
Pathogenicity and Volatile Nematicidal Metabolites from Duddingtonia flagrans against Meloidogyne incognita


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Normal Culture
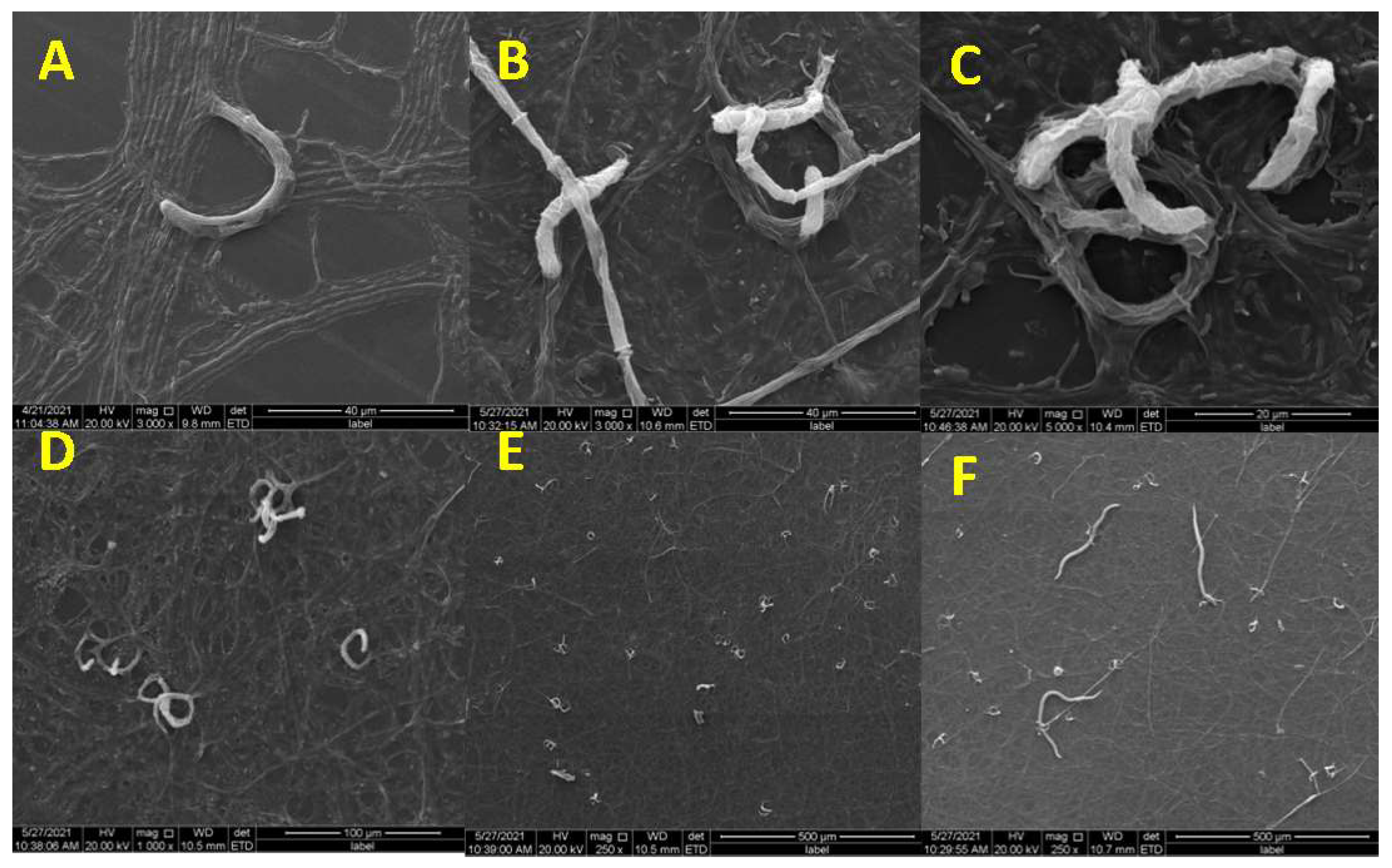
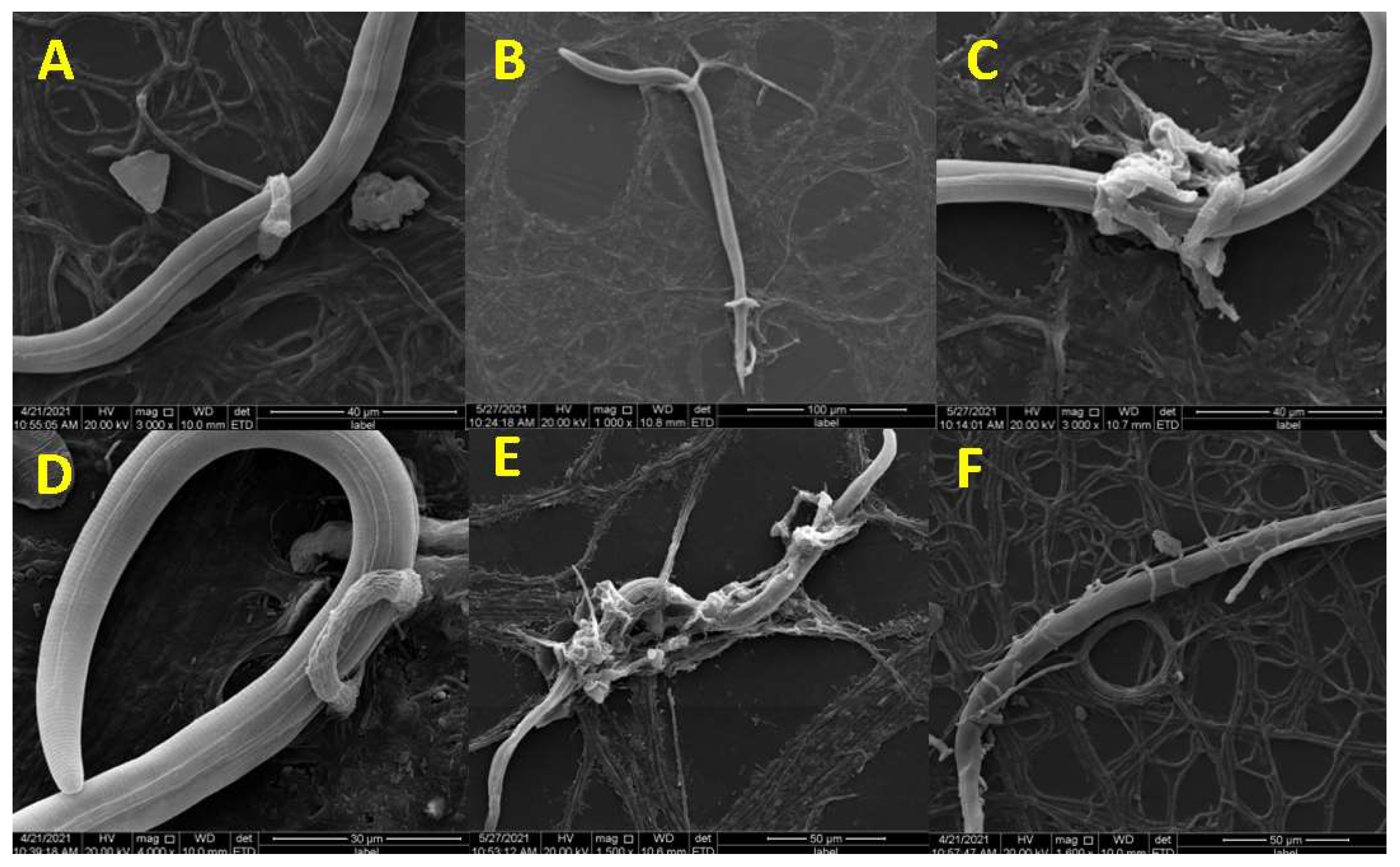
2.2. Observation of Trap Formation and Predatory Process Using Scanning Electron Microscopy
2.3. Gas Chromatography–Mass Spectrometry Assay
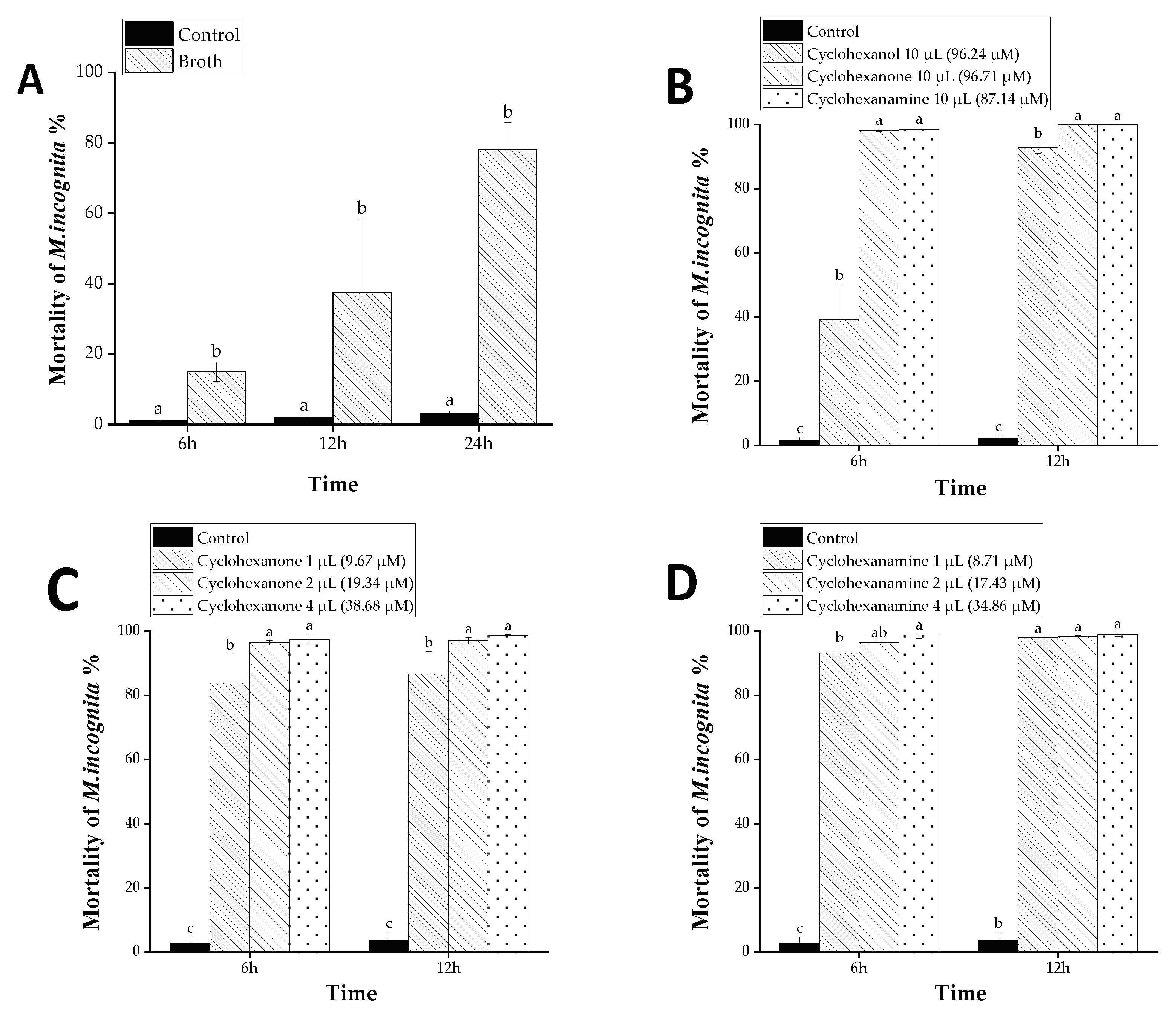
2.4. Assays of Nematicidal Activity
2.4.1. Nematicidal Activity of the Broth’s VOCs and Compounds
2.4.2. Inhibition of Egg Hatching Activity by VOCs
3. Results
3.1. Trap Formation and Pathogenicity Process of D. flagrans against M. incognita
3.2. Nematicidal Activity of D. flagrans’ Broth VOCs against M. incognita
3.3. Identification of the VOCs Produced by D. flagrans
3.4. Nematicidal Activity of VOCs against M. incognita
3.5. Inhibition of Egg Hatching by VOCs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elling, A.A. Major emerging problems with minor Meloidogyne species. Phytopathology 2013, 103, 1092–1102. [Google Scholar] [CrossRef]
- Oka, Y.; Koltai, H.; Bar-Eyal, M.; Mor, M.; Sharon, E.; Chet, I.; Spiegel, Y. New strategies for the control of plant-parasitic nematodes. Pest Manag. Sci. 2000, 56, 983–988. [Google Scholar] [CrossRef]
- Youssar, L.; Wernet, V.; Hensel, N.; Yu, X.; Hildebrand, H.G.; Schreckenberger, B.; Kriegler, M.; Hetzer, B.; Frankino, P.; Dillin, A.; et al. Intercellular communication is required for trap formation in the nematode-trapping fungus Duddingtonia flagrans. PLoS Genet 2019, 15, e1008029. [Google Scholar] [CrossRef] [PubMed]
