
Functional Anatomy of the Thoracic Limb of the Komodo Dragon (Varanus komodoensis)

 , ,
, ,  , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
Active and Passive Movement System—Between Past and Present Research
2. Materials and Methods
2.1. Animals
2.2. Anatomical and Physiological Evaluation
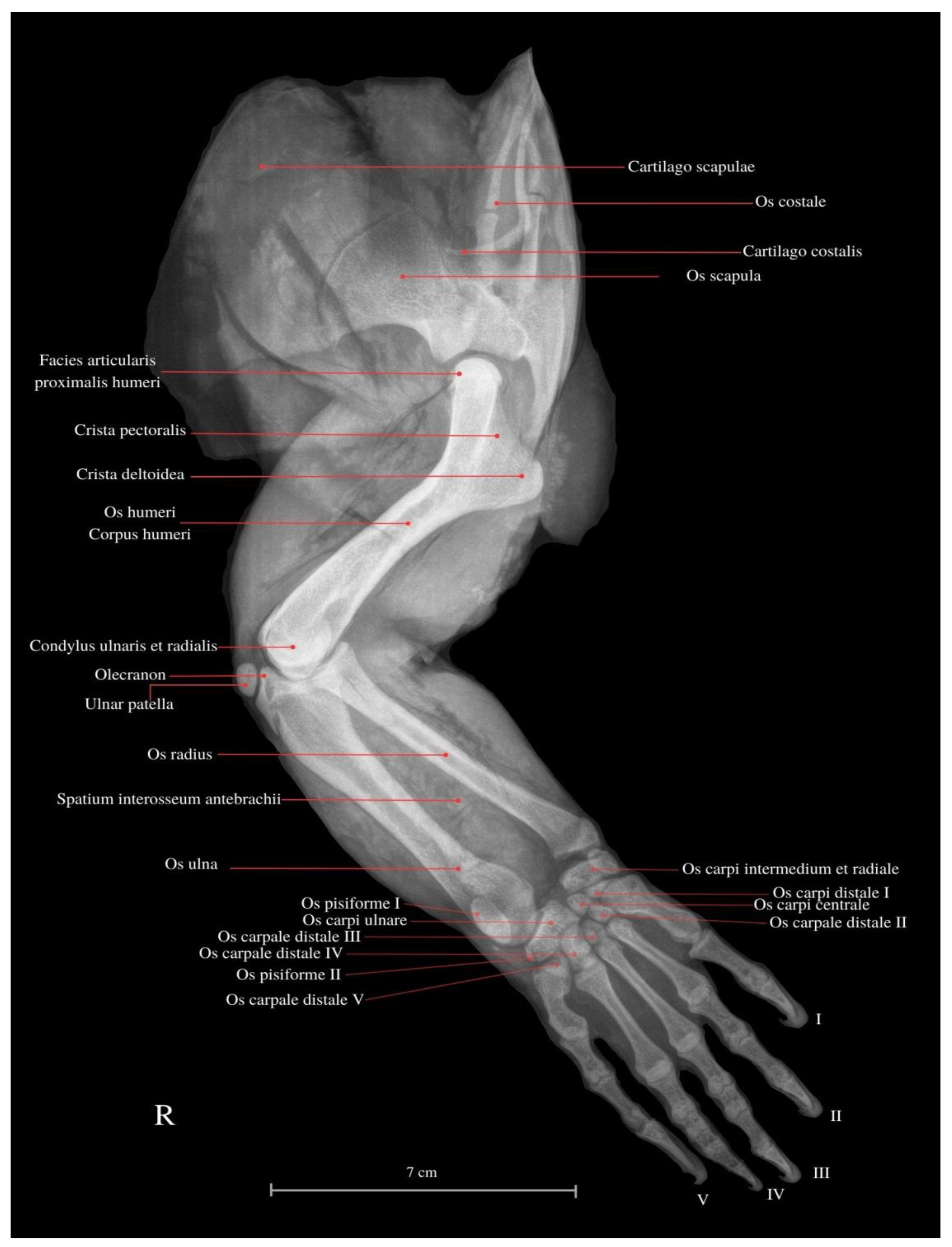
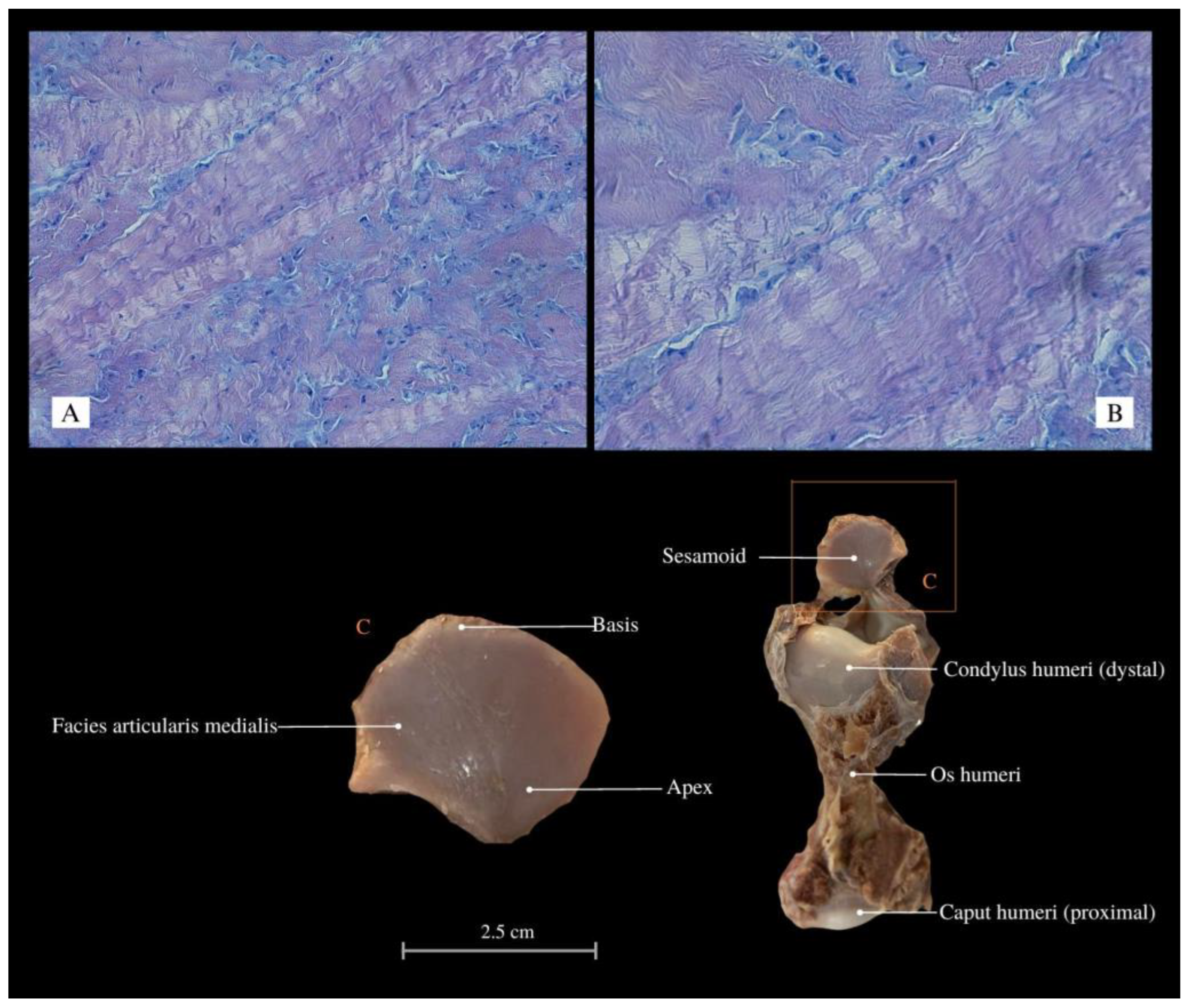
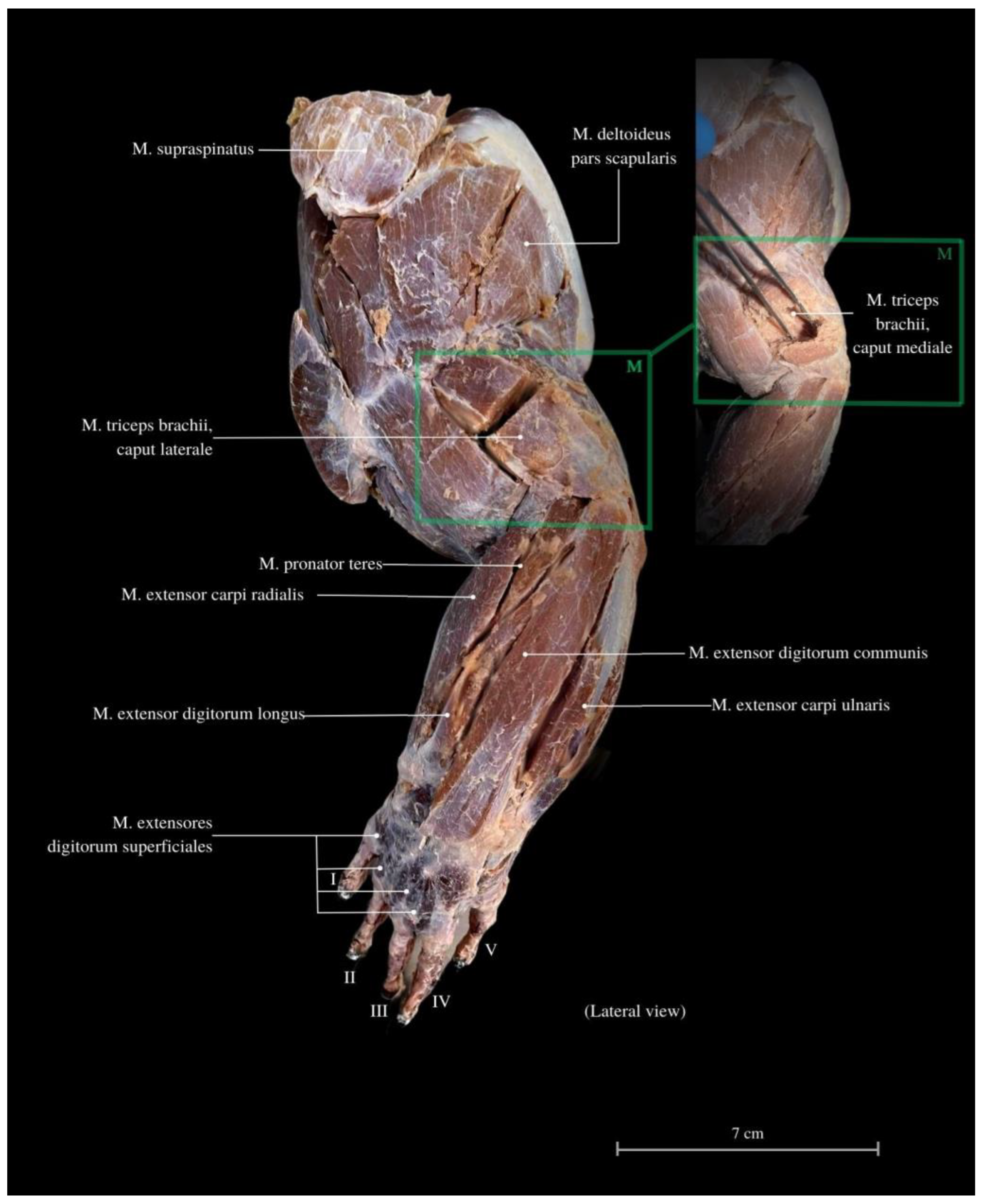
