
Electrochemical Characterization of Two Gut Microbial Strains Cooperatively Promoting Multiple Sclerosis Pathogenesis




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture Preparation
2.2. Transmission Electron Microscopy
2.3. Whole-Cell Electrochemical Analysis of Pure Strains
2.4. Supernatant Exchange Experiments during the Current Producing Condition
2.5. Scanning Electron Microscopy (SEM)
3. Results and Discussion
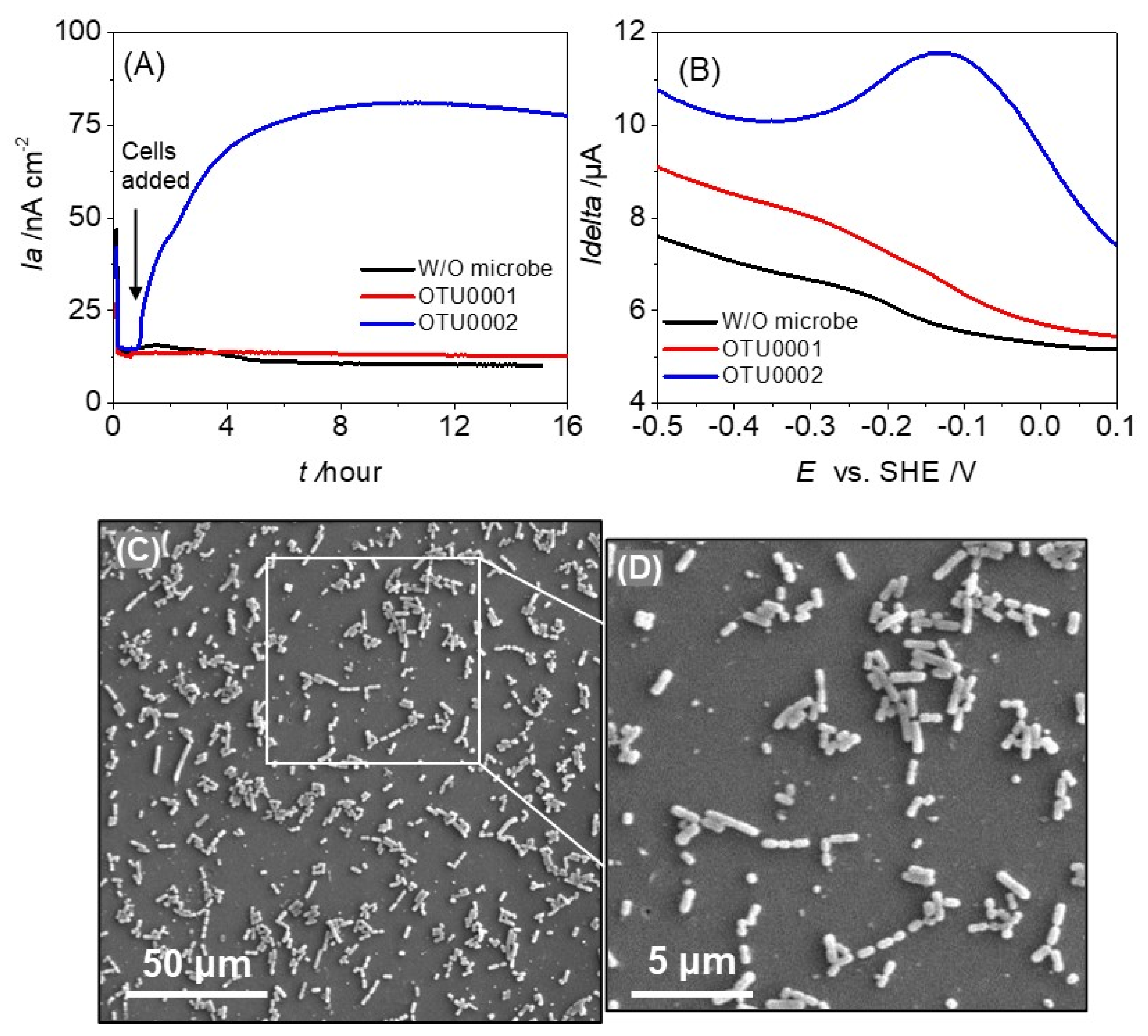
3.1. Electrochemical Characterization of OTU0001 and OTU0002
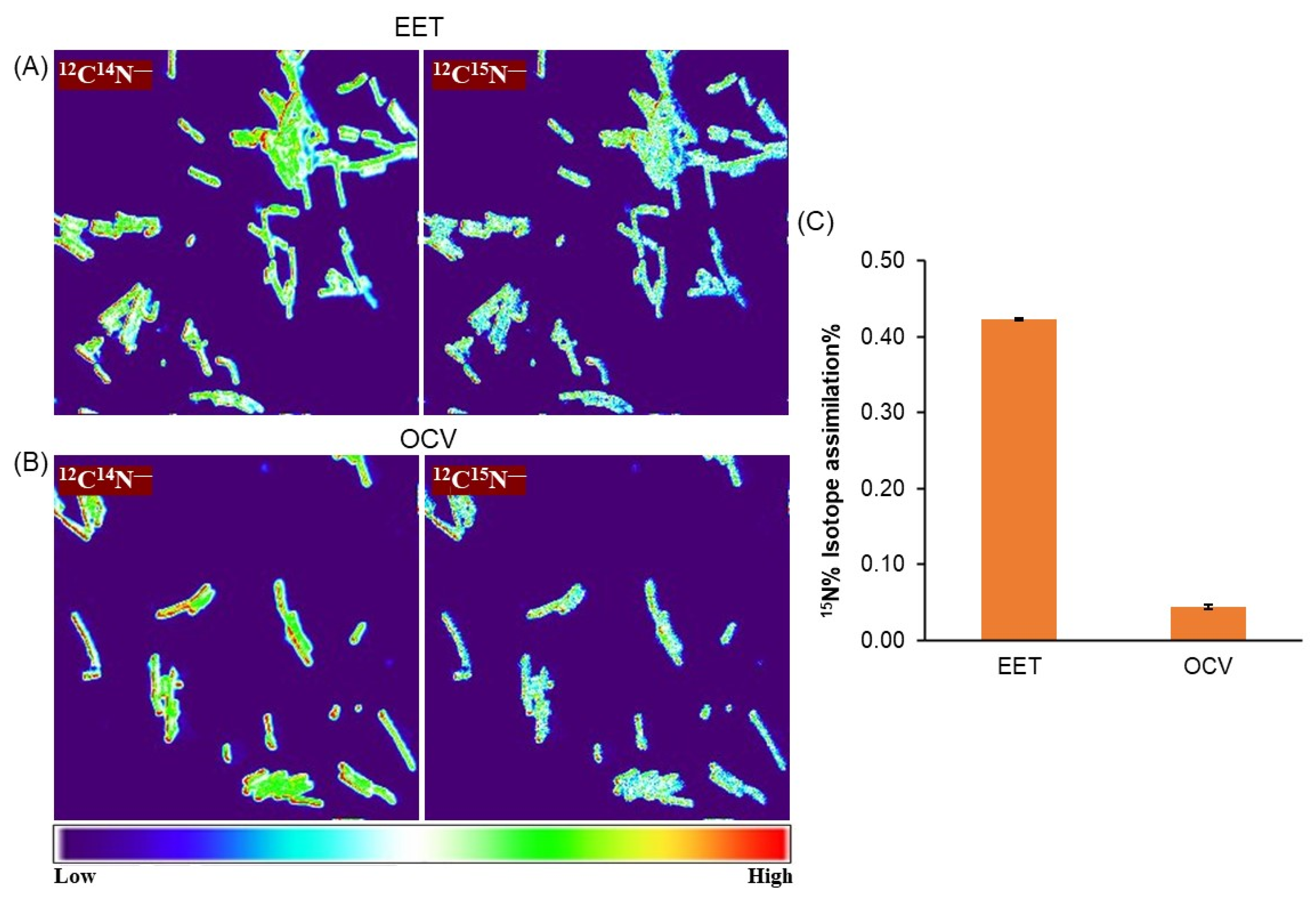
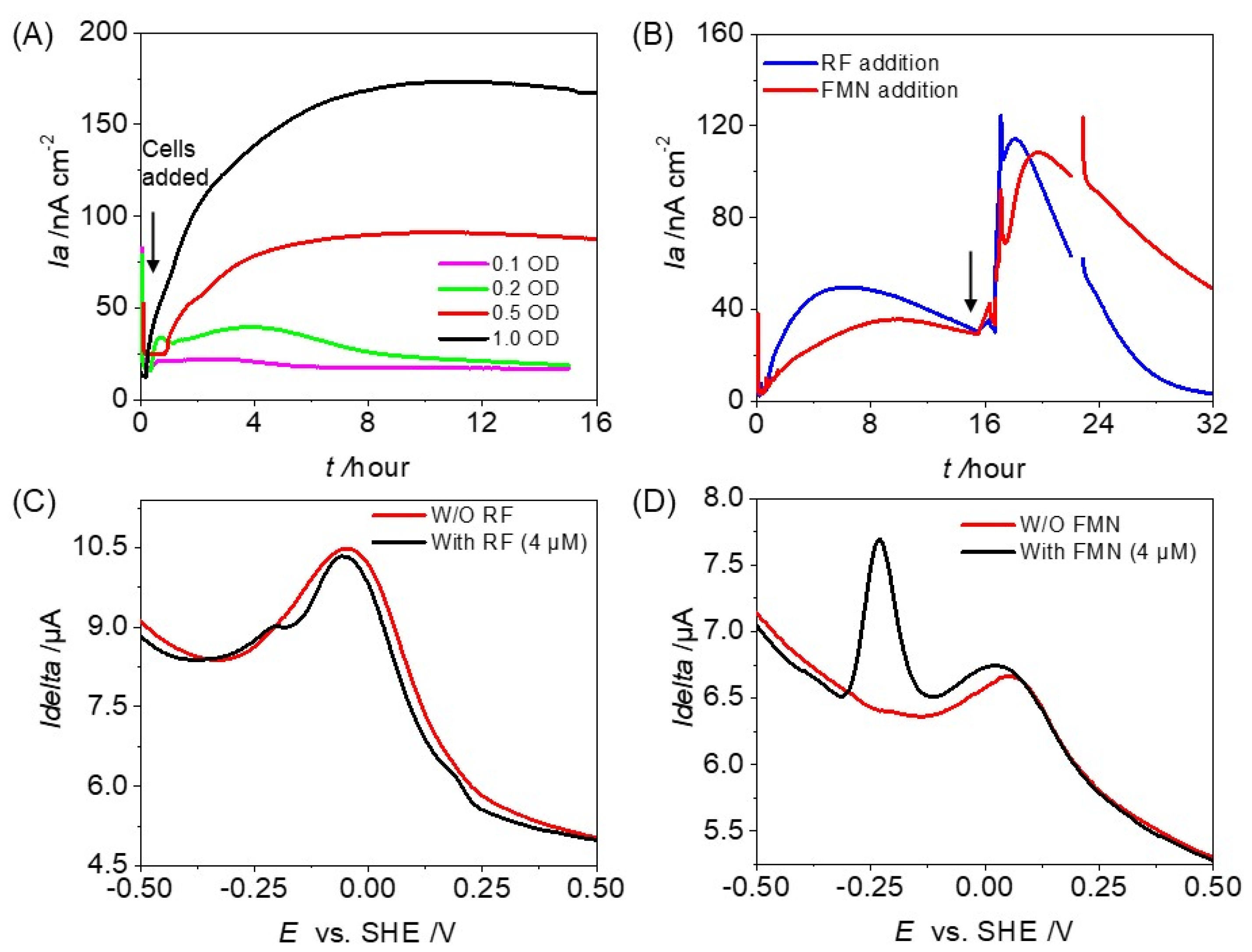
3.2. Potential Involvement of Exogenous and Endogenous Redox Mediators on the Microbial Current Production of OTU0002
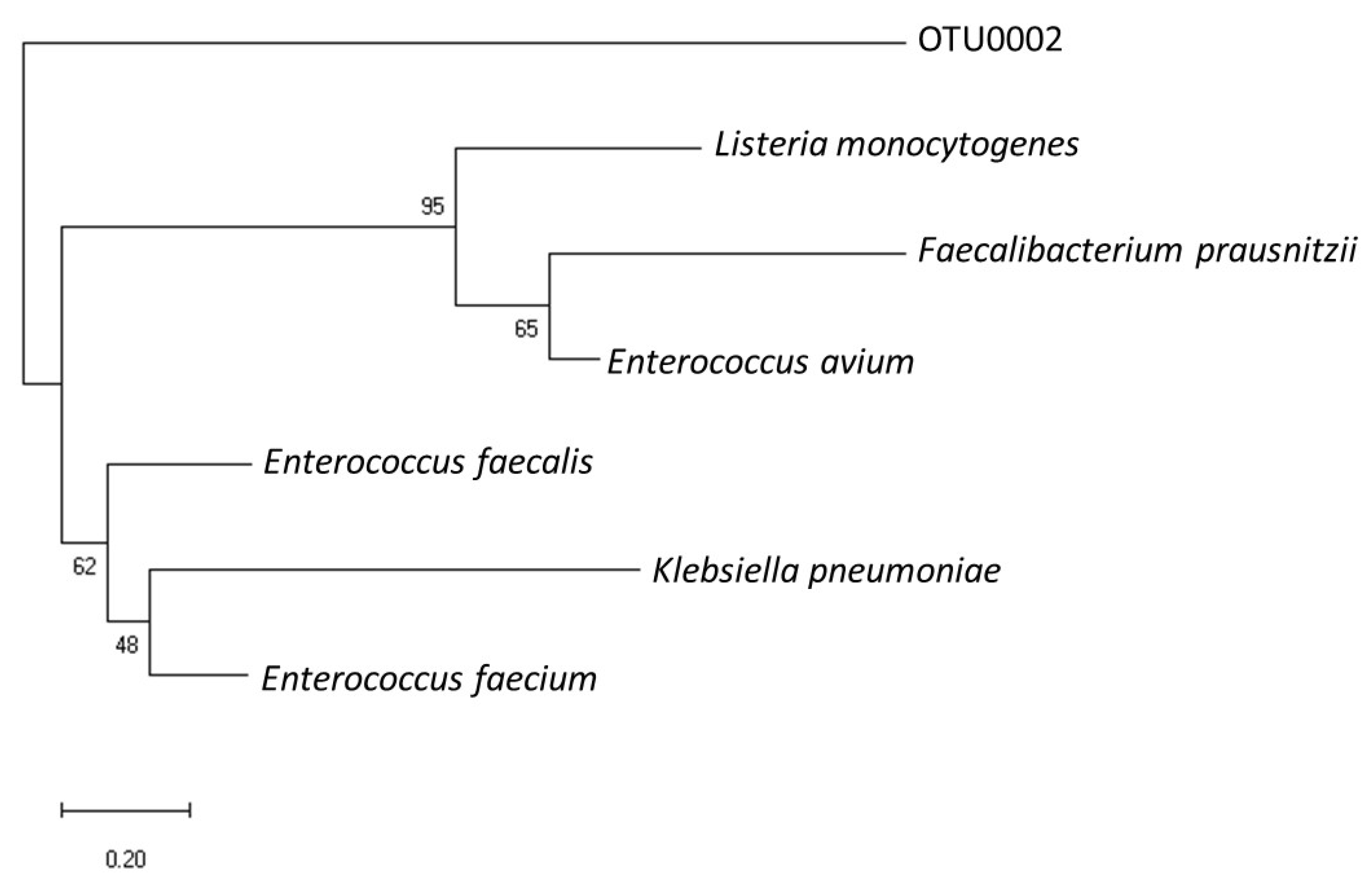
3.3. Phylogenetic Relevance of OTU0002 with Other EET-Capable Pathogens
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nealson, K.H.; Little, B. Breathing manganese and iron: Solid-state respiration. Adv. Appl. Microbiol. 1997, 45, 213–239. [Google Scholar] [CrossRef]
- Myers, C.R.; Nealson, K.H. Bacterial Manganese Reduction and Growth with Manganese Oxide as the Sole Electron Acceptor. Science 1988, 240, 1319–1321. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J.P. Novel Mode of Microbial Energy Metabolism: Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or Manganese. Appl. Environ. Microbiol. 1988, 54, 1472–1480. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Dong, H.L.; Reguera, G.; Beyenal, H.; Lu, A.H.; Liu, J.; Yu, H.Q.; Fredrickson, J.K. Extracellular electron transfer mechanisms between microorganisms and minerals. Nat. Rev. Microbiol. 2016, 14, 651–662. [Google Scholar] [CrossRef]