- Pandit, R.; Patelm, R.; Patel, N.; Bhatt, V.; Joshi, C.; Singh, P.K.; Kunjadia, A. RNA-Seq reveals the molecular mechanism of trapping and killing of root-knot nematodes by nematode-trapping fungi. World J. Microbiol. Biotechnol. 2017, 33, 65. [Google Scholar] [CrossRef]
- Nordbring-Hertz, B.; Jansson, H.B.; Tunlid, A. Nematophagous Fungi. Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2006. [Google Scholar]
- Lopez-Llorca, L.V.; Olivares-Bernabeu, C.; Salinas, J.; Jansson, H.B.; Kolattukudy, P.E. Prepenetration events in fungal parasitism of nematode eggs. Mycol. Res. 2002, 106, 499–506. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, E.; An, Z.; Liu, X. Evolution of nematode-trapping cells of predatory fungi of the Orbiliaceae based on evidence from rRNA-encoding DNA and multiprotein sequences. Proc. Natl. Acad. Sci. USA 2007, 104, 8379–8384. [Google Scholar] [CrossRef]
- Hertzberg, H.; Larsen, M.; Maurer, V. Biological control of helminths in grazing animals using nematophagous fungi. Berl. Munch. Tierarztl. Wochenschr. 2001, 115, 278–285. [Google Scholar]
- Sahoo, A.; Khan, F. Nutritional and biological control synergism against gastrointestinal nematodes in small ruminants. J. Vet Sci. Anim. Husb. 2016, 4, 104. [Google Scholar]
- Cai, K.Z.; Liu, J.L.; Liu, W.; Wang, B.B.; Xu, Q.; Sun, L.J.; Chen, M.Y.; Zhao, M.W.; Wu, J.Y.; Li, X.S. Screening of different sample types associated with sheep and cattle for the presence of nematophagous fungi in China. J. Basic Microbiol. 2016, 56, 214–228. [Google Scholar] [CrossRef]
- Céspedes-Gutiérrez, E.; Aragón-Novoa, D.M.; Gómez-Lvarez, M.I.; Cortés-Rojas, D.F. In vitro evaluation of physicochemical variables on the nematophagous fungus Duddingtonia flagrans. J. Basic Microbiol. 2021, 61, 547–556. [Google Scholar] [CrossRef]