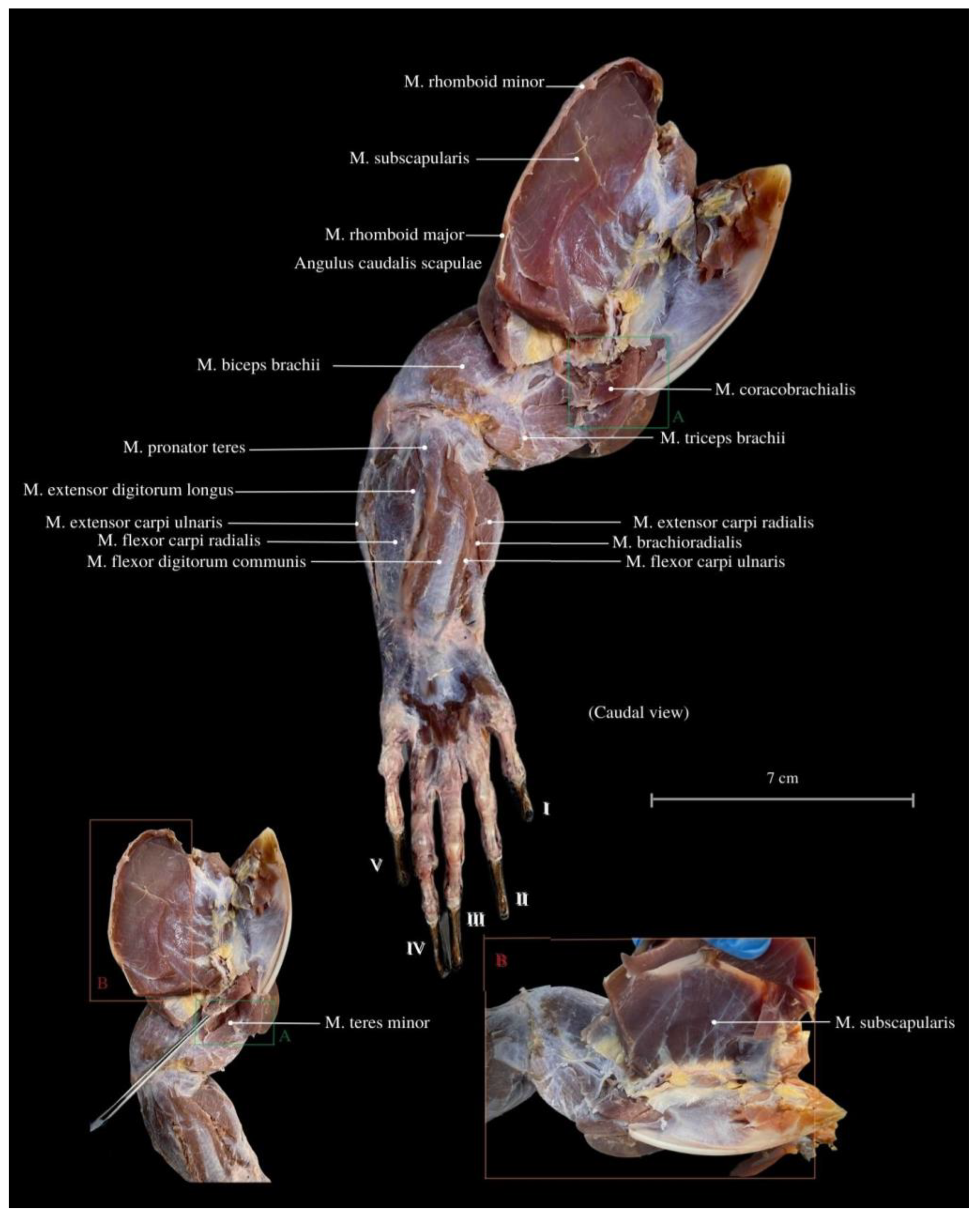
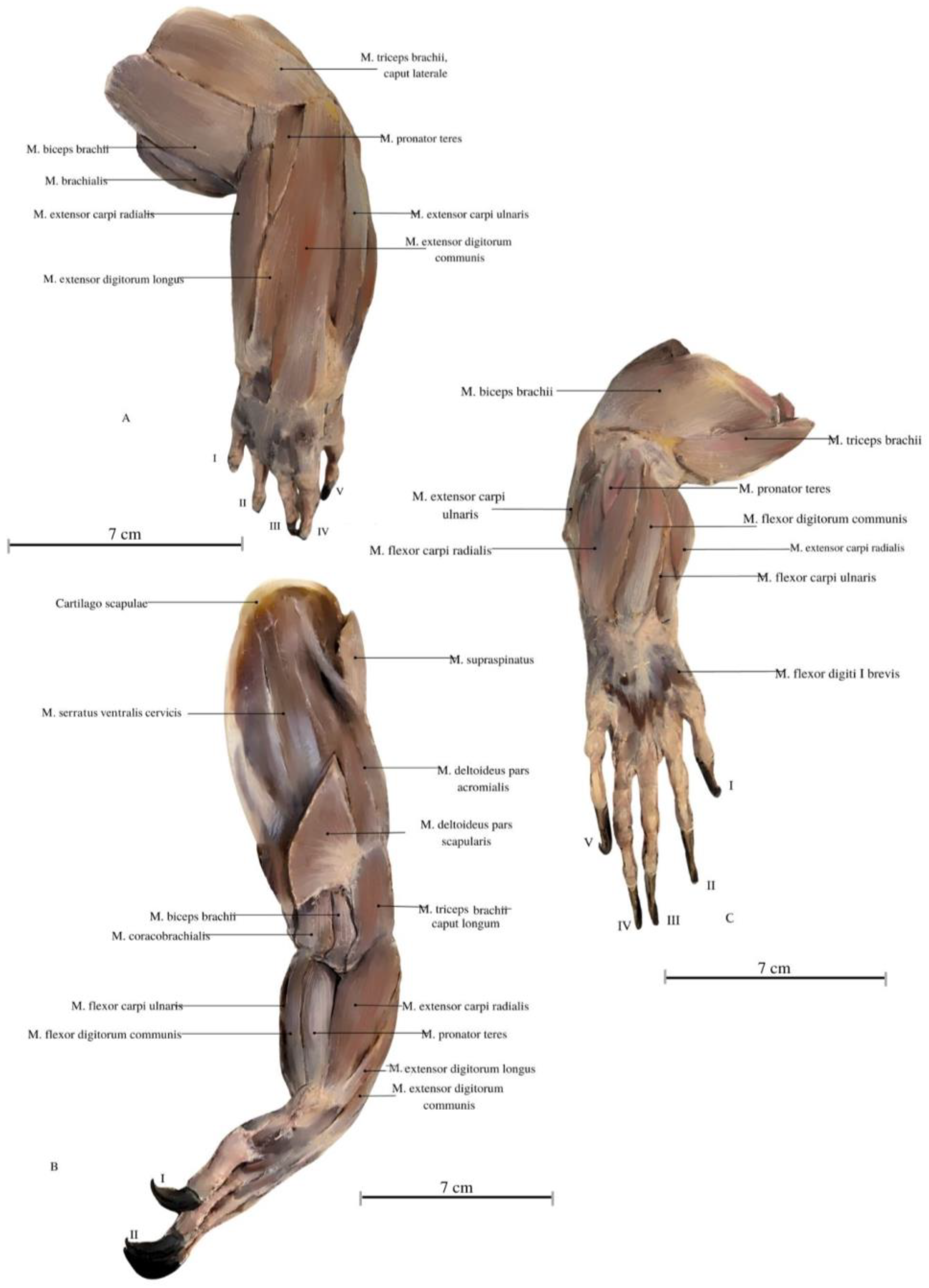
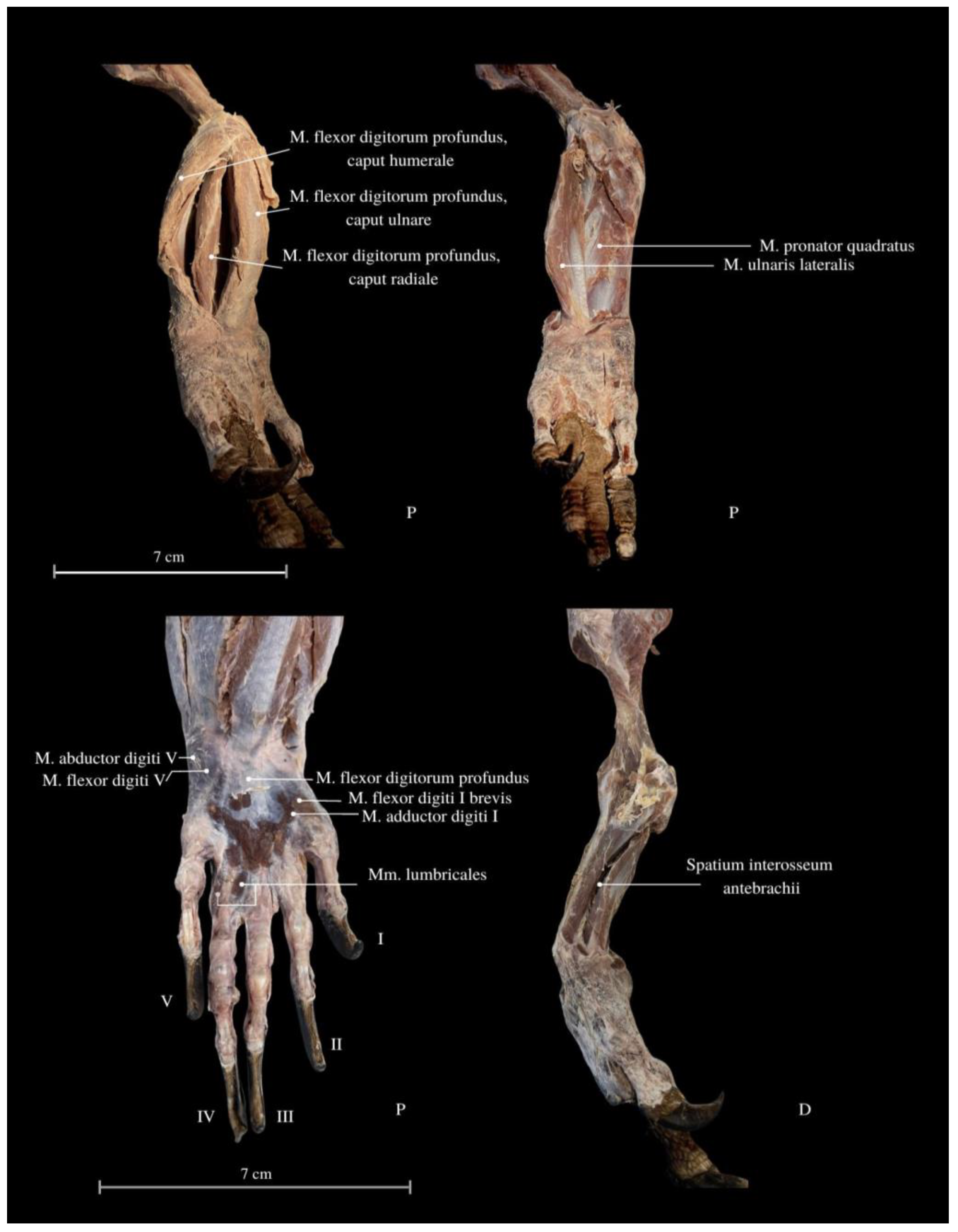
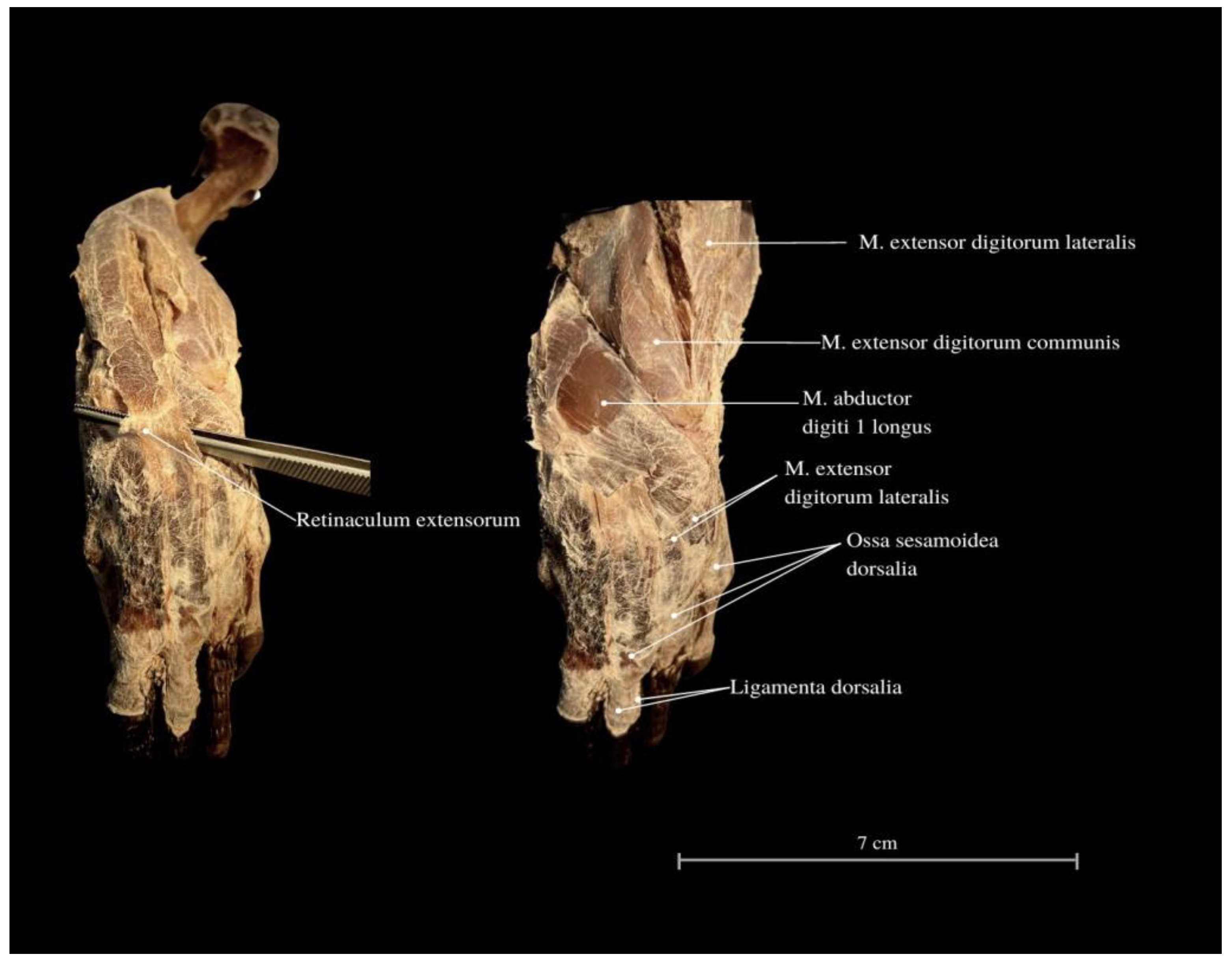
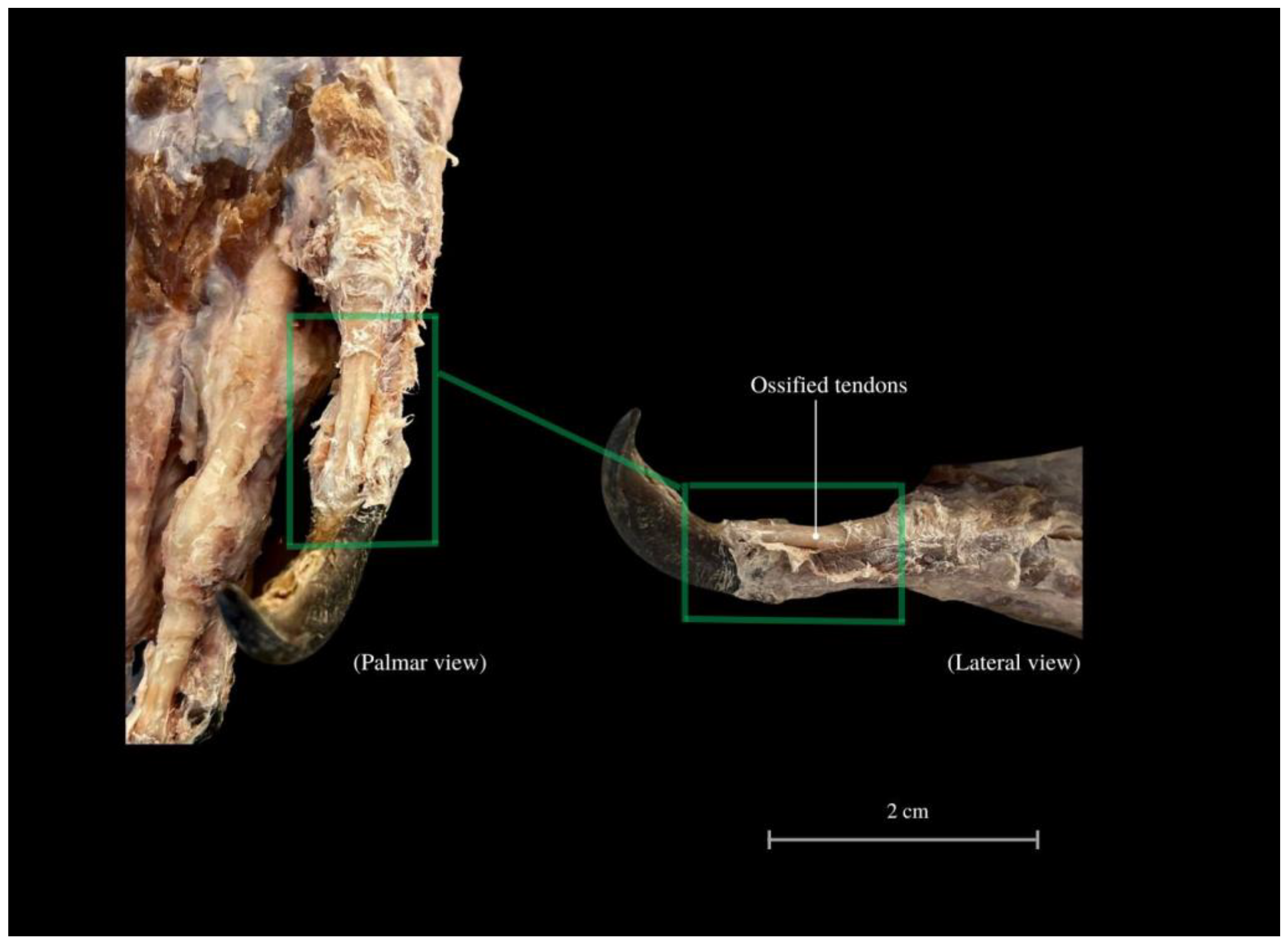
