
Methane Emission, Carbon Footprint and Productivity of Specialized Dairy Cows Supplemented with Bitter Cassava (Manihot esculenta Crantz)

 , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment Location
2.2. Treatments Evaluated/Assessed
2.3. Animal Characteristics
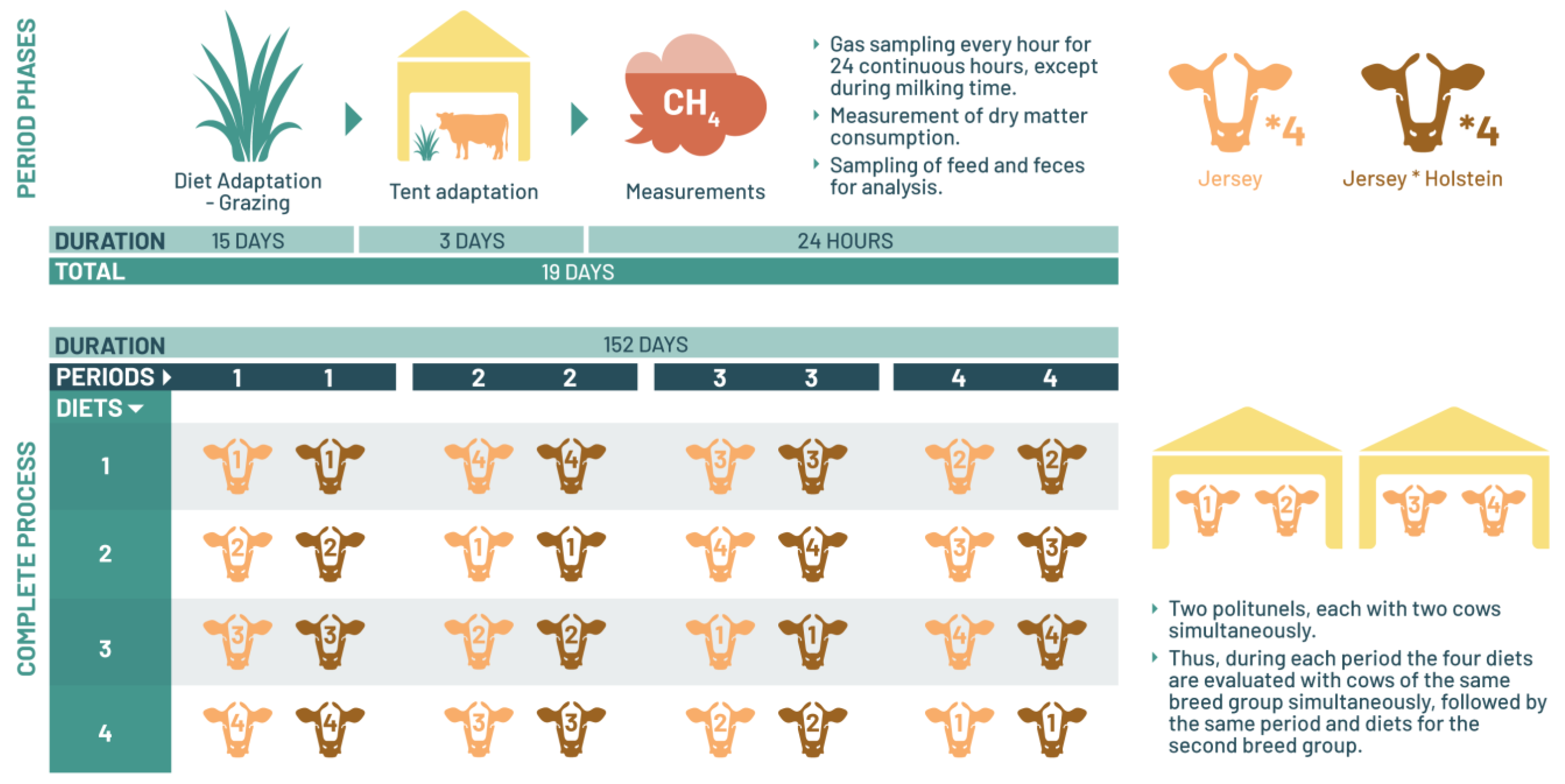
2.4. Experimental Design and Animal Management
2.5. Nutritional Quality of the Treatments
2.6. Polytunnel Conditions and Quantification of Methane Emissions
2.7. Dry Matter and Nutrient Consumption
2.8. Milk Production and Quality
2.9. Carbon Footprint Estimation/Estimate
2.10. Data Analysis
3. Results
3.1. Nutritional Quality of the Components and the Treatments Evaluated
3.2. Nutrient and Energy Intake
3.3. Milk Production and Quality
3.4. Enteric Methane Emissions
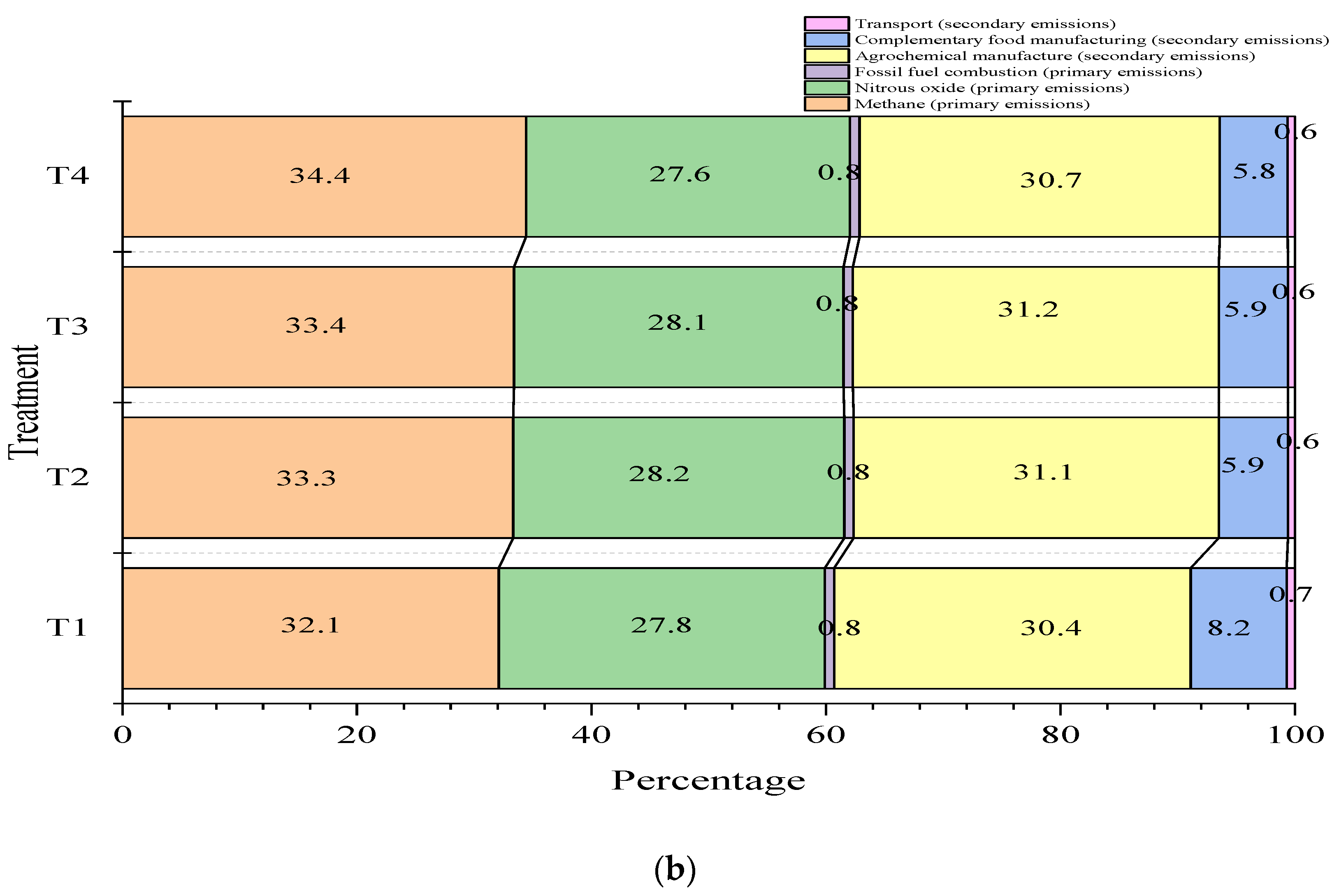
3.5. Carbon Footprint
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Global Warming of 1.5 °C: IPCC Special Report on Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels in Context of Strengthening Response to Climate Change, Sustainable Development, and Efforts to Eradicate Poverty, 1st ed.; Cambridge University Press: Cambridge, UK, 2022; ISBN 978-1-00-915794-0. [Google Scholar]
- Naranjo Ramírez, J.F. En defensa del metano (de las vacas). CES Med. Vet. Zootec. 2019, 14, 80–86. [Google Scholar] [CrossRef]
- Almeida, A.K.; Hegarty, R.S.; Cowie, A. Meta-Analysis Quantifying the Potential of Dietary Additives and Rumen Modifiers for Methane Mitigation in Ruminant Production Systems. Anim. Nutr. 2021, 7, 1219–1230. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty Years of Research on Rumen Methanogenesis: Lessons Learned and Future Challenges for Mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef] [PubMed]
- González-Quintero, R.; van Wijk, M.T.; Ruden, A.; Gómez, M.; Pantevez, H.; Castro-Llanos, F.; Notenbaert, A.; Arango, J. Yield gap analysis to identify attainable milk and meat productivities and the potential for greenhouse gas emissions mitigation in cattle systems of Colombia. Agric. Syst. 2022, 195, 103303. [Google Scholar] [CrossRef]
