Simple Summary
Feeding approaches to increase production efficiency using new feed additives are essential for sustainable poultry production. This study aimed to evaluate the dietary addition of microbial muramidase (MMUR) on broiler chickens’ nutritional and health aspects. The study suggested that MMUR could enhance the chicken growth rate and improve their antioxidant, inflammatory, and anti-inflammatory responses.
Abstract
The impact of microbial muramidase (MMUR) addition to broiler chicken rations was evaluated through growth parameters, liver histoarchitecture, antioxidant status, biochemical analysis, and expression of pro-inflammatory cytokines for 35 days. Four hundred three-day-old chicks (97.68 ± 0.59 g) were distributed to four distinct groups with ten duplicates each (100 chicks/group) consisting of: group 1 (G1): a basal diet without MMUR (control group); G2: a basal diet + 200 mg MMUR kg−1 G3: a basal diet + 400 mg MMUR kg−1; and G4: a basal diet + 600 mg MMUR kg−1. The results showed that the final body weight and total weight gain were increased (p = 0.015) in birds fed with diets supplemented with MMUR at 600 mg kg−1. The feed conversion ratio (FCR) was improved in all treatment groups compared with the control group. Birds fed with a diet supplemented with 600 mg MMUR kg−1 showed the highest body weight gain and improved FCR. The values of thyroxin hormones and growth hormones were increased in all MMUR-supplemented groups. Dietary MMUR increased the activities of antioxidant enzymes (total antioxidant activity, catalase, and superoxide dismutase) and decreased the activity of malondialdehyde (p < 0.05). In addition, it increased the values of interleukin 1 beta and interferon-gamma compared with the control group. Furthermore, dietary MMUR increased the expression of transforming growth factor-beta immunostaining in the liver and spleen tissues. Our results show that supplementing broilers’ diets with 600 mg MMUR kg−1 could enhance the chicken growth rate and improve their antioxidant, inflammatory, and anti-inflammatory responses.
1. Introduction
The addition of exogenous enzymes to broiler chicken rations is a public practice in nutrition, as it helps in digestion and destroys the antinutritional compounds in the feeds, thus improving their nutritional value and the birds’ performance [1], while also minimizing the feed costs and environmental pollution [2,3]. Almost all market feed enzymes target specific substances in the feed ingredients (e.g., carbohydrates, phytases, and proteases) [2]. However, microbial muramidase (MMUR) is considered one of the enzyme categories that target constituents existing in the lumen of the intestine [4,5].
Muramidase, recognized as N-acetylmuramidase or Lysozyme, is an antimicrobial enzyme that forms a portion of an animal’s innate immune response. It occurs naturally in many body fluids and secretions, including tears, saliva, egg whites, human milk, and gastrointestinal tract (GIT) [6]. Hen egg white contains a high level of Lysozyme enzyme that is thermally stable with a high melting point, reaching 72 °C at pH 5.0 and remaining stable at pH (6–9). It catalyzes the peptidoglycan (PGN) component of the cell envelope of gram-positive microbes by cleaving 1,4-beta-linkages between N-acetylmuramic acid and N-acetyl-D-glucosamine residues, causing lysis of the bacteria [7]. PGN fragments associated with bacterial cell wall lysis are regularly discharged in the gastrointestinal tract and accumulate in the lumen of the intestine [8]. They are recognized as a microbe-associated molecular pattern (MAMP) by immunity cells and activated inflammatory processes [9]. Also, it leads to the production of pro-inflammatory cytokines, antimicrobial peptides, and a protective response to microbes responsible for infection [10,11]. Microbial muramidase hydrolyzes PGN, enhances nutrient absorption, minimizes inflammation, and redirects nutrients for the growth of chickens. The fortification of broilers’ diets with muramidase increases weight gain and improves FCR [7,12]. Also, the supplementation of nursery pigs’ diets with hen egg white muramidase improves the gain rate and the efficiency of feed utilization due to the improvement in small intestine histology [13,14,15] or owing to a strong general immune response [4,5,14]. Muramidase can be a safe feed additive for weaning pigs [13]. Microbial muramidase has recently been investigated for improving intestinal histomorphology, meat quality, immune response, and its hypolipidemic effect in broiler chickens [15]. The current study aimed to evaluate the possible effects of dietary MMUR on the growth parameters, liver histoarchitecture, antioxidant status, biochemical analysis, and the expression of pro-inflammatory cytokines of broiler chickens after a feeding period of 35 days.
2. Material and Methods
2.1. Feed Additive Used
Microbial muramidase, EC Number 3.2.1.17, lysozyme or N-acetylmuramidase, is produced by fermentation with a genetically modified strain of Trichoderma reesei (Accession number DSM 32338), Balancius™, Isando. The enzyme activity in the product is 60,000 LSU(F)/g.
2.2. Birds
This trial was undertaken in the Poultry Research Division of the Faculty of Veterinary Medicine at Zagazig University, Egypt. Four hundred one-day-old male Ross-308 chickens were acquired from a native hatchery. On arrival, they were reared on a broad range antibiotic (neomycin) and a dehydrated solution for three successive days and attained an average weight of 97.68 ± 0.59 g. The chicks were nourished in a logically aired house with sawdust litter. The building temperature was adjusted by heaters and was maintained at 34 °C through the initial week and gradually minimized until it reached 25 °C in the last week. The lighting program was 24 h/day for the first week, then changed to 16 h of light and 8 h of darkness in the period of 7–35 days. All chicks were vaccinated against Gumboro and New Castle diseases and checked daily for health problems.