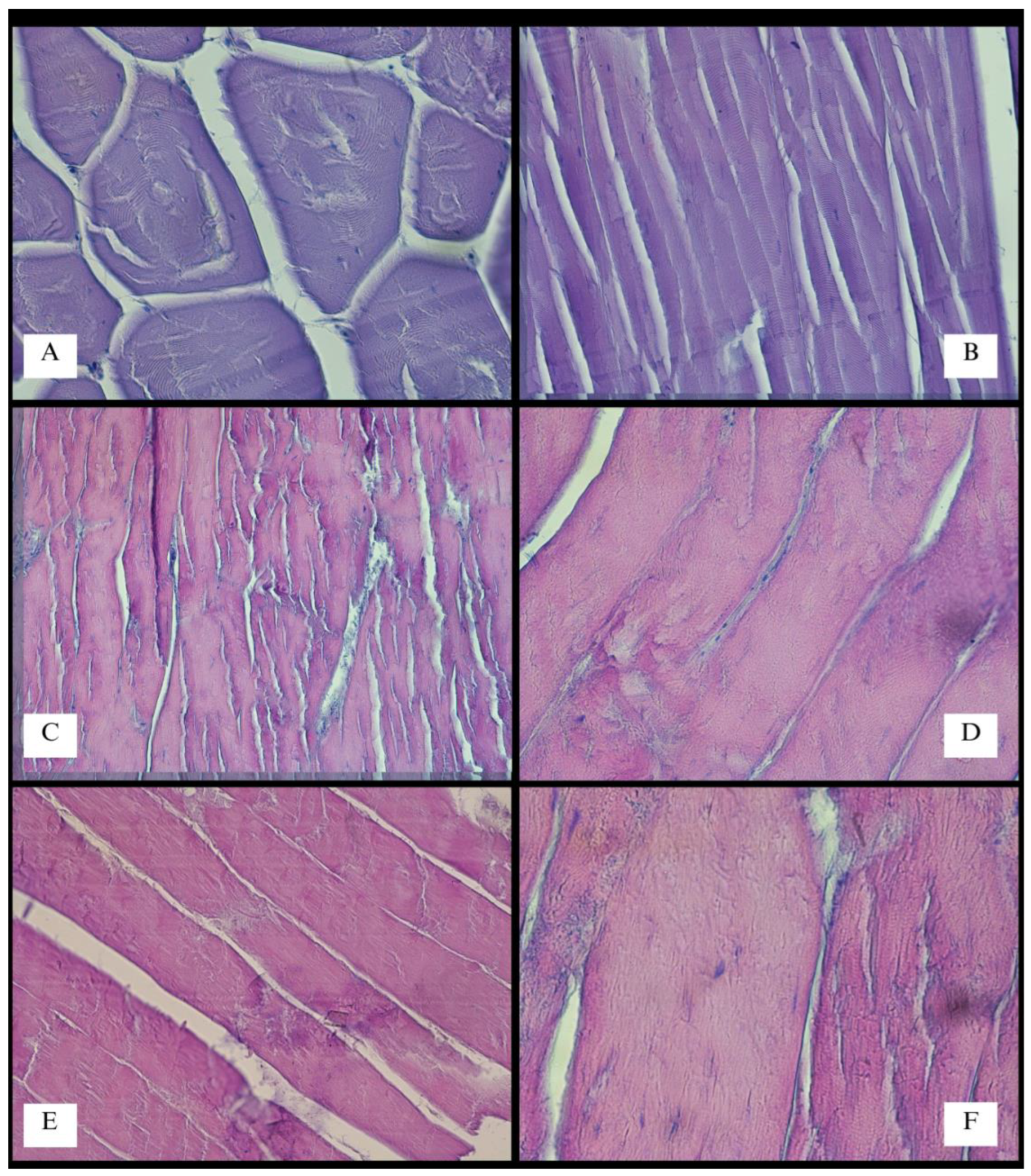
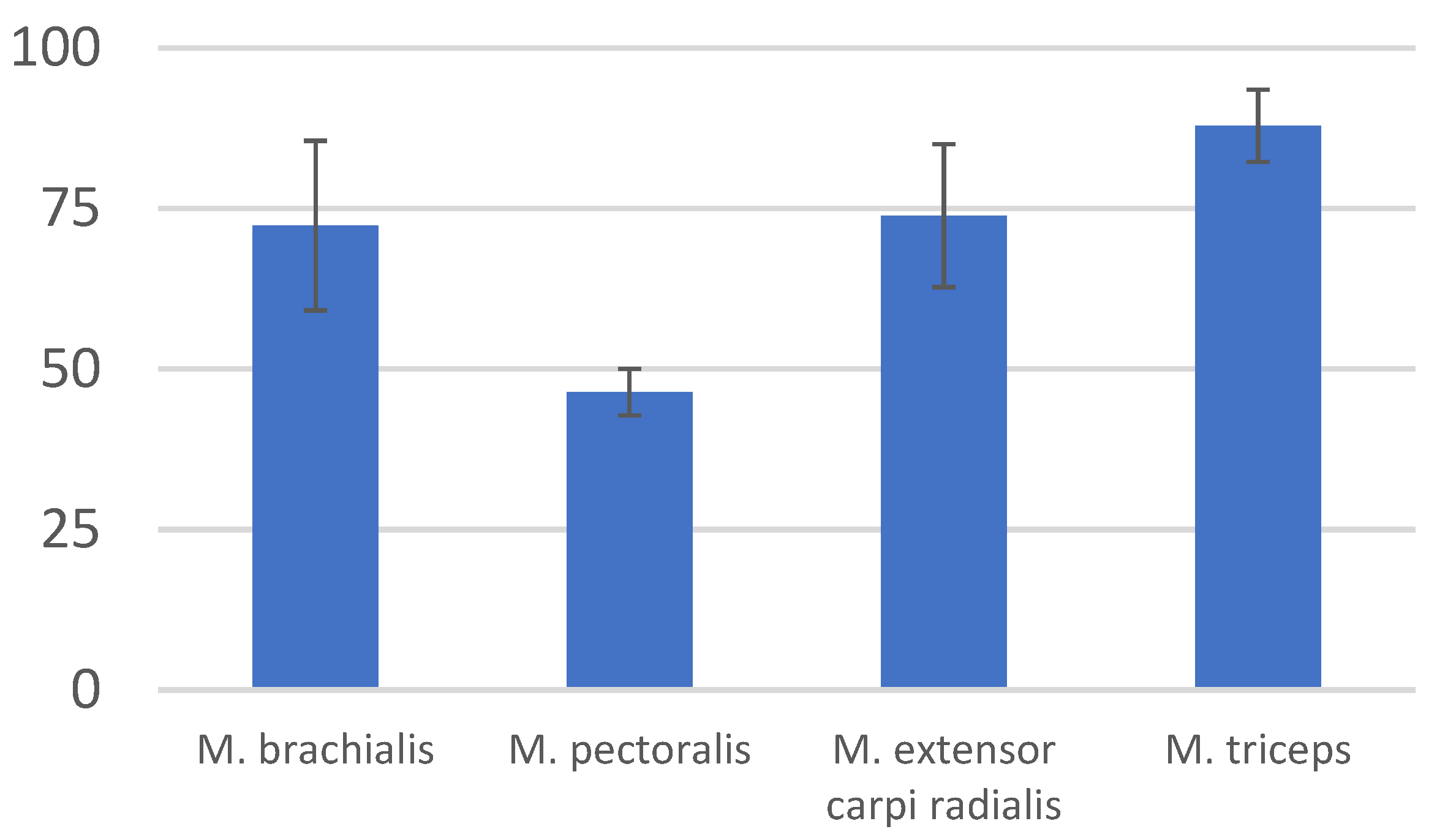
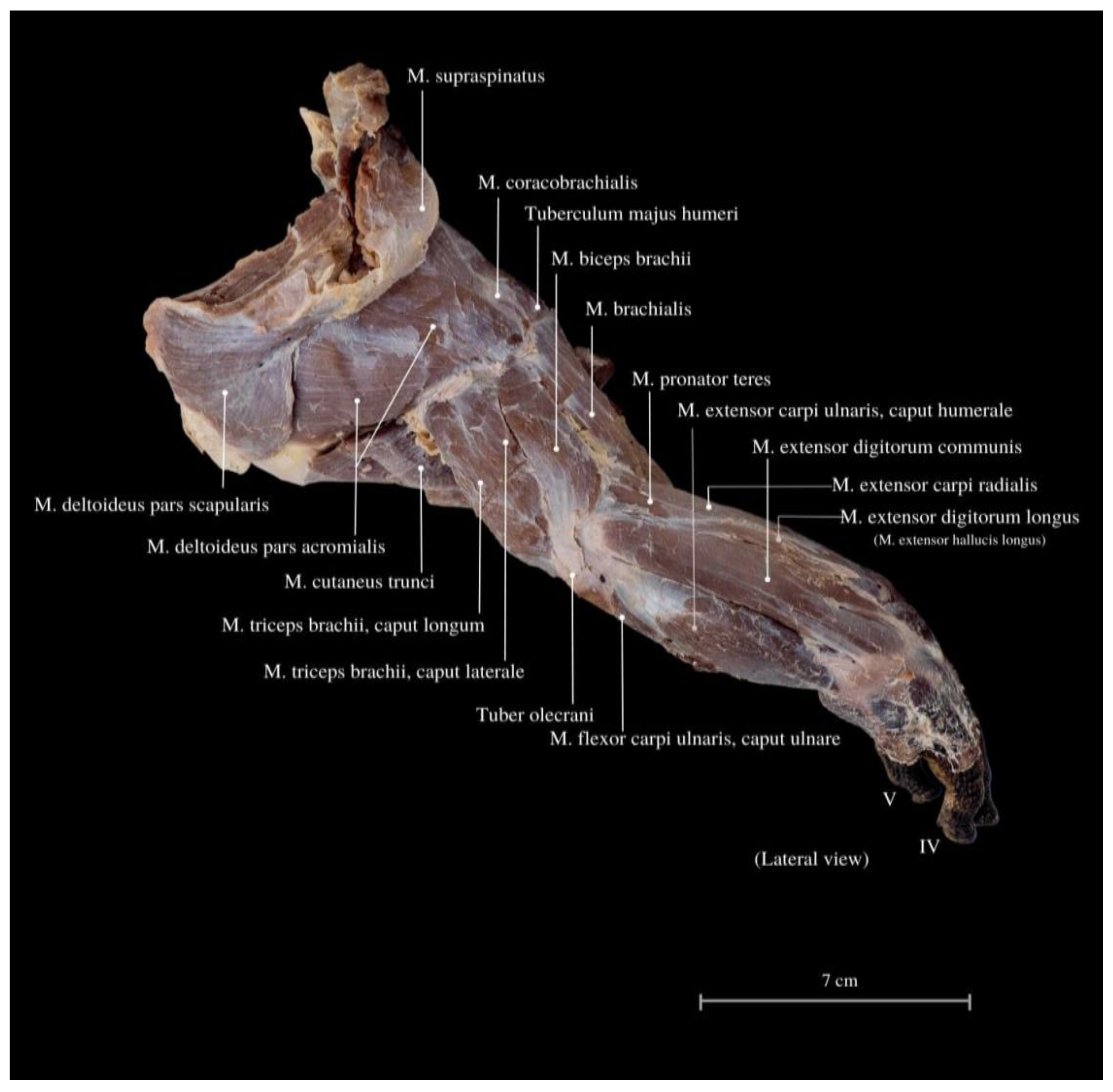
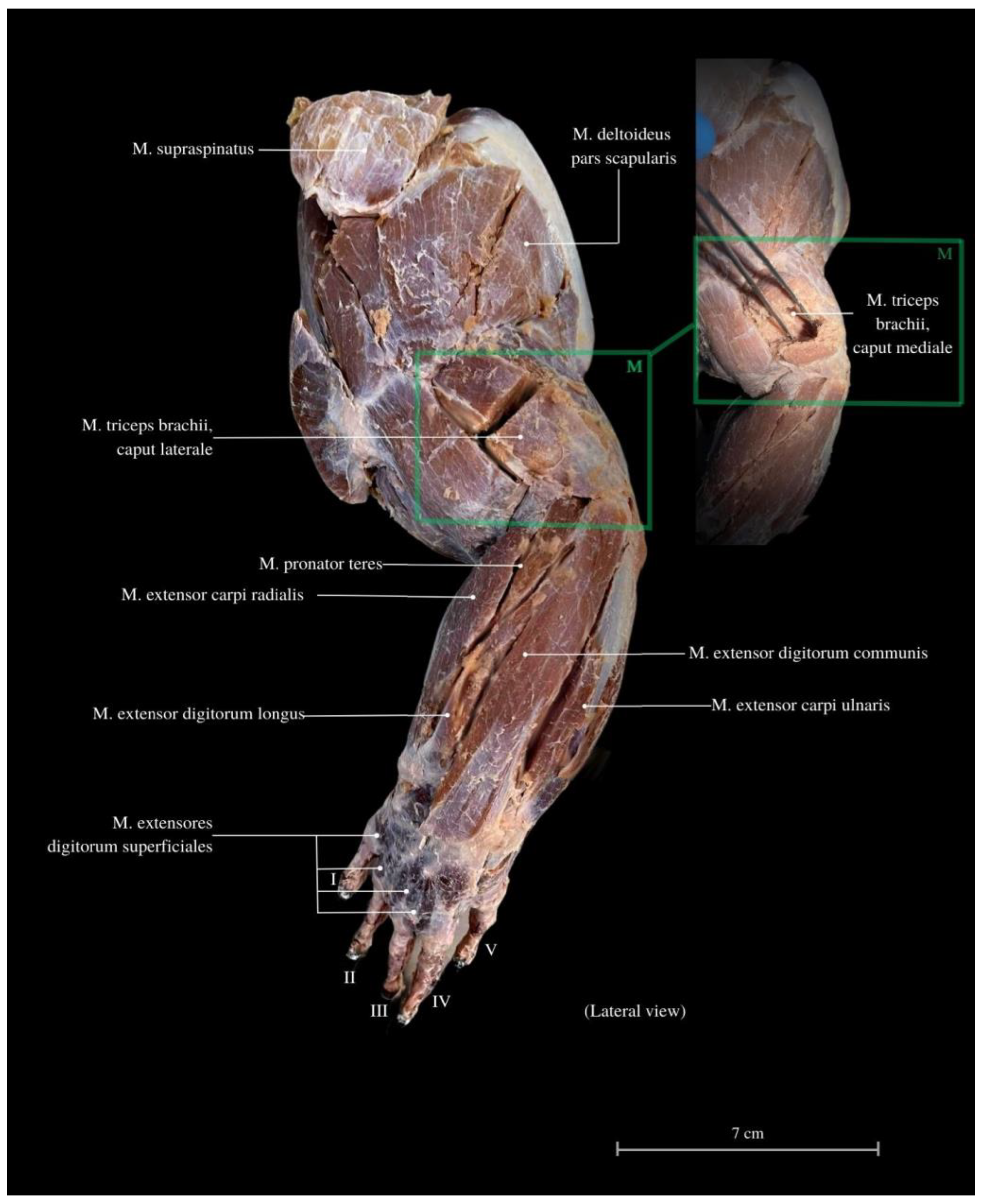
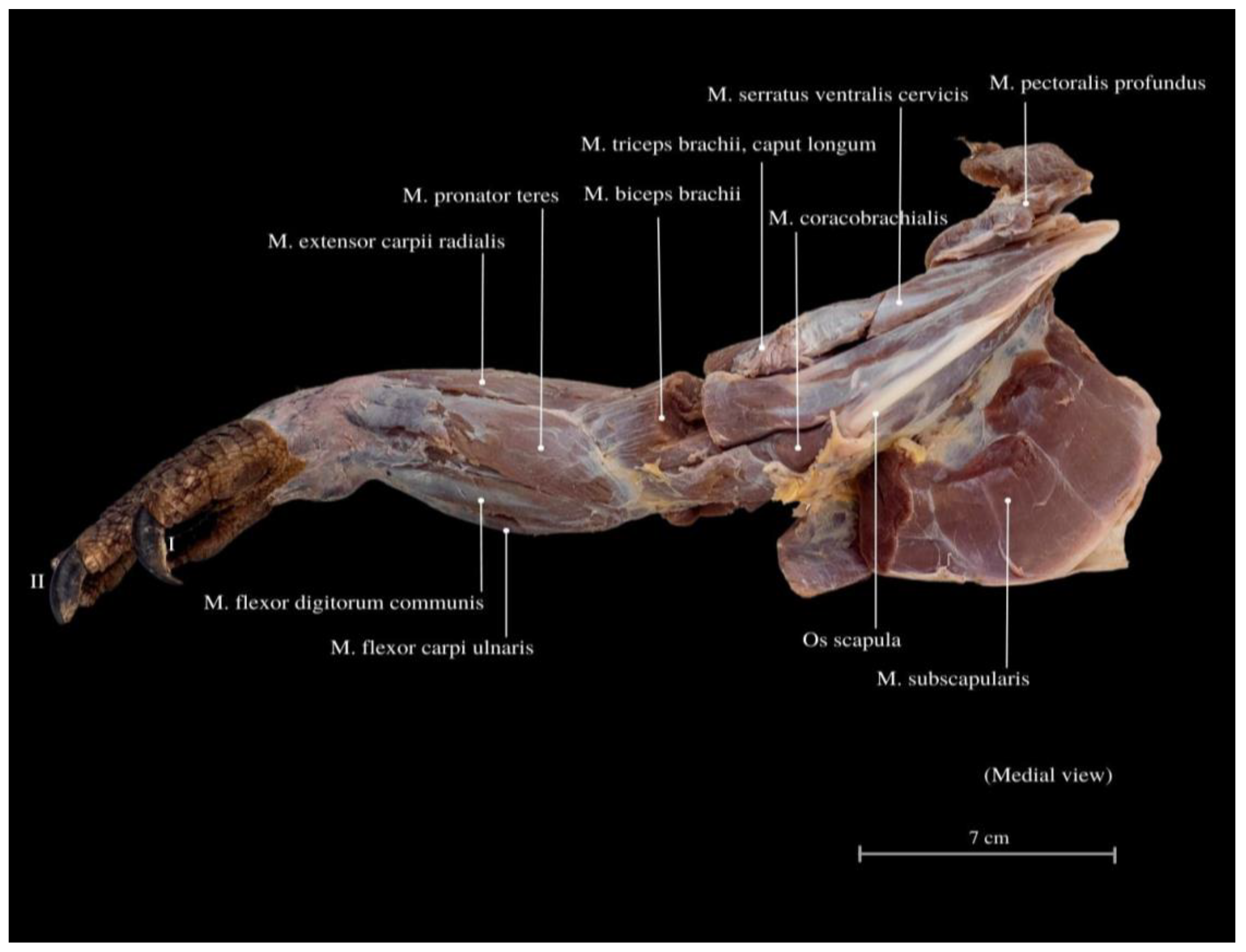
