Do Domestic Pigs Acquire a Positive Perception of Humans through Observational Social Learning?

 ,
,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
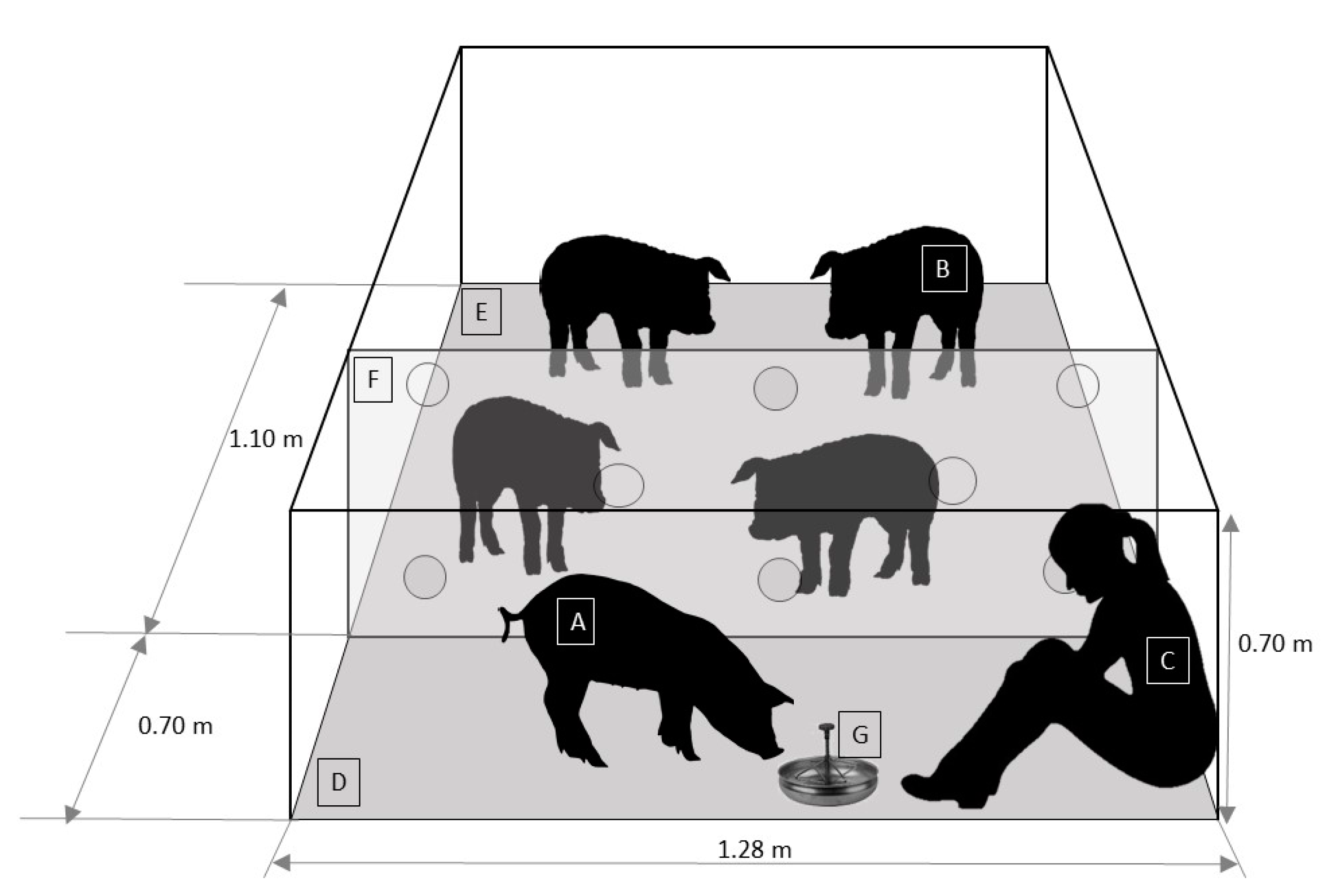
2.2. Experimental Set-Up
2.2.1. Determination of Social Dominance Order
2.2.2. Treatments
Gentle Handling Procedure
2.2.3. Open-field Arena Test
Behavioral Reactions toward the Stockperson in the Open-Field Arena Test
Behavioral Observations
Physiological Changes on Cardiac Activity in Response to Interaction with the Stockperson
Heart Rate Variability Analysis
2.3. Statistics
3. Results
3.1. Behavioral Reaction in Open-Field Arena Test
3.2. Physiological Reaction in the Open-Field Arena Test
4. Discussion
4.1. Behavioral Response of Observers during the Interaction with the Stockperson in the Open-Field Arena Test
4.2. Physiological Response to the Interaction with the Stockperson in the Open-Field Arena Test
4.3. Effect of Social Rank of Demonstrator Pig on Observer Pigs’ Response in the Open-Field Arena Test
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Statement
References
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 Five Domains Model: Including Human-Animal Interactions in Assessments of Animal Welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef] [PubMed]
- Waiblinger, S.; Boivin, X.; Pedersen, V.; Tosi, M.-V.; Janczak, A.M.; Visser, E.K.; Jones, R.B. Assessing the human-animal relationship in farmed species: A critical review. Appl. Anim. Behav. Sci. 2006, 101, 185–242. [Google Scholar] [CrossRef]
- Bertenshaw, C.; Rowlinson, P.; Edge, H.; Douglas, S.; Shiel, R. The effect of different degrees of ‘positive’ human-animal interaction during rearing on the welfare and subsequent production of commercial dairy heifers. Appl. Anim. Behav. Sci. 2008, 114, 65–75. [Google Scholar] [CrossRef]
- Coulon, M.; Nowak, R.; Peyrat, J.; Chandèze, H.; Boissy, A.; Boivin, X. Do lambs perceive regular human stroking as pleasant? Behavior and heart rate variability analyses. PLoS ONE 2015, 10, e0118617. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, Y.; Bi, Y.; Zhao, P.; Sun, H.; Li, J.; Liu, H.; Zhang, R.; Li, X.; Bao, J. Effects of Long-Term Gentle Handling on Behavioral Responses, Production Performance, and Meat Quality of Pigs. Animals 2020, 10, 330. [Google Scholar] [CrossRef]
- Gonyou, H.W.; Hemsworth, P.H.; Barnett, J.L. Effects of frequent interactions with humans on growing pigs. Appl. Anim. Behav. Sci. 1986, 16, 269–278. [Google Scholar] [CrossRef]
- Breuer, K.; Hemsworth, P.H.; Coleman, G.J. The effect of positive or negative handling on the behavioural and physiological responses of nonlactating heifers. Appl. Anim. Behav. Sci. 2003, 84, 3–22. [Google Scholar] [CrossRef]
- Brajon, S.; Laforest, J.P.; Bergeron, R.; Tallet, C.; Hötzel, M.J.; Devillers, N. Persistency of the piglet’s reactivity to the handler following a previous positive or negative experience. Appl. Anim. Behav. Sci. 2015, 162, 9–19. [Google Scholar] [CrossRef]