- Gaillac, R.; Marbach, S. The Carbon Footprint of Meat and Dairy Proteins: A Practical Perspective to Guide Low Carbon Footprint Dietary Choices. J. Clean. Prod. 2021, 321, 128766. [Google Scholar] [CrossRef]
- Mazzetto, A.M.; Falconer, S.; Ledgard, S. Mapping the Carbon Footprint of Milk Production from Cattle: A Systematic Review. J. Dairy Sci. 2022, 105, 9713–9725. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, D.; Capper, J.L.; Garnsworthy, P.C.; Grainger, C.; Shalloo, L. A Case Study of the Carbon Footprint of Milk from High-Performing Confinement and Grass-Based Dairy Farms. J. Dairy Sci. 2014, 97, 1835–1851. [Google Scholar] [CrossRef] [PubMed]
- Jayasundara, S.; Worden, D.; Weersink, A.; Wright, T.; VanderZaag, A.; Gordon, R.; Wagner-Riddle, C. Improving Farm Profitability Also Reduces the Carbon Footprint of Milk Production in Intensive Dairy Production Systems. J. Clean. Prod. 2019, 229, 1018–1028. [Google Scholar] [CrossRef]
- Arango, J.; Ruden, A.; Martinez-Baron, D.; Loboguerrero, A.M.; Berndt, A.; Chacón, M.; Torres, C.F.; Oyhantcabal, W.; Gomez, C.A.; Ricci, P.; et al. Ambition Meets Reality: Achieving GHG Emission Reduction Targets in the Livestock Sector of Latin America. Front. Sustain. Food Syst. 2020, 4, 65. [Google Scholar] [CrossRef]
- Ku-Vera, J.C.; Castelán-Ortega, O.A.; Galindo-Maldonado, F.A.; Arango, J.; Chirinda, N.; Jiménez-Ocampo, R.; Valencia-Salazar, S.S.; Flores-Santiago, E.J.; Montoya-Flores, M.D.; Molina-Botero, I.C.; et al. Review: Strategies for Enteric Methane Mitigation in Cattle Fed Tropical Forages. Animal 2020, 14, s453–s463. [Google Scholar] [CrossRef]
- Vargas, J.; Ungerfeld, E.; Muñoz, C.; DiLorenzo, N. Feeding Strategies to Mitigate Enteric Methane Emission from Ruminants in Grassland Systems. Animals 2022, 12, 1132. [Google Scholar] [CrossRef] [PubMed]
- de Souza Congio, G.F.; Bannink, A.; Mayorga Mogollón, O.L.; Hristov, A.N.; Jaurena, G.; Gonda, H.; Gere, J.I.; Cerón-Cucchi, M.E.; Ortiz-Chura, A.; Tieri, M.P.; et al. Enteric Methane Mitigation Strategies for Ruminant Livestock Systems in the Latin America and Caribbean Region: A Meta-Analysis. J. Clean. Prod. 2021, 312, 127693. [Google Scholar] [CrossRef]
