
Expression Profiles of the Progesterone Receptor, Cyclooxygenase-2, Growth Differentiation Factor 9, and Bone Morphogenetic Protein 15 Transcripts in the Canine Oviducts during the Oestrous Cycle



Abstract
Simple Summary
Abstract
1. Introduction
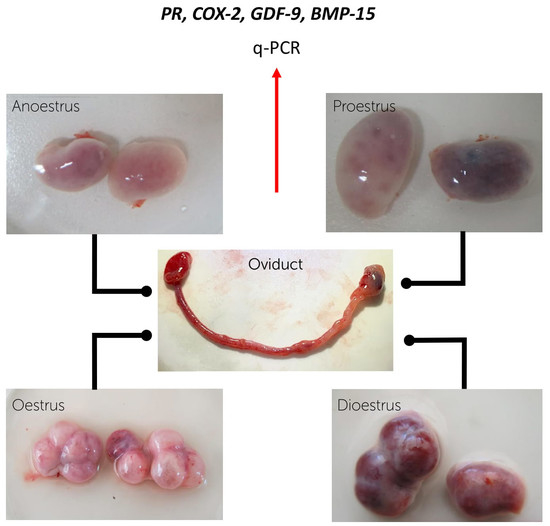
2. Materials and Methods
2.1. Sample Collection
2.2. Recovery of the Oviductal Cells
2.3. RNA Extraction and Quantitative Real-Time RT-qPCR Analysis
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wollenhaupt, K.; Tomek, W.; Brüssow, K.P.; Tiemann, U.; Viergutz, T.; Schneider, F.; Nürnberg, G. Effects of ovarian steroids and epidermal growth factor (EGF) on expression and bioactivation of specific regulators of transcription and translation in oviductal tissue in pigs. Reproduction 2002, 123, 87–96. [Google Scholar] [CrossRef]
- Cerny, K.L.; Garrett, E.; Walton, A.J.; Anderson, L.H.; Bridges, P.J. A transcriptomal analysis of bovine oviductal epithelial cells collected during the follicular phase versus the luteal phase of the estrous cycle. Reprod. Biol. Endocrinol. 2015, 13, 84. [Google Scholar] [CrossRef] [PubMed]
- Barton, B.E.; Herrera, G.G.; Anamthathmakula, P.; Rock, J.K.; Willie, A.M.; Harris, E.A.; Takemaru, K.; Winuthayanon, W. Roles of steroid hormones in oviductal function. Reproduction 2020, 159, R125–R137. [Google Scholar] [CrossRef] [PubMed]
- Hermoso, M.; Barrera, N.; Morales, B.; Perez, S.; Villalon, M. Platelet-activating factor increases ciliary activity in the hamster oviduct through epithelial production of prostaglandin E2. Pflugers Arch. 2001, 442, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Gauvreau, D.; Moisan, V.; Roy, M.; Fortier, M.A.; Bilodeau, J.F. Expression of prostaglandin E synthases in the bovine oviduct. Theriogenology 2010, 73, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Wijayagunawardane, M.P.; Kodithuwakku, S.P.; Yamamoto, D.; Miyamoto, A. Vascular endothelial growth factor system in the cow oviduct: Possible involvement in the regulation of the oviductal motility and embryo transport. Mol. Reprod. Dev. 2005, 72, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Martínez, S.; Hermoso, M.; Farina, M.; Ribeiro, M.L.; Rapanelli, M.; Espinosa, M.; Villalón, M.; Franchi, A. 17-β-Estradiol upregulates COX-2 in the rat oviduct. Prostaglandins Lipid Mediat. 2006, 80, 155–164. [Google Scholar]
- Hunter, R.H. Components of oviduct physiology in eutherian mammals. Biol. Rev. Camb. Philos. Soc. 2012, 87, 244–255. [Google Scholar] [CrossRef]
- Concannon, P.W. Endocrinologic control of normal canine ovarian function. Reprod. Domest. Anim. 2009, 44 (Suppl. 2), 3–15. [Google Scholar] [CrossRef]
- Hardy, D.B.; Janowski, B.A.; Corey, D.R.; Mendelson, C.R. Progesterone receptor plays a major anti-inflammatory role in human myometrial cells by antagonism of nuclear factor-B activation of cyclooxygenase 2 expressions. Mol. Endocrinol. 2006, 20, 2724–2733. [Google Scholar] [CrossRef]
- Akison, L.K.; Boden, M.J.; Kennaway, D.J.; Russell, D.L.; Rocker, R.L. Progesterone receptor-dependent regulation of genes in the oviducts of female mice. Physiol. Genom. 2014, 46, 583–592. [Google Scholar] [CrossRef]