- Breuer, M.; Rosso, K.M.; Blumberger, J.; Butt, J.N. Multi-haem cytochromes in Shewanella oneidensis MR-1: Structures, functions and opportunities. J. R. Soc. Interface 2015, 12, 20141117. [Google Scholar] [CrossRef] [PubMed]
- Garber, A.I.; Nealson, K.H.; Okamoto, A.; McAllister, S.M.; Chan, C.S.; Barco, R.A.; Merino, N. FeGenie: A Comprehensive Tool for the Identification of Iron Genes and Iron Gene Neighborhoods in Genome and Metagenome Assemblies. Front. Microbiol. 2020, 11, 37. [Google Scholar] [CrossRef]
- Edwards, M.J.; White, G.F.; Butt, J.N.; Richardson, D.J.; Clarke, T.A. The Crystal Structure of a Biological Insulated Transmembrane Molecular Wire. Cell 2020, 181, 665–673.e610. [Google Scholar] [CrossRef] [PubMed]
- Costa, N.L.; Hermann, B.; Fourmond, V.; Faustino, M.M.; Teixeira, M.; Einsle, O.; Paquete, C.M.; Louro, R.O. How Thermophilic Gram-Positive Organisms Perform Extracellular Electron Transfer: Characterization of the Cell Surface Terminal Reductase OcwA. MBio 2019, 10, 10–1128. [Google Scholar] [CrossRef]
- Okamoto, A.; Hashimoto, K.; Nealson, K.H.; Nakamura, R. Rate enhancement of bacterial extracellular electron transport involves bound flavin semiquinones. Proc. Natl. Acad. Sci. USA 2013, 110, 7856–7861. [Google Scholar] [CrossRef]
