


Wide Distribution of Teratosphaeria epicoccoides and T. destructans Associated with Diseased Eucalyptus Leaves in Plantations in Southern China


Abstract
1. Introduction
2. Materials and Methods
2.1. Disease Symptoms, Samples, and Fungal Isolations
2.2. DNA Extraction, PCR Amplification, and Sequencing
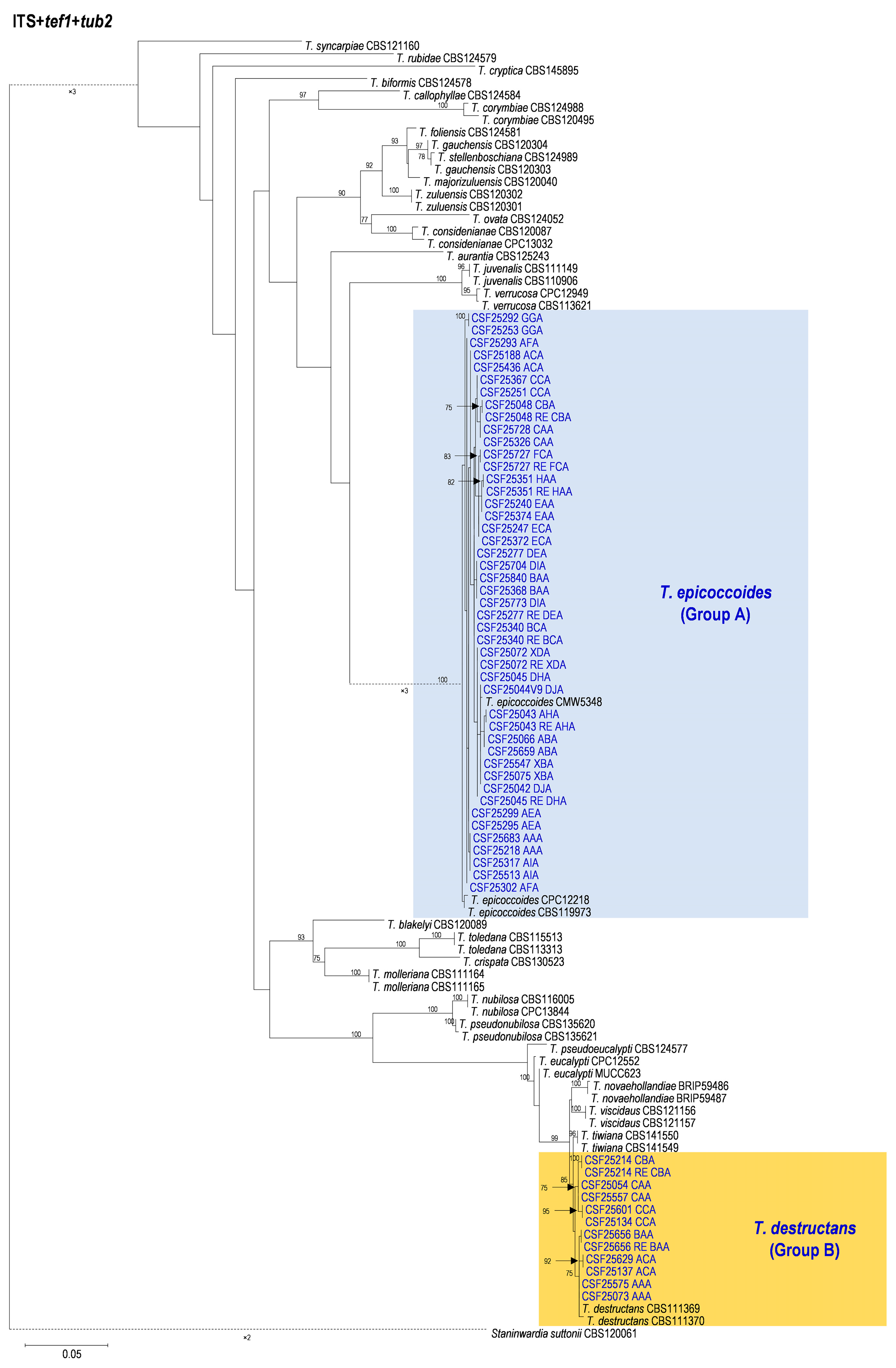
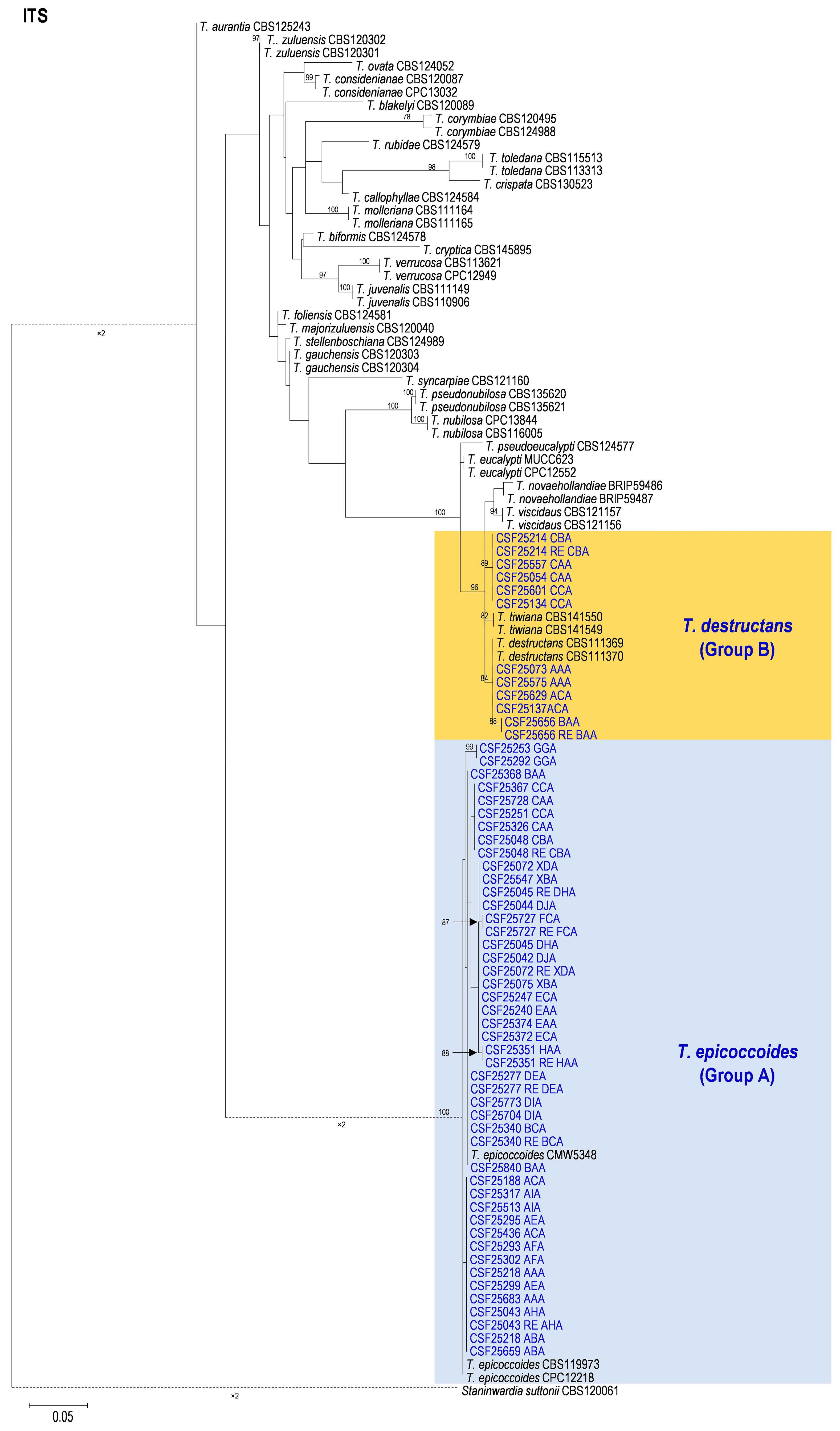
2.3. Multi-Gene Phylogenetic Analyses and Species Identification

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site No. a | Province | Isolate No. b,c | Sample and Isolate Information d | Species | GenBank Accession No. e | Genotype g | ||
|---|---|---|---|---|---|---|---|---|
| ITS | tef1 | tub2 | ||||||
| 1 | Yunnan | CSF25654 | 20230315-2-(2) | T. destructans | OR961532 | OR980975 | OR973016 | AAA |
| 1 | Yunnan | CSF25655 | 20230315-2-(3) | T. destructans | OR961533 | OR980976 | OR973017 | AAA |
| 1 | Yunnan | CSF25656 c | 20230315-2-(4) | T. destructans | OR961534 | OR980977 | OR973018 | BAA |
| 1 | Yunnan | CSF25657 | 20230315-2-(6) | T. destructans | OR961535 | OR980978 | OR973019 | AAA |
| 1 | Yunnan | CSF25658 | 20230315-2-(7) | T. destructans | OR961536 | OR980979 | OR973020 | AAA |
| 1 | Yunnan | CSF25659 c | 20230315-2-(8) | T. epicoccoides | OR961537 | OR980980 | OR973021 | ABA |
| 1 | Yunnan | CSF25660 | 20230315-2-(9) | T. destructans | OR961538 | OR980981 | OR973022 | AAA |
| 1 | Yunnan | CSF25661 | 20230315-2-(10) | T. destructans | OR961539 | OR980982 | OR973023 | AAA |
| 1 | Yunnan | CSF25686 | 20230315-2-(12) | T. epicoccoides | OR961540 | OR980983 | OR973024 | AAA |
| 1 | Yunnan | CSF25687 | 20230315-2-(13) | T. epicoccoides | OR961541 | OR980984 | OR973025 | ECA |
| 1 | Yunnan | CSF25688 | 20230315-2-(15) | T. epicoccoides | OR961542 | OR980985 | OR973026 | ECA |
| 1 | Yunnan | CSF25689 | 20230315-2-(16) | T. epicoccoides | OR961543 | OR980986 | OR973027 | ACA |
| 1 | Yunnan | CSF25690 | 20230315-2-(17) | T. epicoccoides | OR961544 | OR980987 | OR973028 | AAA |
| 1 | Yunnan | CSF25691 | 20230315-2-(19) | T. epicoccoides | OR961545 | OR980988 | OR973029 | AAA |
| 2 | Yunnan | CSF25640 | 20230315-1-(1) | T. destructans | OR961546 | OR980989 | OR973030 | AAA |
| 2 | Yunnan | CSF25641 | 20230315-1-(2) | T. destructans | OR961547 | OR980990 | OR973031 | AAA |
| 2 | Yunnan | CSF25642 | 20230315-1-(3) | T. destructans | OR961548 | OR980991 | OR973032 | AAA |
| 2 | Yunnan | CSF25643 | 20230315-1-(4) | T. destructans | OR961549 | OR980992 | OR973033 | AAA |
| 2 | Yunnan | CSF25644 | 20230315-1-(5) | T. destructans | OR961550 | OR980993 | OR973034 | AAA |
| 2 | Yunnan | CSF25645 | 20230315-1-(6) | T. destructans | OR961551 | OR980994 | OR973035 | AAA |
| 2 | Yunnan | CSF25646 | 20230315-1-(7) | T. destructans | OR961552 | OR980995 | OR973036 | AAA |
| 2 | Yunnan | CSF25647 | 20230315-1-(8) | T. destructans | OR961553 | OR980996 | OR973037 | AAA |
| 2 | Yunnan | CSF25648 | 20230315-1-(9) | T. destructans | OR961554 | OR980997 | OR973038 | AAA |
| 2 | Yunnan | CSF25649 | 20230315-1-(10) | T. destructans | OR961555 | OR980998 | OR973039 | AAA |
| 2 | Yunnan | CSF25650 | 20230315-1-(11) | T. destructans | OR961556 | OR980999 | OR973040 | AAA |
| 2 | Yunnan | CSF25651 | 20230315-1-(12) | T. destructans | OR961557 | OR981000 | OR973041 | AAA |
| 2 | Yunnan | CSF25652 | 20230315-1-(13) | T. destructans | OR961558 | OR981001 | OR973042 | AAA |
| 2 | Yunnan | CSF25653 | 20230315-1-(14) | T. destructans | OR961559 | OR981002 | OR973043 | AAA |