2.3. Experimental Design and Diets
The chicks were allotted randomly to 4 groups with ten replicates (100 birds/group). The trial groups comprised of G1: a basal diet without MMUR (control group), G2: a basal diet + 200 mg MMUR kg−1, G3: basal diet + 400 mg MMUR kg−1, and G4: basal diet + 600 mg MMUR kg−1, with an enzyme activity of 0, 12,000, 24,000, and 36,000 LSU(F)/kg diet, respectively. The trial was extended to 35 days with water and feeds offered ad libitum. All feeds were crushed and formulated according to the Ross manual guide published by AVIAGEN [16], as represented in Table 1. Dry matter, crude protein, crude fiber, and fat were analyzed in all feedstuffs and rations used by AOAC [17] The Ca and P content of the diets were measured using an Element Analyzer with an Energy Dispersive X-ray fluorescence system, JSX 3222, JEOL, Japan. The amino acid content of the diets was assessed following the methodology described by Simpson et al. [18], using an amino acid analyzer system (Harvard Bioscience, Inc., Holliston, United States).

Table 1.
Proximate and chemical composition of the experimental diets (%).
2.4. Growth Performance
The chicks were weighed separately on the fourth day to determine their initial weights, and the average body weight (BW) from each group was assessed on days 10, 23, and 35. The body weight gain (BWG) was estimated as the difference between the final and the initial weight at each stage interval. The average feed consumption per bird in each replicate was calculated as the difference between the amount of feed offered and the amount of feed left, then divided by the birds’ numbers. The feed conversion ratio (FCR) was determined according to Wagner et al. [19]. FCR = feed intake (g)/body weight gain (g).
2.5. Sample Collection
Ten blood samples were taken from the brachial vein of ten random birds from each group (n =10) on the 35th day in a dry clean tube without anticoagulant and left to clot at room temperature and then centrifuged at 3000 rpm for 5 min to separate the serum. The collected serum was preserved at −20 °C in Eppendorf tubes prior to analysis.
2.6. Clinico-Biochemical Analysis
The serum levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were measured using analytic kits (Spinreact, Santa Coloma, Spain). The glucose levels were assessed with colorimetric investigative kits from Spectrum-Bioscience (Egyptian Company for Biotechnology, Cairo, Egypt), as discussed by [20]. ELISA kits (My Biosource Co. of CAT. NO. MBS269454, MBS265796, MBS266317, and MBS025331) were used for the analysis of triiodothyronine hormone (T3), thyroxin (T4), growth hormone (GH), and leptin.
2.7. Immune and Antioxidant Indices
ELISA kits (Cat. No. MBS2024496) were used to measure the serum interleukin 1beta (IL1β) levels. The activities of Malondialdehyde (MDA) (nmol/mL), total antioxidant capacity (TAC) (U/mL), catalase (CAT) (U/mL), and superoxide dismutase (SOD) (U/mL) were measured in the serum following the methodology of Mcdonald and Hultin [21], Rice-Evans and Miller [22], Aebi [23], and Nishikimi et al. [24], respectively.
2.8. Histological and Immunohistochemical Examination
Three liver specimens from each group (n = 3) were taken and fixated in 10% neutral formalin. Samples were dehydrated with (75–100%) ascending grades of ethanol, located in xylol I and II, and embedded in paraffin. Then, they were sliced into 4 µm slices (cross and longitudinal) by a microtome (Leica RM 2155, England, UK) and stained by hematoxylin and eosin (H and E) [25]. Pictures from each bird in each treatment (25 pictures/group) were taken by an AmScope 5.0 MP microscope digital camera adjusted on (100× and 400× magnification) a high power field. Stained tissues were inspected for pathological changes such as inflammation, degeneration, apoptosis, and necrosis.
The inflammatory response of chickens was explored in the leucocyte populations using transforming growth factor-beta (TGF-β) in the liver and spleen. On the 35th day, samples from the liver and spleen (three samples for each group) were taken to observe the expression of TGF-β, following the methodology of Saber et al. [26]. Tissue segments were nurtured in an endogenous peroxidase-blocking mixture containing hydrogen peroxide and sodium azide (DAKO peroxidase blocking reagent, Cat. No. S2001). Then, one to two drops of primary monoclonal antibody supersensitive against TGF-β were added (Cat. BAF240, Novus Biologicals, Briarwood Avenue, Centennial, CO, USA); after that, the slides were stained by hematoxylin and examined under a microscope. Finally, they were analyzed for different immune-positive cells using three high power fields (HPFs) using Image J 1.49 software bundled with 64-bit Java 1.8.0_172 (National Institutes of Health, MD, USA) [27].
2.9. Statistical Analysis
The results were evaluated by using a one-way analysis of variance (ANOVA) and a SPSS general linear model (SPSS Inc., Chicago, IL, USA) after applying homogeneity and normality among the different groups using Levene and Shapiro–Wilk tests. The linear and quadratic effects of MMUR were exhibited using orthogonal polynomial contrasts. A Tukey test was used to calculate the significance between the mean values and adjusted at a p < 0.05 significance level.
The statistical model is given by:
where Yik = observed value of the response variable, U = observed mean for the response variable, Ti = the fixed effect of the treatment group, Eijk = random error.
