
Response to Incremental Replacement of Palm Oil with Fish Oil in Starter Diet on Growth Performance, Plasma Metabolites, Ruminal Fermentation, and Behavior of Dairy Calves

 , , ,
, , ,  , and
, and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
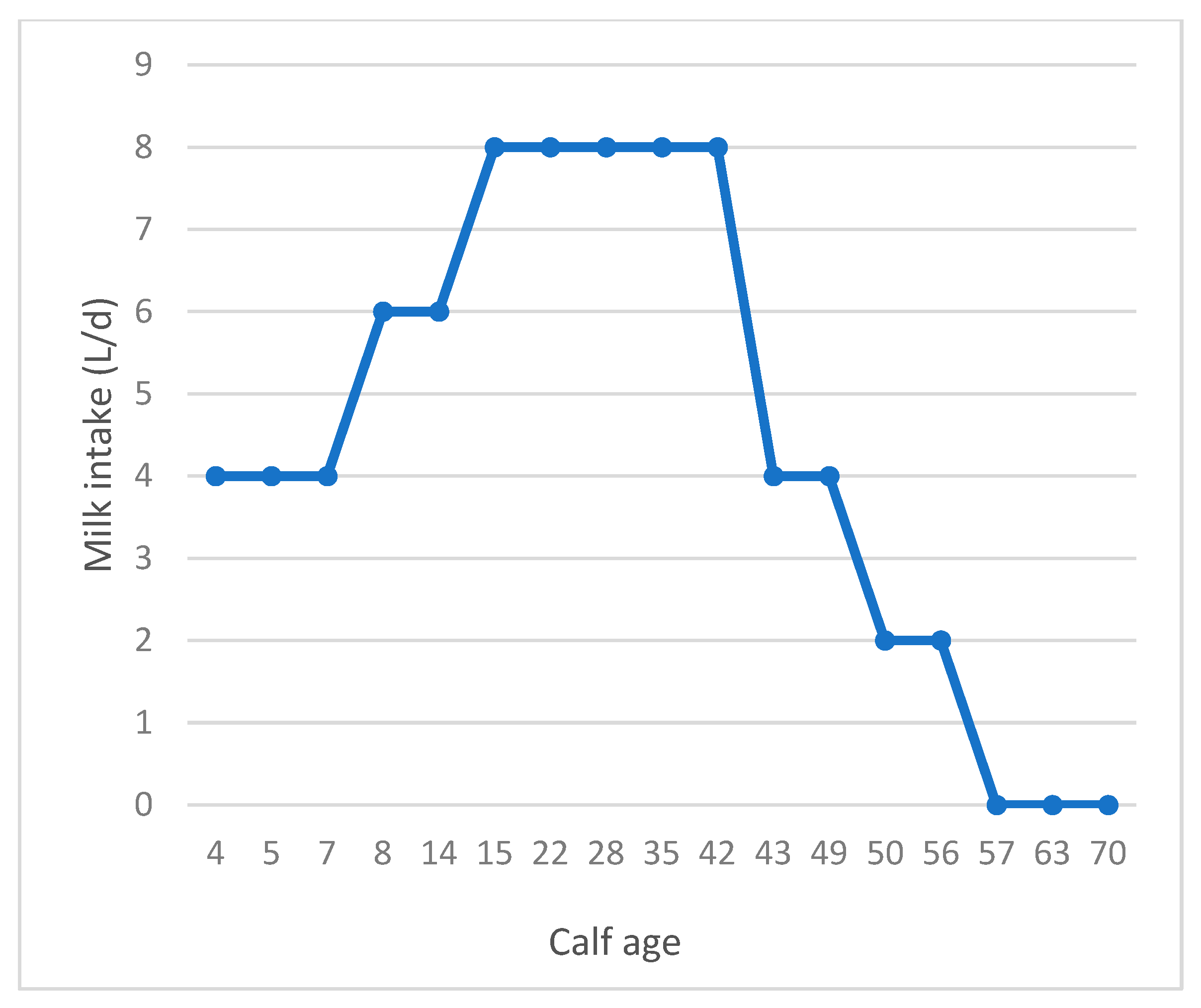
2.1. Animals, Treatments, and Management
2.2. Sampling
2.3. Statistical Analysis
3. Results
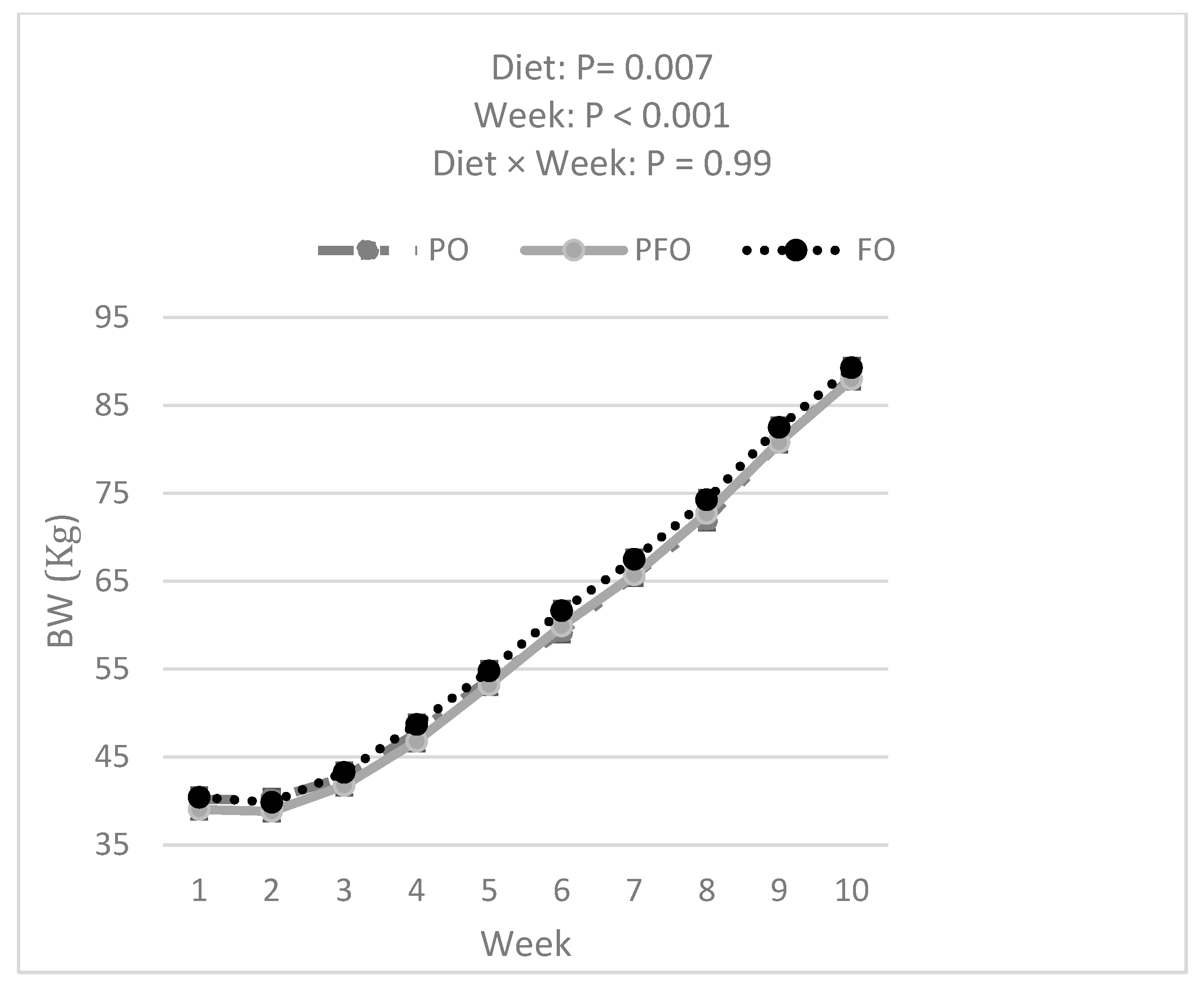
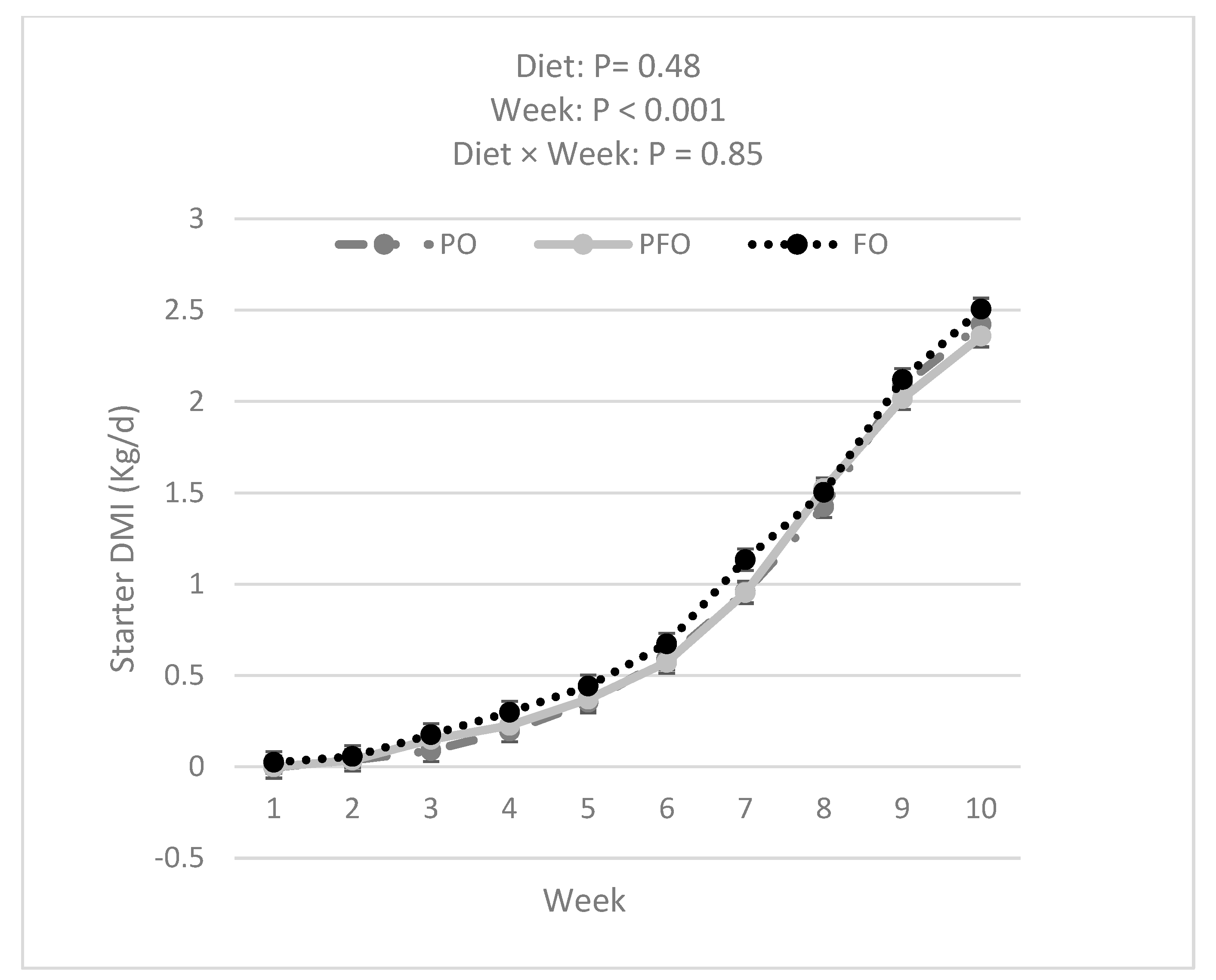
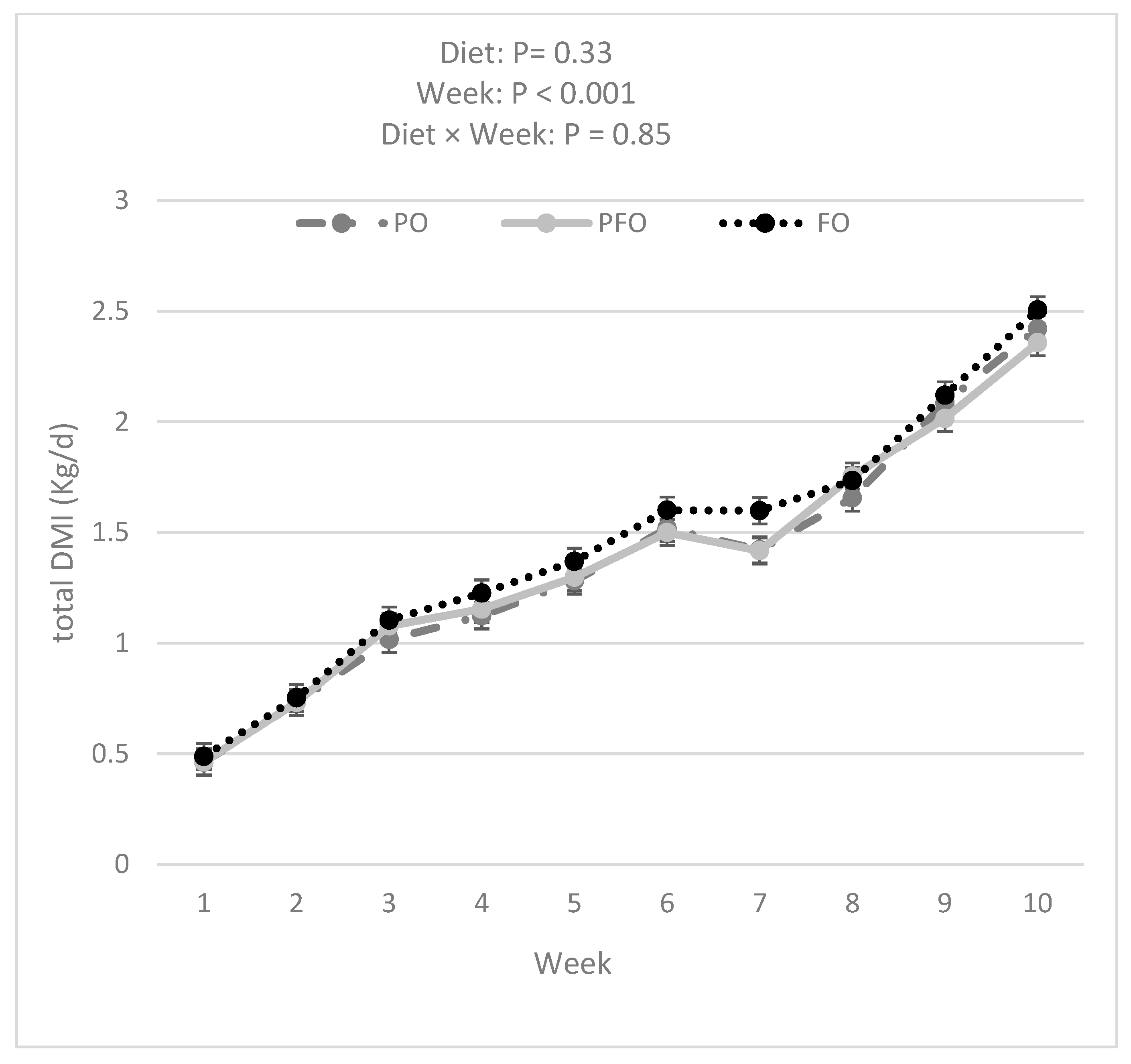
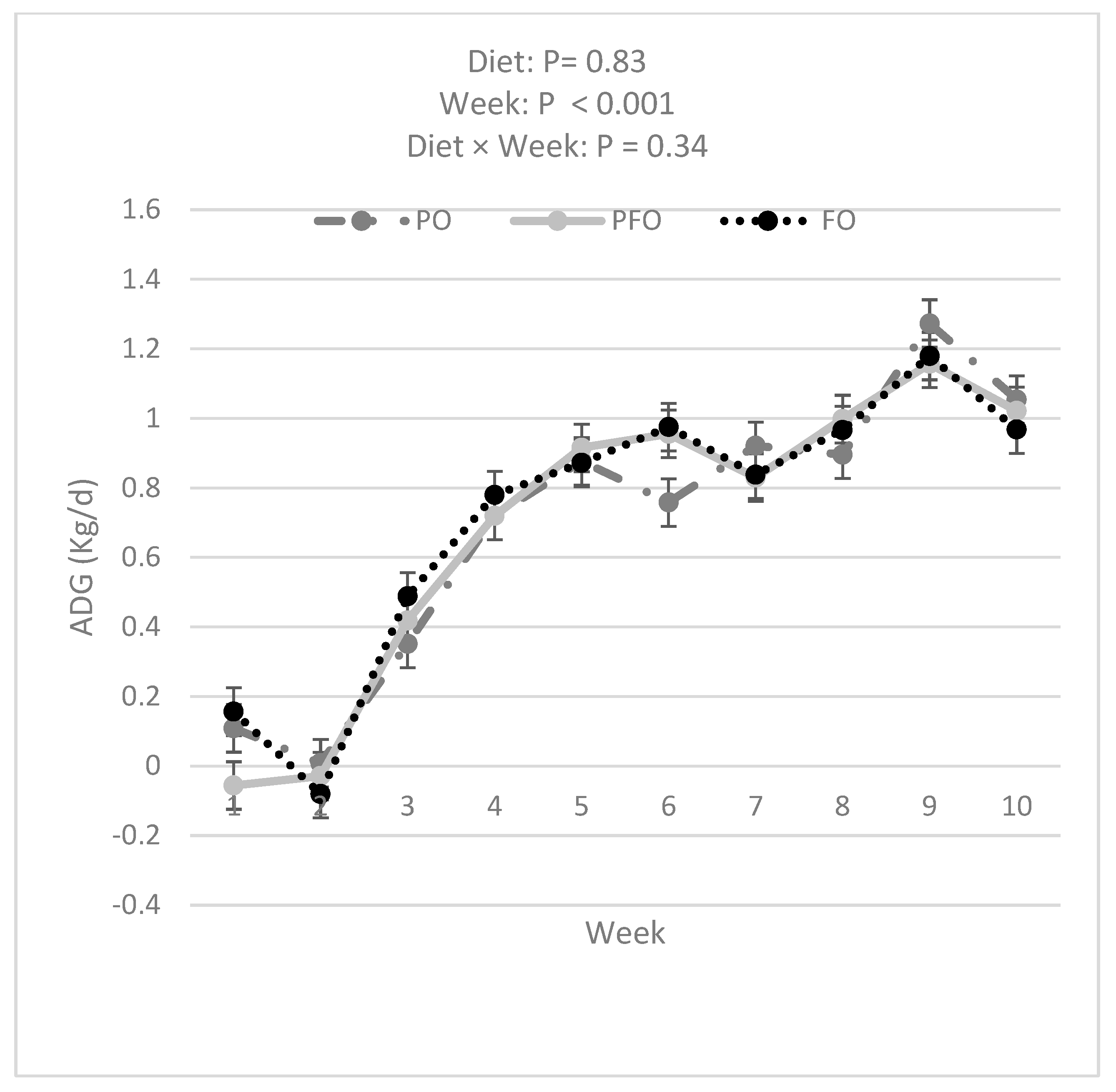
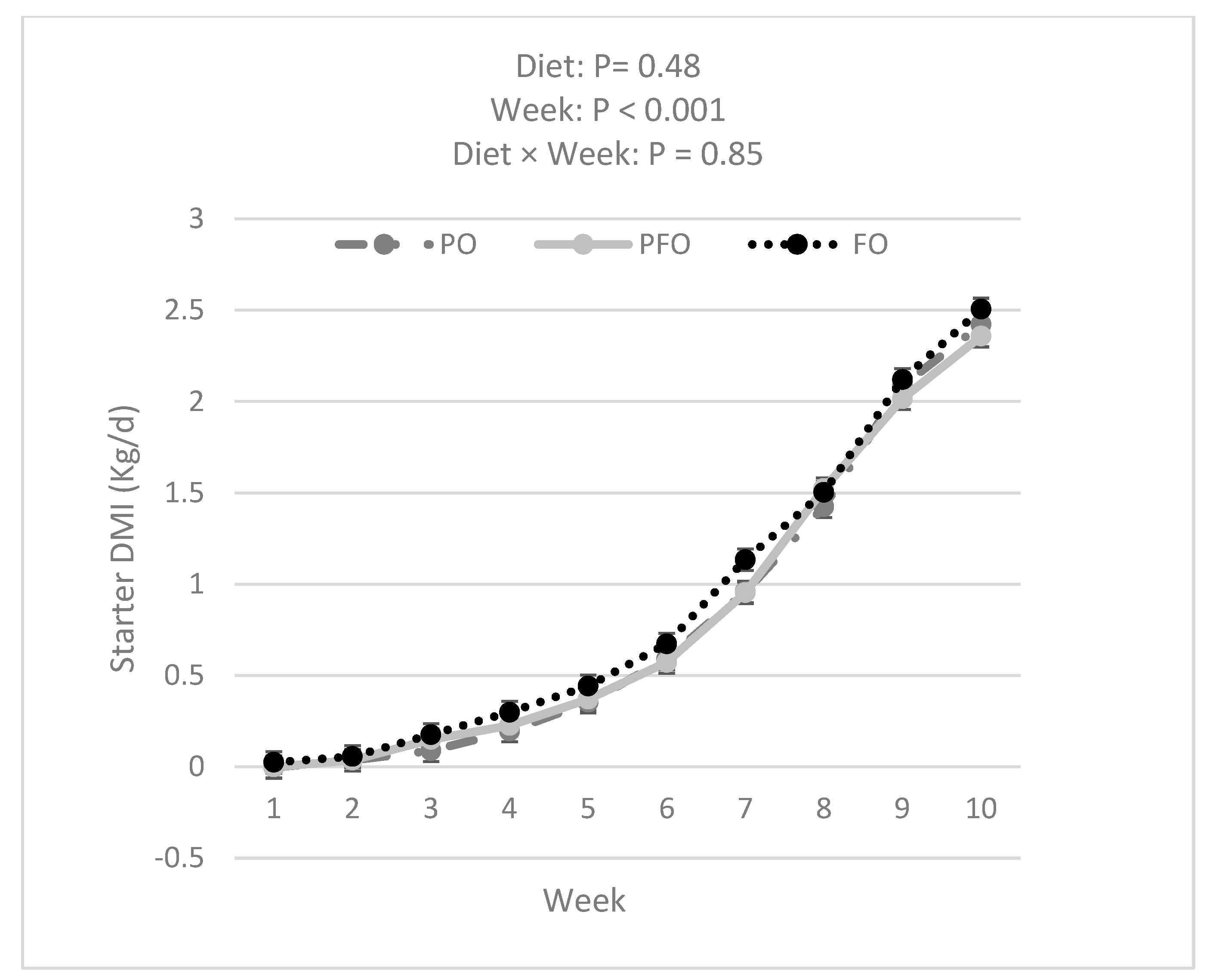
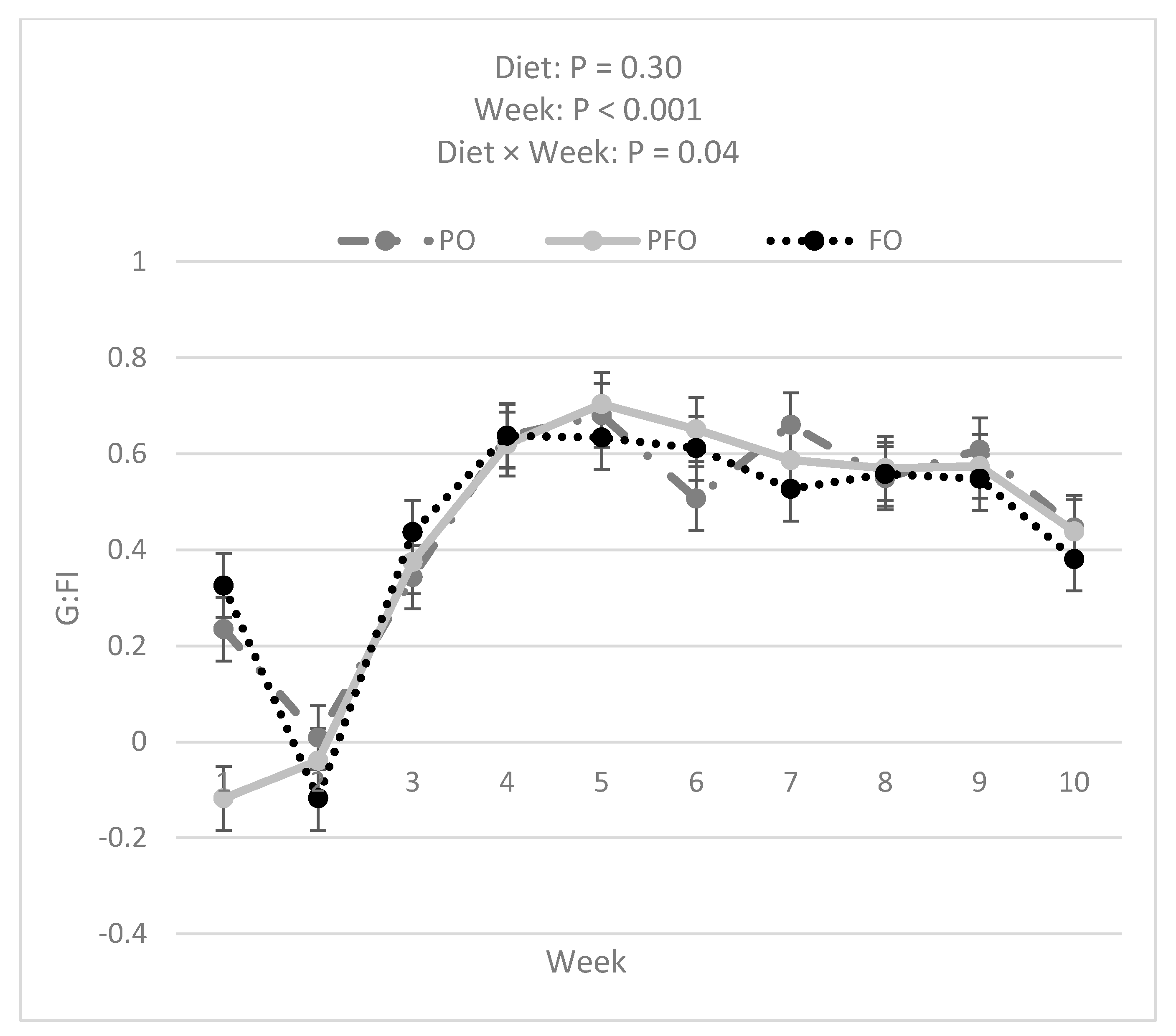
3.1. Weekly BW, ADG, Starter DMI, Total DMI, and G:FI
3.2. Growth Indices, Blood Variables, and Ruminal Fermentation Characteristics
3.3. Animal Behavioral Parameters and Calf Scour
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sordillo, L.M. Nutritional strategies to optimize dairy cattle immunity. J. Dairy. Sci. 2016, 99, 4967–4982. [Google Scholar] [CrossRef] [PubMed]
- Worden, L.C.; Erickson, M.G.; Gramer, S.; Tap, C.; Ylioja, C.; Trottier, N.; Preseault, C.L.; VandeHaar, M.J.; Lock, A.L.; Karcher, E.L. Short communication: Decreasing the dietary ratio of n-6 to n-3 fatty acids increases the n-3 concentration of peripheral blood mononuclear cells in weaned Holstein heifer calves. J. Dairy. Sci. 2018, 101, 1227–1233. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.M.; Aldrich, J.M.; Schlotterbeck, R.L.; Bateman, H.G. Effects of changing the fat and fatty acid composition of milk replacers fed to neonatal calves. Prof. Anim. Sci. 2007, 23, 135–143. [Google Scholar] [CrossRef]
- Hill, T.M.; Bateman, H.G.; Aldrich, J.M.; Schlotterbeck, R.L. Effects of changing the essential and functional fatty acid intake of dairy calves. J. Dairy. Sci. 2009, 92, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.M.; Bateman, H.G.; Aldrich, J.M.; Schlotterbeck, R.L. Effect of various fatty acids on dairy calf performance. Appl. Anim. Sci. 2011, 27, 167–175. [Google Scholar] [CrossRef]
- Panahiha, P.; Mirzaei-Alamouti, H.; Kazemi-Bonchenari, M.; Aschenbach, J.R. Growth performance, nutrient digestibility, and ruminal fermentation of dairy calves fed starter diets with alfalfa hay versus corn silage as forage and soybean oil versus palm fatty acids as fat source. J. Dairy. Sci. 2022, 105, 9597–9609. [Google Scholar] [CrossRef] [PubMed]
- Panahiha, P.; Mirzaei-Alamouti, H.; Kazemi-Bonchenari, M.; Poorhamdollah, M.; Vazirigohar, M.; Aschenbach, J.R. The type of lipid supplement has crucial implications for forage particle size in calf starter diets. J. Anim. Sci. Biotechnol. 2023, 14, 109. [Google Scholar] [CrossRef]