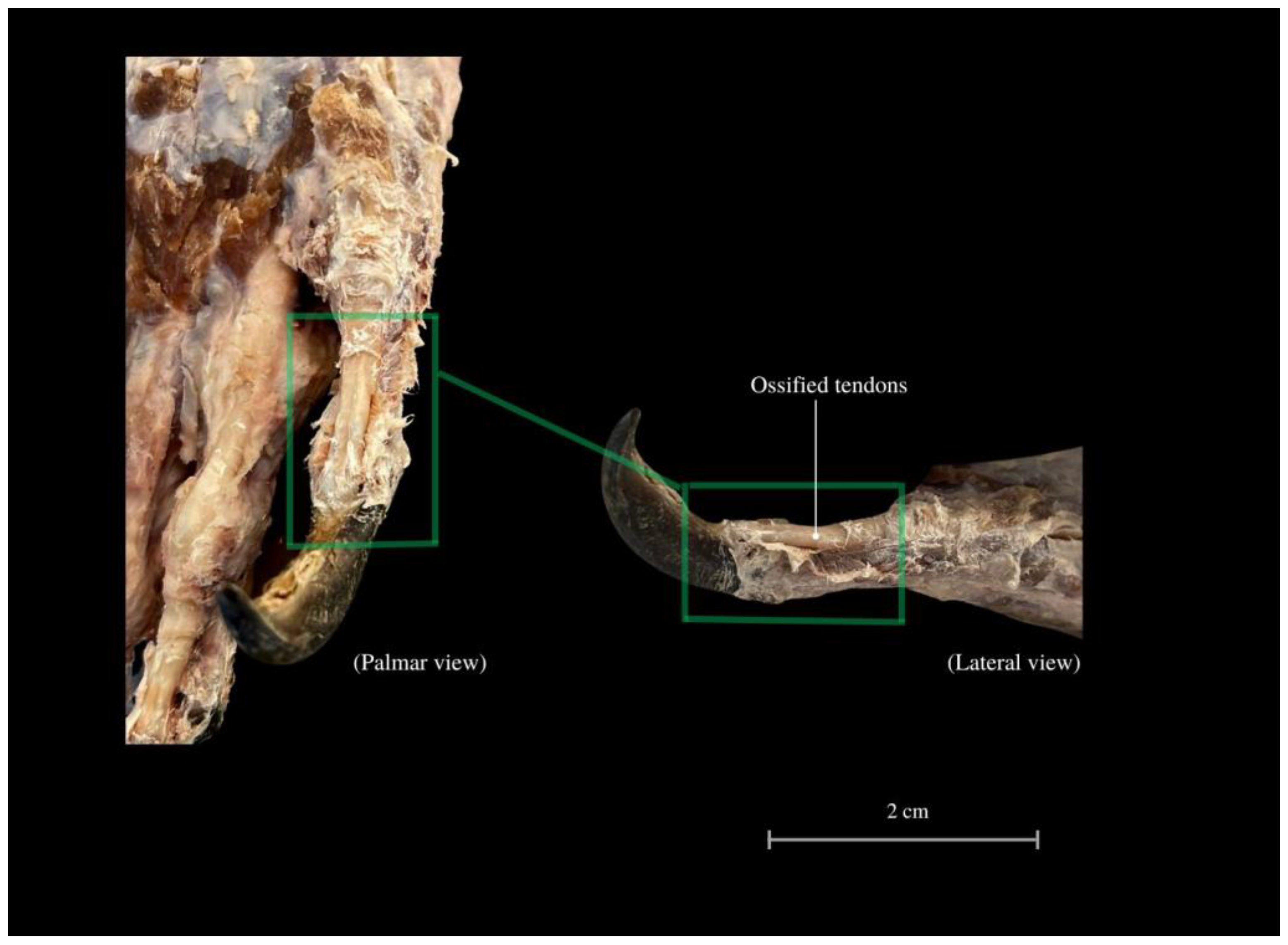
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Integrated Taxonomic Information System (ITIS). Available online: https://doi.org/10.5066/F7KH0KBK (accessed on 13 December 2022).