- Hemsworth, P.H.; Barnett, J.L.; Hansen, C.; Gonyou, H.W. The influence of early contact with humans on subsequent behavioural response of pigs to humans. Appl. Anim. Behav. Sci. 1986, 15, 55–63. [Google Scholar] [CrossRef]
- Sankey, C.; Richard-Yris, M.A.; Leroy, H.; Henry, S.; Hausberger, M. Positive interactions lead to lasting positive memories in horses, Equus caballus. Anim. Behav. 2010, 79, 869–875. [Google Scholar] [CrossRef]
- Tallet, C.; Sy, K.; Prunier, A.; Nowak, R.; Boissy, A.; Boivin, X. Behavioural and physiological reactions of piglets to gentle tactile interactions vary according to their previous experience with humans. Livest. Sci. 2014, 167, 331–341. [Google Scholar] [CrossRef]
- Tallet, C.; Brajon, S.; Devillers, N.; Lensink, J. Pig–human interactions: Creating a positive perception of humans to ensure pig welfare. In Advances in Pig Welfare, 1st ed.; Špinka, M., Ed.; Woodhead Publishing: Duxford, UK, 2018; pp. 381–398. [Google Scholar] [CrossRef]
- Brajon, S.; Laforest, J.P.; Schmitt, O.; Devillers, N. The way humans behave modulates the emotional state of piglets. PLoS ONE 2015, 10, e0133408. [Google Scholar] [CrossRef]
- Hemsworth, P.H.; Barnett, J.L. The effects of aversively handling pigs, either individually or in groups, on their behaviour, growth and corticosteroids. Appl. Anim. Behav. Sci. 1991, 30, 61–72. [Google Scholar] [CrossRef]
- Day, J.E.L.; Spoolder, H.A.M.; Burfoot, A.; Chamberlain, H.L.; Edwards, S.A. The separate and interactive effects of handling and environmental enrichment on the behaviour and welfare of growing pigs. Appl. Anim. Behav. Sci. 2002, 75, 177–192. [Google Scholar] [CrossRef]
- Nicol, C.J. The social transmission of information and behaviour. Appl. Anim. Behav. Sci. 1995, 44, 79–98. [Google Scholar] [CrossRef]
- Heyes, C.M. Social learning in animals: Categories and mechanisms. Biol. Rev. 1994, 69, 207–231. [Google Scholar] [CrossRef] [PubMed]
- Nicol, C.J. Development, direction, and damage limitation: Social learning in domestic fowl. Anim. Learn. Behav. 2004, 32, 72–81. [Google Scholar] [CrossRef]
- Munksgaard, L.; De Passillé, A.M.; Rushen, J.; Herskin, M.S.; Kristensen, A.M. Dairy cows’ fear of people: Social learning, milk yield and behaviour at milking. Appl. Anim. Behav. Sci. 2001, 73, 15–26. [Google Scholar] [CrossRef]
- Jones, R.B. Reduction of the domestic chick’s fear of human beings by regular handling and related treatments. Anim. Behav. 1993, 46, 991–998. [Google Scholar] [CrossRef]
- Zulkifli, I.; Azah, A.S.N. Fear and stress reactions, and the performance of commercial broiler chickens subjected to regular pleasant and unpleasant contacts with human being. Appl. Anim. Behav. Sci. 2004, 88, 77–87. [Google Scholar] [CrossRef]
- Henry, S.; Hemery, D.; Richard-Yris, M.A.; Hausberger, M. Human-mare relationships and behaviour of foals toward humans. Appl. Anim. Behav. Sci. 2005, 93, 341–362. [Google Scholar] [CrossRef]
