 0.5 | Synergism/Additive Effect | |
| NS + CEF | 0.4 | 0.5 | 0.7 | 0.6 | Synergism/Additive Effect | |
| MRSA 4 | ||||||
| NS + OXA | 0.5 | 0.6 | 0.7 | 0.8 | Synergism/Additive Effect | |
| NS + AUG | 0.5 | 0.5 | 0.6 | 0.8 | Synergism/Additive Effect | |
| NS + CEF | 0.5 | 0.5 | 0.7 | 0.7 | Synergism/Additive Effect | |
| MRSA 5 | ||||||
| NS + OXA | 0.5 | 0.5 | 0.5 | 0.8 | Synergism/Additive Effect | |
| NS + AUG | 0.5 | 0.5 | 0.5 | 0.7 | Synergism/Additive Effect | |
| NS + CEF | 0.7 | 0.6 | 0.6 | 0.7 | Additive Effect | |
| Com Pound Name | Rt | Area (%) |
|---|---|---|
| p-Cymene | 9.841 | 5.18 |
| Linalool | 13.498 | 1.12 |
| Thymoquinone | 18.018 | 7.85 |
| trans-Anethole | 19.023 | 1.52 |
| m-Thymol | 19.872 | 0.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badger-Emeka, L.I.; Emeka, P.M.; Ibrahim, H.I.M. A Molecular Insight into the Synergistic Mechanism of Nigella sativa (Black Cumin) with β-Lactam Antibiotics against Clinical Isolates of Methicillin-Resistant Staphylococcus aureus. Appl. Sci. 2021, 11, 3206. https://doi.org/10.3390/app11073206
Badger-Emeka LI, Emeka PM, Ibrahim HIM. A Molecular Insight into the Synergistic Mechanism of Nigella sativa (Black Cumin) with β-Lactam Antibiotics against Clinical Isolates of Methicillin-Resistant Staphylococcus aureus. Applied Sciences. 2021; 11(7):3206. https://doi.org/10.3390/app11073206
Chicago/Turabian StyleBadger-Emeka, Lorina I., Promise Madu Emeka, and Hairul Islam M. Ibrahim. 2021. "A Molecular Insight into the Synergistic Mechanism of Nigella sativa (Black Cumin) with β-Lactam Antibiotics against Clinical Isolates of Methicillin-Resistant Staphylococcus aureus" Applied Sciences 11, no. 7: 3206. https://doi.org/10.3390/app11073206
APA StyleBadger-Emeka, L. I., Emeka, P. M., & Ibrahim, H. I. M. (2021). A Molecular Insight into the Synergistic Mechanism of Nigella sativa (Black Cumin) with β-Lactam Antibiotics against Clinical Isolates of Methicillin-Resistant Staphylococcus aureus. Applied Sciences, 11(7), 3206. https://doi.org/10.3390/app11073206