
Antioxidation Defenses of Apis mellifera Queens and Workers Respond to Imidacloprid in Different Age-Dependent Ways: Old Queens Are Resistant, Foragers Are Not





{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rearing of Workers
2.2. Rearing of Queens
2.3. Imidacloprid Contamination
2.4. Laboratory Procedures
2.5. Statistical Analysis
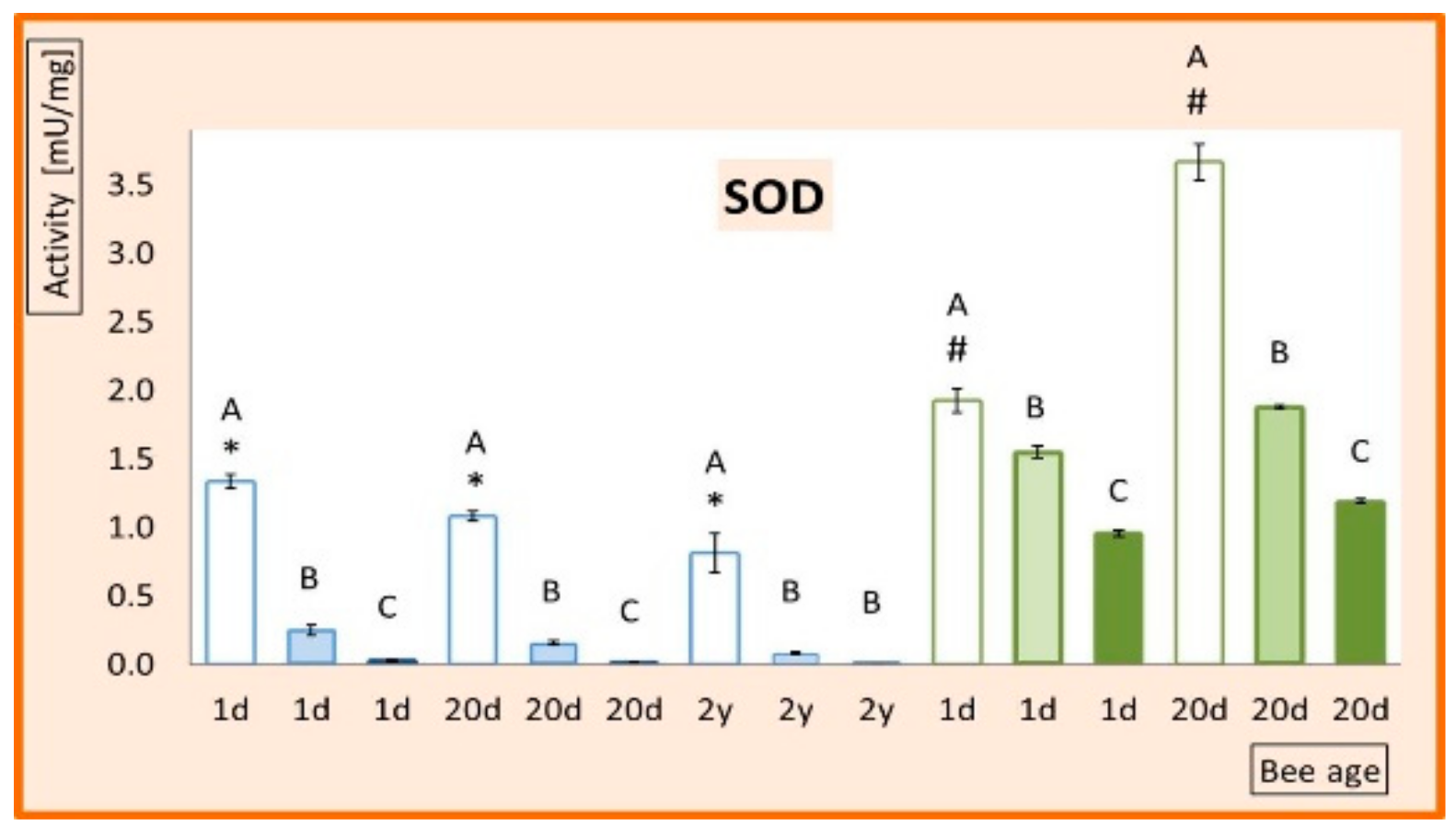
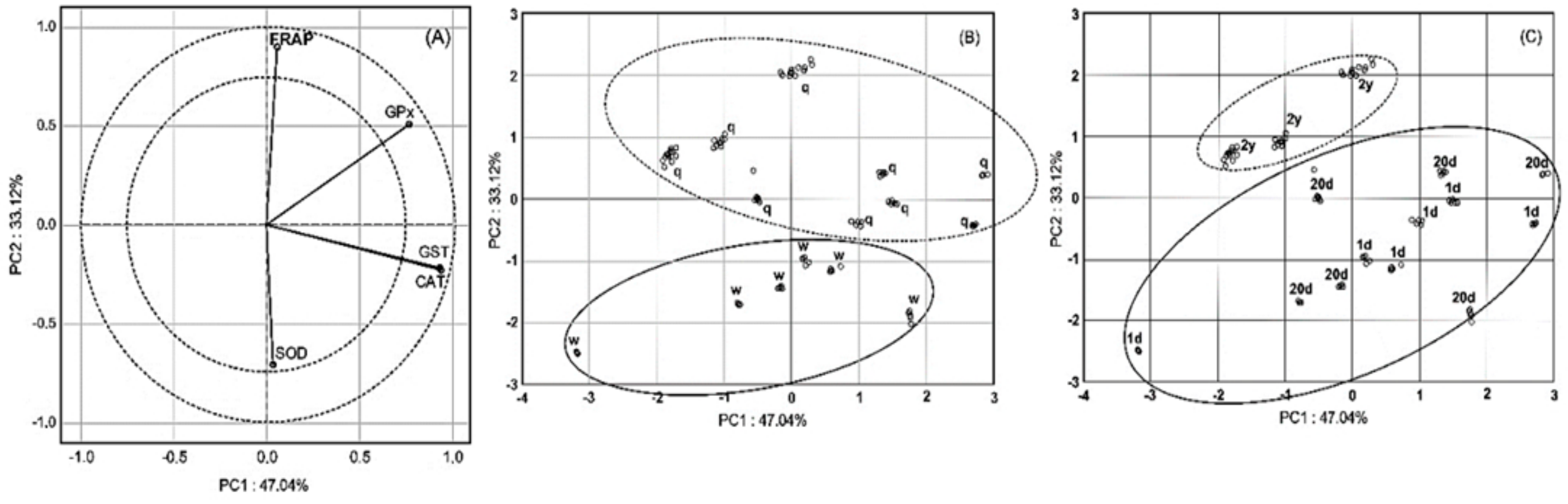
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Felton, G.W. Antioxidant defenses of invertebrates and vertebrates. In Oxidative Stress and Antioxidant Defenses in Biology, 1st ed.; Ahmad, S., Ed.; Chapman & Hall: New York, NY, USA, 1995; pp. 356–434. [Google Scholar]
- Kramer, B.H.; Nehring, V.; Buttstedt, A.; Heinze, J.; Korb, J.; Libbrech, R.; Meusemann, K.; Paxton, R.J.; Séguret, A.; Schaub, F.; et al. Oxidative stress and senescence in social insects: A significant but inconsistent link? Philos. Trans. R. Soc. B 2021, 376, 20190732. [Google Scholar] [CrossRef]
- Gregorc, A.; Alburaki, M.; Rinderer, N.; Sampson, B.; Knight, P.R.; Karim, S.; Adamczyk, J. Effects of coumaphos and imidacloprid on honey bee (Hymenoptera: Apidae) lifespan and antioxidant gene regulations in laboratory experiments. Sci. Rep. 2018, 8, 15003. [Google Scholar] [CrossRef] [PubMed]
- Muller, F.L.; Lustgarten, M.S.; Jang, Y.; Richardson, A.; Van Remmen, H. Trends in oxidative aging theories. Free. Radic. Biol. Med. 2007, 43, 477–503. [Google Scholar] [CrossRef] [PubMed]
- Münch, D.; Amdam, G.; Wolschin, F. Aging in a eusocial insect: Molecular and physiological characteristics of life span plasticity in the honey bee. Funct. Ecol. 2008, 22, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Liochev, S.I. Reactive oxygen species and the free radical theory of aging. Free. Radic. Biol. Med. 2013, 60, 1–4. [Google Scholar] [CrossRef]
- Costantini, D. Understanding diversity in oxidative status and oxidative stress: The opportunities and challenges ahead. J. Exp. Biol. 2019, 222, 13. [Google Scholar] [CrossRef]
- De Smet, L.; Hatjina, F.; Ioannidis, P.; Hamamtzoglou, A.; Schoonvaere, K.; Francis, F.; Meeus, I.; Smagghe, D.; de Graf, D.C. Stress indicator gene expression profiles, colony dynamics, and tissue development of honey bees exposed to sub-lethal doses of imidacloprid in laboratory and field experiments. PLoS ONE 2017, 12, e0171529. [Google Scholar] [CrossRef] [PubMed]