- Pandit, R.J.; Kunjadia, P.D.; Mukhopadhyaya, P.N.; Joshi, C.G. Isolation, molecular characterization and predatory activity of two Indian isolates of nematode-trapping fungi. Appl. Biol. Res. 2014, 16, 1–11. [Google Scholar] [CrossRef]
- Monteiro, T.S.A.; Balbino, H.M.; de Mello, I.N.K.; Coutinhoa, R.R.; de Araújo, J.V.; Freitasa, L.G. Duddingtonia flagrans preying a plant parasitic nematode. Braz. J. Biol. 2020, 80, 197–198. [Google Scholar] [CrossRef]
- Monteiro, T.S.A.; Valadares, S.V.; de Mello, I.N.K.; Moreira, B.C.; Kasuya, M.C.M.; de Araújo, J.V.; de Freitas, L.G. Nematophagus fungi increasing phosphorus uptake and promoting plant growth. Biol. Control 2018, 123, 71–75. [Google Scholar] [CrossRef]
- Campos, V.P.; Pinho, R.S.C.; Freire, E.S. Volatiles produced by interacting microorganisms potentially useful for the control of plant pathogens. Ciênc E Agrotecnol. 2010, 34, 525–535. [Google Scholar] [CrossRef]
- Terra, W.C.; Campos, V.P.; Martins, S.J.; Costa, L.S.A.S.; da Silva, J.C.P.; Barros, A.F.; Lopez, L.E.; Santos, T.C.N.; Smant, G.; Oliveira, D.F. Volatile organic molecules from Fusarium oxysporum strain 21 with nematicidal activity against Meloidogyne incognita. Crop Prot. 2018, 106, 125–131. [Google Scholar] [CrossRef]
- Liarzi, O.; Bucki, P.; Miyara, S.B.; Ezra, D. Bioactive volatiles from an endophytic Daldinia cf. concentrica isolate affect the viability of the plant parasitic nematode Meloidogyne javanica. PLoS ONE 2016, 11, e0168437. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Dou, G.; Gao, M.; Ren, F.; Li, R.; Zhang, X.; Yan, D.H. Annulohypoxylon sp. FPYF3050 produces volatile organic compounds against the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2020, 22, 245–255. [Google Scholar] [CrossRef]