- Kaya, H.S.; Hantak, A.M.; Stubbs, L.J.; Taylor, R.N.; Bagchi, I.C.; Bagchi, M.K. Roles of progesterone receptor A and B isoforms during human endometrial decidualization. Mol. Endocrinol. 2015, 29, 882–895. [Google Scholar] [CrossRef]
- Teilmann, S.C.; Clement, C.A.; Thorup, J.; Byskov, A.G.; Christensen, S.T. Expression and localization of the progesterone receptor in mouse and human reproductive organs. J. Endocrinol. 2006, 191, 525–535. [Google Scholar] [CrossRef]
- Akison, L.K.; Robker, R.L. The critical roles of progesterone receptor (PGR) in ovulation, oocyte developmental competence and oviductal transport in mammalian reproduction. Reprod. Domest. Anim. 2012, 47 (Suppl. 4), 288–296. [Google Scholar] [CrossRef]
- Tahir, M.; Reynaud, K.; Grimard, B.; Thoumire, S.; Chastant-Maillard, S.; Saint-Dizier, M. Expression of nuclear and membrane progesterone receptors in the canine oviduct during the periovulatory period. Reprod. Fertil. Dev. 2013, 25, 1065–1076. [Google Scholar] [CrossRef]
- Kim, J.; Bagchi, I.C.; Bagchi, M.K. Control of ovulation in mice by progesterone receptor-regulated gene networks. Mol. Hum. Reprod. 2009, 15, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.L.; DeWitt, D.L.; Garavito, R.M. Cyclooxygenases: Structural, cellular, and molecular biology. Annu. Rev. Biochem. 2000, 69, 145–182. [Google Scholar] [CrossRef] [PubMed]
- Tai, H.H.; Ensor, C.M.; Tong, M.; Zhou, H.; Yan, F. Prostaglandin catabolizing enzymes. Prostaglandins Lipid Mediat. 2002, 68–69, 483–493. [Google Scholar] [CrossRef]
- Odau, S.; Gabler, C.; Holder, C.; Einspanie, R. Differential expression of cyclooxygenase 1 and cyclooxygenase 2 in the bovine oviduct. J. Endocrinol. 2006, 191, 263–274. [Google Scholar] [CrossRef]
- Kowalewski, M.P.; Fox, B.; Gram, A.; Boos, A.; Reichler, I. Prostaglandin E2 functions as a luteotrophic factor in the dog. Reproduction 2013, 145, 213–226. [Google Scholar] [CrossRef]
- De los Reyes, M.; Araujo, A.; Flores, J.; Ramirez, G.; Palomino, J.; Parraguez, V.H.; Aspee, K. Cyclooxygenase-2 messenger RNA levels in canine follicular cells: Interrelationship with GDF-9, BMP-15 and progesterone. Domest. Anim. Endocrinol. 2021, 74, 106529. [Google Scholar] [CrossRef]
- Takahashi, T.; Jason, D.; Morrow, J.D.; Wang, H.; Dey, S.K. Cyclooxygenase-2-derived Prostaglandin E2 Directs Oocyte Maturation by Differentially Influencing Multiple Signaling Pathways. J. Biol. Chem. 2007, 281, 37117–37129. [Google Scholar] [CrossRef]
- Takahashi, T.; Igarashi, H.; Amita, M.; Hara, S.; Kurachi, H. Roles of prostaglandins during oocyte maturation: Lessons from Knockout mice. J. Mamm. Ova Res. 2010, 27, 11–20. [Google Scholar] [CrossRef]
- Viggiano, M.; Cebral, E.; Gimeno, A.L.; Gimeno, M.A. Probable influence of ova and embryo prostaglandins in the differential ovum transport in pregnant and cycling rats. Prostaglandins Leukot. Essent. Fat. Acids 1992, 45, 211–215. [Google Scholar] [CrossRef]
- Zhao, Y.; Chegini, N.; Flanders, K.C. Human fallopian tube expresses transforming growth factor (TGF beta) isoforms, TGF beta type I-III receptor messenger ribonucleic acid and protein, and contains [125I] TGF beta-binding sites. J. Clin. Endocrinol. Metab. 1994, 79, 1177–1184. [Google Scholar] [PubMed]