- Vargas, A.C. El sector lechero de pequeña escala en el Departamento de Caldas: Estado actual, retos y modelo para la asistencia técnica universitaria. Universidad de Caldas. Rural. Soc. Prod. Environ. 2019, 19, 165–170. [Google Scholar]
- Cheng, M.; McCarl, B.; Fei, C. Climate Change and Livestock Production: A Literature Review. Atmosphere 2022, 13, 140. [Google Scholar] [CrossRef]
- Wanapat, M.; Kang, S. Cassava Chip (Manihot esculenta Crantz) as an Energy Source for Ruminant Feeding. Anim. Nutr. 2015, 1, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Vera Arteaga, T.A.; Zambrano Vélez, M.I.; Muñoz Murillo, J.P. Raciones suplementarias con follaje de yuca (Manihot esculenta Crantz) en la alimentación de vacas lecheras brown swiss. Sci. Rev. Prod. Cienc. E Investig. 2019, 3, 10–15. [Google Scholar] [CrossRef]
- Leguizamón, A.J.; Rompato, K.M.; Hoyos, R.E.; Audisio, M.C. Nutritional Evaluation of Three Varieties of Cassava Leaves (Manihot esculenta Crantz) Grown in Formosa, Argentina. J. Food Compos. Anal. 2021, 101, 103986. [Google Scholar] [CrossRef]
- Suharti, S.; Oktafiani, H.; Sudarman, A.; Baik, M.; Wiryawan, K.G. Effect of Cyanide-Degrading Bacteria Inoculation on Performance, Rumen Fermentation Characteristics of Sheep Fed Bitter Cassava (Manihot esculenta Crantz) Leaf Meal. Ann. Agric. Sci. 2021, 66, 131–136. [Google Scholar] [CrossRef]
- Granum, G.; Wanapat, M.; Pakdee, P.; Wachirapakorn, C.; Toburan, W. A Comparative Study on the Effect of Cassava Hay Supplementation in Swamp Buffaloes (Bubalus Bubalis) and Cattle (Bos Indicus). Asian-Australas. J. Anim. Sci. 2007, 20, 1389–1396. [Google Scholar] [CrossRef]
- Hue, K.T.; Van, D.T.T.; Ledin, I.; Wredle, E.; Spörndly, E. Effect of Harvesting Frequency, Variety and Leaf Maturity on Nutrient Composition, Hydrogen Cyanide Content and Cassava Foliage Yield. Asian-Australas. J. Anim. Sci. 2012, 25, 1691–1700. [Google Scholar] [CrossRef]
- Ministerio de Agricultura y Desarrollo Rural de Colombia. Reporte: Área, Producción y rendimiento Nacional del cultivo de Yuca. Available online: https://www.agronet.gov.co/estadistica/Paginas/home.aspx?cod=1 (accessed on 12 December 2023).