- Aurori, C.M.; Buttstedt, A.; Dezmirean, D.S.; Liviu, A.; Mărghitaş, L.A.; Robin, F.A.; Moritz, R.F.; Erler, S. What is the main driver of aging in long-lived winter honey bees: Antioxidant enzymes, innate immunity, or vitellogenin? J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Lemanski, N.J.; Bansal, S.; Fefferman, N.H. The sensitivity of a honey bee colony to worker mortality depends on season and resource availability. BMC Evol. Biol. 2020, 20, 139. [Google Scholar] [CrossRef]
- Dively, G.P.; Embrey, M.S.; Kamel, A.; Hawthorne, D.J.; Pettis, J.S. Assessment of chronic sublethal effects of imidacloprid on honey bee colony health. PLoS ONE 2015, 10, e0118748. [Google Scholar] [CrossRef]
- Mužinić, V.; Želježić, D. Non-target toxicity of novel insecticides. Arh. Hig. Rada Toksikol. 2018, 69, 86–102. [Google Scholar] [CrossRef]
- O’Neal, S.T.; Anderson, T.D.; Wu-Smart, J.Y. Interactions between pesticides and pathogen susceptibility in honey bees. Curr. Opin. Insect Sci. 2018, 26, 57–62. [Google Scholar] [CrossRef]
- Williams, G.R.; Troxler, A.; Retschnig, G.; Roth, K.; Yanez, O.; Shutle, D.; Neumann, P.; Gauthier, L. Neonicotinoid pesticides severely affect honey bee queens. Sci. Rep. 2015, 5, 14621. [Google Scholar] [CrossRef]
- Zhu, Y.C.; Yao, J.; Adamczyk, J.; Luttrell, R. Synergistic toxicity and physiological impact of imidacloprid alone and binary mixtures with seven representative pesticides on honey bee (Apis mellifera). PLoS ONE 2017, 12, e0176837. [Google Scholar] [CrossRef]
- Ciereszko, A.; Wilde, J.; Dietrich, G.J.; Siuda, M.; Bąk, B.; Judycka, S.; Karol, H. Sperm parameters of honey bee drone exposed to imidacloprid. Apidologie 2017, 48, 211–222. [Google Scholar] [CrossRef]
- Paleolog, J.; Wilde, J.; Siuda, M.; Bąk, B.; Wójcik, Ł.; Strachecka, A. Imidacloprid markedly affects hemolymph proteolysis, biomarkers, DNA global methylation, and the cuticle proteolytic layer in Western honey bees. Apidologie 2020, 51, 620–630. [Google Scholar] [CrossRef]
- Balieira, K.V.B.; Mazzo, M.; Bizerra, P.F.V.; Guimarães, A.R.D.J.S.; Nicodemo, D.; Mingatto, F.E. Imidacloprid-induced oxidative stress in honey bees and the antioxidant action of caffeine. Apidologie 2018, 49, 562–572. [Google Scholar] [CrossRef]