- Henry, S.; Briefer, S.; Richard-Yris, M.A.; Hausberger, M. Are 6-month-old foals sensitive to dam’s influence? Dev. Psychobiol. 2007, 49, 514–521. [Google Scholar] [CrossRef]
- De Lima Rocha, A.D.; Menescal-de-Oliveira, L.; da Silva, L.F.S. Effects of human contact and intra-specific social learning on tonic immobility in guinea pigs, Cavia porcellus. Appl. Anim. Behav. Sci. 2017, 191, 1–4. [Google Scholar] [CrossRef]
- Held, S.; Mendl, M.; Devereux, C.; Byrne, R.W. Social tactics of pigs in a competitive foraging task: The ‘informed forager’ paradigm. Anim. Behav. 2000, 59, 569–576. [Google Scholar] [CrossRef]
- Morgan, C.A.; Lawrence, A.B.; Chirnside, J.; Deans, L.A. Can information about solid food be transmitted from one piglet to another? Anim. Sci. 2001, 73, 471–478. [Google Scholar] [CrossRef]
- Figueroa, J.; Solà-Oriol, D.; Manteca, X.; Pérez, J.F. Social learning of feeding behaviour in pigs: Effects of neophobia and familiarity with the demonstrator conspecific. Appl. Anim. Behav. Sci. 2013, 148, 120–127. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012; 400p. [Google Scholar]
- Langbein, J.; Puppe, B. Analysing dominance relationships by sociometric methods—A plea for a more standardised and precise approach in farm animals. Appl. Anim. Behav. Sci. 2004, 87, 293–315. [Google Scholar] [CrossRef]
- Stukenborg, A.; Traulsen, I.; Puppe, B.; Presuhn, U.; Krieter, J. Agonistic behaviour after mixing in pigs under commercial farm conditions. Appl. Anim. Behav. Sci. 2011, 129, 28–35. [Google Scholar] [CrossRef]
- Meese, G.B.; Ewbank, R. The establishment and nature of the dominance hierarchy in the domesticated pig. Anim. Behav. 1973, 21, 326–334. [Google Scholar] [CrossRef]
- Friend, T.H.; Knabe, D.A.; Tanskley, T.D., Jr. Behavior and performance of pigs grouped by three different methods at weaning. J. Anim. Sci. 1983, 57, 1406–1411. [Google Scholar] [CrossRef]
- McGlone, J.J. Influence of resources on pig aggression and dominance. Behav. Process. 1986, 12, 135–144. [Google Scholar] [CrossRef]
- Friard, O.; Gamba, M. BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Figueroa, J.; Frías, D.; Solà-Oriol, D.; Tadich, T.; Franco-Rosselló, R.; Nuñez, V.; Dwyer, D.M. Palatability in pigs, the pleasure of consumption. J. Anim. Sci. 2019, 97, 2165–2174. [Google Scholar] [CrossRef]
- De Oliveira, D.; Paranhos da Costa, M.J.R.; Zupan, M.; Rehn, T.; Keeling, L.J. Early human handling in non-weaned piglets: Effects on behaviour and body weight. Appl. Anim. Behav. Sci. 2015, 164, 56–63. [Google Scholar] [CrossRef]
- Marchant-Forde, R.M.; Marlin, D.J.; Marchant-Forde, J.N. Validation of a cardiac monitor for measuring heart rate variability in adult female pigs: Accuracy, artefacts and editing. Physiol. Behav. 2004, 80, 449–458. [Google Scholar] [CrossRef]