- Lee, S.H.; Oh, H.J.; Kim, M.J.; Kim, G.A.; Choi, Y.B.; Jo, Y.K.; Nugraha, E.M.; Setyawan, N.; Lee, B.C. Oocyte maturation-related gene expression in the canine oviduct, cumulus cells, and oocytes and effect of co-culture with oviduct cells on in vitro maturation of oocytes. J. Assist. Reprod. Genet. 2017, 34, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.J. Oocyte regulation of metabolic cooperativity between mouse cumulus cells and oocytes: BMP15 and GDF9 control cholesterol biosynthesis in cumulus cells. Development 2008, 135, 111–121. [Google Scholar]
- Chang, H.M.; Qiao, J.; Leung, P.C. Oocyte-somatic cell interactions in the human ovary-novel role of bone morphogenetic proteins and growth differentiation factors. Hum. Reprod. Update 2016, 23, 1–18. [Google Scholar] [CrossRef]
- Belli, M.; Shimasaki, S. Molecular aspects and clinical Relevance of GDF9 and BMP15 in ovarian function. Vitam. Horm. 2018, 107, 317–348. [Google Scholar]
- Garcia, P.; Aspee, K.; Ramirez, G.; Dettleff, P.; Palomino, J.; Peralta, O.A.; Parraguez, V.H.; De los Reyes, M. Influence of growth differentiation factor 9 and bone morphogenetic protein 15 on in vitro maturation of canine oocytes. Reprod. Domest. Anim. 2019, 54, 373–380. [Google Scholar] [CrossRef]
- Elvin, J.A.; Clarck, A.T.; Wang, P.; Wolfman, N.M.; Matzuk, M.M. Paracrine Actions of Growth Differentiation Factor-9 in the mammalian ovary. Mol. Endocrinol. 1999, 13, 1035–1048. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, J.S.A.; Vannucchi, C.I.; Braga, F.C.; Paula-Lopes, F.F.; Milazzotto, M.P.; Assumpcao, M.E.O.A.; Visintin, J.A. Oestrogen and Progesterone receptor gene expression in canine oocytes and cumulus cells throughout the oestrous cycle. Reprod. Domest. Anim. 2009, 44 (Suppl. 2), 239–242. [Google Scholar] [CrossRef] [PubMed]
- Palomino, J.; De los Reyes, M. Temporal expression of GDF-9 and BMP-15 in canine ovarian follicles. Theriogenology 2016, 86, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, G.; Palomino, J.; Aspee, K.; De los Reyes, M. GDF-9 and BMP-15 mRNA levels in canine cumulus cells related to cumulus expansion and the maturation process. Animals 2020, 10, 462. [Google Scholar] [CrossRef] [PubMed]
- Brugger, N.; Otzdorff, C.; Walter, B.; Hoffmann, B.; Braun, J. Quantitative determination of progesterone (P4) in canine blood serum using an enzyme-linked fluorescence assay. Reprod. Domest. Anim. 2011, 46, 870–873. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed]
- Fuentealba, B.; Nieto, M.; Croxatto, H.B. Estrogen and Progesterone Receptors in the Oviduct during Egg Transport in Cyclic and Pregnant Rats. Biol. Reprod. 1988, 9, 751–757. [Google Scholar] [CrossRef]
- Vermeirsch, H.; Van Den Broec, W.; Coryn, M.; Simoens, P. Immunolocalization of sex steroid hormone receptors in the canine uterine tube and their relation to sex steroid hormone concentrations levels. Reprod. Fertil. Dev. 2002, 14, 241–250. [Google Scholar] [CrossRef]
- Slobodin, B.; Han, R.; Calderone, V.; Oude, V.; Loayza-Puch, F.; Elkon, R.; Agami, R. Transcription Impacts the Efficiency of mRNA Translation via co-transcriptional N6-adenosine Methylation. Cell 2017, 169, 326–337. [Google Scholar] [CrossRef]
- Pasqualini, J.R.; Nguyen, B.L. Progesterone receptors in the foetal uterus of guinea-pig: Its stimulation after oestradiol treatment. Endocrinology 1980, 106, 1160–1165. [Google Scholar] [CrossRef]