- Xu, S.; Jangir, Y.; El-Naggar, M. Disentangling the roles of free and cytochrome-bound flavins in extracellular electron transport from Shewanella oneidensis MR-1. Electrochim. Acta 2016, 198, 49–55. [Google Scholar] [CrossRef]
- Yi, Y.; Zhao, T.; Zang, Y.; Xie, B.; Liu, H. Different mechanisms for riboflavin to improve the outward and inward extracellular electron transfer of Shewanella loihica. Electrochem. Commun. 2021, 124, 106966. [Google Scholar] [CrossRef]
- Glasser, N.R.; Saunders, S.H.; Newman, D.K. The Colorful World of Extracellular Electron Shuttles. Annu. Rev. Microbiol. 2017, 71, 731–751. [Google Scholar] [CrossRef]
- Marsili, E.; Baron, D.B.; Shikhare, I.D.; Coursolle, D.; Gralnick, J.A.; Bond, D.R. Shewanella secretes flavins that mediate extracellular electron transfer. Proc. Natl. Acad. Sci. USA 2008, 105, 3968–3973. [Google Scholar] [CrossRef] [PubMed]
- Light, S.H.; Su, L.; Rivera-Lugo, R.; Cornejo, J.A.; Louie, A.; Iavarone, A.T.; Ajo-Franklin, C.M.; Portnoy, D.A. A flavin-based extracellular electron transfer mechanism in diverse Gram-positive bacteria. Nature 2018, 562, 140–144. [Google Scholar] [CrossRef]
- Miran, W.; Naradasu, D.; Okamoto, A. Pathogens electrogenicity as a tool for in-situ metabolic activity monitoring and drug assessment in biofilms. iScience 2021, 24, 102068. [Google Scholar] [CrossRef] [PubMed]
- Deneer, H.G.; Healey, V.; Boychuk, I. Reduction of exogenous ferric iron by a surface-associated ferric reductase of Listeria spp. Microbiology 1995, 141 Pt 8, 1985–1992. [Google Scholar] [CrossRef] [PubMed]