- Ciofi, C. The Komodo dragon. Sci. Am. 1999, 280, 84–91. [Google Scholar] [CrossRef]
- The American Museum of Natural History 1996. Available online: https://web.archive.org (accessed on 18 November 2022).
- Sahu, M.A. Tingkat Perjumpaan Komodo (Varanus komodoensis Ouwens, 1912). Pada Jalur-Jalur-Wisata di Loh Liang, Taman Nasional Komodo; Skripsi; Universitas Atma Jaya Yogyakarta: Bogor, Indonesia, 2018; pp. 6–16. [Google Scholar]
- Jones, A.R.; Jessop, T.S.; Ariefiandy, A.; Brook, B.W.; Brown, C.S.; Ciofi, C.; Benu, Y.J.; Parwandana, D.; Sitorus, T.; Wigley, T.M.L.; et al. Indentifying island safe havens to prevent the extinction of the World’s largest lizard from global warming. Ecol. Evol. 2020, 10, 10492–10507. [Google Scholar] [CrossRef] [PubMed]
- Ciofi, C.; Puswati, J.; Winana, D.; de Boer, M.; Chelazzi, G.; Sastrawan, P. Preliminary Analysis of Home Range Structure in the Komodo Monitor, Varanus komodoensis. Copeia 2007, 2007, 462–470. [Google Scholar] [CrossRef]
- Available online: Colchester-zoo.com (accessed on 4 February 2023).
- Ciofi, C.; De Boer, M.E. Distribution and conservation of the Komodo monitor (Varanus komodoensis). Herpetol. J. 2004, 14, 99–107. [Google Scholar]
- Jessop, T.; Ariefiandy, A.; Azmi, M.; Ciofi, C.; Imansyah, J.; Purwandana, D. Varanus komodensis, Komodo Dragon; The IUNC Red List of Threatened Species™: Gland, Switzerland, 2021; pp. 1–12. Available online: https://www.iucnredlist.org/species/22884/123633058 (accessed on 17 November 2022).
- Good, D.A.; Bauer, A.M.; Günther, R. An Annotated Type Catalogue of the Anguimorph Lizards (Squamata: Anguidae, Helodermatidae, Varanidae, Xenosauridae) in the Zoological Museum, Berlin. Zoosystematics Evol. 1993, 69, 45–56. [Google Scholar] [CrossRef]
- Srikulnath, K.; Uno, Y.; Nishida, C.; Matsuda, Y. Karyotype evolution in monitor lizards: Cross-species chromosome mapping of cDNA reveals highly conserved synteny and gene order in the Toxicofera clade. Chromosome Res. 2013, 21, 805–819. [Google Scholar] [CrossRef]
- Skawiński, T.; Skórzewski, G.; Borczyk, B. Embryonic development and perinatal skeleton in a limbless, viviparous lizard, Anguis fragilis (Squamata: Anguimorpha). PeerJ 2021, 9, e11621. [Google Scholar] [CrossRef] [PubMed]
- Talanda, M. Early Evolution of Lizards in the Fossil Record. Ph.D. Thesis, Faculty of Biology of University of Warsaw, Warsaw, Poland, 2016; pp. 1–144. [Google Scholar]
- Palci, A.; Caldwell, M.W. Vestigial Forelimbs and Axial Elongation in a 95 Million-Year-Old Non-Snake Squamate. J. Vertebr. Paleontol. 2007, 27, 1–7. [Google Scholar] [CrossRef]
- Simões, T.; Pyron, R.A. The Squamate Tree of Life. Bull. Mus. Comp. Zool. 2021, 163, 47–95. [Google Scholar] [CrossRef]
- Laver, R.J.; Purwandana, D.; Ariefiandy, A.; Imansyah, J.; Forsyth, D.; Ciofi, C.; Jessop, T.S. Life-History and Spatial Determinants of Somatic Growth Dynamics in Komodo Dragon Populations. PLoS ONE 2012, 7, e45399. [Google Scholar] [CrossRef]
- Jessop, T.S.; Madsen, T.; Sumner, J.; Rudiharto, H.; Philips, J.A.; Ciofi, C. Maximum body size among insular Komodo dragon populations covaries with large prey density. OIKOS Adv. Ecol. 2006, 112, 422–429. [Google Scholar] [CrossRef]
- Collar, D.C.; Schulte, J.A., II; Losos, J.B. Evolution of extreme body size disparity in monitor lizards (Varanus). Evolution 2011, 69, 2664–2680. [Google Scholar] [CrossRef] [PubMed]
- Hocknull, S.A.; Piper, P.J.; van den Bergh, G.D.; Due, R.A.; Morwood, M.J.; Kurniawan, I. Dragon’s Paradise Lost: Palaeobiogeography, Evolution and Extinction of the Largest-Ever Terrestrial Lizards (Varanidae). PLoS ONE 2009, 4, E7241. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.B.; Ciofi, C. (Eds.) Komodo Dragons Biology and Conservation; Smithsonian Institution: Washington, DC, USA, 2002; pp. 10–270. [Google Scholar]
- Benson, K. Monitors: The Biology of Varanid Lizards. Am. Zool. 2000, 40, 156. [Google Scholar]
- Pfeffer, P. Observations sur le Varan de komodo. In Varanus Komodoensis, Ouwens 1912; Sociéte Nationale D’Acclimatation: Paris, France, 1912; pp. 195–243. Available online: https://www.usda.gov/ (accessed on 23 November 2022).
- Available online: https://nasional.kompas.com/read/2011/02/04/13481641/~Sains~Biologi?page=all (accessed on 14 January 2023).
- Available online: https://www.theatlantic.com/science/archive/2015/05/the-dragon-autopsy/393890/ (accessed on 16 January 2023).
