
The Effect of Different Feeding Systems on Salivary Cortisol Levels during Gestation in Sows on Herd Level


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Design
2.3. Management Routines
2.4. Saliva Collection
2.5. Data on Farrowing Sows
2.6. Recordings of Piglets
2.7. Salivary Cortisol Analysis
2.8. Data Management and Statistical Analysis
3. Results
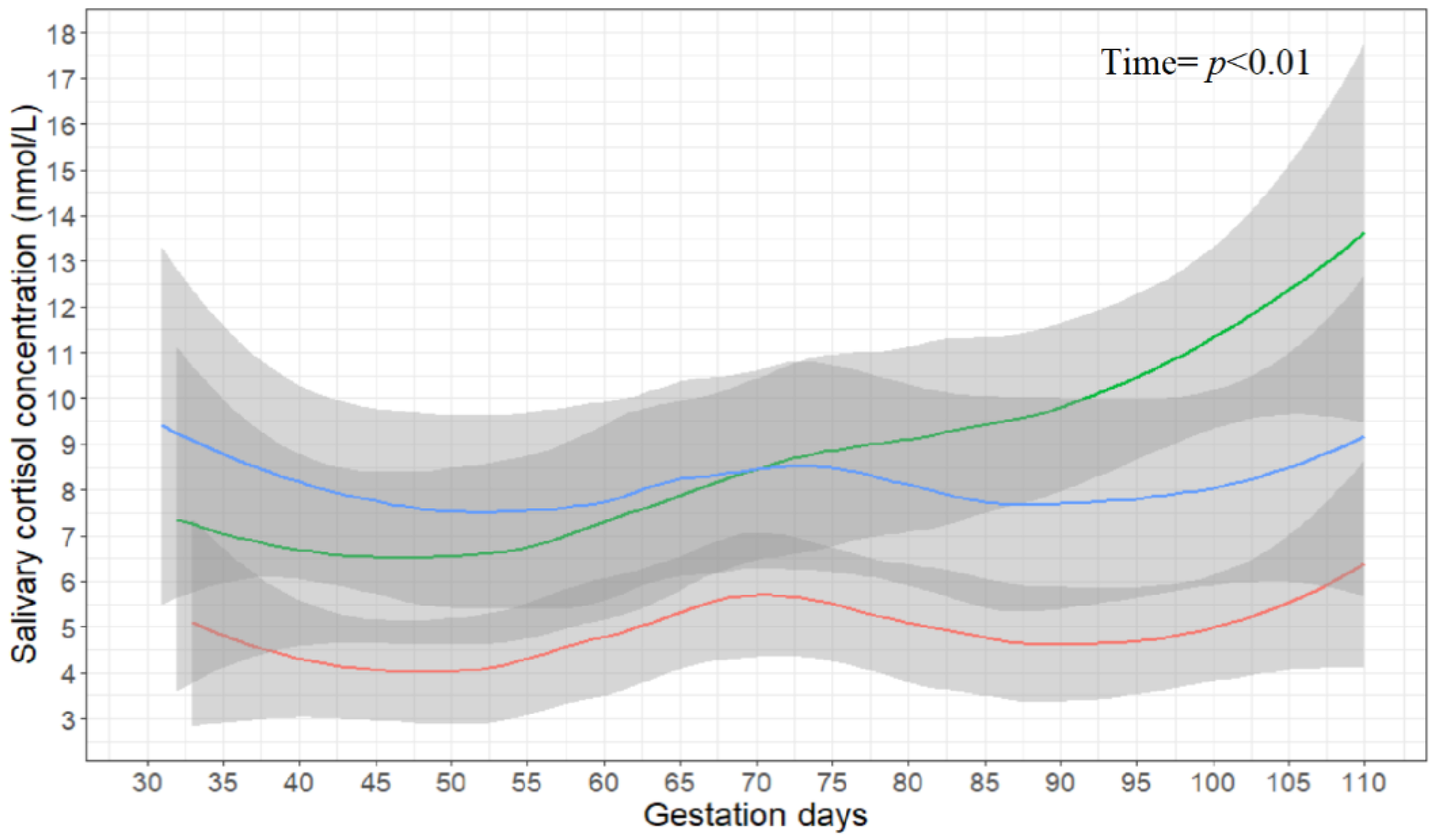
3.1. Salivary Cortisol Concentrations
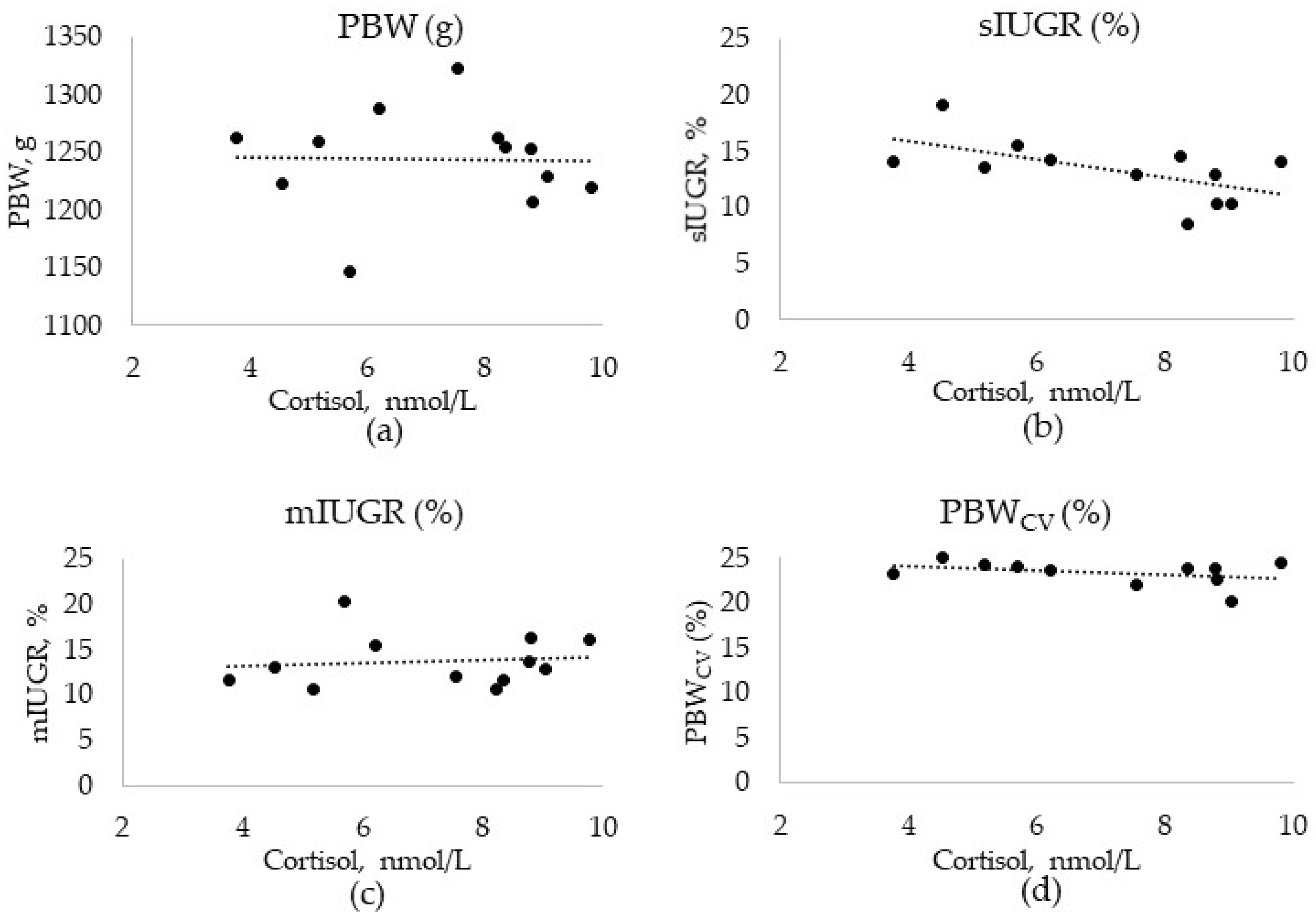
3.2. Production Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Milligan, B.N.; Fraser, D.; Kramer, D.L. Within-litter birth weight variation in the domestic pig and its relation to pre-weaning survival, weight gain, and variation in weaning weights. Livest. Prod. Sci. 2002, 76, 181–191. [Google Scholar] [CrossRef]
- Hansen, C.F.; Hales, J.; Amdi, C.; Moustsen, V.A. Intrauterine growth-restricted piglets defined by their head shape have impaired survival and growth during the suckling period. Anim. Prod. Sci. 2018, 59, 1056–1062. [Google Scholar] [CrossRef]
- Quiniou, N.; Dagorn, J.; Gaudré, D. Variation of piglets’ birth weight and consequences on subsequent performance. Livest. Prod. Sci. 2002, 78, 63–70. [Google Scholar] [CrossRef]
- Roza, S.J.; Steegers, E.A.P.; Verburg, B.O.; Jaddoe, V.W.V.; Moll, H.A.; Hofman, A.; Verhulst, F.C.; Tiemeier, H. What is spared by fetal brain-sparing? Fetal circulatory redistribution and behavioral problems in the general population. Am. J. Epidemiol. 2008, 168, 1145–1152. [Google Scholar] [CrossRef]
- Hales, J.; Moustsen, V.A.; Nielsen, M.B.F.; Hansen, C.F. Individual physical characteristics of neonatal piglets affect preweaning survival of piglets born in a noncrated system. J. Anim. Sci. 2013, 91, 4991–5003. [Google Scholar] [CrossRef] [PubMed]
- Amdi, C.; Krogh, U.; Flummer, C.; Oksbjerg, N.; Hansen, C.F.; Theil, P.K. Intrauterine growth restricted piglets defined by their head shape ingest insufficient amounts of colostrum. J. Anim. Sci. 2013, 91, 5605–5613. [Google Scholar] [CrossRef]
- Sjaastad, Ø.V.; Sand, O.; Hove, K. Physiology of Domestic Animals, 3rd ed.; Scandinavian Veterinary Press: Drøbak, Norway, 2016. [Google Scholar]
- Otten, W.; Kanitz, E.; Tuchscherer, M.; Schneider, F.; Brüssow, K.-P. Effects of adrenocorticotropin stimulation on cortisol dynamics of pregnant gilts and their fetuses: Implications for prenatal stress studies. Theriogenology 2004, 61, 1649–1659. [Google Scholar] [CrossRef] [PubMed]