| 2 | Yunnan | CSF25683 c | 20230315-1-(24) | T. epicoccoides | OR961560 | OR981003 | OR973044 | AAA |
| 2 | Yunnan | CSF25685 | 20230315-1-(29) | T. epicoccoides | OR961561 | OR981004 | OR973045 | AAA |
| 3 | Yunnan | CSF25671 | 20230314-2-(12) | T. epicoccoides | OR961562 | OR981005 | OR973046 | AAA |
| 3 | Yunnan | CSF25672 | 20230314-2-(13) | T. epicoccoides | OR961563 | OR981006 | OR973047 | ACA |
| 3 | Yunnan | CSF25673 | 20230314-2-(14) | T. epicoccoides | OR961564 | OR981007 | OR973048 | AAA |
| 3 | Yunnan | CSF25674 | 20230314-2-(15) | T. epicoccoides | OR961565 | OR981008 | OR973049 | EAA |
| 3 | Yunnan | CSF25675 | 20230314-2-(16) | T. epicoccoides | OR961566 | OR981009 | OR973050 | AAA |
| 3 | Yunnan | CSF25676 | 20230314-2-(17) | T. epicoccoides | OR961567 | OR981010 | OR973051 | AAA |
| 4 | Yunnan | CSF25637 | 20230314-1-(4) | T. destructans | OR961568 | OR981011 | OR973052 | AAA |
| 4 | Yunnan | CSF25638 | 20230314-1-(10) | T. destructans | OR961569 | OR981012 | OR973053 | AAA |
| 4 | Yunnan | CSF25639 | 20230314-1-(13) | T. destructans | OR961570 | OR981013 | OR973054 | AAA |
| 5 | Yunnan | CSF25662 | 20230315-3-(1) | T. destructans | OR961571 | OR981014 | OR973055 | AAA |
| 5 | Yunnan | CSF25663 | 20230315-3-(2) | T. destructans | OR961572 | OR981015 | OR973056 | AAA |
| 5 | Yunnan | CSF25664 | 20230315-3-(3) | T. destructans | OR961573 | OR981016 | OR973057 | AAA |
| 5 | Yunnan | CSF25665 | 20230315-3-(4) | T. destructans | OR961574 | OR981017 | OR973058 | AAA |
| 5 | Yunnan | CSF25666 | 20230315-3-(5) | T. destructans | OR961575 | OR981018 | OR973059 | AAA |
| 5 | Yunnan | CSF25667 | 20230315-3-(6) | T. destructans | OR961576 | OR981019 | OR973060 | AAA |
| 5 | Yunnan | CSF25668 | 20230315-3-(7) | T. destructans | OR961577 | OR981020 | OR973061 | AAA |
| 5 | Yunnan | CSF25669 | 20230315-3-(8) | T. destructans | OR961578 | OR981021 | OR973062 | AAA |
| 5 | Yunnan | CSF25670 | 20230315-3-(9) | T. destructans | OR961579 | OR981022 | OR973063 | AAA |
| 5 | Yunnan | CSF25698 | 20230315-3-(16) | T. epicoccoides | OR961580 | OR981023 | OR973064 | AAA |
| 5 | Yunnan | CSF25699 | 20230315-3-(17) | T. epicoccoides | OR961581 | OR981024 | OR973065 | ACA |
| 5 | Yunnan | CSF25700 | 20230315-3-(18) | T. epicoccoides | OR961582 | OR981025 | OR973066 | EAA |
| 5 | Yunnan | CSF25701 | 20230315-3-(19) | T. epicoccoides | OR961583 | OR981026 | OR973067 | EAA |
| 5 | Yunnan | CSF25702 | 20230315-3-(21) | T. epicoccoides | OR961584 | OR981027 | OR973068 | ACA |
| 6 | Yunnan | CSF25703 | 20230316-2-(1) | T. epicoccoides | OR961585 | OR981028 | OR973069 | DIA |
| 6 | Yunnan | CSF25704 c | 20230316-2-(2) | T. epicoccoides | OR961586 | OR981029 | OR973070 | DIA |
| 6 | Yunnan | CSF25705 | 20230316-2-(4) | T. epicoccoides | OR961587 | OR981030 | OR973071 | DIA |
| 6 | Yunnan | CSF25706 | 20230316-2-(6) | T. epicoccoides | OR961588 | OR981031 | OR973072 | DIA |
| 6 | Yunnan | CSF25707 | 20230316-2-(7) | T. epicoccoides | OR961589 | OR981032 | OR973073 | DIA |
| 6 | Yunnan | CSF25708 | 20230316-2-(8) | T. epicoccoides | OR961590 | OR981033 | OR973074 | DIA |
| 7 | Yunnan | CSF25713 | 20230317-1-(1) | T. epicoccoides | OR961591 | OR981034 | OR973075 | AAA |
| 7 | Yunnan | CSF25714 | 20230317-1-(2) | T. epicoccoides | OR961592 | OR981035 | OR973076 | AAA |
| 7 | Yunnan | CSF25715 | 20230317-1-(4) | T. epicoccoides | OR961593 | OR981036 | OR973077 | EAA |
| 7 | Yunnan | CSF25716 | 20230317-1-(6) | T. epicoccoides | OR961594 | OR981037 | OR973078 | EAA |