- Keogh, D.; Lam, L.N.; Doyle, L.E.; Matysik, A.; Pavagadhi, S.; Umashankar, S.; Low, P.M.; Dale, J.L.; Song, Y.Y.; Ng, S.P.; et al. Extracellular Electron Transfer Powers Enterococcus faecalis Biofilm Metabolism. MBio 2018, 9, e00626-17. [Google Scholar] [CrossRef] [PubMed]
- Pankratova, G.; Leech, D.; Gorton, L.; Hederstedt, L. Extracellular Electron Transfer by the Gram-Positive Bacterium Enterococcus faecalis. Biochemistry 2018, 57, 4597–4603. [Google Scholar] [CrossRef]
- Naradasu, D.; Miran, W.; Sakamoto, M.; Okamoto, A. Isolation and Characterization o f Human Gut Bacteria Capable of Extracellular Electron Transport by Electrochemical Techniques. Front. Microbiol. 2019, 9, 3267. [Google Scholar] [CrossRef]
- Saunders, S.H.; Tse, E.C.M.; Yates, M.D.; Otero, F.J.; Trammell, S.A.; Stemp, E.D.A.; Barton, J.K.; Tender, L.M.; Newman, D.K. Extracellular DNA Promotes Efficient Extracellular Electron Transfer by Pyocyanin in Pseudomonas aeruginosa Biofilms. Cell 2020, 182, 919–932.e919. [Google Scholar] [CrossRef]
- Mark Welch, J.L.; Rossetti, B.J.; Rieken, C.W.; Dewhirst, F.E.; Borisy, G.G. Biogeography of a human oral microbiome at the micron scale. Proc. Natl. Acad. Sci. USA 2016, 113, E791–E800. [Google Scholar] [CrossRef]
- Kato, S.; Hashimoto, K.; Watanabe, K. Microbial interspecies electron transfer via electric currents through conductive minerals. Proc. Natl. Acad. Sci. USA 2012, 109, 10042–10046. [Google Scholar] [CrossRef]