- Garcia, M.; Greco, L.F.; Favoreto, M.G.; Marsola, R.S.; Wang, D.; Shin, J.H.; Staples, C.R. Effect of supplementing essential fatty acids to pregnant nonlactating Holstein cows and their preweaned calves on calf performance, immune response, and health. J. Dairy. Sci. 2014, 97, 5045–5064. [Google Scholar] [CrossRef] [PubMed]
- Ballou, M.A.; DePeters, E.J. Supplementing milk replacer with omega-3 fatty acids from fish oil on immunocompetence and health of Jersey calves. J. Dairy. Sci. 2008, 91, 3488–3500. [Google Scholar] [CrossRef]
- Ghasemi, E.; Golabadi, D.; Piadeh, A. Effect of supplementing palmitic acid and altering the dietary ratio of n-6:n-3 fatty acids in low-fibre diets on production responses of dairy cows. Br. J. Nutr. 2021, 126, 355–365. [Google Scholar] [CrossRef]
- Karcher, E.L.; Hill, T.M.; Bateman, H.G.; Schlotterbeck, R.L.; Vito, N.; Sordillo, L.M.; VandeHaar, M.J. Comparison of supplementation of n-3 fatty acids from fish and flax oil on cytokine gene expression and growth of milk-fed Holstein calves. J. Dairy. Sci. 2014, 97, 2329–2337. [Google Scholar] [CrossRef]
- Melendez, P.; Roeschmann, C.F.; Baudo, A.; Tao, S.; Pinedo, P.; Kalantari, A.; Coarsey, M.; Bernard, J.K.; Naikare, H. Effect of fish oil and canola oil supplementation on immunological parameters, feed intake, and growth of Holstein calves. J. Dairy Sci. 2022, 105, 2509–2520. [Google Scholar] [CrossRef] [PubMed]
- Kazemi-Bonchenari, M.; Dehghan-Banadaky, M.; Fattahnia, F.; Saleh-Bahmanpour, A.; Jahani-Moghadam, M.; Mirzaei, M. Effects of linseed oil and rumen undegradable protein: Rumen degradable protein ratio on performance of Holstein dairy calves. Br. J. Nutr. 2020, 123, 1247–1257. [Google Scholar] [CrossRef]
- Griswold, D.E.; Adams, J.L. Constitutive cyclooxygenase (COX-1) and inducible cyclooxygenase (COX-2): Rationale for selective inhibition and progress to date. Med. Res. Rev. 1996, 16, 181–206. [Google Scholar] [CrossRef]
- Goldman, D.W.; Pickett, W.C.; Goetzl, E.J. Human neutrophil chemotactic and degranulating activities of leukotriene B5 (LTB5) derived from eicosapentaenoic acid. Biochem. Biophys. Res. Commun. 1983, 117, 282–288. [Google Scholar] [CrossRef]
- Raphael, W.; Sordillo, L.M. Dietary polyunsaturated fatty acids and inflammation: The role of phospholipid biosynthesis. Int. J. Mol. Sci. 2013, 14, 21167–21188. [Google Scholar] [CrossRef] [PubMed]
- Scaioli, E.; Liverani, E.; Belluzzi, A. The imbalance between n-6/n-3 polyunsaturated fatty acids and inflammatory bowel disease: A comprehensive review and future therapeutic perspectives. Int. J. Mol. Sci. 2017, 18, 2619. [Google Scholar] [CrossRef]