| 7 | Yunnan | CSF25717 | 20230317-1-(7) | T. epicoccoides | OR961595 | OR981038 | OR973079 | ECA |
| 7 | Yunnan | CSF25718 | 20230317-1-(8) | T. epicoccoides | OR961596 | OR981039 | OR973080 | AAA |
| 8 | Yunnan | CSF25725 | 20230318-1-(1) | T. epicoccoides | OR961597 | OR981040 | OR973081 | ECA |
| 8 | Yunnan | CSF25726 | 20230318-1-(2) | T. epicoccoides | OR961598 | OR981041 | OR973082 | EAA |
| 8 | Yunnan | CSF25727 c | 20230318-1-(3) | T. epicoccoides | OR961599 | OR981042 | OR973083 | FCA |
| 8 | Yunnan | CSF25728 c | 20230318-1-(4) | T. epicoccoides | OR961600 | OR981043 | OR973084 | CAA |
| 8 | Yunnan | CSF25729 | 20230318-1-(5) | T. epicoccoides | OR961601 | OR981044 | OR973085 | EAA |
| 8 | Yunnan | CSF25730 | 20230318-1-(6) | T. epicoccoides | OR961602 | OR981045 | OR973086 | EAA |
| 9 | Yunnan | CSF25737 | 20230318-2-(1) | T. epicoccoides | OR961603 | OR981046 | OR973087 | AAA |
| 9 | Yunnan | CSF25738 | 20230318-2-(2) | T. epicoccoides | OR961604 | OR981047 | OR973088 | EAA |
| 9 | Yunnan | CSF25739 | 20230318-2-(3) | T. epicoccoides | OR961605 | OR981048 | OR973089 | EAA |
| 9 | Yunnan | CSF25740 | 20230318-2-(4) | T. epicoccoides | OR961606 | OR981049 | OR973090 | AAA |
| 9 | Yunnan | CSF25741 | 20230318-2-(5) | T. epicoccoides | OR961607 | OR981050 | OR973091 | EAA |
| 9 | Yunnan | CSF25742 | 20230318-2-(6) | T. epicoccoides | OR961608 | OR981051 | OR973092 | EAA |
| 10 | Yunnan | CSF25750 | 20230318-3-(1) | T. epicoccoides | OR961609 | OR981052 | OR973093 | EAA |
| 10 | Yunnan | CSF25751 | 20230318-3-(2) | T. epicoccoides | OR961610 | OR981053 | OR973094 | EAA |
| 10 | Yunnan | CSF25752 | 20230318-3-(3) | T. epicoccoides | OR961611 | OR981054 | OR973095 | EAA |
| 10 | Yunnan | CSF25753 | 20230318-3-(4) | T. epicoccoides | OR961612 | OR981055 | OR973096 | CAA |
| 10 | Yunnan | CSF25754 | 20230318-3-(7) | T. epicoccoides | OR961613 | OR981056 | OR973097 | AAA |
| 10 | Yunnan | CSF25755 | 20230318-3-(8) | T. epicoccoides | OR961614 | OR981057 | OR973098 | AAA |
| 11 | Yunnan | CSF25763 | 20230319-1-(2) | T. epicoccoides | OR961615 | OR981058 | OR973099 | EAA |
| 11 | Yunnan | CSF25764 | 20230319-1-(3) | T. epicoccoides | OR961616 | OR981059 | OR973100 | ACA |
| 11 | Yunnan | CSF25765 | 20230319-1-(5) | T. epicoccoides | OR961617 | OR981060 | OR973101 | AAA |
| 11 | Yunnan | CSF25766 | 20230319-1-(6) | T. epicoccoides | OR961618 | OR981061 | OR973102 | CAA |
| 11 | Yunnan | CSF25767 | 20230319-1-(7) | T. epicoccoides | OR961619 | OR981062 | OR973103 | AAA |
| 12 | Guangxi | CSF25066 c | 20220703-1-(1) | T. epicoccoides | OR961620 | OR981063 | OR973104 | ABA |
| 12 | Guangxi | CSF25067 | 20220703-1-(3) | T. destructans | OR961621 | OR981064 | OR973105 | CAA |
| 12 | Guangxi | CSF25068 | 20220703-1-(4) | T. destructans | OR961622 | OR981065 | OR973106 | CAA |
| 12 | Guangxi | CSF25069 | 20220703-1-(5) | T. destructans | OR961623 | OR981066 | OR973107 | AAA |
| 12 | Guangxi | CSF25070 | 20220703-1-(6) | T. destructans | OR961624 | OR981067 | OR973108 | CAA |
| 12 | Guangxi | CSF25071 | 20220703-1-(8) | T. destructans | OR961625 | OR981068 | OR973109 | AAA |
| 12 | Guangxi | CSF25072 c | 20220703-1-(10) | T. epicoccoides | OR961626 | OR981069 | OR973110 | XDA |
| 12 | Guangxi | CSF25218 c | 20220703-1-(12) | T. epicoccoides | OR961627 | OR981070 | OR973111 | AAA |
| 12 | Guangxi | CSF25219 | 20220703-1-(14) | T. epicoccoides | OR961628 | OR981071 | OR973112 | CAA |
| 12 | Guangxi | CSF25220 | 20220703-1-(15) | T. epicoccoides | OR961629 | OR981072 | OR973113 | ECA |