- Amdi, C.; Giblin, L.; Hennessy, A.A.; Ryan, T.; Stanton, C.; Stickland, N.C.; Lawlor, P.G. Feed allowance and maternal backfat levels during gestation influence maternal cortisol levels, milk fat composition and offspring growth. J. Nutr. Sci. 2013, 2, e1. [Google Scholar] [CrossRef] [PubMed]
- Chiesa, C.; Osborn, J.F.; Haass, C.; Natale, F.; Spinelli, M.; Scapillati, E.; Spinelli, A.; Pacifico, L. Ghrelin, leptin, IGF-1, IGFBP-3, and insulin concentrations at birth: Is there a relationship with fetal growth and neonatal anthropometry? Clin. Chem. 2008, 54, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Roelofs, S.; Godding, L.; de Haan, J.R.; van der Staay, F.J.; Nordquist, R.E. Effects of parity and litter size on cortisol measures in commercially housed sows and their offspring. Physiol. Behav. 2019, 201, 83–90. [Google Scholar] [CrossRef]
- Strawford, M.L.; Li, Y.Z.; Gonyou, H.W. The effect of management strategies and parity on the behaviour and physiology of gestating sows housed in an electronic sow feeding system. Can. J. Anim. Sci. 2008, 88, 559–567. [Google Scholar] [CrossRef]
- Anil, L.; Anil, S.S.; Deen, J.; Baidoo, S.K.; Wheaton, J.E. Evaluation of well-being, productivity, and longevity of pregnant sows housed in groups in pens with an electronic sow feeder or separately in gestation stalls. Am. J. Vet. Res. 2005, 66, 1630–1638. [Google Scholar] [CrossRef]
- Holt, J.P.; Johnston, L.J.; Baidoo, S.K.; Shurson, G.C. Effects of a high-fiber diet and frequent feeding on behavior, reproductive performance, and nutrient digestibility in gestating sows. J. Anim. Sci. 2006, 84, 946–955. [Google Scholar] [CrossRef]
- Merlot, E.; Pastorelli, H.; Prunier, A.; Père, M.C.; Louveau, I.; Lefaucheur, L.; Perruchot, M.H.; Meunier-Salaün, M.C.; Gardan-Salmon, D.; Gondret, F.; et al. Sow environment during gestation: Part I. Influence on maternal physiology and lacteal secretions in relation with neonatal survival. Animal 2018, 13, 1432–1439. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.C.; Hong, J.S.; Jin, S.S.; Kim, Y.Y. Comparing gestating sows housing between electronic sow feeding system and a conventional stall over three consecutive parities. Livest. Sci. 2017, 199, 37–45. [Google Scholar] [CrossRef]