- Von Borell, E.; Langbein, J.; Després, G.; Hansen, S.; Leterrier, C.; Marchant-Forde, J.; Marchant-Forde, R.; Minero, M.; Mohr, E.; Prunier, A.; et al. Heart rate variability as a measure of autonomic regulation of cardiac activity for assessing stress and welfare in farm animals—A review. Physiol. Behav. 2007, 92, 293–316. [Google Scholar] [CrossRef]
- Malik, M.; Bigger, J.T.; Camm, A.J.; Kleiger, R.E.; Malliani, A.; Moss, A.J.; Schwartz, P.J. Heart rate variability: Standards of measurement, physiological interpretation, and clinical use. Eur. Heart J. 1996, 17, 354–381. [Google Scholar] [CrossRef]
- Byrd, C.J.; Johnson, J.S.; Radcliffe, J.S.; Craig, B.A.; Eicher, S.D.; Lay, D.C. Nonlinear analysis of heart rate variability for evaluating the growing pig stress response to an acute heat episode. Animal 2020, 14, 379–387. [Google Scholar] [CrossRef]
- Poletto, R.; Janczak, A.M.; Robichat-Forde, R.M.; Marchant-Forde, J.N.; Matthews, D.L.; Dowell, C.A.; Hogan, D.F.; Freeman, L.J.; Lay, D.C., Jr. Identification of low and high frequency ranges for heart rate variability and blood pressure analyses using pharmacological autonomic blockade with atropine and propranolol in swine. Physiol. Behav. 2011, 103, 188–196. [Google Scholar] [CrossRef]
- Howell, D.C. Multiple comparisons among treatment means. In Statistical Methods for Psychology, 8th ed.; Wadsworth Cengage Learning: Belmont, CA, USA, 2012; pp. 369–410. [Google Scholar]
- Martin, P.; Bateson, P. Statistical analysis. In Measuring Behaviour: An Introductory Guide, 3rd ed.; Cambridge University Press: Cambridge, UK, 1994; pp. 103–120. [Google Scholar] [CrossRef]
- Jahn-Eimermacher, A.; Lasarzik, I.; Raber, J. Statistical analysis of latency outcomes in behavioral experiments. Behav. Brain Res. 2011, 221, 271–275. [Google Scholar] [CrossRef]
- Beasley, T.M.; Schumacker, R.E. Multiple regression approach to analyzing contingency tables: Post hoc and planned comparison procedures. J. Exp. Educ. 1995, 64, 79–93. [Google Scholar] [CrossRef]
- Leliveld, L.M.; Düpjan, S.; Tuchscherer, A.; Puppe, B. Behavioural and physiological measures indicate subtle variations in the emotional valence of young pigs. Physiol. Behav. 2016, 157, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Miura, A.; Tanida, H.; Tanaka, T.; Yoshimoto, T. The influence of human posture and movement on the approach and escape behaviour of weanling pigs. Appl. Anim. Behav. Sci. 1996, 49, 247–256. [Google Scholar] [CrossRef]
- Tallet, C.; Veissier, I.; Boivin, X. Human contact and feeding as rewards for the lamb’s affinity to their stockperson. Appl. Anim. Behav. Sci. 2005, 94, 59–73. [Google Scholar] [CrossRef]
- Lürzel, S.; Windschnurer, I.; Futschik, A.; Waiblinger, S. Gentle interactions decrease the fear of humans in dairy heifers independently of early experience of stroking. Appl. Anim. Behav. Sci. 2016, 178, 16–22. [Google Scholar] [CrossRef]
- Rault, J.L.; Waiblinger, S.; Boivin, X.; Hemsworth, P. The power of a positive human-animal relationship for animal welfare. Front. Vet. Sci. 2020, 7, 590867. [Google Scholar] [CrossRef]