- Jin, J.; Boeglin, W.E.; Brash, A.R. Analysis of 12/15-lipoxygenase metabolism of EPA and DHA with special attention to authentication of docosatrienes. J. Lipid Res. 2021, 62, 100088. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Lee, H.J.; Lee, W.S.; Kim, H.S.; Kim, S.B.; Ki, K.S.; Choi, Y.J. Pre-and postweaning performance of Holstein female calves fed milk through step-down and conventional methods. J. Dairy. Sci. 2007, 90, 876–885. [Google Scholar] [CrossRef]
- Heinrichs, A.J.; Jones, C.M.; VanRoekel, L.R.; Fowler, M.A. Calf Track: A system of dairy calf workforce management, training, and evaluation and health evaluation. J. Dairy. Sci. 2003, 86, 10–3168. [Google Scholar]
- Mirzaei-Alamouti, H.; Moradi, S.; Shahalizadeh, Z.; Razavian, M.; Amanlou, H.; Harkinezhad, T.; Aschenbach, J.R. Both monensin and plant extract alter ruminal fermentation in sheep but only monensin affects the expression of genes involved in acid-base transport of the ruminal epithelium. Anim. Feed. Sci. Technol. 2016, 219, 132–143. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; AOAC: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Ebrahimi, M.; Rajion, M.A.; Jafari, S.; Faseleh Jahromi, M.; Oskoueian, E.; Qurni Sazili, A.; Ghaffari, M.H. Effects of dietary n-6: N-3 polyunsaturated fatty acid ratios on meat quality, carcass characteristics, tissue fatty acid profiles, and expression of lipogenic genes in growing goats. PLoS ONE 2018, 13, e0188369. [Google Scholar] [CrossRef] [PubMed]
- Miltko, R.; Majewska, M.P.; Bełżecki, G.; Kula, K.; Kowalik, B. Growth performance, carcass and meat quality of lambs supplemented different vegetable oils. Asian-Australas. J. Anim. Sci. 2019, 32, 767. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Adesogan, A.T.; Badinga, L.; Staples, C.R. Effects of dietary n-6:n-3 fatty acid ratio on feed intake, digestibility, and fatty acid profiles of the ruminal contents, liver, and muscle of growing lambs. J. Anim. Sci. 2007, 85, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Karami, M.; Ponnampalam, E.N.; Hopkins, D.L. The effect of palm oil or canola oil on feedlot performance, plasma and tissue fatty acid profile and meat quality in goats. Meat Sci. 2013, 94, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, E.; Azad-Shahraki, M.; Khorvash, M. Effect of different fat supplements on performance of dairy calves during cold season. J. Dairy. Sci. 2017, 100, 5319–5328. [Google Scholar] [CrossRef] [PubMed]
- Śpitalniak-Bajerska, K.; Szumny, A.; Pogoda-Sewerniak, K.; Kupczyński, R. Effects of n-3 fatty acids on growth, antioxidant status, and immunity of preweaned dairy calves. J. Dairy Sci. 2020, 103, 2864–2876. [Google Scholar] [CrossRef]