- Lassen, J.; Difford, G.F. Review: Genetic and Genomic Selection as a Methane Mitigation Strategy in Dairy Cattle. Animal 2020, 14, s473–s483. [Google Scholar] [CrossRef] [PubMed]
- Maciel, I.C.D.F.; Barbosa, F.A.; Tomich, T.R.; Ribeiro, L.G.P.; Alvarenga, R.C.; Lopes, L.S.; Malacco, V.M.R.; Rowntree, J.E.; Thompson, L.R.; Lana, Â.M.Q. Could the Breed Composition Improve Performance and Change the Enteric Methane Emissions from Beef Cattle in a Tropical Intensive Production System? PLoS ONE 2019, 14, e0220247. [Google Scholar] [CrossRef]
- Instituto Geográfico Agustín Codazzi (IGAC). Estudio General de Suelos y Zonificación de Tierras Departamento de Antioquia; Imprenta Nacional de Colombia: Bogotá, Colombia, 2007; Tomos 1–3; 992p. [Google Scholar]
- ISO 6496; Animal Feeding Stuffs—Determination of Moisture and Other Volatile Matter Content. International Organization for Standardization (ISO): Geneva, Switzerland, 1999.
- AOAC. Determination of ash in animal feed: AOAC official method 942.05 revisited. In Official Methods of Analysis, 18th ed.; Oxford University Press: Gaithersburg, MD, USA, 2005; p. 1397. [Google Scholar] [CrossRef]
- AOAC. Protein (Crude) Determination in Animal Feed: Copper Catalyst Kjeldahl Method 984.13. In Official Methods of Analysis, 15th ed.; Oxford University Press: Gaithersburg, MD, USA, 1990. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- ISO 9831:1998; Animal Feeding Stuffs, Animal Products, and Faeces or Urine—Determination of Gross Calorific Value Bomb Calorimeter Method. International Organization for Standardization (ISO): Geneva, Switzerland, 1998.
- Mestres, C. Gelification d’amidons de Mais Modifies Thermiquement. Application a La Fabrication de Pates Alimentaires sans Gluten. Ph.D. Thesis, Université de Nantes, Montpellier, France, 1982. [Google Scholar]
- Nennich, T.D.; Harrison, J.H.; VanWieringen, L.M.; Meyer, D.; Heinrichs, A.J.; Weiss, W.P.; St-Pierre, N.R.; Kincaid, R.L.; Davidson, D.L.; Block, E. Prediction of Manure and Nutrient Excretion from Dairy Cattle. J. Dairy Sci. 2005, 88, 3721–3733. [Google Scholar] [CrossRef]
- Lockyer, D. Methane Emissions from Grazing Sheep and Calves. Agric. Ecosyst. Environ. 1997, 66, 11–18. [Google Scholar] [CrossRef]
- Murray, P.J.; Chadwick, D.C.; Newbold, C.J.; Lockyer, D.R. Measurement of Methane from Grazing Animals—The Tunnel Method. In Measuring Methane Production from Ruminants; Makkar, H.P.S., Vercoe, P.E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 105–109. ISBN 978-1-4020-6132-5. [Google Scholar]
- Hill, J.; McSweeney, C.; Wright, A.-D.G.; Bishop-Hurley, G.; Kalantar-zadeh, K. Measuring Methane Production from Ruminants. Trends Biotechnol. 2016, 34, 26–35. [Google Scholar] [CrossRef]
- ISO 14637:2004; Determination of Urea Content. Enzymatic Method Using Difference in pH (Reference Method). International Organization for Standardization (ISO): Geneva, Switzerland, 2004.
- Tyrrell, H.F.; Reid, J.T. Prediction of the Energy Value of Cow’s Milk. J. Dairy Sci. 1965, 48, 1215–1223. [Google Scholar] [CrossRef]
- Gavrilova, O.; Leip, A.; Dong, H.; MacDonald, J.D.; Gomez Bravo, C.A.; Amon, B.; Barahona Rosales, R.; Prado, A.D.; de Lima, M.A.; Oyhantcabal, W.; et al. Emissions from livestock and manure management. In 2019 Refinement to the 2006 Guidelines for National Greenhouse Gas Inventories. Agriculture, Forestry and Other Land Use; IPCC: Geneve, Switzerland, 2019; Chapter 10; Volume 4. [Google Scholar]
- Hergoualc’h, K.; Akiyama, H.; Bernoux, M.; Chirinda, N.; Prado, A.D.; Kasimir, Å.; MacDonald, J.D.; Ogle, S.M.; Regina, K.; Weerden, T.J.V.D. N2O emissions from managed soils, and CO2 emissions from lime and urea application. In 2019 Refinement to the 2006 Guidelines for National Greenhouse Gas Inventories. Agriculture, Forestry and Other Land Use; IPCC: Geneve, Switzerland, 2019; Chapter 11; Volume 4. [Google Scholar]
- International Dairy Federation (IDF). A common carbon footprint approach for the dairy sector. The IDF guide to standard life cycle assessment methodology. Bull. Int. Dairy Fed. 2015, 479.
- SAS Institute. User’s Guide: Statistics Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2012. [Google Scholar]
- Kusmartono; Retnaningrum, S.; Mashudi; Harper, K.J.; Poppi, D.P. Improving Live Weight Gain of Crossbred Limousin Bulls with Cassava Peel Silage. Animal 2022, 16, 100524. [Google Scholar] [CrossRef]
- Phonethep, P.; Preston, T.R.; Leng, R.A. Effect on feed intake, digestibility, N retention and methane emissions in goats of supplementing foliages of cassava (Manihot esculenta Crantz) and Tithonia diversifolia with water spinach (Ipomoea aquatica). Livest. Res. Rural. Dev. 2016, 28, 72. Available online: http://www.lrrd.org/lrrd28/5/phon28072.html. (accessed on 5 January 2023).