- Zupan, M.; Framstad, T.; Zanella, A.J. Behaviour, heart rate, and heart rate variability in pigs exposed to novelty. Rev. Bras. Zootec. 2016, 45, 121–129. [Google Scholar] [CrossRef]
- Bensoussan, S.; Tigeot, R.; Meunier-Salaün, M.C.; Tallet, C. Broadcasting human voice to piglets (Sus scrofa domestica) modifies their behavioural reaction to human presence in the home pen and in arena tests. Appl. Anim. Behav. Sci. 2020, 225, 104965. [Google Scholar] [CrossRef]
- Weary, D.M.; Ross, S.; Fraser, D. Vocalizations by isolated piglets: A reliable indicator of piglet need directed towards the sow. Appl. Anim. Behav. Sci. 1997, 53, 249–257. [Google Scholar] [CrossRef]
- Düpjan, S.; Schön, P.C.; Puppe, B.; Tuchscherer, A.; Manteuffel, G. Differential vocal responses to physical and mental stressors in domestic pigs (Sus scrofa). Appl. Anim. Behav. Sci. 2008, 114, 105–115. [Google Scholar] [CrossRef]
- Pairis, M.; Young, A.; Millman, S.T.; Garvey, J.; Johnson, A.K. Can fear be effectively assessed in swine? A study measuring fear levels during a human approach test. Anim. Ind. Rep. 2009, 655. [Google Scholar] [CrossRef]
- Forkman, B.; Boissy, A.; Meunier-Salaün, M.C.; Canali, E.; Jones, R.B. A critical review of fear tests used on cattle, pigs, sheep, poultry and horses. Physiol. Behav. 2007, 92, 340–374. [Google Scholar] [CrossRef] [PubMed]
- Reimert, I.; Rodenburg, T.B.; Ursinus, W.W.; Kemp, B.; Bolhuis, J.E. Responses to novel situations of female and castrated male pigs with divergent social breeding values and different backtest classifications in barren and straw-enriched housing. Appl. Anim. Behav. Sci. 2014, 151, 24–35. [Google Scholar] [CrossRef]
- Reimert, I.; Rodenburg, T.B.; Ursinus, W.W.; Duijvesteijn, N.; Camerlink, I.; Kemp, B.; Bolhuis, J.E. Backtest and novelty behavior of female and castrated male piglets with diverging social breeding values for growth. J. Anim. Sci. 2013, 91, 4589–4597. [Google Scholar] [CrossRef]
- Mineka, S.; Davidson, M.; Cook, M.; Keir, R. Observational conditioning of snake fear in rhesus monkeys. J. Abnorm. Psychol. 1984, 93, 355–372. [Google Scholar] [CrossRef]
- Cook, M.; Mineka, S.; Wolkenstein, B.; Laitsch, K. Observational conditioning of snake fear in unrelated rhesus monkeys. J. Abnorm. Psychol. 1985, 94, 591–610. [Google Scholar] [CrossRef]
- Imfeld-Mueller, S.; Van Wezemael, L.; Stauffacher, M.; Gygax, L.; Hillmann, E. Do pigs distinguish between situations of different emotional valences during anticipation? Appl. Anim. Behav. Sci. 2011, 131, 86–93. [Google Scholar] [CrossRef]
- Zupan, M.; Buskas, J.; Altimiras, J.; Keeling, L.J. Assessing positive emotional states in dogs using heart rate and heart rate variability. Physiol. Behav. 2015, 155, 102–111. [Google Scholar] [CrossRef]
- Baciadonna, L.; Nawroth, C.; McElligott, A.G. Judgement bias in goats (Capra hircus): Investigating the effects of human grooming. Peer J. 2016, 4, e2485. [Google Scholar] [CrossRef]