Yik = U + Ti + Eijk
3. Results
3.1. Growth Performance
The performance of broilers is shown in Table 2. Throughout the starter period (4th–10th day), the BW, BWG, and FCR were non-significantly different in broilers fed feeds enriched with MMUR at 200 and 400 mg kg−1 compared with the control group. Nevertheless, these parameters were significantly enhanced (linear p = 0.041, 0.041, and 0.033, respectively) by dietary MMUR at a level of 600 mg kg−1. The addition of MMUR at different levels did not affect the feed intake (FI) compared to the control group. During the grower period (11th–23rd day), birds fed on the 600 mg MMUR kg−1 diet showed an increase in BW (p = 0.018) and BWG (p = 0.033) compared to the control group. The FI was indifferent (p = 0.790) among all experimental groups. The MMUR levels at 200 and 600 mg kg−1 showed an improved FCR (linear p = 0.002). Throughout the finisher period (24th–35th day), dietary MMUR at 600 mg kg−1 increased the BW (p = 0.015) without impacting the BWG, FI, and FCR. The overall performance results showed that the final BW and BWG were linearly increased (p = 0.015) in birds fed MMUR-supplemented diets at 600 mg kg−1. The overall FCR decreased (p = 0.004) with MMUR at 600 mg kg−1. The various levels of MMUR addition had no significant effect on the total FI. The ultimate body weight was at its maximum at 600 mg MMUR kg−1, while the smallest weight was detected in the control group.
Table 2.
Effect of MMUR on the growth performance of broiler chicks during the feeding periods.
3.2. Clinic-Biochemical Indices
The MMUR effect on the blood biochemical indices of the broilers is presented in Table 3. The enrichment of diets with different MMUR levels did not influence the serum values of AST, ALT, and uric acid but linearly increased the creatinine level (p = 0.001) compared with the control group. The addition of MMUR at different doses linearly increased the serum levels of T3 (p = 0.001), T4 (p < 0.001), and growth hormones (p < 0.001). MMUR supplementation at all levels did not affect glucose levels compared with the control group. The serum leptin linearly increased (p = 0.017) with MMUR addition compared with the control group.
Table 3.
Effect of MMUR on the blood biochemical parameters of broiler chickens.
3.3. Antioxidant and Inflammatory Responses
The effect of MMUR on serum antioxidant enzyme activities and serum inflammatory indices of the chickens are displayed in Table 4. The TAC, CAT, and SOD activities increased linearly (p ≤ 0.001). In contrast, the MDA level decreased either linearly or quadratically in all MMUR-enriched groups compared to the control group. The levels of IL1β and IFN-γ were linearly amplified (p ≤ 0.001) in all MMUR treatments. The highest MMUR supplementation level (600 mg/kg) induced the highest values of these parameters.
Table 4.
Effect of MMUR on antioxidant and pro-inflammatory responses in broiler chickens.
3.4. Histopathological Findings
The inspected liver sections from the various treated groups (0, 200, 400, 600 mg MMUR kg−1) show typical histological characteristics of various structures, including portal area and hepatocytes, which are seen as small masses around the central veins. Several round cells were observed around the hepatic portal area, indicating a natural immune response (Figure 1). Mild to moderate portal lymphoplasmacytic aggregations and biliary proliferation were seen in the control group (Figure 1A). Mildly aggregated immune-responsive lymphoplasmacytic cells were observed for MMUR at levels 400 and 600 mg kg−1 (Figure 1C,D). The aggregated inflammatory cells appear to be an immune surveillance and protective device instead of a destructive inflammatory procedure (Figure 1).
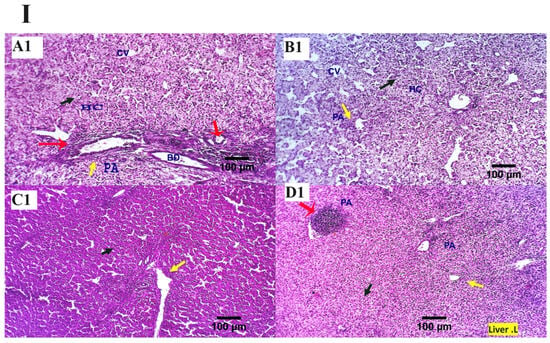
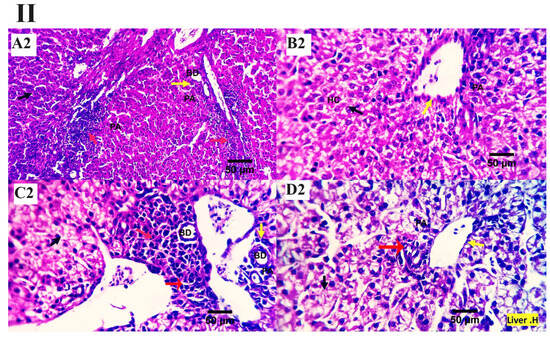
Figure 1.
(I,II) Photomicrographs from the liver ((I): H&E × 100 magnification, (II): H&E × 400 magnification) of various chicken experimental groups, which received 0 mg/kg MMUR (A1,A2), 200 mg/kg MMUR (B1,B2), 400 mg/kg MMUR (C1,C2), and 600 mg/kg MMUR (D1,D2). These show typical histological characteristics of various structures, including portal area (PA, yellow arrow) and hepatocytes (HC, black arrow), which are seen as a small masses around the central veins (CV), with a few round cells observed around the portal area indicating a natural immune response. Mild to moderate portal lymphoplasmacytic aggregations (red arrows) and biliary proliferations (BD, yellow arrow) are detected in group (A). Mildly aggregated immune-responsive lymphoplasmacytic cells can be observed in groups (C,D) (red arrows).