- Cardoso, A.P.; Mirione, E.; Ernesto, M.; Massaza, F.; Cliff, J.; Rezaul Haque, M.; Bradbury, J.H. Processing of Cassava Roots to Remove Cyanogens. J. Food Compos. Anal. 2005, 18, 451–460. [Google Scholar] [CrossRef]
- Kanjanapruthipong, J. The Use of Cassava in Cattle Feeding; Latin American and Caribbean Consortium to Support Cassava Research and Development (CLAYUCA): Cali, Columbia, 1998. [Google Scholar]
- Thang, C.M.; Ledin, I.; Bertilsson, J. Effect of Using Cassava Products to Vary the Level of Energy and Protein in the Diet on Growth and Digestibility in Cattle. Livest. Sci. 2010, 128, 166–172. [Google Scholar] [CrossRef]
- Knowles, M.M.; Pabón, M.L.; Carulla, J.E. Uso de la yuca (Manihot esculenta Crantz) y otras fuentes de almidones no convencionales en la alimentación de rumiantes. Rev. Colomb. Cienc. Pecu. 2012, 25, 488–499. [Google Scholar]
- Li, S.; Cui, Y.; Zhou, Y.; Luo, Z.; Liu, J.; Zhao, M. The Industrial Applications of Cassava: Current Status, Opportunities and Prospects. J. Sci. Food Agric. 2017, 97, 2282–2290. [Google Scholar] [CrossRef]
- Mejía-Agüero, L.E.; Galeno, F.; Hernández-Hernández, O.; Matehus, J.; Tovar, J. Starch Determination, Amylose Content and Susceptibility to in Vitro Amylolysis in Flours from the Roots of 25 Cassava Varieties. J. Sci. Food Agric. 2012, 92, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Anyanwu, C.N.; Ibeto, C.N.; Ezeoha, S.L.; Ogbuagu, N.J. Sustainability of Cassava (Manihot esculenta Crantz) as Industrial Feedstock, Energy and Food Crop in Nigeria. Renew. Energy 2015, 81, 745–752. [Google Scholar] [CrossRef]
- Marsetyo; Sulendre, I.W.; Takdir, M.; Harper, K.J.; Poppi, D.P. Formulating Diets Based on Whole Cassava Tuber. Anim. Prod. Sci. 2021, 61, 761–769. [Google Scholar] [CrossRef]
- Binsulong, B.; Gunha, T.; Kongphitee, K.; Maeda, K.; Sommart, K. Enteric Methane Emissions, Rumen Fermentation Characteristics, and Energetic Efficiency of Holstein Crossbred Bulls Fed Total Mixed Ration Silage with Cassava Instead of Rice Straw. Fermentation 2023, 9, 850. [Google Scholar] [CrossRef]
- Suyitman; Warly, L.; Rachmat, A. Effect of cassava leaf meal supplementation on in vitro digestibility of ammoniated palm leaf enriched with sulfur and phosphorus minerals. Pak. J. Nutr. 2017, 16, 249–252. [Google Scholar] [CrossRef][Green Version]
- Lunsin, R.; Wanapat, M.; Rowlinson, P. Effect of Cassava Hay and Rice Bran Oil Supplementation on Rumen Fermentation, Milk Yield and Milk Composition in Lactating Dairy Cows. Asian-Australas. J. Anim. Sci. 2012, 25, 1364–1373. [Google Scholar] [CrossRef]
- Coffey, E.L.; Horan, B.; Evans, R.D.; Berry, D.P. Milk Production and Fertility Performance of Holstein, Friesian, and Jersey Purebred Cows and Their Respective Crosses in Seasonal-Calving Commercial Farms. J. Dairy Sci. 2016, 99, 5681–5689. [Google Scholar] [CrossRef] [PubMed]
- Hristov, A.N.; Hristova, K.A.; Price, W.J. Relationship between dry matter intake, body weight, and milk yield in dairy cows: A summary of published data. J. Dairy Sci. 2000, 83 (Suppl. S1), 260. [Google Scholar]
- Nascimento, T.V.C.; Oliveira, R.L.; Menezes, D.R.; De Lucena, A.R.F.; Queiroz, M.A.Á.; Lima, A.G.V.O.; Ribeiro, R.D.X.; Bezerra, L.R. Effects of Condensed Tannin-Amended Cassava Silage Blend Diets on Feeding Behavior, Digestibility, Nitrogen Balance, Milk Yield and Milk Composition in Dairy Goats. Animal 2021, 15, 100015. [Google Scholar] [CrossRef] [PubMed]
- Pertiwi, H.; Yulanda Maharsedyo, N.; Amaro, L.; Bhawono Dadi, T. Nutritional Evaluation of Cassava (Manihot esculenta) Peels as a Dietary Supplement in Tropical Friesian Holstein Cross Breed Dairy Cattle. Vet. Med. Int. 2019, 2019, 1–4. [Google Scholar] [CrossRef]