- Słowińska, M.; Nynca, J.; Wilde, J.; Bąk, B.; Siuda, M.; Ciereszko, A. Total antioxidant capacity of honey bee hemolymph in relation to age and exposure to pesticide, and comparison to antioxidant capacity of seminal plasma. Apidologie 2016, 47, 227–236. [Google Scholar] [CrossRef]
- Łoś, A.; Strachecka, A. Fast and cost-effective biochemical spectrophotometric analysis of solution of insect “blood” and body surface elution. Sensors 2018, 18, 1494. [Google Scholar] [CrossRef]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis, 2nd ed.; Bergmeyer, H.V., Ed.; Verlag Chemie/Academic Press Inc.: Weinheim, NY, USA, 1974; Volume 2, pp. 673–684. [Google Scholar]
- Chance, B.; Maehly, A. Assay of catalases and peroxidases. In Methods in Enzymology, 1st ed.; Colowick, S., Kaplan, N., Eds.; Academic Press Inc.: New York, NY, USA, 1955; pp. 764–775. [Google Scholar] [CrossRef]
- Warholm, M.; Guthenberg, C.; von Bahr, C.; Mannervik, B. Glutathione transferases from human liver. Method. Enzymol. 1985, 113, 499–504. [Google Scholar] [CrossRef]
- Podczasy, J.; Wei, R. Reduction of iodonitrotetrazolium violet by superoxide radicals. Biochem. Biophys. Res. Commun. 1988, 150, 1294–1301. [Google Scholar] [CrossRef]
- Benzie, I.; Strain, J. The ferric reducing ability of plasma (FRAP) as a measure of antioxidant power: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Strachecka, A.; Olszewski, K.; Paleolog, J. Varroa treatment with bromfenvinphos markedly suppresses honey bee biochemical defense levels. Entomol. Exp. Appl. 2016, 160, 57–71. [Google Scholar] [CrossRef]
- Załuski, R.; Bittarello, A.C.; Vieira, J.C.S.; Braga, C.P.; de Magalhaes Padilha, P.; da Silva Fernandes, M.; de Souza Bovi, T.; de Oliveira Orsi, R. Modification of the head proteome of nurse honey bees (Apis mellifera) exposed to field-relevant doses of pesticides. Sci. Rep. 2020, 10, 2190. [Google Scholar] [CrossRef] [PubMed]
- Nikolić, T.V.; Purać, J.; Orčić, S.; Kojić, D.; Vujanović, D.; Stanimirović, Z.; Gržetić, I.; Ilijević, K.; Šikoparija, B.; Blagojević, D.P. Environmental effects on superoxide dismutase and catalase activity and expression in honey bee. Arch. Insect Biochem. Physiol. 