3.5. TGF-β Immunostaining
The analysis of the examined liver slices exposed an average percentage of positive cells in each of the three high power fields (HPF) to the anti-inflammatory marker (TGF-β) as follows: 0.09, 0.13, 3.96, and 4.00 for dietary MMUR levels of 0, 200, 400, 600 mg kg−1, respectively (Figure 2 and Figure 3). A morphometric analysis of the inspected spleen segments revealed an average percentage of positive cells in each of the three high power fields (HPFs) to the pro-inflammatory marker (TGF-β) in all groups as follows: 1.19, 1.01, 1.66, and 7.21 for dietary MMUR levels of 0, 200, 400, 600 mg kg−1, respectively (Figure 2 and Figure 4).
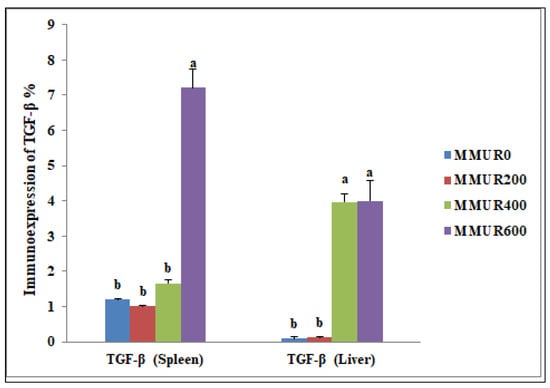
Figure 2.
The morphometric analytic data of the immunostained TGF-β cells in the spleen and liver of various experimental groups. Bars with different letters (a,b) significantly differ at p < 0.05. (MMUR0) group received 0 mg/kg MMUR, (MMUR200) group received 200 mg/kg MMUR, (MMUR400) group received 400 mg/kg MMUR, (MMUR600) group received 600 mg/kg MMUR.
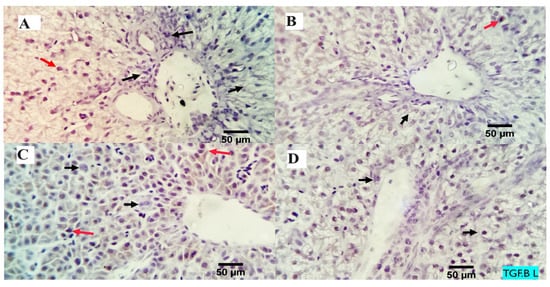
Figure 3.
The immunostained positive TGF-β cells (red arrows) in the liver of different experimental chicken groups. Black arrows: negative cells. ×400 magnification power. (A) group received 0 mg/kg MMUR, (B) group received 200 mg/kg MMUR, (C) group received 400 mg/kg MMUR, (D) group received 600 mg/kg MMUR.
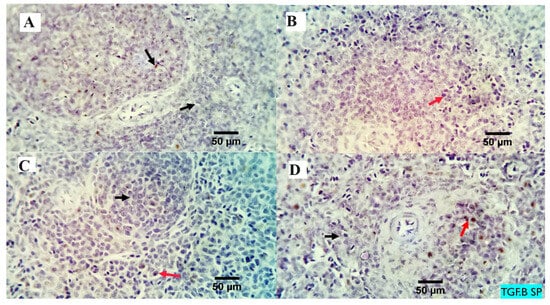
Figure 4.
The immunostained positive TGF beta cells (red arrows) in the spleen of different experimental chicken groups. Black arrows: negative cells. × 400 magnification power. (A) group received 0 mg/kg MMUR, (B) group received 200 mg/kg MMUR, (C) group received 400 mg/kg MMUR, (D) group received 600 mg/kg MMUR.
4. Discussion
The current study aimed to investigate the impact of dietary supplementation of graded levels of MMUR on the growth performance parameters, liver histoarchitecture, antioxidant status, biochemical analysis, and expression of pro-inflammatory cytokines of broiler chickens. The results showed that adding MMUR to the broilers’ diets encourages the growth performance of broilers’ BW, BWG, and FCR without changing their feed consumption. The highest results were in chickens fed on MMUR at a level of 600 mg/kg, and the lowest was in those fed the control group. This improvement in the performance parameters may be attributed to the excellent condition of the chickens and the improved gastrointestinal tract morphology represented by higher villus length, crypt depth, and absorptive area, as reported in our previous study [15], as well as effective gastrointestinal tract function (good digestion and absorption efficacy) [5], and also the increase in intraepithelial lymphocyte count (IEL), goblet cell numbers, and mucus secretion [5]. Mucus acts as a fence against microbes, physical and chemical attacks, as a lubricant of the GIT tract and its constituents, and as a main component of innate immunity which results in a better feed utilization efficiency [28]. The improved performance caused by MMUR may be due to the increase in the apparent ileal digestibility of dry matter, ether extract, and ash, and decreased intestinal permeability [12,29].
Moreover, MMUR increases the digestibility of fat, protein, and phosphorus [7]. It activates other feed enzymes, such as phytase enzymes [7]. Supplementation with MMUR maintains good intestinal tight junctions, prevents translocation of harmful particles from the lumen of the intestine to the bloodstream, and reduces the exposure of the birds to microbes and their compounds, toxins, and antigens [29,30]. Also, it increases the lactobacillus numbers in broilers’ caecum [12]. Its inclusion with lysozyme maintains healthy gut function, enhances the immune reaction, and also enhances production characteristics in pigs [31,32]. It also minimizes the pathogen counts in broilers’ cecum [32,33], improves growth parameters, and augments antioxidant activity and general immunity of broilers [33].