- Henao-Velásquez, A.F.; Múnera-Bedoya, O.D.; Herrera, A.C.; Agudelo-Trujillo, J.H.; Cerón-Muñoz, M.F. Lactose and Milk Urea Nitrogen: Fluctuations during Lactation in Holstein Cows. Rev. Bras. Zootec. 2014, 43, 479–484. [Google Scholar] [CrossRef]
- Sánchez-Salas, J. Nitrógeno ureico en leche: Importancia, determinación y relación con otros componentes lácteos. Nutr. Anim. Trop. 2016, 10, 20. [Google Scholar] [CrossRef][Green Version]
- de Souza Congio, G.F.; Bannink, A.; Mayorga, O.L.; Rodrigues, J.; Bougouin, A.; Kebreab, E.; Silva, R.R.; Maurício, R.M.; da Silva, S.C.; Oliveira, P.P.A.; et al. Prediction of enteric methane production and yield in dairy cattle using a Latin America and Caribbean database. Sci. Total Environ. 2022, 825, 153982. [Google Scholar] [CrossRef]
- Wheatley, C.; Chuzel, G.; Zakhia, N. Cassava—The Nature of the Tuber. In Encyclopedia of Food Science, and Food Technology and Nutrition; Academic Press: San Diego, CA, USA, 2003; pp. 734–743. [Google Scholar]
- Hristov, A.N.; Oh, J.; Firkins, J.L.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Makkar, H.P.S.; Adesogan, A.T.; Yang, W.; Lee, C.; et al. SPECIAL TOPICS—Mitigation of Methane and Nitrous Oxide Emissions from Animal Operations: I. A Review of Enteric Methane Mitigation Options1. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef]
- Mills, J.A.; Dijkstra, J.; Bannink, A.; Cammell, S.B.; Kebreab, E.; France, J. A Mechanistic Model of Whole-Tract Digestion and Methanogenesis in the Lactating Dairy Cow: Model Development, Evaluation, and Application. J. Anim. Sci. 2001, 79, 1584. [Google Scholar] [CrossRef]
- Ogbuji, C.A.; Ogbuji, N.P. Phytochemical, antinutrient and mineral compositions of leaf extracts of some cassava varieties. J. Environ. Sci. Toxic. Food Technol. 2016, 10, 5–8. [Google Scholar]
- Makkar, H.P.S.; Blummel, M.; Becker, K. In vitro effects of and interactions between tannins and saponins and fate of saponins tannins in the rumen, J. Sci. Food Agric. 1995, 69, 481–493. [Google Scholar] [CrossRef]
- Grainger, C.; Clarke, T.; Auldist, M.J.; Beauchemin, K.A.; McGinn, S.M.; Waghorn, G. Potential use of Acacia mearnsii condensed tannins to reduce methane emissions and nitrogen. excretion from grazing dairy cows. Can. J. Anim. Sci. 2009, 89, 241–251. [Google Scholar] [CrossRef]
- Hatew, B.; Podesta, S.C.; Van Laar, H.; Pellikaan, W.F.; Ellis, J.L.; Dijkstra, J.; Bannink, A. Effects of dietary starch content and rate of fermentation on methane production in lactating dairy cows. J. Dairy Sci. 2015, 98, 486–499. [Google Scholar] [CrossRef] [PubMed]
- Barahona, R.; Sánchez, M.S. Limitaciones físicas y químicas de la digestibilidad de pastos tropicales y estrategias para aumentarla. Rev. CORPOICA Cienc. Tecnol. Agrop. 2005, 6, 69–82. [Google Scholar]
- Patra, A.K.; Saxena, J. A new perspective on the use of plant secondary metabolites to inhibit methanogenesis in the rumen. Phytochemistry 2010, 71, 1198–1222. [Google Scholar] [CrossRef]
- Van Wyngaard, J.D.V.; Meeske, R.; Erasmus, L.J. Effect of Dietary Nitrate on Enteric Methane Emissions, Production Performance and Rumen Fermentation of Dairy Cows Grazing Kikuyu-Dominant Pasture during Summer. Anim. Feed Sci. Technol. 2018, 244, 76–87. [Google Scholar] [CrossRef]
- Lemos, D.; Vanessa, G. Evaluación de las Emisiones Entéricas de Metano en Vacas Lecheras Bajo Trópico Alto Con o Sin la Inclusión de Botón de Oro (Tithonia diversifolia). Mater’s Thesis, Universidad Nacional de Colombia, Bogotá, Colombia, 2015. [Google Scholar]