2015, 90, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Amdam, G.V. Social context, stress, and plasticity of aging. Aging Cell 2011, 10, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Pamplona, R.; Costantini, D. Molecular and structural antioxidant defenses against oxidative stress in animals. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R843–R863. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.; Carey, J.R. Hierarchy and connectedness as determinants of health and longevity in social insects. In Sociality, Hierarchy, Health. Comparative Biodemography: A Collection of Papers, 1st ed.; Weinstein, M., Lane, M.A., Eds.; National Academies Press: Washington, WA, USA, 2014; pp. 269–295. [Google Scholar]
- Margotta, J.W.; Robert, S.P.; Elekonich, M.M. Effects of flight activity and age on oxidative damage in the honey bee, Apis mellifera. J. Exp. Biol. 2018, 221, jeb183228. [Google Scholar] [CrossRef] [PubMed]
- Prezenská, M.; Sobeková, A.; Sabová, L. Antioxidant enzymes of honey bee larvae exposed to oxamyl. Folia Veterinaria 2019, 63, 9–14. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Goulson, D.; Pennacchio, F.; Nazzi, F.; Goka, K.; Desneux, N. Are bee diseases linked to pesticides? A brief review. Environ. Int. 2016, 89–90, 7–11. [Google Scholar] [CrossRef]
- Tarek, H.; Hamiduzzaman, M.M.; Morfin, N.; Guzman-Novoa, E. Sub-lethal doses of neonicotinoid and carbamate insecticides reduce the lifespan and alter the expression of immune health and detoxification related genes of honey bees (Apis mellifera). Mol. Res. 2018, 17, gmr16039908. [Google Scholar] [CrossRef]
- Corona, M.; Robinson, G.E. Genes of the antioxidant system of the honey bee: Annotation and phylogeny. Insect Mol. Biol. 2006, 15, 687–701. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Hsieh, Y.S. Oxidative stress decreases in the trophocytes and fat cells of worker honey bees during aging. Biogerontology 2014, 15, 129–137. [Google Scholar] [CrossRef]
- Rubio, C.P.; Hernández-Ruiz, J.; Martinez-Subiela, S.; Tvarijonaviciute, A.; Ceron, J.J. Spectrophotometric assays for total antioxidant capacity (TAC) in dog serum: An update. BMC Vet. Res. 2016, 12, 166. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Wang, T.M.; Vance, T.M.; Hurbert, P.; Dae-Ok, K.; Sung, I.; Koo, S.I.K.; Chun, O.K. Validation of analytical methods for plasma total antioxidant capacity by comparing with urinary 8-isoprostane level. J. Microbiol. Biotechnol. 2017, 27, 388–394. [Google Scholar] [CrossRef]