Previous studies reported a positive effect of microbial muramidase 007 additions to broiler diets on FCR and body weight gain [5,7,12]. Also, the inclusion of MUR in broilers at 35,000 LSU (F)/kg [29] or at 45,000 LSU (F)/kg [34] had an improved effect on body growth and FCR in broilers at 1–42 d. Moreover, muramidase from various origins, for example, the addition of 10% modified rice expressing lysozyme [35] and hen egg white lysozyme [33,36], improved the FCR and BWG of the broilers. On the other hand, previous researchers observed a non-significant effect of hen egg white lysozyme or MMUR inclusion on the performance parameters of broilers [15,37] and pigs [38].
Alanine transaminase (ALT) and aspartate transaminase (AST) are intracellular liver enzymes discharged in high quantities into the blood flow after liver damage. In our experiment, fortifying diets with MMUR at different levels did not affect the serum values of liver enzymes (AST and ALT) and uric acid. However, it increased the creatinine level compared with the control without influencing the serum glucose level. These results confirmed the results of liver histopathology where MMUR supplementation did not cause any harmful effects on the hepatic histoarchitecture, which appeared normal in the non-supplemented groups. Microbial muramidase is an antioxidant that guards cells against the destructive effect of free radicals, lessens toxicity, and maintains liver function [39]. Consistent with our data, Lichtenberg et al. [12] reported no effect of MMUR addition on serum AST value in broilers. Also, Schliffka et al. [38] detected no impact of muramidase addition at 65,000, 325,000, and 650,000 LSU (F)/kg of feed on the serum values of AST, ALT, urea, and glucose in swine. Our results showed that the addition of MMUR to broiler diets resulted in increased values of T3, T4, and growth hormones, which were confirmed by earlier results which stated that the addition of exogenous lysozyme enzyme at 50, 100, and 150 mg/kg of rabbit’s diets increased the serum levels of triiodothyronine and thyroxine [40]. These results also explain the improved growth performance of birds by MMUR supplementation due to the beneficial role of these hormones in controlling growth and other metabolic functions [41].
Lysozyme is an essential ingredient in non-specific humoral immunity. It exerts bactericidal activity by hydrolysis of the β-1,4-glycosidic link between N-acetyl muramic acid and N-acetyl glucosamine of microbial wall peptidoglycans, along with activation of the complement system and phagocytic activity leading to the destruction of the glucosidic bonds of Escherichia coli and Staphylococcus walls and the prevention of contamination and disease [42]. Our results showed increased serum levels of IL1β and IFN-γ at all MMUR supplementation levels, in addition to the rise in the immunoexpression of TGF-β in the liver and spleen tissues by MMUR supplementation in a level-dependent way. Transforming growth factor beta (TGF-β) is a multifunctional cytokine secreted by numerous kinds of cells, including macrophages. It contains three mammalian isoforms (TGF-β 1 to 3, HGNC symbols TGFB1, TGFB2, TGFB3) and several other signaling proteins. The first line of immune response is the release of cytokines, which have a parallel action on the metabolism of nutrients. Pro-inflammatory cytokines forward valuable nutrients far from production and towards the immune reply [43,44]. Cooper et al. [45] documented that TGF-β1 was amplified in unaffected pigs consuming a transgenic goats’ milk lysozyme. The histological examination of the hepatic tissues in the current study showed mildly aggregated immune-responsive lymphoplasmacytic cells at MMUR supplementation levels of 400 and 600 mg kg−1. These aggregated inflammatory cells seem to be an immune surveillance and protective device instead of a destructive inflammatory response.
It has been reported that hen egg white lysozyme (HEWL) is highly effective against a wide range of gram-positive microbes, while killing gram-negative microorganisms [46] and minimizing the numbers of C. perfringens, lactobacilli and E. coli in the ileum of chickens [36]. In addition, the transgenic human lysozyme has activity against numerous pathogens of the digestive tract besides Clostridium perfringens [47]. Moreover, it results in the improvement of nonspecific immunity, including the upgrade of the gene appearance of interferon-gamma, interleukin-10 and 18, and the formation of intestinal glutathione peroxidase [4,14,33,37,48]. Our previous study reported linear increases in the serum levels of IL10, complement 3, and lysozyme activity by MMUR supplementation (200–600 mg/kg), in addition to the increase in the immunoexpression of lymphocyte subpopulation biomarkers (CD3 and CD20) in the spleen tissues [15]. Muramidase can split PGN into its ultimate active component, muramyl dipeptide (MDP) [49]. Negroni et al. [50] confirmed that the intracellular nucleotide-binding oligomerization domain-containing protein 2 (NOD2) receptor could sense MDP. Severe stimulation of NOD2 triggers nuclear factor kappa-B cells (NF-κB) and mitogen-activated protein kinase pathways, resulting in cytokine production and a pro-inflammatory response [51].