- Kranendonk, G.; Hopster, H.; Fillerup, M.; Ekkel, E.D.; Mulder, E.J.H.; Wiegant, V.M.; Taverne, M.A.M. Lower birth weight and attenuated adrenocortical response to ACTH in offspring from sows that orally received cortisol during gestation. Domest. Anim. Endocrinol. 2006, 30, 218–238. [Google Scholar] [CrossRef] [PubMed]
- Tybirk, P.; Sloth, N.M.; Kjelsen, N.J.; Vinther, J. Normer for Næringsstoffer; SEGES Svineproduktion: Copenhagen, Denmark, 2019; pp. 1–14. [Google Scholar]
- Kirschbaum, C.; Hellhammer, D.H. Salivary cortisol in psychobiological research: An overview. Neuropsychobiology 1989, 22, 150–169. [Google Scholar] [CrossRef]
- Cook, N.J.; Schaefer, A.L.; Lepage, P.; Jones, S.M. Salivary vs. serum cortisol for the assessment of adrenal activity in swine. Can. J. Anim. Sci. 1996, 76, 329–335. [Google Scholar] [CrossRef]
- Chevaux, E.; Sacy, A.; Le Treut, Y.; Martineau, G. IntraUterine growth retardation (IUGR): Morphological and behavioural description. In Proceedings of the 21st IPVS Congress, Vancouver, BC, Canada, 18–21 July 2010. [Google Scholar]
- Engelsmann, M.N.; Hansen, C.F.; Nielsen, M.N.; Kristensen, A.R.; Amdi, C. Glucose injections at birth, warmth and placing at a nurse sow improve the growth of IUGR piglets. Animals 2019, 9, 519. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2014, 67. [Google Scholar] [CrossRef]
- Lenth, R. Least-squares means: The R package lsmeans. J. Stat. Softw. 2016, 69. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Meunier-Salaün, M.C.; Gort, F.; Prunier, A.; Schouten, W.P.G. Behavioural patterns and progesterone, cortisol and prolactin levels around parturition in European (Large-White) and Chinese (Meishan) sows. Appl. Anim. Behav. Sci. 1991, 31, 43–59. [Google Scholar] [CrossRef]
- Amdi, C.; Moustsen, V.A.; Oxholm, L.C.; Baxter, E.M.; Sørensen, G.; Eriksson, K.B.; Diness, L.H.; Nielsen, M.F.; Hansen, C.F. Comparable cortisol, heart rate and milk let-down in nurse sows and non-nurse sows. Livest. Sci. 2017, 198, 174–181. [Google Scholar] [CrossRef]
- Haussmann, M.F.; Carroll, J.A.; Weesner, G.D.; Daniels, M.J.; Matteri, R.L.; Lay, D.C., Jr. Administration of ACTH to restrained, pregnant sows alters their pigs’ hypothalamic-pituitary-adrenal (HPA) axis. J. Anim. Sci. 2000, 78, 2399–2411. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, S.; D’Eath, R.B.; Robson, S.K.; Lawrence, A.B. The effect of confinement during lactation on the hypothalamic-pituitary-adrenal axis and behaviour of primiparous sows. Physiol. Behav. 2006, 87, 345–352. [Google Scholar] [CrossRef]