- McGlynn, S.E.; Chadwick, G.L.; Kempes, C.P.; Orphan, V.J. Single cell activity reveals direct electron transfer in methanotrophic consortia. Nature 2015, 526, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, P.M.; Rotaru, A.-E.; Summers, Z.M.; Shrestha, M.; Liu, F.; Lovley, D.R. Transcriptomic and genetic analysis of direct interspecies electron transfer. Appl. Environ. Microbiol. 2013, 79, 2397–2404. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, E.; Kim, S.W.; Suda, W.; Kawasumi, M.; Onawa, S.; Taguchi-Atarashi, N.; Morita, H.; Taylor, T.D.; Hattori, M.; Ohno, H. Gut microorganisms act together to exacerbate inflammation in spinal cords. Nature 2020, 585, 102–106. [Google Scholar] [CrossRef]
- Roh, Y.; Gao, H.; Vali, H.; Kennedy, D.W.; Yang, Z.K.; Gao, W.; Dohnalkova, A.C.; Stapleton, R.D.; Moon, J.W.; Phelps, T.J.; et al. Metal reduction and iron biomineralization by a psychrotolerant Fe(III)-reducing bacterium, Shewanella sp. strain PV-4. Appl Environ. Microbiol 2006, 72, 3236–3244. [Google Scholar] [CrossRef] [PubMed]
- Rifkin, S.C.; Evans, D.H. Analytical evaluation of differential pulse voltammetry at stationary electrodes using computer-based instrumentation. Anal. Chem. 1976, 48, 2174–2179. [Google Scholar] [CrossRef]
- Litwin, J.A. Transition Metal-Catalyzed Oxidation of 3,3’-Diaminobenzidine [Dab] in a Model System. Acta Histochem. 1982, 71, 111–117. [Google Scholar] [CrossRef]
- Sheik, A.R.; Muller, E.E.; Audinot, J.N.; Lebrun, L.A.; Grysan, P.; Guignard, C.; Wilmes, P. In situ phenotypic heterogeneity among single cells of the filamentous bacterium Candidatus Microthrix parvicella. ISME J. 2016, 10, 1274–1279. [Google Scholar] [CrossRef]