In the current study, the supplementation of broilers’ diets with MMUR increased the total antioxidant capacity, catalase, and SOD activities while decreasing the level of MDA. The high serum values of TAC, SOD, and GST in broilers fed with MMUR-enriched diets led to the increased ability of the broilers’ antioxidant defense systems to hunt oxidative stress. Exogenous lysozyme is a valuable source of antioxidant compounds that can protect body cells from damage by free radicals and lessen poisonousness [39]. Researchers have previously reported that the enrichment of poultry diets with MMUR has a positive impact on the antioxidant ability of the gastrointestinal tract and increases the gene appearance of intestinal glutathione peroxidase (SOD and GSH-Px) [4,14,33,36,37,48]. Similarly, the addition of the lysozyme enzyme to the diets of rabbits at 50, 100, and 150 mg/kg increased the total antioxidant capacity values, glutathione S-transferase, SOD, and catalase, while decreasing the value of malondialdehyde [52]. Moreover, an increased serum SOD activity and decreased MDA levels of the intestine, accompanied by increased numbers of goblet cells and height of microvilli, were observed following the addition of muramidase to the feeds of juvenile gibel carp [53].
5. Conclusions
The enrichment of broiler chicken diets with MMUR at 600 mg/ kg is recommended in practical poultry feeding due to its low cost (8 USD/kg of product at the time of the experiment, while the cost of 600 mg MMUR was 0.48 Cent) and positive influence on the bird’s productive performance, represented by improved body weight, weight gain, and FCR through increased metabolic hormone levels (T3, T4, and growth hormones). Dietary MMUR had no harmful effects on liver tissues and liver function tests. Moreover, it upgraded the antioxidant activity and prevented lipid peroxidation as indicated through increased TAC, catalase, and SOD activities and reduced MDA levels. Dietary MMUR acts as an immune-enhancing substance through increasing IL1β and IFN-γ levels and TGF-β immunoexpression in the liver and spleen.
Author Contributions
Conceptualization: S.A.A. (Shimaa A. Amer); Formal analysis: S.A.A. (Shimaa A. Amer) and A.E.O.; Methodology: S.A.A. (Shimaa A. Amer), A.E.O., G.I.A.E.-R., A.G., S.A.A. (Samar A. Abdo), A.E., A.S.K., A.-W.A.A.-W. and E.M.Y.; Writing—original draft: S.A.A. (Shimaa A. Amer) and A.E.O.; Writing—review & editing: S.A.A. (Shimaa A. Amer), A.E.O. and S.J.D. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Zagazig University and Researchers Supporting project number (RSP2023R36), King Saud University, Riyadh, Saudi Arabia.
Institutional Review Board Statement
The ethical approval of the experimental protocol was obtained from the Institutional Animal Care and Use Committee of Zagazig University, Egypt (Approval No. ZU-IACUC/2/F/153/2022). All animal experiments were performed based on the recommendations described in “The Guide for the Care and Use of Laboratory Animals in Scientific Investigations”.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated or analyzed during the current study are not publicly available but are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Bedford, M.; Cowieson, A. Exogenous enzymes and their effects on intestinal microbiology. Anim. Feed Sci. Technol. 2012, 173, 76–85. [Google Scholar] [CrossRef]
- Adeola, O.; Cowieson, A. Board-invited review: Opportunities and challenges in using exogenous enzymes to improve nonruminant animal production. J. Anim. Sci. 2011, 89, 3189–3218. [Google Scholar] [CrossRef]
- Meale, S.J.; Beauchemin, K.A.; Hristov, A.N.; Chaves, A.; McAllister, T. Board-invited review: Opportunities and challenges in using exogenous enzymes to improve ruminant production. J. Anim. Sci. 2014, 92, 427–442. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Lin, S.; Zhu, J.; Pang, X.; Fang, Z.; Lin, Y.; Che, L.; Xu, S.; Li, J.; Huang, Y. Effects of dietary lysozyme levels on growth performance, intestinal morphology, non-specific immunity and mRNA expression in weanling piglets. Anim. Sci. J. 2016, 87, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Sais, M.; Barroeta, A.C.; López-Colom, P.; Nofrarías, M.; Majó, N.; Lopez-Ulibarri, R.; Calvo, E.P.; Martín-Orúe, S.M. Evaluation of dietary supplementation of a novel microbial muramidase on gastrointestinal functionality and growth performance in broiler chickens. Poult. Sci. 2020, 99, 235–245. [Google Scholar] [CrossRef]
- Sahoo, N.; Kumar, P.; Bhusan, B.; Bhattacharya, T.; Dayal, S.; Sahoo, M. Lysozyme in livestock: A guide to selection for disease resistance: A review. J. Anim. Sci. Adv 2012, 2, 347–360. [Google Scholar]
- Boroojeni, F.G.; Männer, K.; Rieger, J.; Calvo, E.P.; Zentek, J. Evaluation of a microbial muramidase supplementation on growth performance, apparent ileal digestibility, and intestinal histology of broiler chickens. Poult. Sci. 2019, 98, 2080–2086. [Google Scholar] [CrossRef]