- Mirzaei-Alamouti, H.; Panahiha, P.; Patra, A.K.; Mansouryar, M. Effects of prepartum diet grain type and postpartum starch level on milk production, milk composition, and plasma metabolites of primiparous and multiparous Holstein cows. Anim. Feed. Sci. Technol. 2022, 291, 115393. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets 1 | |||
|---|---|---|---|
| Item | PO | PFO | FO |
| Ground barley grain | 15.9 | 15.9 | 16.0 |
| Ground corn grain | 42.5 | 42.5 | 42.5 |
| Wheat bran | 4.50 | 4.80 | 4.90 |
| Soybean meal | 24.7 | 24.6 | 24.5 |
| Extruded soybean | 1.60 | 1.60 | 1.60 |
| Corn gluten meal | 1.60 | 1.60 | 1.60 |
| Prilled palm fatty acids 2 | 3.30 | 1.70 | 0.00 |
| Fish oil (calcium salt) 3 | 0.00 | 1.90 | 3.90 |
| Sodium bicarbonate | 1.30 | 1.30 | 1.30 |
| Salt | 0.40 | 0.40 | 0.40 |
| Calcium carbonate | 0.90 | 0.40 | 0.00 |
| Minerals and vitamins 4 | 2.00 | 2.00 | 2.00 |
| Sodium bentonite | 1.30 | 1.30 | 1.30 |
| Chemical composition | |||
| Dry matter (DM) | 91.0 | 91.0 | 91.0 |
| Crude protein (CP) | 18.6 | 18.6 | 18.6 |
| Ether extract (EE) | 6.60 | 6.60 | 6.60 |
| Neutral detergent fiber (NDF) | 12.6 | 12.7 | 12.8 |
| Non-fiber carbohydrate 5 | 54.2 | 54.3 | 54.4 |
| Ash | 9.10 | 8.90 | 8.70 |
| Diets 1 | |||
|---|---|---|---|
| Name | PO | PFO | FO |
| C12:0 | 0.00 | 1.00 | 2.10 |
| C14:0 | 0.70 | 0.90 | 1.20 |
| C16:0 | 44.2 | 31.1 | 17.2 |
| C16:1 | 0.00 | 1.30 | 2.70 |
| C18:0 | 3.70 | 6.30 | 8.90 |
| C18:1 | 18.1 | 20.6 | 23.3 |
| C18:2 | 29.4 | 30.7 | 32.3 |
| C18:3 | 1.90 | 3.20 | 4.50 |
| C20:5 + C22:6 | 0.0 | 3.60 | 7.30 |
| Other FA | 2.00 | 1.20 | 0.40 |
| Total FA | 100 | 100 | 100 |
| Total n-3 FA | 1.90 | 6.80 | 11.8 |
| n-6/n-3 | 15.5 | 4.50 | 2.70 |
| Diet 1 | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Item | PO | PFO | FO | SEM | Diet | Linear | Quadratic | Diet × Time | Time |
| Body length, cm | |||||||||
| (days 3–70) | 89.6 | 86.9 | 89.3 | 2.3 | 0.44 | 0.93 | 0.21 | 0.41 | <0.0001 |
| (day 3) | 77.9 | 76.1 | 77.6 | 4.1 | 0.65 | 0.25 | 0.21 | ||
| (day 43) | 90.7 a | 79.8 b | 87.9 ab | 4.2 | 0.02 | 0.30 | 0.24 | ||
| (day 56) | 92.0 | 93.3 | 93.5 | 4.1 | 0.74 | 0.93 | 0.21 | ||
| (day 70) | 97.7 | 98.3 | 98.3 | 4.1 | 0.96 | 0.30 | 0.22 | ||
| Body girth, cm | |||||||||
| (days 3–70) | 104.4 | 101.0 | 104.5 | 2.4 | 0.26 | 0.97 | 0.11 | 0.37 | <0.0001 |
| (day 3) | 78.7 | 77.4 | 78.4 | 4.7 | 0.78 | 0.16 | 0.11 | ||
| (day 43) | 102.3 a | 91.1 b | 104.3 a | 4.8 | 0.03 | 0.16 | 0.12 | ||
| (day 56) | 112.7 | 111.8 | 112.7 | 4.7 | 1.00 | 0.97 | 0.11 | ||
| (day 70) | 124.0 | 123.6 | 122.6 | 4.7 | 0.83 | 0.16 | 0.11 | ||
| Withers height, cm | |||||||||
| (days 3–70) | 85.5 | 82.4 | 85.0 | 2.0 | 0.26 | 0.78 | 0.11 | 0.33 | <0.0001 |
| (day 3) | 77.2 | 76.6 | 77.2 | 3.6 | 0.86 | 0.13 | 0.11 | ||
| (day 43) | 86.2 a | 75.2 b | 83.4 a | 3.6 | 0.03 | 0.22 | 0.15 | ||