- Long, X.Z.; Tokunou, Y.; Okamoto, A. Mechano-control of Extracellular Electron Transport Rate Modification of Inter-heme Coupling in Bacterial Surface Cytochrome. Environ. Sci. Technol. 2023, 57, 7421–7430. [Google Scholar] [CrossRef]
- Hühner, J.; Ingles-Prieto, A.; Neusüss, C.; Lämmerhofer, M.; Janovjak, H. Quantification of riboflavin, flavin mononucleotide, and flavin adenine dinucleotide in mammalian model cells by CE with LED-induced fluorescence detection. Electrophoresis 2015, 36, 518–525. [Google Scholar] [CrossRef]
- Powers, H.J. Riboflavin (vitamin B-2) and health. Am. J. Clin. Nutr. 2003, 77, 1352–1360. [Google Scholar] [CrossRef]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in Human Health and Diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef] [PubMed]
- Moscoviz, R.; Flayac, C.; Desmond-Le Quéméner, E.; Trably, E.; Bernet, N. Revealing extracellular electron transfer mediated parasitism: Energetic considerations. Sci. Rep. 2017, 7, 7766. [Google Scholar] [CrossRef] [PubMed]
- Chong, G.W.; Pirbadian, S.; El-Naggar, M.Y. Surface-Induced Formation and Redox-Dependent Staining of Outer Membrane Extensions in Shewanella oneidensis MR-1. Front. Energy Res. 2019, 7, 87. [Google Scholar] [CrossRef]
- Malvankar, N.S.; Vargas, M.; Nevin, K.P.; Franks, A.E.; Leang, C.; Kim, B.-C.; Inoue, K.; Mester, T.; Covalla, S.F.; Johnson, J.P.; et al. Tunable metallic-like conductivity in microbial nanowire networks. Nat. Nanotechnol. 