- Lee, W.-J.; Hase, K. Gut microbiota–generated metabolites in animal health and disease. Nat. Chem. Biol. 2014, 10, 416–424. [Google Scholar] [CrossRef]
- Broom, L.J.; Kogut, M.H. Inflammation: Friend or foe for animal production? Poult. Sci. 2018, 97, 510–514. [Google Scholar] [CrossRef]
- Lee, M.S.; Kim, Y.-J. Signaling pathways downstream of pattern-recognition receptors and their cross talk. Annu. Rev. Biochem. 2007, 76, 447–480. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Lichtenberg, J.; Calvo, E.P.; Madsen, K.; Lund, T.Ø.; Birkved, F.K.; van Cauwenberghe, S.; Mourier, M.; Wulf-Andersen, L.; Jansman, A.; Lopez-Ulibarri, R. Safety evaluation of a novel muramidase for feed application. Regul. Toxicol. Pharmacol. 2017, 89, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Oliver, W.; Wells, J. Lysozyme as an alternative to antibiotics improves growth performance and small intestinal morphology in nursery pigs. J. Anim. Sci. 2013, 91, 3129–3136. [Google Scholar] [CrossRef] [PubMed]
- Oliver, W.; Wells, J. Lysozyme as an alternative to growth promoting antibiotics in swine production. J. Anim. Sci. Biotechnol. 2015, 6, 35. [Google Scholar] [CrossRef] [PubMed]
- Amer, S.A.; Farahat, M.; Gouda, A.; Abdel-Wareth, A.A.; Abdel-Warith, A.-W.A.; Younis, E.M.; Elshopakey, G.E.; Baher, W.M.; Saleh, G.K.; Davies, S.J. New Insights into the Effects of Microbial Muramidase Addition in the Diets of Broiler Chickens. Animals 2023, 13, 1356. [Google Scholar] [CrossRef]
- Aviagen, R. Ross Broiler Management Manual, 2009; ROSS: Richmond, VA, USA, 2014; Volume 9, pp. 350–364. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International; AOAC: Rockville, MD, USA, 2000. [Google Scholar]
- Simpson, R.J.; Neuberger, M.R.; Liu, T.-Y. Complete amino acid analysis of proteins from a single hydrolysate. J. Biol. Chem. 1976, 251, 1936–1940. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.; Furrow, R.; Bradley, B. Subchronic toxicity of monensin in broiler chickens. Vet. Pathol. 1983, 20, 353–359. [Google Scholar] [CrossRef]
- Trinder, P. Determination of glucose in blood using glucose oxidase with an alternative oxygen acceptor. Ann. Clin. Biochem. 1969, 6, 24–27. [Google Scholar] [CrossRef]
- Mcdonald, R.E.; Hultin, H.O. Some characteristics of the enzymic lipid peroxidation system in the microsomal fraction of flounder skeletal muscle. J. Food Sci. 1987, 52, 15–21. [Google Scholar] [CrossRef]
- Rice-Evans, C.; Miller, N.J. 241 Total antioxidant status in plasma and body fluids. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1994; Volume 234, pp. 279–293. [Google Scholar]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Bancroft, J.; Layton, C.; Suvarna, S. Bancroft’s Theory and Practice of Histological Techniques; Churchill Livingstone: London, UK, 2013. [Google Scholar]
- Saber, S.; Khalil, R.M.; Abdo, W.S.; Nassif, D.; El-Ahwany, E. Olmesartan ameliorates chemically-induced ulcerative colitis in rats via modulating NFκB and Nrf-2/HO-1 signaling crosstalk. Toxicol. Appl. Pharmacol. 2019, 364, 120–132. [Google Scholar] [CrossRef]
- Rizzardi, A.E.; Johnson, A.T.; Vogel, R.I.; Pambuccian, S.E.; Henriksen, J.; Skubitz, A.P.; Metzger, G.J.; Schmechel, S.C. Quantitative comparison of immunohistochemical staining measured by digital image analysis versus pathologist visual scoring. Diagn. Pathol. 2012, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Deplancke, B.; Gaskins, H.R. Microbial modulation of innate defense: Goblet cells and the intestinal mucus layer. Am. J. Clin. Nutr. 2001, 73, 1131S–1141S. [Google Scholar] [CrossRef] [PubMed]
- Goes, E.C.; Dal Pont, G.C.; Maiorka, A.; Bittencourt, L.C.; Bortoluzzi, C.; Fascina, V.B.; Lopez-Ulibarri, R.; Calvo, E.P.; Beirão, B.C.; Caron, L.F. Effects of a microbial muramidase on the growth performance, intestinal permeability, nutrient digestibility, and welfare of broiler chickens. Poult. Sci. 2022, 101, 102232. [Google Scholar] [CrossRef] [PubMed]
- Kogut, M.H.; Genovese, K.J.; Swaggerty, C.L.; He, H.; Broom, L. Inflammatory phenotypes in the intestine of poultry: Not all inflammation is created equal. Poult. Sci. 2018, 97, 2339–2346. [Google Scholar] [CrossRef]
- Lee, M.; Kovacs-Nolan, J.; Yang, C.; Archbold, T.; Fan, M.Z.; Mine, Y. Hen egg lysozyme attenuates inflammation and modulates local gene expression in a porcine model of dextran sodium sulfate (DSS)-induced colitis. J. Agric. Food Chem. 2009, 57, 2233–2240. [Google Scholar] [CrossRef] [PubMed]
- May, K.; Wells, J.; Maxwell, C.; Oliver, W. Granulated lysozyme as an alternative to antibiotics improves growth performance and small intestinal morphology of 10-day-old pigs. J. Anim. Sci. 2012, 90, 1118–1125. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, M.A.; El-Far, A.H.; Elbestawy, A.R.; Ghanem, R.; Mousa, S.A.; Abd El-Hamid, H.S. Exogenous dietary lysozyme improves the growth performance and gut microbiota in broiler chickens targeting the antioxidant and non-specific immunity mRNA expression. PLoS ONE 2017, 12, e0185153. [Google Scholar] [CrossRef]