- Gätschenberger, H.; Azzami, K.; Tautz, J.; Beier, H. Antibacterial immune competence of honey bees (Apis mellifera) is adapted to different life stages and environmental risks. PLoS ONE 2013, 8, e66415. [Google Scholar] [CrossRef] [PubMed]
- Hulbert, A.J.; Pamplona, R.; Buffenstein, R.; Buttemer, W.A. Life and death: Metabolic rate, membrane composition, and life span of animals. Physiol. Rev. 2007, 87, 1175–1213. [Google Scholar] [CrossRef] [PubMed]
- Cocchi, M.; Lercker, G.; Frega, N.G.; Gabrielli, F.; Quaranta, M. Is a low concentration of linoleic acid related to the extended longevity of the queen honey bee? Prog. Nutr. 2019, 21, 729–734. [Google Scholar] [CrossRef]
- Park, H.G.; Lee, K.S.; Kim, B.Y.; Yoon, H.J.; Choi, S.Y.; Lee, K.Y.; Wan, H.; Li, J.; Jin, B.R. Honey bee (Apis cerana) vitellogenin acts as an antimicrobial and antioxidant agent in the body and venom. Dev. Comp. Immunol. 2018, 85, 51–61. [Google Scholar] [CrossRef]
- Corona, M.; Velarde, R.A.; Remolina, S.; Moran-Lauter, A.; Wang, Y.; Kimberly, A.; Hughes, A.K.; Robinson, G.E. Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity. Proc. Natl. Acad. Sci. USA 2007, 104, 7128–7133. [Google Scholar] [CrossRef] [PubMed]
- Amdam, G.V.; Ihle, K.; Page, R.E. Regulation of honey bee worker (Apis mellifera) life histories by vitellogenin. In Hormones, Brain, and Behavior, 2nd ed.; Pfaff, D.W., Arnold, A.P., Edgen, A.M., Fahrbach, S.E., Rubin, R.T., Eds.; Academic Press: San Diego, CA, USA, 2009; Volume 2, pp. 1003–1027. [Google Scholar]
- Santos, D.E.; Souza, A.O.; Tibério, G.J.; Alberici, L.C.; Hartfelder, K. Differential expression of antioxidant system genes in honey bee (Apis mellifera L.) caste development mitigates ROS-mediated oxidative damage in queen larvae. Genet. Mol. Biol. 2020, 43, e20200173. [Google Scholar] [CrossRef] [PubMed]
- Negroni, M.A.; Feldmeyer, B.; Foitzik, S. Experimental increase in fecundity causes upregulation of fecundity and body maintenance genes in the fat body of ant queens. Biol. Lett. 2021, 17, 20200909. [Google Scholar] [CrossRef]
- Kennedy, A.; Herman, J.; Rueppell, O. Reproductive activation in honeybee (Apis mellifera) workers protects against abiotic and biotic stress. Philos. Trans. R. Soc. B 2021, 376. [Google Scholar] [CrossRef]