| 12 | Guangxi | CSF25222 | 20220703-1-(18) | T. epicoccoides | OR961630 | OR981073 | OR973114 | CAA |
| 12 | Guangxi | CSF25223 | 20220703-1-(19) | T. epicoccoides | OR961631 | OR981074 | OR973115 | BAA |
| 13 | Guangxi | CSF25083 | 20220703-3-(1) | T. destructans | OR961632 | OR981075 | OR973116 | AAA |
| 13 | Guangxi | CSF25084 | 20220703-3-(2) | T. destructans | OR961633 | OR981076 | OR973117 | CAA |
| 13 | Guangxi | CSF25085 | 20220703-3-(3) | T. destructans | OR961634 | OR981077 | OR973118 | AAA |
| 13 | Guangxi | CSF25086 | 20220703-3-(4) | T. destructans | OR961635 | OR981078 | OR973119 | CAA |
| 13 | Guangxi | CSF25087 | 20220703-3-(5) | T. destructans | OR961636 | OR981079 | OR973120 | CAA |
| 13 | Guangxi | CSF25088 | 20220703-3-(6) | T. epicoccoides | OR961637 | OR981080 | OR973121 | XBA |
| 13 | Guangxi | CSF25089 | 20220703-3-(7) | T. destructans | OR961638 | OR981081 | OR973122 | AAA |
| 13 | Guangxi | CSF25090 | 20220703-3-(9) | T. destructans | OR961639 | OR981082 | OR973123 | CAA |
| 13 | Guangxi | CSF25091 | 20220703-3-(10) | T. destructans | OR961640 | OR981083 | OR973124 | AAA |
| 13 | Guangxi | CSF25240 c | 20220703-3-(17) | T. epicoccoides | OR961641 | OR981084 | OR973125 | EAA |
| 13 | Guangxi | CSF25241 | 20220703-3-(19) | T. epicoccoides | OR961642 | OR981085 | OR973126 | EAA |
| 13 | Guangxi | CSF25242 | 20220703-3-(20) | T. epicoccoides | OR961643 | OR981086 | OR973127 | EAA |
| 13 | Guangxi | CSF25243 | 20220703-3-(22) | T. epicoccoides | OR961644 | OR981087 | OR973128 | ACA |
| 13 | Guangxi | CSF25244 | 20220703-3-(23) | T. epicoccoides | OR961645 | OR981088 | OR973129 | AAA |
| 13 | Guangxi | CSF25245 | 20220703-3-(24) | T. epicoccoides | OR961646 | OR981089 | OR973130 | ACA |
| 14 | Guangxi | CSF25073 c | 20220703-2-(1) | T. destructans | OR961647 | OR981090 | OR973131 | AAA |
| 14 | Guangxi | CSF25074 | 20220703-2-(2) | T. destructans | OR961648 | OR981091 | OR973132 | AAA |
| 14 | Guangxi | CSF25075 c | 20220703-2-(3) | T. epicoccoides | OR961649 | OR981092 | OR973133 | XBA |
| 14 | Guangxi | CSF25076 | 20220703-2-(4) | T. destructans | OR961650 | OR981093 | OR973134 | AAA |
| 14 | Guangxi | CSF25077 | 20220703-2-(5) | T. destructans | OR961651 | OR981094 | OR973135 | AAA |
| 14 | Guangxi | CSF25078 | 20220703-2-(6) | T. destructans | OR961652 | OR981095 | OR973136 | AAA |
| 14 | Guangxi | CSF25079 | 20220703-2-(7) | T. epicoccoides | OR961653 | OR981096 | OR973137 | ABA |
| 14 | Guangxi | CSF25080 | 20220703-2-(8) | T. destructans | OR961654 | OR981097 | OR973138 | CAA |
| 14 | Guangxi | CSF25081 | 20220703-2-(9) | T. epicoccoides | OR961655 | OR981098 | OR973139 | XBA |
| 14 | Guangxi | CSF25082 | 20220703-2-(10) | T. destructans | OR961656 | OR981099 | OR973140 | CAA |
| 14 | Guangxi | CSF25231 | 20220703-2-(21) | T. epicoccoides | OR961657 | OR981100 | OR973141 | AAA |
| 14 | Guangxi | CSF25232 | 20220703-2-(22) | T. epicoccoides | OR961658 | OR981101 | OR973142 | AAA |
| 14 | Guangxi | CSF25233 | 20220703-2-(25) | T. epicoccoides | OR961659 | OR981102 | OR973143 | EAA |
| 15 | Guangxi | CSF25092 | 20220703-4-(1) | T. destructans | OR961660 | OR981103 | OR973144 | AAA |
| 15 | Guangxi | CSF25093 | 20220703-4-(3) | T. destructans | OR961661 | OR981104 | OR973145 | AAA |
| 15 | Guangxi | CSF25094 | 20220703-4-(4) | T. destructans | OR961662 | OR981105 | OR973146 | CAA |
| 15 | Guangxi | CSF25095 | 20220703-4-(5) | T. destructans | OR961663 | OR981106 | OR973147 | CAA |