- González-Quintero, R.; Kristensen, T.; Sánchez-Pinzón, M.S.; Bolívar-Vergara, D.M.; Chirinda, N.; Arango, J.; Pantevez, H.; Barahona-Rosales, R.; Knudsen, M.T. Carbon Footprint, Non-Renewable Energy and Land Use of Dual-Purpose Cattle Systems in Colombia Using a Life Cycle Assessment Approach. Livest. Sci. 2021, 244, 104330. [Google Scholar] [CrossRef]
- Uddin, M.E.; Aguirre-Villegas, H.A.; Larson, R.A.; Wattiaux, M.A. Carbon Footprint of Milk from Holstein and Jersey Cows Fed Low or High Forage Diet with Alfalfa Silage or Corn Silage as the Main Forage Source. J. Clean. Prod. 2021, 298, 126720. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Input | Breed: Jersey * Holstein | Breed: Jersey |
|---|---|---|
| Fertilizer 1 a, kg ha−1 yr−1 | 1500 | 1500 |
| Fertilizer 2 b, kg ha−1 yr−1 | 1500 | 1500 |
| Fertilized area, ha | 130 | 70 |
| Petrol, L ha−1 yr−1 | 17.7 | 9.5 |
| Diesel, L ha−1 yr−1 | 17.7 | 9.5 |
| Ingredients | Treatment | |||||||
|---|---|---|---|---|---|---|---|---|
| Items | Cassava Root | Cassava Leaves | Concentrate | Kikuyu | T1 | T2 | T3 | T4 |
| Dry matter (g/kg DM) | 860 | 837 | 872 | 141 | 552 | 569 | 570 | 565 |
| Crude protein (g/kg DM) | 33.6 | 212 | 171 | 154 | 164 | 171 | 140 | 156 |
| Neutral detergent fiber (g/kg DM) | 108 | 587 | 396 | 675 | 518 | 564 | 460 | 503 |
| Acid detergent fiber (g/kg DM) | 92.2 | 471 | 65.3 | 370 | 199 | 248 | 195 | 228 |
| Gross energy (MJ/kg DM) | 15.9 | 19.5 | 18.3 | 17.5 | 17.9 | 17.8 | 17.5 | 17.9 |
| Ashes (g/kg DM) | 49.6 | 76.5 | 77.3 | 115 | 93.9 | 100.1 | 88.1 | 90.6 |
| Starch (g/100 g DM) | 49.0 | -- | 45.9 | -- | -- | -- | -- | -- |
| Item | Breed: Jersey | Breed: Jersey * Holstein | MSE | Significance Level (p) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 | T2 | T3 | T4 | T1 | T2 | T3 | T4 | Treatments | Breed | T*B | ||
| Intake | ||||||||||||
| Dry matter (kg/d) | 8.21 | 8.11 | 7.85 | 7.93 | 8.89 | 8.03 | 8.56 | 8.51 | 0.946 | 0.488 | 0.176 | 0.913 |
| Dry matter (kg/MW) | 9.03 | 8.90 | 8.63 | 8.71 | 8.41 | 7.60 | 8.10 | 8.04 | 0.990 | 0.496 | 0.272 | 0.932 |
| Organic matter (kg/d) | 7.47 | 7.33 | 7.19 | 7.24 | 8.08 | 7.24 | 7.83 | 7.76 | 0.826 | 0.540 | 0.172 | 0.929 |
| Crude protein (g/d) | 1.40 a | 1.44 a | 1.15 b | 1.29 a,b | 1.51 a | 1.44 a | 1.27 b | 1.39 a,b | 0.156 | 0.012 | 0.159 | 0.914 |
| NDF (kg/d) | 4.32 | 4.51 | 3.64 | 4.04 | 4.80 | 4.46 | 4.14 | 4.45 | 0.669 | 0.074 | 0.169 | 0.999 |
| ADF (kg/d) | 1.47 | 1.99 | 1.37 | 1.66 | 1.71 | 1.95 | 1.60 | 1.86 | 0.329 | 0.559 | 0.164 | 0.883 |
| Digestible nutrient intake | ||||||||||||
| Dry matter (kg/d) | 4.49 | 4.42 | 4.25 | 4.31 | 4.92 | 4.37 | 4.71 | 4.68 | 0.613 | 0.787 | 0.189 | 0.8627 |
| Organic Matter (kg/d) | 4.19 | 4.07 | 4.03 | 4.02 | 4.63 | 4.08 | 4.49 | 4.48 | 0.514 | 0.635 | 0.073 | 0.752 |
| Crude protein (g/d) | 748 a | 750 a | 526 b | 642 a,b | 702 a | 691 a | 483 b | 557 a,b | 135.3 | 0.009 | 0.236 | 0.989 |
| NDF (kg/d) | 1.86 a,B | 2.08 a,B | 1.25 b,B | 1.62 a,b,B | 2.33 a,A | 2.18 a,A | 1.72 b,A | 2.04 a,b,A | 0.397 | 0.011 | 0.013 | 0.756 |
| ADF (kg/d) | 0.16 c | 0.54 a | 0.11 c | 0.37 b | 0.27 c | 0.44 a | 0.27 c | 0.38 b | 0.124 | 0.001 | 0.541 | 0.442 |
| Energy consumption (MJ/d) | ||||||||||||
| Gross energy | 149 | 149 | 139 | 149 | 161 | 147 | 152 | 1531 | 16.97 | 0.702 | 0.177 | 0.821 |
| Digestible energy | 76.5 | 77.6 | 70.3 | 70.3 | 82.5 | 75.4 | 76.8 | 75.1 | 10.66 | 0.570 | 0.323 | 0.806 |
| Item | Breed: Jersey | Breed: Jersey * Holstein | MSE | Significance Level (p) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 | T2 | T3 | T4 | T1 | T2 | T3 | T4 | Treatments | Breed | T*B | ||
| Quantity (L/d) | 15.0 B | 12.3 B | 15.5 B | 16.0 B | 18.8 A | 17.3 A | 17.5 A | 17.0 A | 3.093 | 0.528 | 0.014 | 0.583 |