- Degenkolb, T.; Vilcinskas, A. Metabolites from nematophagous fungi and nematicidal natural products from fungi as an alternative for biological control. Part I: metabolites from nematophagous ascomycetes. Appl. Microbiol. Biotechnol. 2016, 100, 3799–3812. [Google Scholar] [CrossRef]
- Anderson, M.G.; Rickards, R.W.; Lacey, E. Structures of flagranones A, B and C, cyclohexenoxide antibiotics from the nematode-trapping fungus Duddingtonia flagrans. J. Antibiot. 1999, 53, 1023–1028. [Google Scholar] [CrossRef][Green Version]
- Lee, Y.S.; Naning, K.W.; Nguyen, X.H.; Kim, S.B.; Moon, J.H.; Kim, K.Y. Ovicidal activity of lactic acid produced by Lysobacter capsici YS1215 on eggs of root-knot nematode, Meloidogyne incognita. J. Microbiol. Biotechnol. 2014, 24, 1510–1515. [Google Scholar] [CrossRef]
- Wan, J.; Dai, Z.; Zhang, K.; Li, G.; Zhao, P. Pathogenicity and metabolites of endoparasitic nematophagous fungus Drechmeria coniospora YMF1.01759 against nematodes. Microorganisms 2021, 9, 1735. [Google Scholar] [CrossRef] [PubMed]
- Su, H.N.; Xu, Y.Y.; Wang, X.; Zhang, K.Q.; Li, G.H. Induction of trap formation in nematode-trapping fungi by bacteria-released ammonia. Lett. Appl. Microbiol. 2016, 62, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Yu, C.; Shao, Z.; Cai, M.; Li, G.; Zheng, L.; Yu, Z.; Zhang, J. Identification and characterization of nematicidal volatile organic compounds from deep sea Virgibacillus dokdonensis MCCC 1A00493. Molecules 2020, 25, 744. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Ferrari, F.; Giannakou, I.; Menkissoglu-Spiroudi, U. Synergistic and antagonistic interactions of terpenes against Meloidogyne incognita and the nematicidal activity of essential oils from seven plants indigenous to Greece. Pest Manag. Sci. 2011, 67, 341–351. [Google Scholar] [CrossRef]