| 15 | Guangxi | CSF25096 | 20220703-4-(6) | T. destructans | OR961664 | OR981107 | OR973148 | AAA |
| 15 | Guangxi | CSF25097 | 20220703-4-(7) | T. destructans | OR961665 | OR981108 | OR973149 | AAA |
| 15 | Guangxi | CSF25098 | 20220703-4-(8) | T. destructans | OR961666 | OR981109 | OR973150 | CAA |
| 15 | Guangxi | CSF25099 | 20220703-4-(9) | T. destructans | OR961667 | OR981110 | OR973151 | CAA |
| 15 | Guangxi | CSF25247 c | 20220703-4-(10) | T. epicoccoides | OR961668 | OR981111 | OR973152 | ECA |
| 15 | Guangxi | CSF25248 | 20220703-4-(11) | T. epicoccoides | OR961669 | OR981112 | OR973153 | AAA |
| 16 | Guangxi | CSF25056 | 20220702-3-(1) | T. destructans | OR961670 | OR981113 | OR973154 | CAA |
| 16 | Guangxi | CSF25057 | 20220702-3-(2) | T. destructans | OR961671 | OR981114 | OR973155 | CAA |
| 16 | Guangxi | CSF25058 | 20220702-3-(3) | T. destructans | – f | OR981115 | OR973156 | -AA |
| 16 | Guangxi | CSF25059 | 20220702-3-(4) | T. destructans | OR961672 | OR981116 | OR973157 | CAA |
| 16 | Guangxi | CSF25060 | 20220702-3-(5) | T. destructans | OR961673 | OR981117 | OR973158 | AAA |
| 16 | Guangxi | CSF25061 | 20220702-3-(6) | T. destructans | OR961674 | OR981118 | OR973159 | CAA |
| 16 | Guangxi | CSF25062 | 20220702-3-(7) | T. destructans | OR961675 | OR981119 | OR973160 | AAA |
| 16 | Guangxi | CSF25063 | 20220702-3-(8) | T. destructans | OR961676 | OR981120 | OR973161 | CAA |
| 16 | Guangxi | CSF25064 | 20220702-3-(9) | T. destructans | OR961677 | OR981121 | OR973162 | CAA |
| 16 | Guangxi | CSF25065 | 20220702-3-(10) | T. destructans | OR961678 | OR981122 | OR973163 | CAA |
| 16 | Guangxi | CSF25214 c | 20220702-3-(17) | T. destructans | OR961679 | OR981123 | OR973164 | CBA |
| 16 | Guangxi | CSF25215 | 20220702-3-(18) | T. epicoccoides | OR961680 | OR981124 | OR973165 | ECA |
| 16 | Guangxi | CSF25216 | 20220702-3-(19) | T. epicoccoides | OR961681 | OR981125 | OR973166 | ECA |
| 16 | Guangxi | CSF25217 | 20220702-3-(20) | T. epicoccoides | OR961682 | OR981126 | OR973167 | EAA |
| 17 | Guangxi | CSF25046 | 20220702-2-(1) | T. destructans | OR961683 | OR981127 | OR973168 | CAA |
| 17 | Guangxi | CSF25047 | 20220702-2-(2) | T. destructans | OR961684 | OR981128 | OR973169 | AAA |
| 17 | Guangxi | CSF25048 c | 20220702-2-(3) | T. epicoccoides | OR961685 | OR981129 | OR973170 | CBA |
| 17 | Guangxi | CSF25049 | 20220702-2-(4) | T. destructans | OR961686 | OR981130 | OR973171 | AAA |
| 17 | Guangxi | CSF25050 | 20220702-2-(5) | T. destructans | OR961687 | OR981131 | OR973172 | AAA |
| 17 | Guangxi | CSF25051 | 20220702-2-(6) | T. destructans | OR961688 | OR981132 | OR973173 | CAA |
| 17 | Guangxi | CSF25052 | 20220702-2-(7) | T. destructans | OR961689 | OR981133 | OR973174 | AAA |
| 17 | Guangxi | CSF25053 | 20220702-2-(8) | T. destructans | OR961690 | OR981134 | OR973175 | AAA |
| 17 | Guangxi | CSF25054 c | 20220702-2-(9) | T. destructans | OR961691 | OR981135 | OR973176 | CAA |
| 17 | Guangxi | CSF25055 | 20220702-2-(10) | T. destructans | OR961692 | OR981136 | OR973177 | AAA |
| 17 | Guangxi | CSF25204 | 20220702-2-(17) | T. epicoccoides | OR961693 | OR981137 | OR973178 | EAA |
| 17 | Guangxi | CSF25205 | 20220702-2-(18) | T. epicoccoides | OR961694 | OR981138 | OR973179 | EAA |
| 17 | Guangxi | CSF25206 | 20220702-2-(19) | T. epicoccoides | OR961695 | OR981139 | OR973180 | EAA |
| 17 | Guangxi | CSF25207 | 20220702-2-(20) | T. epicoccoides | OR961696 | OR981140 | OR973181 | ECA |
| 18 | Guangxi | CSF25036 | 20220702-1-(1) | T. destructans | OR961697 | OR981141 | – | AA- |