| (day 56) | 87.5 | 86.6 | 87.9 | 3.6 | 0.72 | 0.78 | 0.11 | ||
| (day 70) | 91.3 | 91.3 | 91.3 | 3.6 | 0.99 | 0.22 | 0.13 | ||
| Heart girth, cm | |||||||||
| (days 3–70) | 92.1 | 89.3 | 92.6 | 2.1 | 0.24 | 0.84 | 0.098 | 0.73 | <0.0001 |
| (day 3) | 77.4 | 76.9 | 76.9 | 4.0 | 0.89 | 0.17 | 0.10 | ||
| (day 43) | 90.7 ab | 82.7 a | 91.8 b | 4.1 | 0.03 | 0.13 | 0.10 | ||
| (day 56) | 97.1 | 95.4 | 97.9 | 4.0 | 0.53 | 0.84 | 0.10 | ||
| (day 70) | 103.4 | 102.2 | 103.7 | 4.0 | 0.71 | 0.13 | 0.10 | ||
| Hip height, cm | |||||||||
| (days 3–70) | 89.4 | 86.4 | 89.4 | 2.0 | 0.24 | 0.99 | 0.10 | 0.43 | <0.0001 |
| (day 3) | 80.8 | 79.8 | 81.2 | 3.8 | 0.79 | 0.14 | 0.10 | ||
| (day 43) | 88.9 a | 78.9 b | 88.5 a | 3.9 | 0.01 | 0.15 | 0.11 | ||
| (day 56) | 92.7 | 91.2 | 92.7 | 3.8 | 0.69 | 1.00 | 0.10 | ||
| (day 70) | 95.1 | 95.7 | 95.3 | 3.8 | 0.97 | 0.15 | 0.10 | ||
| Hip width, cm | |||||||||
| (days 3–70) | 20.0 | 20.2 | 20.2 | 0.3 | 0.78 | 0.51 | 0.83 | 0.53 | <0.0001 |
| (day 3) | 17.3 | 17.3 | 17.1 | 0.4 | 0.98 | 0.60 | 0.83 | ||
| (day 43) | 19.6 | 19.6 | 19.8 | 0.4 | 0.85 | 0.88 | 0.98 | ||
| (day 56) | 21.1 | 21.2 | 21.2 | 0.4 | 0.80 | 0.51 | 0.83 | ||
| (day 70) | 22.2 | 22.7 | 22.8 | 0.4 | 0.80 | 0.88 | 0.91 | ||
| Diets 1 | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Item | PO | PFO | FO | SEM | Diet | Linear | Quadratic | Diet × Time | Time |
| Rumen pH | |||||||||
| Entire period | 5.98 | 6.05 | 5.98 | 0.19 | 0.91 | 0.98 | 0.67 | 0.38 | 0.024 |
| Pre-weaning period (day 42) | 5.96 | 5.84 | 5.80 | 0.24 | 0.62 | 0.72 | 0.67 | ||
| Post-weaning period (day 70) | 6.01 | 6.27 | 6.16 | 0.24 | 0.53 | 0.70 | 0.67 | ||
| Total SCFA, mM | |||||||||
| Entire period | 38.5 | 39.8 | 40.3 | 4.7 | 0.93 | 0.71 | 0.91 | 0.56 | 0.009 |
| Pre-weaning period (day 42) | 35.8 | 32.4 | 35.4 | 6.5 | 0.95 | 0.78 | 0.91 | ||
| Post-weaning period (day 70) | 41.3 | 47.3 | 45.2 | 6.3 | 0.74 | 0.93 | 0.100 | ||
| Acetate, mol/100 mol SCFA | |||||||||
| Entire period | 47.9 | 48.4 | 48.4 | 1.3 | 0.92 | 0.73 | 0.84 | 0.77 | 0.78 |
| Pre-weaning period (day 42) | 48.0 | 48.5 | 47.8 | 1.8 | 0.77 | 0.73 | 0.84 | 0.84 | |
| Post-weaning period (day 70) | 47.8 | 48.2 | 49.0 | 1.7 | 0.51 | 1.00 | 0.92 | 0.92 | |
| Propionate, mol/100 mol SCFA | |||||||||
| Entire period | 33.6 | 33.2 | 31.9 | 1.3 | 0.43 | 0.22 | 0.68 | 0.69 | 0.0008 |
| Pre-weaning period (day 42) | 31.6 | 30.7 | 30.5 | 2.0 | 0.61 | 0.78 | 0.68 | ||
| Post-weaning period (day 70) | 35.6 | 35.7 | 33.2 | 1.9 | 0.96 | 0.33 | 0.46 | ||
| Butyrate, mol/100 mol SCFA | |||||||||
| Entire period | 10.4 | 10.3 | 11.3 | 0.8 | 0.42 | 0.28 | 0.47 | 0.95 | <0.0001 |
| Pre-weaning period (day 42) | 11. 7 | 11.7 | 12.8 | 1.1 | 0.94 | 0.96 | 0.47 | ||