| Fat (kg/d) | 0.62 | 0.57 | 0.67 | 0.69 | 0.75 | 0.83 | 0.70 | 0.68 | 0.167 | 0.998 | 0.087 | 0.637 |
| Protein (kg/d) | 0.47 | 0.39 | 0.50 | 0.50 | 0.56 | 0.52 | 0.55 | 0.51 | 0.103 | 0.358 | 0.053 | 0.405 |
| Solids (kg/d) | 1.09 | 0.95 | 1.18 | 1.19 | 1.31 | 1.35 | 1.25 | 1.19 | 0.259 | 0.977 | 0.062 | 0.512 |
| MUN (mg/dL) | 15.5 | 16.6 | 14.6 | 16.1 | 14.9 | 15.5 | 16.2 | 14.6 | 1.605 | 0.739 | 0.456 | 0.235 |
| ECM (kg/d) | 47.8 B | 40.2 B | 51.2 B | 51.5 B | 57.6 A | 54.9 A | 55.6 A | 52.1A | 9.894 | 0.634 | 0.046 | 0.501 |
| Item | Breed: Jersey | Breed: Jersey * Holstein | MSE | Significance Level (p) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 | T2 | T3 | T4 | T1 | T2 | T3 | T4 | Treatments | Breed | T*B | ||
| Methane (g/d) | 187 B | 193 B | 193B | 205 B | 234A | 232 A | 224 A | 250 A | 32.46 | 0.657 | 0.002 | 0.959 |
| Methane (g/d)/DMI (kg) | 23.5 | 23.8 | 25.2 | 25.7 | 26.4 | 30.5 | 26.4 | 30.0 | 5.276 | 0.693 | 0.055 | 0.763 |
| Methane (g/d)/DMId (kg) | 43.4 | 43.7 | 46.8 | 47.6 | 47.8 | 57.4 | 48.1 | 55.1 | 10.56 | 0.676 | 0.084 | 0.685 |
| Methane (g/d)/OMId (kg) | 46.5 | 47.3 | 49.1 | 50.9 | 50.7 | 60.4 | 50.4 | 57.6 | 10.72 | 0.643 | 0.109 | 0.732 |
| Methane (g/d)/NDFId (kg) | 112 | 93 | 189 | 138 | 102 | 128 | 136 | 151 | 49.58 | 0.103 | 0.842 | 0.362 |
| Methane (g/d)/CPId (kg) | 0.27 b,B | 0.26 b,B | 0.39 a,B | 0.32 a,B | 0.35 b,A | 0.38 b,A | 0.49 a,A | 0.50 a,A | 0.105 | 0.049 | 0.003 | 0.661 |
| Ym (%) | 7.20 | 7.22 | 7.91 | 7.93 | 8.11 | 9.23 | 8.29 | 9.28 | 1.61 | 0.696 | 0.057 | 0.773 |
| Methane (g)/Milk (kg) | 12.7 | 16.7 | 13.1 | 13.5 | 12.8 | 13.7 | 13.4 | 14.7 | 3.857 | 0.614 | 0.803 | 0.727 |
| Methane (g)/ECM kg | 3.97 | 5.13 | 3.96 | 4.30 | 4.19 | 4.35 | 4.23 | 4.83 | 1.21 | 0.625 | 0.874 | 0.713 |
| Jersey * Holstein | Jersey | |||||||
|---|---|---|---|---|---|---|---|---|
| Item | T1 | T2 | T3 | T4 | T1 | T2 | T3 | T4 |
| Carbon footprint (kgCO2eq)/FPCM (kg) | 1.35 | 1.30 | 1.34 | 1.42 | 1.65 | 1.75 | 1.54 | 1.54 |
| Carbon footprint (kgCO2eq)/LWG (kg) | 10.6 | 10.2 | 10.7 | 11.5 | 12.3 | 13.4 | 11.4 | 11.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molina-Botero, I.C.; Gaviria-Uribe, X.; Rios-Betancur, J.P.; Medina-Campuzano, M.; Toro-Trujillo, M.; González-Quintero, R.; Ospina, B.; Arango, J. Methane Emission, Carbon Footprint and Productivity of Specialized Dairy Cows Supplemented with Bitter Cassava (Manihot esculenta Crantz). Animals 2024, 14, 19. https://doi.org/10.3390/ani14010019
Molina-Botero IC, Gaviria-Uribe X, Rios-Betancur JP, Medina-Campuzano M, Toro-Trujillo M, González-Quintero R, Ospina B, Arango J. Methane Emission, Carbon Footprint and Productivity of Specialized Dairy Cows Supplemented with Bitter Cassava (Manihot esculenta Crantz). Animals. 2024; 14(1):19. https://doi.org/10.3390/ani14010019
Chicago/Turabian StyleMolina-Botero, Isabel Cristina, Xiomara Gaviria-Uribe, Juan Pablo Rios-Betancur, Manuela Medina-Campuzano, Mercedes Toro-Trujillo, Ricardo González-Quintero, Bernardo Ospina, and Jacobo Arango. 2024. "Methane Emission, Carbon Footprint and Productivity of Specialized Dairy Cows Supplemented with Bitter Cassava (Manihot esculenta Crantz)" Animals 14, no. 1: 19. https://doi.org/10.3390/ani14010019
APA StyleMolina-Botero, I. C., Gaviria-Uribe, X., Rios-Betancur, J. P., Medina-Campuzano, M., Toro-Trujillo, M., González-Quintero, R., Ospina, B., & Arango, J. (2024). Methane Emission, Carbon Footprint and Productivity of Specialized Dairy Cows Supplemented with Bitter Cassava (Manihot esculenta Crantz). Animals, 14(1), 19. https://doi.org/10.3390/ani14010019