| 18 | Guangxi | CSF25037 | 20220702-1-(2) | T. destructans | OR961698 | OR981142 | OR973182 | AAA |
| 18 | Guangxi | CSF25038 | 20220702-1-(3) | T. destructans | OR961699 | OR981143 | OR973183 | CAA |
| 18 | Guangxi | CSF25039 | 20220702-1-(4) | T. destructans | OR961700 | OR981144 | OR973184 | AAA |
| 18 | Guangxi | CSF25040 | 20220702-1-(5) | T. destructans | OR961701 | OR981145 | OR973185 | AAA |
| 18 | Guangxi | CSF25041 | 20220702-1-(6) | T. destructans | OR961702 | OR981146 | OR973186 | AAA |
| 18 | Guangxi | CSF25042 c | 20220702-1-(7) | T. epicoccoides | OR961703 | OR981147 | OR973187 | DJA |
| 18 | Guangxi | CSF25043 c | 20220702-1-(8) | T. epicoccoides | OR961704 | OR981148 | OR973188 | AHA |
| 18 | Guangxi | CSF25044 c | 20220702-1-(9) | T. epicoccoides | OR961705 | OR981149 | OR973189 | DJA |
| 18 | Guangxi | CSF25045 c | 20220702-1-(10) | T. epicoccoides | OR961706 | OR981150 | OR973190 | DHA |
| 18 | Guangxi | CSF25188 c | 20220702-1-(11) | T. epicoccoides | OR961707 | OR981151 | OR973191 | ACA |
| 18 | Guangxi | CSF25189 | 20220702-1-(12) | T. epicoccoides | OR961708 | OR981152 | OR973192 | ACA |
| 18 | Guangxi | CSF25192 | 20220702-1-(15) | T. epicoccoides | OR961709 | OR981153 | OR973193 | ACA |
| 18 | Guangxi | CSF25196 | 20220702-1-(19) | T. epicoccoides | OR961710 | OR981154 | OR973194 | AEA |
| 19 | Guangxi | CSF25100 | 20220704-1-(1) | T. destructans | OR961711 | OR981155 | OR973195 | CAA |
| 19 | Guangxi | CSF25101 | 20220704-1-(2) | T. destructans | OR961712 | OR981156 | OR973196 | CAA |
| 19 | Guangxi | CSF25102 | 20220704-1-(3) | T. destructans | OR961713 | OR981157 | OR973197 | CCA |
| 19 | Guangxi | CSF25103 | 20220704-1-(4) | T. destructans | OR961714 | OR981158 | OR973198 | AAA |
| 19 | Guangxi | CSF25104 | 20220704-1-(5) | T. destructans | OR961715 | OR981159 | OR973199 | CAA |
| 19 | Guangxi | CSF25105 | 20220704-1-(6) | T. destructans | OR961716 | OR981160 | OR973200 | CAA |
| 19 | Guangxi | CSF25106 | 20220704-1-(7) | T. destructans | OR961717 | OR981161 | OR973201 | CAA |
| 19 | Guangxi | CSF25107 | 20220704-1-(8) | T. destructans | OR961718 | OR981162 | OR973202 | CAA |
| 19 | Guangxi | CSF25108 | 20220704-1-(12) | T. destructans | OR961719 | OR981163 | OR973203 | CAA |
| 19 | Guangxi | CSF25109 | 20220704-1-(13) | T. destructans | OR961720 | OR981164 | OR973204 | CAA |
| 19 | Guangxi | CSF25250 | 20220704-1-(17) | T. epicoccoides | OR961721 | OR981165 | OR973205 | ECA |
| 19 | Guangxi | CSF25251 c | 20220704-1-(18) | T. epicoccoides | OR961722 | OR981166 | OR973206 | CCA |
| 19 | Guangxi | CSF25252 | 20220704-1-(19) | T. epicoccoides | OR961723 | OR981167 | OR973207 | ACA |
| 19 | Guangxi | CSF25253 c | 20220704-1-(20) | T. epicoccoides | OR961724 | OR981168 | OR973208 | GGA |
| 19 | Guangxi | CSF25254 | 20220704-1-(21) | T. epicoccoides | OR961725 | OR981169 | OR973209 | ACA |
| 20 | Guangxi | CSF25110 | 20220704-2-(1) | T. destructans | OR961726 | OR981170 | OR973210 | CAA |
| 20 | Guangxi | CSF25111 | 20220704-2-(2) | T. destructans | OR961727 | OR981171 | OR973211 | AAA |
| 20 | Guangxi | CSF25112 | 20220704-2-(4) | T. destructans | OR961728 | OR981172 | OR973212 | AAA |
| 20 | Guangxi | CSF25113 | 20220704-2-(5) | T. destructans | OR961729 | OR981173 | OR973213 | AAA |
| 20 | Guangxi | CSF25114 | 20220704-2-(6) | T. destructans | OR961730 | OR981174 | OR973214 | CAA |
| 20 | Guangxi | CSF25115 | 20220704-2-(7) | T. destructans | OR961731 | OR981175 | OR973215 | AAA |
| 20 | Guangxi | CSF25116 | 20220704-2-(8) | T. destructans | OR961732 | OR981176 | OR973216 | CAA |