| Post-weaning period (day 70) | 9.1 | 8.9 | 9.8 | 1.0 | 0.42 | 0.24 | 0.32 | ||
| Valerate, mol/100 mol SCFA | |||||||||
| Entire period | 5.06 | 5.05 | 5.24 | 0.44 | 0.87 | 0.66 | 0.79 | 0.80 | 0.047 |
| Pre-weaning period (day 42) | 5.62 | 5.44 | 5.50 | 0.70 | 0.87 | 0.99 | 0.79 | ||
| Post-weaning period (day 70) | 4.49 | 4.65 | 4.98 | 0.72 | 0.80 | 0.65 | 0.70 | ||
| Isovalerate, mol/100 mol SCFA | |||||||||
| Entire period | 3.08 | 3.04 | 3.18 | 0.31 | 0.90 | 0.79 | 0.72 | 0.47 | 0.029 |
| Pre-weaning period (day 42) | 3.22 | 3.61 | 3.44 | 0.51 | 0.72 | 0.86 | 0.72 | ||
| Post-weaning period (day 70) | 2.94 | 2.48 | 2.94 | 0.50 | 0.34 | 0.65 | 0.67 | ||
| Acetate/propionate ratio | |||||||||
| Entire period | 1.45 | 1.48 | 1.64 | 0.16 | 0.48 | 0.26 | 0.68 | 0.93 | 0.085 |
| Pre-weaning period (day 42) | 1.53 | 1.60 | 1.79 | 0.24 | 0.78 | 0.82 | 0.68 | ||
| Post-weaning period (day 70) | 1.36 | 1.37 | 1.49 | 0.23 | 0.97 | 0.35 | 0.47 | ||
| Diet 1 | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Item | PO | PFO | FO | SEM | Diet | Linear | Quadratic | Diet × Time | Time |
| Glucose, mg/dL | |||||||||
| Entire period | 87.4 | 97.0 | 94.1 | 4.3 | 0.084 | 0.12 | 0.100 | 0.73 | <0.001 |
| Pre-weaning period (day 15) | 94.3 | 109.0 | 99.9 | 7.6 | 0.47 | 0.030 | 0.100 | ||
| Pre-weaning period (day 43) | 95.2 | 107.0 | 104.8 | 7.4 | 0.77 | 0.51 | 0.24 | ||
| Post-weaning period (day 71) | 72.7 | 74.9 | 77.7 | 7.4 | 0.71 | 0.12 | 0.100 | ||
| Total protein, g/dL | |||||||||
| Entire period | 6.73 | 6.80 | 6.67 | 0.20 | 0.79 | 0.76 | 0.54 | 0.42 | 0.20 |
| Pre-weaning period (day 15) | 6.70 | 6.91 | 6.43 | 0.26 | 0.42 | 0.70 | 0.54 | ||
| Pre-weaning period (day 42) | 6.74 | 6.72 | 6.64 | 0.25 | 0.94 | 0.49 | 0.50 | ||
| Post-weaning period (day 70) | 6.74 | 6.78 | 6.93 | 0.25 | 0.55 | 0.76 | 0.54 | ||
| Albumin, g/dL | |||||||||
| Entire period | 3.49 | 3.44 | 3.50 | 0.06 | 0.61 | 0.89 | 0.32 | 0.96 | <0.001 |
| Pre-weaning period (day 15) | 3.12 | 3.06 | 3.13 | 0.11 | 0.59 | 0.44 | 0.32 | ||
| Pre-weaning period (day 42) | 3.74 | 3.71 | 3.70 | 0.11 | 0.79 | 0.35 | 0.32 | ||
| Post-weaning period (day 70) | 3.61 | 3.55 | 3.67 | 0.11 | 0.59 | 0.89 | 0.32 | ||
| BHBA, mmol/L | |||||||||
| Entire period | 0.33 | 0.28 | 0.29 | 0.04 | 0.42 | 0.28 | 0.45 | 0.99 | <0.001 |
| Pre-weaning period (day 15) | 0.21 | 0.15 | 0.17 | 0.06 | 0.50 | 0.23 | 0.45 | ||
| Pre-weaning period (day 42) | 0.27 | 0.25 | 0.24 | 0.06 | 0.64 | 0.91 | 0.67 | ||
| Post-weaning period (day 70) | 0.50 | 0.46 | 0.46 | 0.06 | 0.46 | 0.28 | 0.45 | ||
| Total leucocytes, G/L | |||||||||
| Entire period | 12.0 | 10.9 | 11.9 | 1.0 | 0.52 | 0.91 | 0.26 | 0.90 | 0.001 |
| Pre-weaning period (day 15) | 13.8 | 12.6 | 13.1 | 1.5 | 0.65 | 0.30 | 0.26 | ||