- Tawfik, A.I.; Ahmed, Z.H.; Abdel-Rahman, M.F.; Moustafa, A.M. Influence of winter feeding on colony development and the antioxidant system of the honey bee, Apis mellifera. J. Apic. Res. 2020, 59, 752–763. [Google Scholar] [CrossRef]
- Harrison, M.C.; Hammond, R.L.; Mallon, E.B. Reproductive workers show queen-like gene expression in an intermediately eusocial insect, the buff-tailed bumble bee Bombus terrestris. Mol. Ecol. 2015, 24, 3043–3063. [Google Scholar] [CrossRef]
- Alonso-Alvarez, C. Oxidative stress as a life-history constraint: The role of reactive oxygen species in shaping phenotypes from conception to death. Funct. Ecol. 2010, 24, 984–996. [Google Scholar] [CrossRef]
- Fairbrother, A.; Purdy, J.; Anderson, T.; Fell, R. Risks of neonicotinoid insecticides to honey bees. Environ. Toxicol. Chem. 2014, 33, 719–731. [Google Scholar] [CrossRef]
- Sagona, S.; Betti, L.; Casini, L.; Palego, L.; Giannaccini, G.; Felicioli, A. Antioxidant enzymes activity during age polyethism in Apis mellifera L., 1758. J. Apic. Res. 2020, 59, 1–11. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paleolog, J.; Wilde, J.; Miszczak, A.; Gancarz, M.; Strachecka, A. Antioxidation Defenses of Apis mellifera Queens and Workers Respond to Imidacloprid in Different Age-Dependent Ways: Old Queens Are Resistant, Foragers Are Not. Animals 2021, 11, 1246. https://doi.org/10.3390/ani11051246
Paleolog J, Wilde J, Miszczak A, Gancarz M, Strachecka A. Antioxidation Defenses of Apis mellifera Queens and Workers Respond to Imidacloprid in Different Age-Dependent Ways: Old Queens Are Resistant, Foragers Are Not. Animals. 2021; 11(5):1246. https://doi.org/10.3390/ani11051246
Chicago/Turabian StylePaleolog, Jerzy, Jerzy Wilde, Artur Miszczak, Marek Gancarz, and Aneta Strachecka. 2021. "Antioxidation Defenses of Apis mellifera Queens and Workers Respond to Imidacloprid in Different Age-Dependent Ways: Old Queens Are Resistant, Foragers Are Not" Animals 11, no. 5: 1246. https://doi.org/10.3390/ani11051246
APA StylePaleolog, J., Wilde, J., Miszczak, A., Gancarz, M., & Strachecka, A. (2021). Antioxidation Defenses of Apis mellifera Queens and Workers Respond to Imidacloprid in Different Age-Dependent Ways: Old Queens Are Resistant, Foragers Are Not. Animals, 11(5), 1246. https://doi.org/10.3390/ani11051246