| 20 | Guangxi | CSF25117 | 20220704-2-(10) | T. destructans | OR961733 | OR981177 | OR973217 | AAA |
| 20 | Guangxi | CSF25118 | 20220704-2-(11) | T. destructans | OR961734 | OR981178 | OR973218 | AAA |
| 20 | Guangxi | CSF25119 | 20220704-2-(12) | T. destructans | OR961735 | OR981179 | OR973219 | AAA |
| 20 | Guangxi | CSF25120 | 20220704-2-(13) | T. destructans | OR961736 | OR981180 | OR973220 | CAA |
| 20 | Guangxi | CSF25121 | 20220704-2-(14) | T. destructans | OR961737 | OR981181 | OR973221 | ACA |
| 20 | Guangxi | CSF25122 | 20220704-2-(15) | T. destructans | OR961738 | OR981182 | OR973222 | AAA |
| 20 | Guangxi | CSF25123 | 20220704-2-(16) | T. destructans | OR961739 | OR981183 | OR973223 | CAA |
| 20 | Guangxi | CSF25258 | 20220704-2-(17) | T. epicoccoides | OR961740 | OR981184 | OR973224 | DIA |
| 20 | Guangxi | CSF25259 | 20220704-2-(18) | T. epicoccoides | OR961741 | OR981185 | OR973225 | ACA |
| 20 | Guangxi | CSF25260 | 20220704-2-(19) | T. epicoccoides | OR961742 | OR981186 | OR973226 | AFA |
| 20 | Guangxi | CSF25261 | 20220704-2-(20) | T. epicoccoides | OR961743 | OR981187 | OR973227 | ACA |
| 20 | Guangxi | CSF25262 | 20220704-2-(21) | T. epicoccoides | OR961744 | OR981188 | OR973228 | ACA |
| 20 | Guangxi | CSF25263 | 20220704-2-(22) | T. epicoccoides | OR961745 | OR981189 | OR973229 | ACA |
| 21 | Guangxi | CSF25124 | 20220704-3-(1) | T. destructans | OR961746 | OR981190 | OR973230 | CAA |
| 21 | Guangxi | CSF25125 | 20220704-3-(2) | T. destructans | OR961747 | OR981191 | OR973231 | CAA |
| 21 | Guangxi | CSF25126 | 20220704-3-(3) | T. destructans | OR961748 | OR981192 | OR973232 | CAA |
| 21 | Guangxi | CSF25127 | 20220704-3-(4) | T. destructans | OR961749 | OR981193 | OR973233 | CAA |
| 21 | Guangxi | CSF25128 | 20220704-3-(5) | T. destructans | – | – | OR973234 | --A |
| 21 | Guangxi | CSF25129 | 20220704-3-(6) | T. destructans | OR961750 | OR981194 | OR973235 | CAA |
| 21 | Guangxi | CSF25130 | 20220704-3-(7) | T. destructans | OR961751 | OR981195 | OR973236 | CAA |
| 21 | Guangxi | CSF25131 | 20220704-3-(8) | T. destructans | OR961752 | OR981196 | OR973237 | CAA |
| 21 | Guangxi | CSF25132 | 20220704-3-(9) | T. destructans | OR961753 | OR981197 | OR973238 | AAA |
| 21 | Guangxi | CSF25133 | 20220704-3-(10) | T. destructans | OR961754 | OR981198 | OR973239 | CAA |
| 21 | Guangxi | CSF25270 | 20220704-3-(11) | T. epicoccoides | OR961755 | OR981199 | OR973240 | ACA |
| 21 | Guangxi | CSF25271 | 20220704-3-(13) | T. epicoccoides | OR961756 | OR981200 | OR973241 | EAA |
| 21 | Guangxi | CSF25272 | 20220704-3-(14) | T. epicoccoides | OR961757 | OR981201 | OR973242 | EAA |
| 21 | Guangxi | CSF25273 | 20220704-3-(16) | T. epicoccoides | OR961758 | OR981202 | OR973243 | ACA |
| 21 | Guangxi | CSF25274 | 20220704-3-(18) | T. epicoccoides | OR961759 | – | OR973244 | A-A |
| 21 | Guangxi | CSF25275 | 20220704-3-(20) | T. epicoccoides | OR961760 | OR981203 | OR973245 | EAA |
| 22 | Hainan | CSF25309 | 20221026-6-(1) | T. epicoccoides | OR961761 | OR981204 | OR973246 | ECA |
| 22 | Hainan | CSF25310 | 20221026-6-(2) | T. epicoccoides | OR961762 | OR981205 | OR973247 | ECA |
| 22 | Hainan | CSF25311 | 20221026-6-(3) | T. epicoccoides | OR961763 | OR981206 | OR973248 | EAA |
| 23 | Hainan | CSF25158 | 20221026-7-(1) | T. destructans | OR961764 | OR981207 | OR973249 | CAA |
| 23 | Hainan | CSF25159 | 20221026-7-(2) | T. destructans | OR961765 | OR981208 | OR973250 | CAA |