2011, 6, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.; Duncan, S.H.; Stams, A.J.M.; van Dijl, J.M.; Flint, H.J.; Harmsen, H.J.M. The gut anaerobe Faecalibacterium prausnitzii uses an extracellular electron shuttle to grow at oxic–anoxic interphases. ISME J. 2012, 6, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, A.; Nakamura, R.; Nealson, K.H.; Hashimoto, K. Bound Flavin Model Suggests Similar Electron-Transfer Mechanisms in Shewanella and Geobacter. Chemelectrochem 2014, 1, 1808–1812. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naradasu, D.; Miran, W.; Okamoto, A. Electrochemical Characterization of Two Gut Microbial Strains Cooperatively Promoting Multiple Sclerosis Pathogenesis. Microorganisms 2024, 12, 257. https://doi.org/10.3390/microorganisms12020257
Naradasu D, Miran W, Okamoto A. Electrochemical Characterization of Two Gut Microbial Strains Cooperatively Promoting Multiple Sclerosis Pathogenesis. Microorganisms. 2024; 12(2):257. https://doi.org/10.3390/microorganisms12020257
Chicago/Turabian StyleNaradasu, Divya, Waheed Miran, and Akihiro Okamoto. 2024. "Electrochemical Characterization of Two Gut Microbial Strains Cooperatively Promoting Multiple Sclerosis Pathogenesis" Microorganisms 12, no. 2: 257. https://doi.org/10.3390/microorganisms12020257
APA StyleNaradasu, D., Miran, W., & Okamoto, A. (2024). Electrochemical Characterization of Two Gut Microbial Strains Cooperatively Promoting Multiple Sclerosis Pathogenesis. Microorganisms, 12(2), 257. https://doi.org/10.3390/microorganisms12020257