- Zhai, Y.; Shao, Z.; Cai, M.; Zheng, L.; Li, G.; Huang, D.; Cheng, W.; Thomashow, L.S.; Weller, D.M.; Yu, Z.; et al. Multiple modes of nematode control by volatiles of Pseudomonas putida 1A00316 from Antarctic soil against Meloidogyne incognita. Front. Microbiol. 2018, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- Maciel, A.S.; Araújo, J.V.; Campos, A.K.; Benjamin, L.A.; Freitas, L.G. Scanning electron microscopy of Ancylostoma spp. dog infective larvae captured and destroyed by the nematophagous fungus Duddingtonia flagrans. Micron 2009, 40, 463–470. [Google Scholar] [CrossRef]
- Andrés, M.F.; González-Coloma, A.; Sanz, J.; Burillo, J.; Sainz, P. Nematicidal activity of essential oils: a review. Phytochem. Rev. 2012, 11, 371–390. [Google Scholar] [CrossRef]
- Campos, A.K.; Araújo, J.V.; Guimares, M.P. Interaction between the nematophagous fungus Duddingtonia flagrans and infective larvae of Haemonchus contortus (Nematoda: Trichostrongyloidea). J. Helminthol. 2008, 82, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Luo, H.L.; Wangm, R.; Yang, L.R.; Zhao, Z.G.; Zhang, W.; Li, B.; Yang, X.Y.; Luo, X.P. Observation on predatory process of Duddingtonia flagrans against infective larvae of Strongylus equinus. Progress Vet. Med. 2018, 39(10), 122–125. [Google Scholar]