- Lay, D.C., Jr.; Kattesh, H.G.; Cunnick, J.E.; Daniels, M.J.; McMunn, K.A.; Toscano, M.J.; Roberts, M.P. Prenatal stress effects on pig development and response to weaning. J. Anim. Sci. 2008, 86, 1316–1324. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, S.; Moinard, C.; Robson, S.K.; Baxter, E.; Ormandy, E.; Douglas, A.J.; Seckl, J.R.; Russell, J.A.; Lawrence, A.B. Programming the offspring of the pig by prenatal social stress: Neuroendocrine activity and behaviour. Horm. Behav. 2006, 49, 68–80. [Google Scholar] [CrossRef]
- Fountas, A.; Karavitaki, N. Hypopituitarism, causes, diagnosis, management and mortality. In Encyclopedia of Endocrine Diseases, 2nd ed.; Huhtaniemi, I., Martini, L., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 301–313. [Google Scholar]
- Murani, E.; Reyer, H.; Ponsuksili, S.; Fritschka, S.; Wimmers, K. A substitution in the ligand binding domain of the porcine glucocorticoid receptor affects activity of the adrenal gland. PLoS ONE 2012, 7, e45518. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Li, H.; Liang, T.; Zhou, B.; Chu, Q.; Schinckel, A.P.; Yang, X.; Zhao, R.; Li, P.; Huang, R. Stocking density affects welfare indicators of growing pigs of different group sizes after regrouping. Appl. Anim. Behav. Sci. 2016, 174, 42–50. [Google Scholar] [CrossRef]
- Hemsworth, P.H.; Morrison, R.S.; Tilbrook, A.J.; Butler, K.L.; Rice, M.; Moeller, S.J. Effects of varying floor space on aggressive behavior and cortisol concentrations in group-housed sows. J. Anim. Sci. 2016, 94, 4809–4818. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, P.H.; Rice, M.; Nash, J.; Giri, K.; Butler, K.L.; Tilbrook, A.J.; Morrison, R.S. Effects of group size and floor space allowance on grouped sows: Aggression, stress, skin injuries, and reproductive performance. J. Anim. Sci. 2013, 91, 4953–4964. [Google Scholar] [CrossRef] [PubMed]
- Merlot, E.; Calvar, C.; Prunier, A. Influence of the housing environment during sow gestation on maternal health, and offspring immunity and survival. Anim. Prod. Sci. 2017, 57, 1751–1758. [Google Scholar] [CrossRef]