| Pre-weaning period (day 42) | 10.4 | 9.3 | 11.2 | 1.5 | 0.56 | 0.36 | 0.29 | ||
| Post-weaning period (day 70) | 11.9 | 10.8 | 11.3 | 1.5 | 0.71 | 0.91 | 0.26 | ||
| Neutrophils, % | |||||||||
| Entire period | 36.8 | 34.3 | 37.2 | 2.9 | 0.56 | 0.90 | 0.29 | 0.71 | 0.19 |
| Pre-weaning period (day 15) | 42.0 | 36.8 | 37.6 | 4.9 | 0.38 | 0.39 | 0.29 | ||
| Pre-weaning period (day 42) | 36.2 | 34.7 | 36.4 | 4.8 | 0.97 | 0.32 | 0.29 | ||
| Post-weaning period (day 70) | 32.2 | 31.4 | 37.5 | 4.8 | 0.80 | 0.90 | 0.29 | ||
| Lymphocytes, % | |||||||||
| Entire period | 57.2 | 60.1 | 56.9 | 2.9 | 0.47 | 0.91 | 0.22 | 0.83 | 0.21 |
| Pre-weaning period (day 15) | 52.7 | 57.9 | 56.2 | 5.0 | 0.72 | 0.32 | 0.22 | ||
| Pre-weaning period (day 42) | 57.3 | 59.3 | 57.3 | 4.8 | 0.68 | 0.26 | 0.23 | ||
| Post-weaning period (day 70) | 61.6 | 63.1 | 57.1 | 4.8 | 0.76 | 0.91 | 0.22 | ||
| Monocytes, % | |||||||||
| Entire period | 3.7 | 3.3 | 3.4 | 0.2 | 0.15 | 0.13 | 0.21 | 0.36 | 0.007 |
| Pre-weaning period (day 15) | 3.2 | 3.0 | 3.2 | 0.3 | 0.90 | 0.07 | 0.21 | ||
| Pre-weaning period (day 42) | 3.8 | 3.5 | 3.7 | 0.3 | 0.74 | 0.74 | 0.43 | ||
| Post-weaning period (day 70) | 4.0 | 3.4 | 3.2 | 0.3 | 0.51 | 0.13 | 0.21 | ||
| Eosinophils, % | |||||||||
| Entire period | 2.4 | 2.4 | 2.6 | 0.1 | 0.38 | 0.20 | 0.60 | 0.11 | 0.19 |
| Pre-weaning period (day 15) | 2.1 | 2.3 | 2.9 | 0.3 | 0.48 | 0.84 | 0.60 | ||
| Pre-weaning period (day 42) | 2.7 | 2.5 | 2.6 | 0.3 | 0.70 | 0.27 | 0.39 | ||
| Post-weaning period (day 70) | 2.3 | 2.4 | 2.2 | 0.3 | 0.70 | 0.20 | 0.60 | ||
| Diet 1 | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Item | PO | PFO | FO | SEM | Diet | Linear | Quadratic | Diet × Time | Time |
| Standing | |||||||||
| Entire period | 89.9 | 91.2 | 78.6 | 7.23 | 0.18 | 0.13 | 0.27 | 0.52 | 0.26 |
| day 40 | 95.0 | 96.0 | 82.5 | 10.0 | 0.92 | 0.85 | 0.27 | ||
| day 47 | 89.5 | 87.0 | 79.0 | 10.0 | 0.80 | 0.092 | 0.14 | ||
| day 61 | 95.5 | 95.5 | 72.5 | 10.0 | 1.00 | 0.13 | 0.27 | ||
| day 68 | 79.5 | 86.5 | 80.5 | 10.0 | 0.55 | 0.091 | 0.19 | ||
| Lying | |||||||||
| Entire period | 90.1 | 88.7 | 101 | 7.23 | 0.18 | 0.14 | 0.27 | 0.52 | 0.26 |
| day 40 | 85.0 | 84.0 | 97.5 | 10.1 | 0.92 | 0.85 | 0.27 | ||
| day 47 | 90.5 | 93.0 | 101 | 10.1 | 0.80 | 0.092 | 0.14 | ||
| day 61 | 84.5 | 84.5 | 107 | 10.1 | 1.00 | 0.13 | 0.27 | ||
| day 68 | 100 | 93.5 | 99.5 | 10.1 | 0.92 | 0.091 | 0.19 | ||
| Eating | |||||||||
| Entire period | 30.9 | 29.9 | 29.1 | 3.63 | 0.89 | 0.63 | 0.97 | 0.22 | <0.0001 |
| day 40 | 15.5 | 15.5 | 21.0 | 6.52 | 1.00 | 0.78 | 0.97 | ||
| day 47 | 28.0 | 31.5 | 34.0 | 6.52 | 0.59 | 0.84 | 0.92 | ||
| day 61 | 46.0 | 38.5 | 30.0 | 6.52 | 0.25 | 0.63 | 0.97 | ||