- Nurhaliza, S.S.; Mustika, A.A. Nurhidayat. Perbandingan Karakteristik Anatomi Skelet Kepala Komodo (Varanus komodoensis) Dan Biawak Air (Varanus salvator); IPB University: Bogor, Indonesia, 2019; pp. 1–24. [Google Scholar]
- Juárez-Campos, I.; Núñez-Altamirano, D.A.; Juárez-Campos, B.; Márquez-Pérez, L.; Juárez-Campos, M.E.; Romero, L. Bioinspired sprawling robotic leg and a path-planning procedure. Int. J. Adv. Robot. Syst. 2018, 15. [Google Scholar] [CrossRef]
- Cooke, S.J.; Sack, L.; Franklin, C.E.; Farrell, A.P.; Beardall, J.; Wikelski, M.; Chown, S.L. What is conservation physiology? Perspectives on an increasingly integrated and essential science. Conserv. Physiol. 2013, 1, cot001. [Google Scholar] [CrossRef] [PubMed]
- Estes, R.; de Queiroz, K.; Gauthier, J. Phylogenic Relationships within Squamata. In Phylogenetic Relationships of the Lizard Families; Stanford University Press: Stanford, CA, USA, 1988; pp. 119–130. [Google Scholar]
- Villa, A.; Abella, J.; Alba, D.M.; Almécija, S.; Bolet, A.; Koufos, G.D.; Knoll, F.; Luján, A.H.; Morales, J.; Robies, J.M.; et al. Revision of Varanus marathonensis (Squamata, Varanidae): Based on historical and new material: Morphology, systematics, and paleobiogeography of the European monitor lizards. PLoS ONE 2018, 13, e0207719. [Google Scholar] [CrossRef]
- Molnar, J.L.; Diogo, R. Evolution, Homology, and Development of Tetrapod Limb Muscles. Evol. Dev. Diversif. Vertebr. Divers. 2021, 13, 393. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 93. [Google Scholar] [CrossRef]
- Al-Haassawi, A.M.A. The Osteology and Myology of the Craniocervical Region in Squamate Reptile: A Comparative Study. Ph.D. Thesis, University of London, London, UK, 2004; pp. 1–568. [Google Scholar]
- Russell, A.P.; Bauer, A.M. The appendicular locomotor apparatur of Sphendon and normal-limbed Squamates. In Biology of Reptilia (The Skull and Appendicular Locomotor Apparatus of Lepidosauria); Gans, C., Gaunt, A.S., Adler, K., Eds.; Society for the Study of Amphibians & Reptiles: Plymouth, MN, USA, 2008; Volume 24, pp. 1–465. [Google Scholar]
- Villaseñor-Amador, D.; Xanat Suárez, N.; Cruz, J.A. Bipedalism in Mexican Albian lizard (Squamata) and the locomotion type in other Cretaceous lizards. J. S. Am. Earth Sci. 2021, 109, 103299. [Google Scholar] [CrossRef]
- Clemente, C.J.; Withers, P.C.; Thompson, G.; Lloyd, D. Why go bipedal? Locomotion and morphology in Australian agamid lizards. J. Exp. Biol. 2008, 211, 2058–2065. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, Y.N.; Fiorillo, A.R.; Lü, J. Lizards ran bipedally 110 million years ago. Sci. Rep. 2018, 8, 2617. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, M. Lizards of the Word; Murdoch Books: Sydney, NSW, Australia, 2020; p. 22. [Google Scholar]
- Rodrigues, M.T.; Pavan, D.; Curcio, F.F. Two New Species of Lizards of the Genus Bachia (Squamata, Gymnophthalmidae) from Central Brazil. J. Herpetol. 2007, 41, 545–553. [Google Scholar] [CrossRef]
- Brennan, P.L.R.; Orbach, D.N. Copulatory behavior and its relationship to genital morphology. Adv. Study Behav. 2020, 52, 65–122. [Google Scholar]
- Gagliano, P.; Sawiec, A.B.; Gibbons, D.E.; Granatosky, M.C. Squamate Locomotion. In Encyclopedia of Animal Cognition and Behavior; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–7. [Google Scholar]
- Chong, B.; Wang, T.; Ericson, E.; Bergmann, P.J.; Goldman, D.I. Coordinating tiny limbs and long bodies: Geometric mechanics of lizard terrestrial swimming. Biophys. Comput. Biol. 2022, 119, e2188456119. [Google Scholar]
- Pianka, E.R.; Vitt, L.J. Lizards: Window to the Evolution of Diversity; University of California Press: Berkeley, CA, USA, 2003; p. 346. [Google Scholar]
- Christian, A.; Garland, T., Jr. Scaling of Limb Proportions in Monitor Lizards (Squamata: Varanidae). J. Herpetol. 1996, 30, 219–230. [Google Scholar] [CrossRef]
- Brocklehurst, R.J.; Fahn-Lai, P.; Regnault, S.; Pierce, S.E. Muscoskeletal modeling of sprawling and parasagittal forelimbs provides insight into synapsid postural transition. iScience 2022, 25, 1–14. [Google Scholar] [CrossRef]
- Gleeson, T.T. A histochemical and enzymatic study of the muscle fiber types in the water monitor, Varanus salvator. J. Exp. Zool. 1983, 227, 191–201. [Google Scholar] [CrossRef]
- Araya-Donoso, R.; Juan, E.S.; Tamburrino, I.; Lamborot, M.; Veloso, C.; Véliz, D. Integrating genetics, physiology and morphology to study desert adaptation in a lizard species. J. Anim. Ecol. 2021, 91, 1148–1162. [Google Scholar] [CrossRef] [PubMed]
- Dick, T.J.M.; Clemente, C.J. How to build your dragon: Saling of muscle architecture from the world’s smallest to the world’s largest monitor lizard. Front. Zool. 2016, 13, 2–17. [Google Scholar] [CrossRef]
- De Souza Junior, P.; de Souza Pahim, A.B.; Viotto-Souza, W.; Pellenz, J.; Bernardes, F.C.; Abidu-Figueiredo, M.; Santos, A.L.Q. Evolutionary history or function? Which preponderates in the expression of the muscle mass of the thoracic limb in Wild carnivorans? Anat. Rec. 2021, 304, 1344–1356. [Google Scholar] [CrossRef]
- Lowie, A.; Herrel, A.; Abdala, V.; Manzano, A.S.; Fabre, A.C. Does the Morphology of the Forelimb Flexor Muscles Differ Between Lizards Using Different Habitats? Anat. Rec. 2018, 301, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Fakhrian, A.M.; Pamungkas, J. Nurhidayat. Komparasi Karakteristik Anatomi Skelet Ekstermitas Kaki Depan Komodo (Varanus komodoensis) degan Biawak Air (Varanus salvator); Skripsi; IPB University: Bogor, Indonesia, 2023. [Google Scholar]
- De Buffrénil, V.; Ineich, I.; Böhme, W. Comparative Data on Epiphyseal Development in the Family of Varanidae. J. Herpetol. 2005, 39, 328–335. [Google Scholar] [CrossRef]
- Allen, V.; Elsey, R.M.; Jones, N.; Wright, J.; Hutchinson, J.R. Functional specialization and ontogenic sailing of limb anatomy in Alligator mississippiensis. J. Anat. 2010, 216, 423–445. [Google Scholar] [CrossRef]
- Pontzer, H. Effective limb length and the scaling of locomotor cost in terrestrial animals. J. Exp. Biol. 2007, 210, 1752–1761. [Google Scholar] [CrossRef]
- Cieri, R.L.; Dick, T.J.M.; Clemente, C.J. Monitoring muscle over three orders of magnitude: Widespread positive allometry in the pectoral girdle of varanid lizards (Varanidae). J. Anat. 2020, 237, 1114–1135. [Google Scholar] [CrossRef]
- Allen, V.; Molnar, J.; Parker, W.; Pollard, A.; Nolan, G.; Hutchinson, J.R. Comparative architectural properties of limp muscles in Crocodylidae and Alligatoridae and their relevance to divergent use of asymmetrical gaits in extant Crocodylia. J. Anat. 2014, 225, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.L.; Higham, T.E.; Collins, C.E.; Garland, T. Determinants of lizard escape performance: Decision, motivation, ability, and opportunity. In Escaping from Predators: An Integrative View of Escape Decisions. Chapter 11.; Cooper, W.E., Blumstein, D.T., Eds.; Cambridge University Press: Cambridge, UK, 2015; pp. 287–310. [Google Scholar]
- Moreno, K.; Wroe, S.; Clausen, P.; McHenry, C.R.; D’Amore, D.C.; Rayfield, E.J.; Cunningham, E. Cranial performance in the Komodo dragon (Varanus komodoensis) as revealed by high-resolution 3-D finite element analysis. J. Anat. 2008, 212, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, N.; MacLean, P.D. (Eds.) Behavior and Neurology of Lizards: An Interdisciplinary Colloquium; US Department of Health, Education, and Welfare, Public Health Service, Alcohol, Drug Abuse, and Mental Health Administration, National Institute of Mental Health: Rockville, MD, USA, 1989; pp. 60–200.
- Available online: https://www.pbs.org/wnet/nature/the-dragon-chronicles-the-komodo-dragons-remarkable-physiology/4523/ (accessed on 21 November 2022).
- Ritter, D. Epaxial muscle function during locomotion in a lizard (Varanus salvator) and the proposal of a key innovation in the vertebrate axial musculoskeletal system. J. Exp. Biol. 1995, 198, 2477–2490. [Google Scholar] [CrossRef]
- Pianka, E.; King, D. Varanoid Lizards of the World; Indiana University Press: Bloomington, IN, USA, 2004; pp. 200–201. [Google Scholar]
- Janeczek, M.; Goździewska-Harłajczuk, K.; Hrabska, L.; Klećkowska-Nawrot, J.; Kuropka, P.; Dobrzyński, M.; Oleksii, M.; Nikodem, A.M. Macroanatomical, histological and microtomographic study of the teeth of the Komodo Dragon (Varanus komodoensis)—Adaptation to hunting. Biology 2023, 12, 247. [Google Scholar] [CrossRef]
- International Committe on Veterinary Gross Anatomical Nomenclature (I.C.V.G.A.N.). Nomina Anatomica Veterinaria, 6th ed.; Editorial Committee: Hanover, Germany; Ghent, Belgium; Colombia, MO, USA; Rio de Janeiro, Brazil, 2017. [Google Scholar]
- Conrad, J.L. An Eocene shinisaurid (Reptilia, Squamata) from Wyoming, USA. J. Vertebr. Paleontol. 2006, 26, 113–126. [Google Scholar] [CrossRef]
- Ast, J. Mitochondrial DNA Evidence and Evolution in Varanoidea (Squamata). Cladistics 2001, 17, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Abdala, V.; Diogo, R. Comparative anatomy, homologies and evolution of the pectoral and forelimb musculature of tetrapods with special attention to extant limbed amphibians and reptiles. J. Anat. 2010, 217, 536–573. [Google Scholar] [CrossRef]
- Diogo, R.; Linde-Medina, M.; Abdala, V.; Ashley-Ross, M.A. New, puzzling insights from comparative myological studies on the old and unsolved forelimb/hindlimb enigma. Biol. Rev. Camb. Philos. Soc. 2013, 88, 196–214. [Google Scholar] [CrossRef] [PubMed]
- Meers, M.B. Crocodylian forelimb musculature and its relevance to Archosauria. Anat. Rec. 2003, 274A, 891–916. [Google Scholar] [CrossRef] [PubMed]
- Freitas, L.M.; Sabec-Pereira, D.K.; Pereira, K.F.; dos Santos, O.P.; Lima, F.C. Muscular Anatomy of the pectoral girdle and forelimb of Iguana i. iguana (Squamata: Iguanidae). Biosci. J. 2017, 33, 1284–1294. [Google Scholar] [CrossRef]
- Taylor, G.; Bonney, V. On the Homology and Morphology of the Popliteus Muscle: A Contribution to Comparative Myology. J. Anat. Physiol. 1905, 40 Pt 1, 34–50. [Google Scholar]
- Jenkins, F.A., Jr.; Goslow, G.E., Jr. The functional anatomy of the shoulder of the savannah monitor lizard (Varanus exanthematicus). J. Morphol. 1983, 175, 195–216. [Google Scholar] [CrossRef]
- Young, B.A. The Subclavian Loop of Varanus salvator. Copeia 1988, 4, 1029–1034. [Google Scholar] [CrossRef]
- Lima, F.C.; Leite, A.V.; Santos, A.L.Q.; Sabec-Pereira, D.K.; Araújo, E.G.; Pereira, K.F. Muscular anatomy of the pectoral and forelimb of Caiman crocodilus crocodilus (Linnaeus, 1758) (Crocodylia: Alligatoridae). Ciência Anim. Bras. 2016, 17, 285–295. [Google Scholar] [CrossRef]
- Landsmeer, J.M.F. The mechanism of forearm rotation in Varanus exanthematicus. J. Morphol. 1983, 175, 119–130. [Google Scholar] [CrossRef] [PubMed]
- El-Bakry, A.M.; Abdeen, A.M.; Abo-Eleneen, R.E. Comparative study of the osteology and locomotion of some reptilian species. Int. J. Biol. Biol. Sci. 2013, 2, 40–58. [Google Scholar]
- Abdala, V.; Manzano, A.S.; Tulli, M.J.; Herrel, A. The Tendinous Patterns in the Palmar Surface of the Lizard Manus: Functional Consequences for Grasping Ability. Anat. Rec. 2009, 292, 842–853. [Google Scholar] [CrossRef]
- Ibáñez-Gimeno, P.; Galtés, I.; Jordana, X.; Assumpció Malgosa, A.; Manyosa, J. Biomechanics of Forearm Rotation: Force and Efficiency of Pronator Teres. PLoS ONE 2014, 9, e90319. [Google Scholar] [CrossRef] [PubMed]
- Regnault, S.; Hutchinson, J.R.; Jones, M.E.H. Sesamoid bones in tuatara (Sphenodon punctatus) investigated with X-ray microtomoghraphy, and implications for sesamoid evolution in Lepidosauria. J. Morphol. 2016, 278, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Otero, T.; Hoyos, J.M. Sesamoid elements in lizards. Herpetol. J. 2013, 23, 105–114. [Google Scholar]
- Ginet, R.; Grassé, P.P. Traité de Zoologie: Anatomie, systematique, biologie. In Mammiféres; Fascicule 1; Téguments, Squelette. Premier Fascicule; Bulletin Mensuel de la Société Linnéenne de Lyon, 36ᵉ année, n°6, juin; Sociéte Nationale D’Acclimatation: Paris, France, 1967; Volume XVI, p. 778. [Google Scholar]
- Yeung, A.Y.; Garg, R. Anatomy, Sesamoid Bones. StatPearls. 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK578171/ (accessed on 20 May 2023).