- Kokras, N.; Dalla, C. Sex differences in animal models of psychiatric disorders. Br. J. Pharmacol. 2014, 171, 4595–4619. [Google Scholar] [CrossRef]
- Lay, D.C., Jr.; Matteri, R.L.; Carroll, J.A.; Fangman, T.J.; Safranski, T.J. Preweaning survival in swine. J. Anim. Sci. 2002, 80, E74–E86. [Google Scholar] [CrossRef]
- Zebunke, M.; Langbein, J.; Manteuffel, G.; Puppe, B. Autonomic reactions indicating positive affect during acoustic reward learning in domestic pigs. Anim. Behav. 2011, 81, 481–489. [Google Scholar] [CrossRef]
- Coussi-Korbel, S.; Fragaszy, D.M. On the relation between social dynamics and social learning. Anim. Behav. 1995, 50, 1441–1453. [Google Scholar] [CrossRef]
- Kendal, R.; Hopper, L.M.; Whiten, A.; Brosnan, S.F.; Lambeth, S.P.; Schapiro, S.J.; Hoppitt, W. Chimpanzees copy dominant and knowledgeable individuals: Implications for cultural diversity. Evol. Hum. Behav. 2015, 36, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Canteloup, C.; Hoppitt, W.; van de Waal, E. Wild primates copy higher-ranked individuals in a social transmission experiment. Nat. Commun. 2020, 11, 459. [Google Scholar] [CrossRef] [PubMed]
- Nicol, C.J.; Pope, S.J. Social learning in small flocks of laying hens. Anim. Behav. 1994, 47, 1289–1296. [Google Scholar] [CrossRef]
- Boogert, N.J.; Reader, S.M.; Laland, K.N. The relation between social rank, neophobia and individual learning in starlings. Anim. Behav. 2006, 72, 1229–1239. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Day of the Experiment | Pigs Age (Days) | Event/Test | Place | Measurements |
|---|---|---|---|---|
| 1 | 21 | Weaning | Commercial farm | - |
| 1–7 | 21–27 | Acclimation to nursery pens | Home pen | - |
| 1–2 | 21–22 | Agonistic behaviors register | Home pen | Behavioral |
| 3–5 | 23–25 | Agonistic behaviors analysis | - | Behavioral |
| 6 | 26 | Dominance Index estimation | - | Dominance index |
| 8–40 | 28–60 | Gentle handling sessions | Home pen | - |
| 27–28 | 47–48 | Habituation to band of Polar device | Home pen | - |
| 29–31 | 49–51 | Habituation to arena and transport cage | Open-field arena | - |
| 34–35 | 54–55 | Basal HRV measurement | Home pen | Baseline HRV |
| 44–46 | 64–66 | Open-field arena test | Open-field arena | Behavioral and HRV |
| Behavioral Observation | Description | Phase |
|---|---|---|
| Latency to first physical contact | Time (s) taken for the pig to make physical contact with any part of body of the stockperson | 2, 3 |
| Latency to approach the area around the stockperson | Time (s) taken for the pig to enter an area within 0.5 m diameter around the point at which the stockperson stood | 2, 3 |
| Time of contact | Time (%) that pig stayed in physical contact with any part of body of the stockperson, either touching or sniffing it | 2, 3 |
| Time in area around the stockperson | Time (%) that pig spent within 0.5 m diameter around of the stockperson, either in contact with her (touching or sniffing) or while exploring the environment | 2, 3 |