- Fortune, J.E.; Willis, E.L.; Bridges, P.J.; Yang, C.S. The periovulatory period in cattle: Progesterone, prostaglandins, oxytocin and ADAMTS proteases. Anim. Reprod. 2009, 6, 60–71. [Google Scholar] [PubMed]
- Reynaud, K.; Saint-Dizier, M.; Tahir, M.Z.; Havard, T.; Harichaux, G.; Labas, V.; Thoumire, S.; Fontbonne, A.; Grimard, B.; Chastant-Maillard, S. Progesterone plays a critical role in canine oocyte maturation and fertilization. Biol. Reprod. 2015, 93, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Park, K.B.; Choi, E.J.; Hyun, S.H.; Kim, N.H.; Jeong, Y.W.; Hwang, W.S. Relationship between time post-ovulation and progesterone on oocyte maturation and pregnancy in canine cloning. Anim. Reprod. Sci. 2018, 185, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Morrill, G.A.; Chien, E.J.; Kostellow, A.B. Progesterone induction of phospholipid methylation and arachidonic acid turnover during the first meiotic division in amphibian oocytes. Life Sci. 1986, 27, 1501–1508. [Google Scholar]
- Wanggren, K.; Lalitkumar, P.G.; Stavreus-Evers, A.; Stabi, B.; Gemzell-Danielsson, K. Prostaglandin E2 and F2alpha receptors in the human Fallopian tube before and after mifepristone treatment. Mol. Hum. Reprod. 2006, 12, 577–585. [Google Scholar] [CrossRef]
- Herschman, H.R. Review: Prostaglandin synthase-2. Biochim. Biophys. Acta 1996, 1299, 125–140. [Google Scholar] [CrossRef]
- Sirois, J.; Simmons, D.L.; Richards, J.S. Hormonal regulation of messenger ribonucleic acid encoding a novel isoform of prostaglandin endo-peroxide H synthase in rat preovulatory follicles Induction in vivo and in vitro. J. Biol. Chem. 1992, 267, 11586–11592. [Google Scholar] [CrossRef]
- Bridges, P.J.; Komar, C.M.; Fortune, J.E. Gonadotropin-Induced Expression of Messenger Ribonucleic Acid for Cyclooxygenase-2 and Production of Prostaglandins E and F2a in Bovine Preovulatory Follicles Are Regulated by the Progesterone Receptor. Endocrinology 2006, 147, 4713–4722. [Google Scholar] [CrossRef][Green Version]
- Salleh, N. Diverse Roles of Prostaglandins in Blastocyst Implantation. Sci. World J. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.Y.; Di, R.; Tang, Q.Q.; Jin, H.H.; Chu, M.X.; Huang, D.W.; He, J.N.; Liu, Q.Y.; Hu, W.P.; Wang, X.Y.; et al. Tissue-specific mRNA expression profiles of GDF9, BMP15, and BMPR1B genes in prolific and non-prolific goat breeds. Czech J. Anim. Sci. 2015, 60, 452–458. [Google Scholar] [CrossRef]
- Hussein, T.S.; Sutton-McDowall, M.L.; Gilchrist, R.B.; Thompson, J.G. Temporal effects of exogenous oocyte-secreted factors on bovine oocyte developmental competence during IVM. Reprod. Fertil. Dev. 2011, 23, 576–584. [Google Scholar] [CrossRef]
- Yeo, C.X.; Gilchrist, R.B.; Thompson, J.G.; Lane, M. Exogenous growth differentiation factor 9 in oocyte maturation media enhances subsequent embryo development and fetal viability in mice. Hum. Reprod. 2008, 23, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Elvin, J.A.; Yan, C.; Matzuk, M.M. Growth differentiation factor-9 stimulates progesterone synthesis in granulosa cells via a prostaglandin E2-EP2 receptor pathway. Proc. Natl. Acad. Sci. USA 2000, 97, 10288–10293. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Arvisais, E.W.; Davis, J.S. Luteinizing hormone stimulates mammalian target of rapamycin signalling in bovine luteal cells via pathways independent of Akt and Mitogen-activated protein kinase: Modulation of glycogen synthase kinase 3 and amp-activated protein kinase. Endocrinology 2010, 151, 2846–2857. [Google Scholar] [CrossRef] [PubMed]