- Xu, C.L.; Liu, W.; Li, Y.Q.; Wang, K.Y.; Qin, G.G.; Wang, D.M.; Wang, B.B.; Wang, H.; Li, X.; Yi, L.X. Observation dynamics of Duddingtonia flagrans against infective larvae of Haemonchus contortus and free-living nematode Caenorhabditis elegans. Chin. Vet. Sci. 2014, 44, 1119–1126. [Google Scholar]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 1–23. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Hatched Worms per Egg Mass ± SD | |||
|---|---|---|---|
| 1 Day | 2 Days | 3 Days | |
| Cyclohexanol 96.24 µM | 29.13 ± 5.95 b | 30.41 ± 4.87 b | 30.69 ± 4.59 b |
| Cyclohexanone 96.71 µM | 2.25 ± 0.82 d | 3.14 ± 0.33 d | 3.14 ± 0.33 d |
| Cyclohexanone 48.36 µM | 10.01 ± 1.21 cd | 10.14 ± 1.23 cd | 10.14 ± 1.23 c |
| Cyclohexanamine 26.14 µM | 7.00 ± 1.21 cd | 8.18 ± 0.64 cd | 8.44 ± 0.69 cd |
| Culture broth 200 µL | 14.13 ± 4.02 c | 14.92 ± 4.01 c | 15.35 ± 4.24 c |
| control | 83.28 ± 2.76 a | 93.50 ± 3.05 a | 99.72 ± 2.71 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, X.; Wang, X.; Li, G. Pathogenicity and Volatile Nematicidal Metabolites from Duddingtonia flagrans against Meloidogyne incognita. Microorganisms 2021, 9, 2268. https://doi.org/10.3390/microorganisms9112268
Mei X, Wang X, Li G. Pathogenicity and Volatile Nematicidal Metabolites from Duddingtonia flagrans against Meloidogyne incognita. Microorganisms. 2021; 9(11):2268. https://doi.org/10.3390/microorganisms9112268
Chicago/Turabian StyleMei, Xiaoyu, Xin Wang, and Guohong Li. 2021. "Pathogenicity and Volatile Nematicidal Metabolites from Duddingtonia flagrans against Meloidogyne incognita" Microorganisms 9, no. 11: 2268. https://doi.org/10.3390/microorganisms9112268
APA StyleMei, X., Wang, X., & Li, G. (2021). Pathogenicity and Volatile Nematicidal Metabolites from Duddingtonia flagrans against Meloidogyne incognita. Microorganisms, 9(11), 2268. https://doi.org/10.3390/microorganisms9112268