- Quesnel, H.; Père, M.C.; Louveau, I.; Lefaucheur, L.; Perruchot, M.H.; Prunier, A.; Pastorelli, H.; Meunier-Salaün, M.C.; Gardan-Salmon, D.; Merlot, E.; et al. Sow environment during gestation: Part II. Influence on piglet physiology and tissue maturity at birth. Animal 2018, 13, 1440–1447. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Stall | Floor | ESF | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Herd | A | C | D | I | B | E | H | K | F | G | J | L |
| Herd size, no. of sows | 1900 | 1250 | 1050 | 1150 | 3050 | 2500 | 1250 | 2000 | 800 | 1200 | 1400 | 1700 |
| Health status 1 | Myc + Ap12 | Myc + Ap6 + Ap12 | Myc | Myc + Ap6 + Ap12 | Myc + PRRS1 + PRRS2 | SPF | Myc + Ap12 | Unknown | Myc + PRRS2 | Myc + Ap12 | Myc + Ap2 + PRRS1 + PRRS2 | Unknown |
| Feedings/day, no. | 1–2 | 1–2 | 2 | 2 | 1 | 2 | 1 | 1 | - | - | - | - |
| Sows/pen in the gestation unit, no. | 60–65 | 50 | 46 | 25–40 | 30–56 | 50–60 | 16 or 40–45 2 | 18 | 45–60 | 60–65 | 48–64 | 75 |
| Feed System | Parity 0–1 | Parity 2–3 | Parity 4–5 | Parity ≥ 6 | ||||
|---|---|---|---|---|---|---|---|---|
| Lsmean | SE | Lsmean | SE | Lsmean | SE | Lsmean | SE | |
| Stall (nmol/L) | 4.97 | 1.15 | 3.60 | 1.13 | 4.21 | 1.15 | 3.03 | 1.30 |
| Floor (nmol/L) | 7.45 | 1.14 | 7.36 | 1.15 | 6.71 | 1.18 | 5.77 | 1.44 |
| ESF (nmol/L) | 7.61 | 1.14 | 6.09 | 1.14 | 5.03 | 1.17 | 8.62 | 1.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bahnsen, I.; Riddersholm, K.V.; de Knegt, L.V.; Bruun, T.S.; Amdi, C. The Effect of Different Feeding Systems on Salivary Cortisol Levels during Gestation in Sows on Herd Level. Animals 2021, 11, 1074. https://doi.org/10.3390/ani11041074
Bahnsen I, Riddersholm KV, de Knegt LV, Bruun TS, Amdi C. The Effect of Different Feeding Systems on Salivary Cortisol Levels during Gestation in Sows on Herd Level. Animals. 2021; 11(4):1074. https://doi.org/10.3390/ani11041074
Chicago/Turabian StyleBahnsen, Ida, Kristina V. Riddersholm, Leonardo V. de Knegt, Thomas S. Bruun, and Charlotte Amdi. 2021. "The Effect of Different Feeding Systems on Salivary Cortisol Levels during Gestation in Sows on Herd Level" Animals 11, no. 4: 1074. https://doi.org/10.3390/ani11041074
APA StyleBahnsen, I., Riddersholm, K. V., de Knegt, L. V., Bruun, T. S., & Amdi, C. (2021). The Effect of Different Feeding Systems on Salivary Cortisol Levels during Gestation in Sows on Herd Level. Animals, 11(4), 1074. https://doi.org/10.3390/ani11041074