| Looking at the stockperson | Time (%) that pig stood still looking at the stockperson, with its body and head oriented toward the stockperson in attention attitude | 2, 3 |
| Frequency of contact | Number of times the pig physically contacted the stockperson | 2, 3 |
| Climbing on the stockperson | Frequency of climbing on the stockperson, with at least the front legs on the thighs, and number of animals that were observed climbing on the stockperson | 3 |
| Accepted strokes | Strokes (%) accepted by each pig based on the total attempts made by the stockperson | 4 |
| Attempts required until accepting the first stroke | Number of attempts made by the stockperson until the pig accepted the first stroke | 4 |
| High-pitched vocalizations | Number of screams, squeals and grunt-squeals | 1, 2, 3, 4 |
| Low-pitched vocalizations | Number of short or long grunts | 1, 2, 3, 4 |
| Locomotor activity | Number of zones that the pig crossed with its front legs | 1, 2, 3, 4 |
| Defecation | Number of animals that were observed defecating in any phase where the stockperson was present | 2, 3, 4 |
| Urination | Number of animals that were observed urinating in any phase where the stockperson was present | 2, 3, 4 |
| Behavior | Treatments | p-Value | ||
|---|---|---|---|---|
| CG | DDG | SDG | ||
| (n = 24) | (n = 20) | (n = 20) | ||
| Phase 1 | ||||
| Locomotion | 35.83 ± 3.20 | 39.45 ± 3.39 | 34.40 ± 3.39 | 0.571 1 |
| Low-pitched vocalization | 20.66 ± 3.78 | 29.40 ± 4.14 | 19.45 ± 4.14 | 0.183 1 |
| High-pitched vocalization | 0 (0–0) | 0 (0–0) | 0 (0–0) | 0.562 3 |
| Phase 2 | ||||
| Locomotion | 11.34 ± 1.80 | 9.00 ± 1.88 | 8.80 ± 1.88 | 0.567 1 |
| Low-pitched vocalization | 16.92 ± 2.74 | 14.05 ± 2.94 | 12.60 ± 2.94 | 0.561 1 |
| High-pitched vocalization | 0 (0–0.75) | 0 (0–3.5) | 0 (0–0) | 0.152 3 |
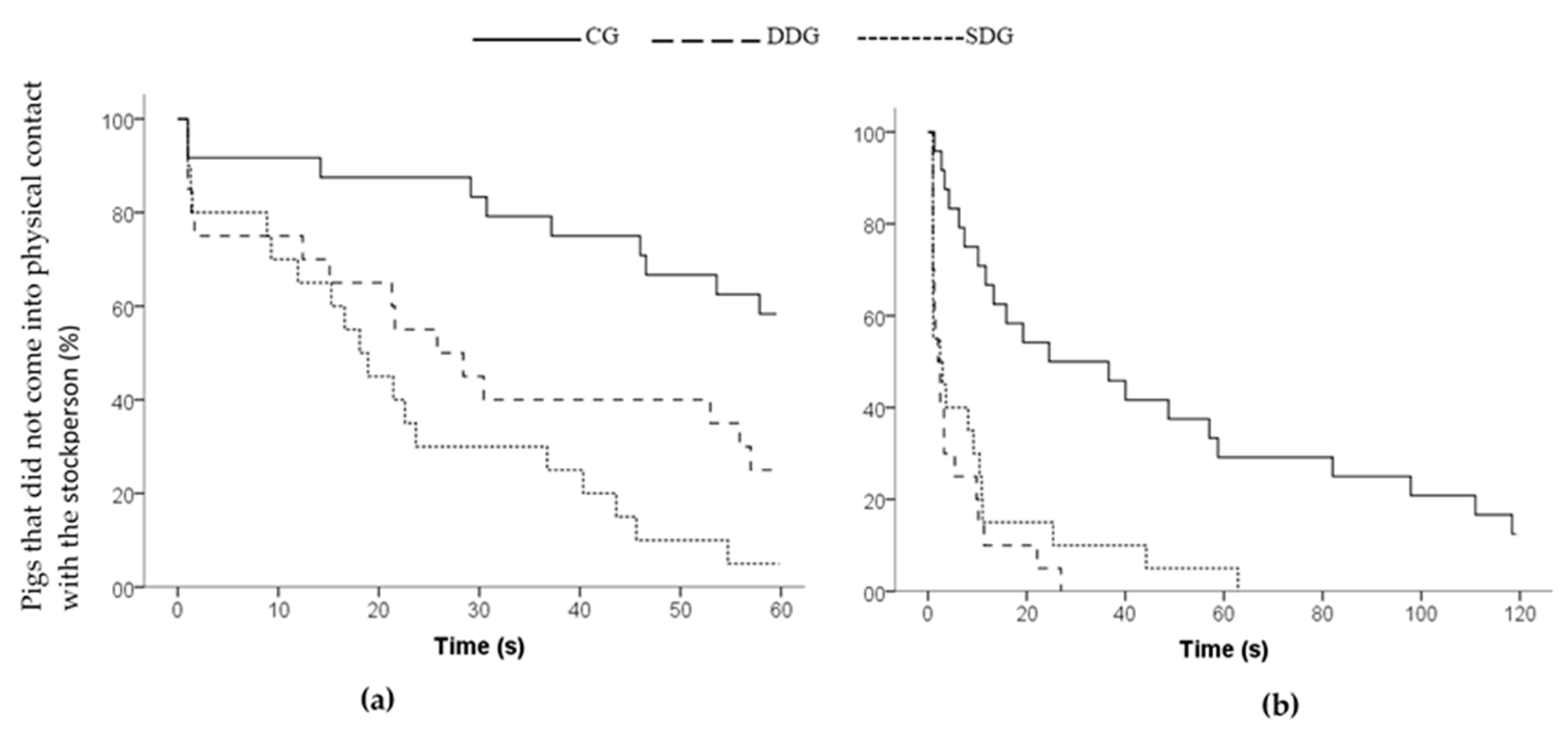
| Latency to approach area (s) | 42.66 ± 4.24 a | 19.18 ± 4.81 b | 18.89 ± 3.66 b | 0.001 2 |
| Latency to first contact (s) | 48.22 ± 3.83 a | 31.34 ± 5.30 b | 22.62 ± 3.98 b | 0.001 2 |
| Time looking at the stockperson (%) | 47.43 ± 6.47 a | 23.30 ± 6.80 b | 22.29 ± 6.80 b | 0.027 1a |
| Time in area (%) | 20.14 ± 6.49 a | 45.77 ± 7.11 b | 45.39 ± 7.11 b | 0.002 1a |
| Time of contact (%) | 8.28 ± 4.79 a | 27.12 ± 5.25 b | 25.60 ± 5.25 b | 0.003 1a |
| Physical contact (frequency) | 0.74 ± 0.35 a | 1.85 ± 0.36 b | 2.15 ± 0.36 b | 0.039 1 |
| Phase 3 | ||||
| Locomotion | 21.98 ± 2.33 p | 13.95 ± 2.40 q | 14.50 ± 2.40 q | 0.058 1 |
| Low-pitched vocalization | 29.71 ± 5.19 p | 14 ± 5.45 q | 15 ± 5.45 q | 0.100 1 |
| High-pitched vocalization | 0 (0–2.75) p | 0 (0–0) q | 0 (0–0) q | 0.080 3 |
| Latency to approach area (s) | 42.21 ± 8.49 a | 4.09 ± 1.47 b | 9.07 ± 3.53 b | <0.001 2 |
| Latency to first contact (s) | 47.11 ± 8.97 a | 5.47 ± 1.64 b | 10.02 ± 3.65 b | <0.001 2 |
| Time looking at the stockperson (%) | 13.30 (3.18–28.05) a | 1.60 (0–8.35) b | 1.04 (0–5.87) b | 0.001 3 |