- Padian, K.; Olsen, P.E. Footprints of the Komodo Monitor and the Trackways of Fossil Reptiles. Copeia 1984, 1984, 662–671. [Google Scholar] [CrossRef]
- Fleming, G.J. Husbandry and Medical Management of Komodo Dragons (Varanus komodoensis) at the White Oak Conservation Centre. Proc. Assoc. Reptil. Amphib. Vet. 1997, 1, 15–22. Available online: https://cdn.ymaws.com/members.arav.org/resource/resmgr/Files/Proceedings_1997/1997_04.pdf (accessed on 12 December 2021).
- D’Amore, D.C.; Clulow, S.; Doody, J.S.; Rhind, D.; McHenry, C.R. Claw morphometrics in monitor lizards: Variable substrate and habitat use correlate to shape diversity within a predator guild. Ecol. Evol. 2018, 3700, 6766–6775. [Google Scholar] [CrossRef]
- Purwandana, D.; Ariefiandy, A.; Imansyah, M.J.; Seno, A.; Ciofi, C.; Letnic, M.; Jessop, T.S. Ecological allometries and niche use dynamics cross Komodo dragon ontogeny. Naturwissenschaften 2016, 103, 27. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Muscle | Weight (W, g) | Wmuscle/Wbody (%) |
|---|---|---|
| M. trapezius | 12.02 | 0.04 |
| M. deltoideus pars scapularis | 36.44 | 0.121 |
| M. deltoideus pars acromialis | 38.87 | 0.129 |
| M. triceps brachii, caput longum | 16.37 | 0.055 |
| M. triceps brachii, caput mediale | 13.87 | 0.046 |
| M. triceps brachii, caput laterale | 10.16 | 0.034 |
| M. biceps brachii, caput laterale | 24.15 | 0.081 |
| M. biceps brachii, caput mediale | 11.73 | 0.039 |
| M. brachialis | 18.27 | 0.061 |
| M. pronator teres | 5.08 | 0.017 |
| M. extensor digitorum communis superficialis | 11.32 | 0.038 |
| M. extensor carpi radialis | 13.45 | 0.045 |
| M. extensor carpi ulnaris | 17.83 | 0.059 |
| M. extensor digitorum longus (hallucis longus) | 11.88 | 0.039 |
| M. flexor digitorum communis superficialis | 13.21 | 0.044 |
| M. brachioradialis | 9.65 | 0.032 |
| M. flexor carpi ulnaris | 17.83 | 0.059 |
| M. flexor carpi radialis | 12.81 | 0.043 |
| M. flexor digitorum profundus, caput humerale | 15.7 | 0.052 |
| M. flexor digitorum profundus, caput radiale | 14.95 | 0.049 |
| M. flexor digitorum profundus, caput ulnare | 13.53 | 0.045 |
| M. pronator quadratus | 5.06 | 0.017 |
| M. ulnaris lateralis | 8.89 | 0.029 |
| M. abductor digiti I longus | 2.53 | 0.008 |
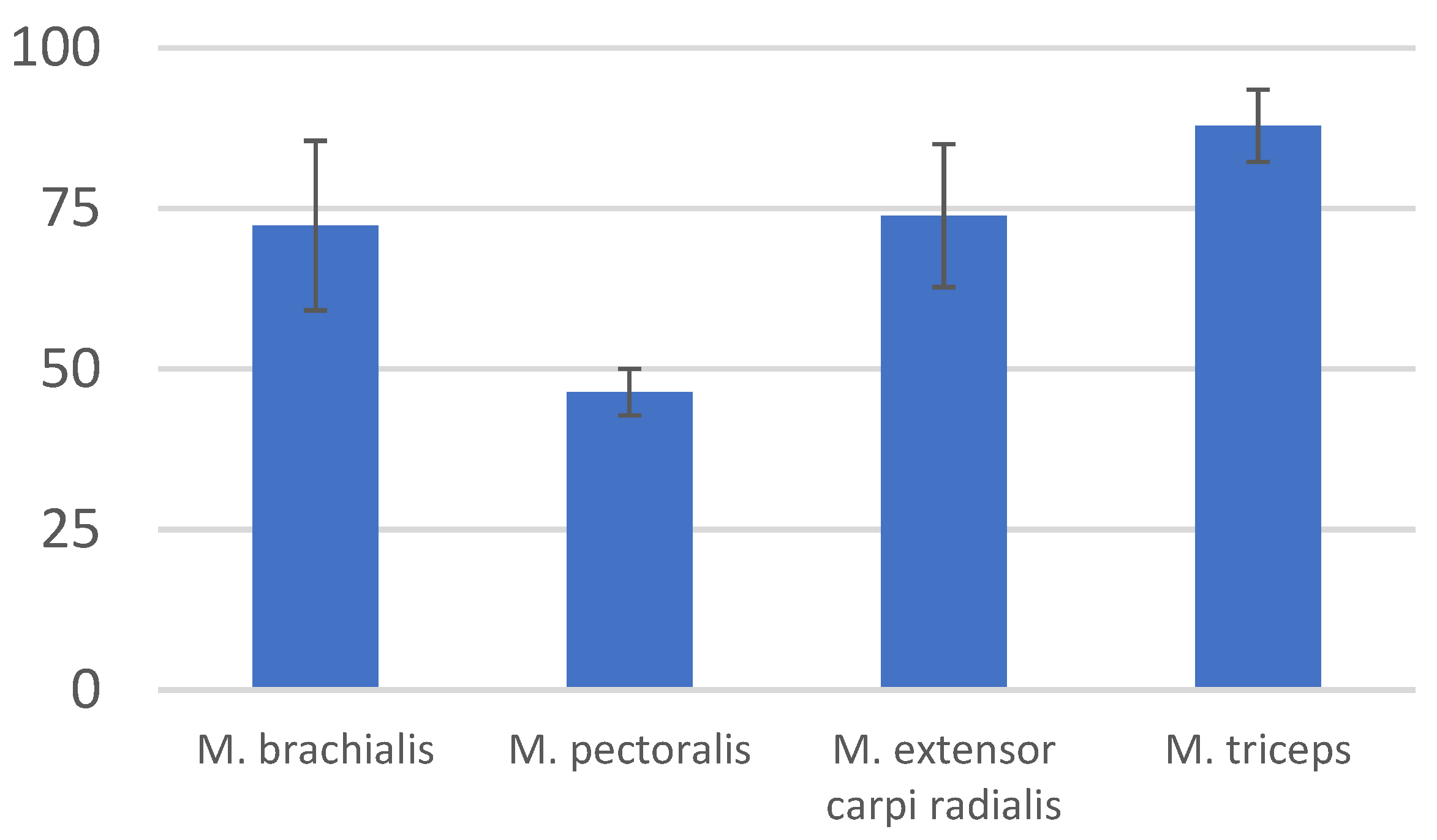
| Muscle | Mean | Standard Deviation (±SD) |
|---|---|---|
| M. brachialis | 72.35 | 13.2 |
| M. pectoralis | 46.41 | 3.635 |
| M. extensor carpi radialis | 73.89 | 11.115 |
| M. biceps brachii | 87.9 | 5.65 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kępa, M.; Tomańska, A.; Staszewska, J.; Tarnowska, M.; Klećkowska-Nawrot, J.; Goździewska-Harłajczuk, K.; Kuźniarski, A.; Gębarowski, T.; Janeczek, M. Functional Anatomy of the Thoracic Limb of the Komodo Dragon (Varanus komodoensis). Animals 2023, 13, 2895. https://doi.org/10.3390/ani13182895
Kępa M, Tomańska A, Staszewska J, Tarnowska M, Klećkowska-Nawrot J, Goździewska-Harłajczuk K, Kuźniarski A, Gębarowski T, Janeczek M. Functional Anatomy of the Thoracic Limb of the Komodo Dragon (Varanus komodoensis). Animals. 2023; 13(18):2895. https://doi.org/10.3390/ani13182895
Chicago/Turabian StyleKępa, Michał, Anna Tomańska, Joanna Staszewska, Małgorzata Tarnowska, Joanna Klećkowska-Nawrot, Karolina Goździewska-Harłajczuk, Amadeusz Kuźniarski, Tomasz Gębarowski, and Maciej Janeczek. 2023. "Functional Anatomy of the Thoracic Limb of the Komodo Dragon (Varanus komodoensis)" Animals 13, no. 18: 2895. https://doi.org/10.3390/ani13182895
APA StyleKępa, M., Tomańska, A., Staszewska, J., Tarnowska, M., Klećkowska-Nawrot, J., Goździewska-Harłajczuk, K., Kuźniarski, A., Gębarowski, T., & Janeczek, M. (2023). Functional Anatomy of the Thoracic Limb of the Komodo Dragon (Varanus komodoensis). Animals, 13(18), 2895. https://doi.org/10.3390/ani13182895