| day 68 | 34.0 | 34.0 | 31.5 | 6.52 | 0.70 | 0.84 | 0.98 | ||
| Non-nutritional oral behaviors | |||||||||
| Entire period | 43.9 | 40.4 | 32.2 | 5.76 | 0.14 | 0.053 | 0.65 | 0.45 | 0.23 |
| day 40 | 44.0 | 44.5 | 36.0 | 7.85 | 0.95 | 0.55 | 0.65 | ||
| day 47 | 46.0 | 36.0 | 24.5 | 7.85 | 0.21 | 0.17 | 0.32 | ||
| day 61 | 44.0 | 35.5 | 31.5 | 7.85 | 0.61 | 0.052 | 0.65 | ||
| day 68 | 41.5 | 45.5 | 37.0 | 7.85 | 0.57 | 0.17 | 0.46 | ||
| Ruminating | |||||||||
| Entire period | 28.4 | 30.6 | 31.0 | 4.67 | 0.83 | 0.58 | 0.82 | 0.73 | 0.12 |
| day 40 | 28.0 | 37.5 | 33.0 | 7.91 | 0.53 | 0.63 | 0.82 | ||
| day 47 | 36.5 | 32.5 | 34.0 | 7.91 | 0.61 | 0.94 | 0.95 | ||
| day 61 | 20.5 | 24.5 | 31.0 | 7.91 | 0.41 | 0.58 | 0.82 | ||
| day 68 | 28.5 | 28.0 | 26.0 | 7.91 | 0.80 | 0.94 | 0.89 | ||
| Idle | |||||||||
| Entire period | 76.9 | 79.1 | 87.6 | 5.67 | 0.15 | 0.068 | 0.53 | 0.71 | 0.33 |
| day 40 | 92.5 | 82.5 | 90.0 | 11.0 | 0.82 | 0.69 | 0.53 | ||
| day 47 | 69.5 | 80.0 | 87.5 | 11.0 | 0.49 | 0.14 | 0.26 | ||
| day 61 | 69.5 | 81.5 | 87.5 | 11.0 | 0.58 | 0.071 | 0.53 | ||
| day 68 | 76.0 | 72.5 | 85.5 | 11.0 | 0.75 | 0.14 | 0.37 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosseini, S.H.; Mirzaei-Alamouti, H.; Mansouryar, M.; Vazirigohar, M.; Rezamand, P.; Mahjoubi, E.; Ramezankhani, J.; Aschenbach, J.R. Response to Incremental Replacement of Palm Oil with Fish Oil in Starter Diet on Growth Performance, Plasma Metabolites, Ruminal Fermentation, and Behavior of Dairy Calves. Animals 2024, 14, 1061. https://doi.org/10.3390/ani14071061
Hosseini SH, Mirzaei-Alamouti H, Mansouryar M, Vazirigohar M, Rezamand P, Mahjoubi E, Ramezankhani J, Aschenbach JR. Response to Incremental Replacement of Palm Oil with Fish Oil in Starter Diet on Growth Performance, Plasma Metabolites, Ruminal Fermentation, and Behavior of Dairy Calves. Animals. 2024; 14(7):1061. https://doi.org/10.3390/ani14071061
Chicago/Turabian StyleHosseini, Seyed Hadi, Hamidreza Mirzaei-Alamouti, Morteza Mansouryar, Mina Vazirigohar, Pedram Rezamand, Ehsan Mahjoubi, Jafar Ramezankhani, and Jörg R. Aschenbach. 2024. "Response to Incremental Replacement of Palm Oil with Fish Oil in Starter Diet on Growth Performance, Plasma Metabolites, Ruminal Fermentation, and Behavior of Dairy Calves" Animals 14, no. 7: 1061. https://doi.org/10.3390/ani14071061
APA StyleHosseini, S. H., Mirzaei-Alamouti, H., Mansouryar, M., Vazirigohar, M., Rezamand, P., Mahjoubi, E., Ramezankhani, J., & Aschenbach, J. R. (2024). Response to Incremental Replacement of Palm Oil with Fish Oil in Starter Diet on Growth Performance, Plasma Metabolites, Ruminal Fermentation, and Behavior of Dairy Calves. Animals, 14(7), 1061. https://doi.org/10.3390/ani14071061