- Brugaletta, G.; De Cesare, A.; Laghi, L.; Manfreda, G.; Zampiga, M.; Oliveri, C.; Pérez-Calvo, E.; Litta, G.; Lolli, S.; Sirri, F. A multi-omics approach to elucidate the mechanisms of action of a dietary muramidase administered to broiler chickens. Sci. Rep. 2022, 12, 5559. [Google Scholar] [CrossRef]
- Humphrey, B.D.; Huang, N.; Klasing, K.C. Rice expressing lactoferrin and lysozyme has antibiotic-like properties when fed to chicks. J. Nutr. 2002, 132, 1214–1218. [Google Scholar]
- Liu, D.; Guo, Y.; Wang, Z.; Yuan, J. Exogenous lysozyme influences Clostridium perfringens colonization and intestinal barrier function in broiler chickens. Avian Pathol. 2010, 39, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Anderson, D.; Rathgeber, B.; MacIsaac, J. The effect of dietary lysozyme with EDTA on growth performance and intestinal microbiota of broiler chickens in each period of the growth cycle. J. Appl. Poult. Res. 2017, 26, 1–8. [Google Scholar] [CrossRef]
- Schliffka, W.; Zhai, H.-X.; Calvo, E.P.; van Cauwenberghe, S.; Walsh, M.C.; Lopez-Ulibarri, R. Safety and efficacy evaluation of a novel dietary muramidase for swine. Heliyon 2019, 5, e02600. [Google Scholar] [CrossRef] [PubMed]
- Robertson, F.P.; Bessell, P.R.; Diaz-Nieto, R.; Thomas, N.; Rolando, N.; Fuller, B.; Davidson, B.R. High serum Aspartate transaminase levels on day 3 postliver transplantation correlates with graft and patient survival and would be a valid surrogate for outcome in liver transplantation clinical trials. Transpl. Int. 2016, 29, 323–330. [Google Scholar] [CrossRef]
- Abdelazeem, A.S.; Fayed, A.M.; Basyony, M.M.; Abu Hafsa, S.H.; Mahmoud, A.E. Hematology profile, digestive enzymes, thyroid hormones, productivity, and nitrogen balance of growing male rabbits supplemented with exogenous dietary lysozyme. Anim. Biotechnol. 2023, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Scanes, C.G.; Harvey, S.; Marsh, J.A.; King, D.B. Hormones and growth in poultry. Poult. Sci. 1984, 63, 2062–2074. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, S.; Xia, Z.; Yuan, J. Effects of different types of polyunsaturated fatty acids on immune function and PGE2 synthesis by peripheral blood leukocytes of laying hens. Anim. Feed Sci. Technol. 2004, 116, 249–258. [Google Scholar] [CrossRef]
- Johnson, R.W. Inhibition of growth by pro-inflammatory cytokines: An integrated view. J. Anim. Sci. 1997, 75, 1244–1255. [Google Scholar] [CrossRef]
- Spurlock, M. Regulation of metabolism and growth during immune challenge: An overview of cytokine function. J. Anim. Sci. 1997, 75, 1773–1783. [Google Scholar] [CrossRef]
- Cooper, C.A.; Brundige, D.R.; Reh, W.A.; Maga, E.A.; Murray, J.D. Lysozyme transgenic goats’ milk positively impacts intestinal cytokine expression and morphology. Transgenic Res. 2011, 20, 1235–1243. [Google Scholar] [CrossRef][Green Version]
- Masschalck, B.; Michiels, C.W. Antimicrobial properties of lysozyme in relation to foodborne vegetative bacteria. Crit. Rev. Microbiol. 2003, 29, 191–214. [Google Scholar] [CrossRef] [PubMed]
- Brundige, D.R.; Maga, E.A.; Klasing, K.C.; Murray, J.D. Lysozyme transgenic goats' milk influences gastrointestinal morphology in young pigs. J. Nutr. 2008, 138, 921–926. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ma, X.; Zhang, S.; Pan, L.; Piao, X. Effects of lysozyme on the growth performance, nutrient digestibility, intestinal barrier, and microbiota of weaned pigs fed diets containing spray-dried whole egg or albumen powder. Can. J. Anim. Sci. 2017, 97, 466–475. [Google Scholar] [CrossRef]
- Wang, Y.; Goossens, E.; Eeckhaut, V.; Calvo, E.P.; Lopez-Ulibarri, R.; Eising, I.; Klausen, M.; Debunne, N.; De Spiegeleer, B.; Ducatelle, R. Dietary muramidase degrades bacterial peptidoglycan to NOD-activating muramyl dipeptides and reduces duodenal inflammation in broiler chickens. Br. J. Nutr. 2021, 126, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Negroni, A.; Pierdomenico, M.; Cucchiara, S.; Stronati, L. NOD2 and inflammation: Current insights. J. Inflamm. Res. 2018, 49–60. [Google Scholar] [CrossRef]
- Strober, W.; Murray, P.J.; Kitani, A.; Watanabe, T. Signalling pathways and molecular interactions of NOD1 and NOD2. Nat. Rev. Immunol. 2006, 6, 9–20. [Google Scholar] [CrossRef]
- Abu Hafsa, S.H.; Mahmoud, A.E.; Fayed, A.M.; Abdel-Azeem, A.-A.S. The Effect of Exogenous Lysozyme Supplementation on Growth Performance, Caecal Fermentation and Microbiota, and Blood Constituents in Growing Rabbits. Animals 2022, 12, 899. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, X.; Yang, Y.; Han, D.; Jin, J.; Xie, S. Effect of dietary lysozyme on growth, immune response, intestine microbiota, intestine morphology and resistance to A eromonas hydrophilia in gibel carp (C arassius auratus gibelio). Aquac. Nutr. 2014, 20, 229–241. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).