| Time in area (%) | 45.93 ± 7.42 a | 84.19 ± 7.82 b | 77.11 ± 7.82 b | 0.008 1a |
| Time of contact (%) | 37.64 ± 8.35 a | 76.35 ± 8.68 b | 65.30 ± 8.68 b | 0.014 1a |
| Physical contact (frequency) | 3.96 ± 0.78 | 3.75 ± 0.80 | 3.99 ± 0.80 | 0.975 1 |
| Climb on the stockperson (frequency) | 1.10 ± 0.54 | 1.65 ± 0.56 | 0.84 ± 0.56 | 0.592 1 |
| Phase 4 | ||||
| Locomotion | 37.90 ± 2.53 a | 28.62 ± 2.69 b | 23.22 ± 2.69 b | 0.005 1 |
| Low-pitched vocalization | 33.50 ± 3.49 a | 17.10 ± 3.77 b | 12.70 ± 3.77 b | 0.003 1 |
| High-pitched vocalization | 2 (0–12.50) a | 0 (0–0) b | 0 (0–0) b | 0.007 3 |
| Accepted strokes (%) | 25.36 ± 5.92 a | 80.51 ± 6.24 b | 84.77 ± 6.35 b | <0.001 1a |
| Attempts until first accepted stroke | 5.50 (4–8.50) a | 1 (1–2) b | 1 (1–2) b | <0.001 3 |
| Parameter (Unit) | Treatments | Sex | |||||
|---|---|---|---|---|---|---|---|
| CG | DDG | SDG | p-Value | Female | Male | p-Value | |
| (n = 21) | (n = 20) | (n = 19) | (n = 30) | (n = 30) | |||
| RR Interval (ms) | −30.02 ± 8.58 | −13.13 ± 8.17 | −24.24 ± 8.32 | 0.358 1 | −13.44 ± 6.43 p | −31.49 ± 6.42 q | 0.057 1 |
| HR (bpm) | 14.05 ± 4.28 | 6.57 ± 4.08 | 11.65 ± 4.15 | 0.465 1 | 7.04 ± 3.21 p | 14.47 ± 3.20 q | 0.100 1 |
| SDNN (ms) | 10.49 ± 1.81 | 5.72 ± 1.87 | 6.46 ± 1.84 | 0.190 1 | 5.31 ± 1.41 a | 9.81 ± 1.42 b | 0.020 1 |
| RMSSD (ms) | 2.71 ± 0.83 | 1.33 ± 0.87 | 1.68 ± 0.85 | 0.506 1 | 1.51 ± 0.66 | 2.31 ± 0.66 | 0.379 1 |
| RMSSD/SDNN (ms) | −0.11 ± 0.04 | −0.08 ± 0.04 | −0.10 ± 0.04 | 0.874 1 | −0.05 ± 0.03 a | −0.14 ± 0.03 b | 0.034 1 |
| LF (ms) | 505.27 ± 67.49 a | 118.76 ± 69.57 b | 109.48 ± 68.30 b | 0.003 1 | 127.20 ± 52.09 a | 361.80 ± 52.28 b | 0.001 1 |
| HF (ms) | 96.76 ± 25.36 | 89.79 ± 26.66 | 89.53 ± 26.02 | 0.975 1 | 72.14 ± 21.24 | 111.92 ± 21.24 | 0.191 1 |
| LF/HF (ms) | 2.023 ± 0.83 a | −2.14 ± 0.85 b | −0.91 ± 0.84 b | 0.015 1 | −1.56 ± 0.63 a | 0.87 ± 0.63 b | 0.004 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luna, D.; González, C.; Byrd, C.J.; Palomo, R.; Huenul, E.; Figueroa, J. Do Domestic Pigs Acquire a Positive Perception of Humans through Observational Social Learning? Animals 2021, 11, 127. https://doi.org/10.3390/ani11010127
Luna D, González C, Byrd CJ, Palomo R, Huenul E, Figueroa J. Do Domestic Pigs Acquire a Positive Perception of Humans through Observational Social Learning? Animals. 2021; 11(1):127. https://doi.org/10.3390/ani11010127
Chicago/Turabian StyleLuna, Daniela, Catalina González, Christopher J. Byrd, Rocío Palomo, Elizabeth Huenul, and Jaime Figueroa. 2021. "Do Domestic Pigs Acquire a Positive Perception of Humans through Observational Social Learning?" Animals 11, no. 1: 127. https://doi.org/10.3390/ani11010127
APA StyleLuna, D., González, C., Byrd, C. J., Palomo, R., Huenul, E., & Figueroa, J. (2021). Do Domestic Pigs Acquire a Positive Perception of Humans through Observational Social Learning? Animals, 11(1), 127. https://doi.org/10.3390/ani11010127