- Norris, R.P.; Freudzon, M.; Mehlmann, L.M.; Cowan, A.E.; Simon, A.M.; Paul, D.; Jaffe, L.A. Luteinizing hormone causes MAP kinase-dependent phosphorylation and closure of connexin 43 gap junctions in mouse ovarian follicles: One of two paths to meiotic resumption. Development 2008, 135, 3229–3238. [Google Scholar] [CrossRef] [PubMed]
- Mester, B.; Ritter, L.; Pitman, J.L.; Bibby, A.H.; Gilchrist, R.B.; McNatty, K.P.; Juengel, J.L.; McIntosh, C.J. Oocyte expression, secretion and somatic cell interaction of mouse bone morphogenetic protein 15 during the peri-ovulatory period. Reprod. Fertil. Dev. 2014, 27, 801–811. [Google Scholar] [CrossRef]
- Yoshino, O.; Osuga, J.S.; Harada, M.; Nishii, O.Y.; Taketani, Y. The function of bone morphogenetic proteins in the human ovary. Reprod. Med. Biol. 2011, 10, 1–7. [Google Scholar] [CrossRef]
- Garciía, E.V.; Valdecantos, P.A.; Barrera, D.; Roldaán-Olarte, M.; Miceli, D.C. Bone morphogenetic proteins in the bovine oviduct: Differential expression of BMP-5 in the isthmus during the estrous cycle. Theriogenology 2014, 81, 1032–1041. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence 5′-3′ | Size (bp) | Reference |
|---|---|---|---|
| ACTB | F:ATTGTCATGGACTCTGGGGATG R:TCCTTGATGTCACGCACGAT | 191 | [33] |
| GDF-9 | F: CAGAAGGGAGGTCTGTCTGC R: TGTTGGGGGAAAAGAAAGTG | 170 | [33] |
| BMP-15 | F: CCCTGCCCCTGATTCGGGAG R: CCGCAAAGGATGCCCAAGGAC | 82 | [33] |
| COX-2 | F:TGAGCGGTTATTCCAGACGAGCAG R:CCAACCCCGCAGCCATTTCCTTCT | 500 | [21] |
| PGR | F:GATGCTATATTTTGCACCTGA R: CTCCTTTTTGCCTCAAGCCA | 266 | [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palomino, J.; Flores, J.; Ramirez, G.; Parraguez, V.H.; De los Reyes, M. Expression Profiles of the Progesterone Receptor, Cyclooxygenase-2, Growth Differentiation Factor 9, and Bone Morphogenetic Protein 15 Transcripts in the Canine Oviducts during the Oestrous Cycle. Animals 2021, 11, 454. https://doi.org/10.3390/ani11020454
Palomino J, Flores J, Ramirez G, Parraguez VH, De los Reyes M. Expression Profiles of the Progesterone Receptor, Cyclooxygenase-2, Growth Differentiation Factor 9, and Bone Morphogenetic Protein 15 Transcripts in the Canine Oviducts during the Oestrous Cycle. Animals. 2021; 11(2):454. https://doi.org/10.3390/ani11020454
Chicago/Turabian StylePalomino, Jaime, Javiera Flores, Georges Ramirez, Victor H. Parraguez, and Monica De los Reyes. 2021. "Expression Profiles of the Progesterone Receptor, Cyclooxygenase-2, Growth Differentiation Factor 9, and Bone Morphogenetic Protein 15 Transcripts in the Canine Oviducts during the Oestrous Cycle" Animals 11, no. 2: 454. https://doi.org/10.3390/ani11020454
APA StylePalomino, J., Flores, J., Ramirez, G., Parraguez, V. H., & De los Reyes, M. (2021). Expression Profiles of the Progesterone Receptor, Cyclooxygenase-2, Growth Differentiation Factor 9, and Bone Morphogenetic Protein 15 Transcripts in the Canine Oviducts during the Oestrous Cycle. Animals, 11(2), 454. https://doi.org/10.3390/ani11020454