| 23 | Hainan | CSF25160 | 20221026-7-(3) | T. destructans | OR961766 | OR981209 | OR973251 | CAA |
| 23 | Hainan | CSF25161 | 20221026-7-(4) | T. destructans | OR961767 | OR981210 | OR973252 | AAA |
| 23 | Hainan | CSF25162 | 20221026-7-(5) | T. destructans | OR961768 | OR981211 | OR973253 | CAA |
| 23 | Hainan | CSF25163 | 20221026-7-(6) | T. destructans | OR961769 | OR981212 | OR973254 | AAA |
| 23 | Hainan | CSF25164 | 20221026-7-(7) | T. destructans | OR961770 | OR981213 | OR973255 | AAA |
| 23 | Hainan | CSF25165 | 20221026-7-(8) | T. destructans | OR961771 | OR981214 | OR973256 | AAA |
| 23 | Hainan | CSF25166 | 20221026-7-(9) | T. destructans | OR961772 | OR981215 | OR973257 | AAA |
| 23 | Hainan | CSF25167 | 20221026-7-(10) | T. destructans | OR961773 | OR981216 | OR973258 | CAA |
| 23 | Hainan | CSF25168 | 20221026-7-(11) | T. destructans | OR961774 | OR981217 | OR973259 | CAA |
| 23 | Hainan | CSF25312 | 20221026-7-(12) | T. epicoccoides | OR961775 | OR981218 | OR973260 | ACA |
| 23 | Hainan | CSF25313 | 20221026-7-(13) | T. epicoccoides | OR961776 | OR981219 | OR973261 | ACA |
| 23 | Hainan | CSF25314 | 20221026-7-(14) | T. epicoccoides | OR961777 | OR981220 | OR973262 | ACA |
| 23 | Hainan | CSF25315 | 20221026-7-(15) | T. epicoccoides | OR961778 | OR981221 | OR973263 | DIA |
| 23 | Hainan | CSF25316 | 20221026-7-(16) | T. epicoccoides | OR961779 | OR981222 | OR973264 | ACA |
| 23 | Hainan | CSF25317 c | 20221026-7-(17) | T. epicoccoides | OR961780 | OR981223 | OR973265 | AIA |
| 24 | Hainan | CSF25144 | 20221026-4-(1) | T. destructans | OR961781 | OR981224 | OR973266 | CAA |
| 24 | Hainan | CSF25145 | 20221026-4-(2) | T. destructans | OR961782 | OR981225 | OR973267 | CAA |
| 24 | Hainan | CSF25146 | 20221026-4-(3) | T. destructans | OR961783 | OR981226 | OR973268 | AAA |
| 24 | Hainan | CSF25147 | 20221026-4-(4) | T. destructans | OR961784 | OR981227 | OR973269 | AAA |
| 24 | Hainan | CSF25292 c | 20221026-4-(5) | T. epicoccoides | OR961785 | OR981228 | OR973270 | GGA |
| 24 | Hainan | CSF25293 c | 20221026-4-(6) | T. epicoccoides | OR961786 | OR981229 | OR973271 | AFA |
| 24 | Hainan | CSF25294 | 20221026-4-(7) | T. epicoccoides | – | OR981230 | OR973272 | -EA |
| 24 | Hainan | CSF25295 c | 20221026-4-(8) | T. epicoccoides | OR961787 | OR981231 | OR973273 | AEA |
| 24 | Hainan | CSF25296 | 20221026-4-(9) | T. epicoccoides | OR961788 | OR981232 | OR973274 | EAA |
| 24 | Hainan | CSF25297 | 20221026-4-(10) | T. epicoccoides | OR961789 | OR981233 | OR973275 | DIA |
| 25 | Hainan | CSF25148 | 20221026-5-(1) | T. destructans | OR961790 | OR981234 | OR973276 | CAA |
| 25 | Hainan | CSF25149 | 20221026-5-(2) | T. destructans | OR961791 | OR981235 | OR973277 | AAA |
| 25 | Hainan | CSF25150 | 20221026-5-(3) | T. destructans | OR961792 | OR981236 | OR973278 | AAA |
| 25 | Hainan | CSF25151 | 20221026-5-(4) | T. destructans | OR961793 | OR981237 | OR973279 | CAA |
| 25 | Hainan | CSF25152 | 20221026-5-(5) | T. destructans | OR961794 | OR981238 | OR973280 | CAA |
| 25 | Hainan | CSF25153 | 20221026-5-(6) | T. destructans | OR961795 | OR981239 | OR973281 | AAA |
| 25 | Hainan | CSF25154 | 20221026-5-(7) | T. destructans | OR961796 | OR981240 | OR973282 | ACA |
| 25 | Hainan | CSF25155 | 20221026-5-(8) | T. destructans | OR961797 | OR981241 | OR973283 | CAA |
| 25 | Hainan | CSF25156 | 20221026-5-(9) | T. destructans | OR961798 | – | OR973284 | C-A |
| 25 | Hainan | CSF25157 | 20221026-5-(10) | T. destructans | OR961799 | OR981242 | OR973285 | AAA |
| 25 | Hainan | CSF25299 c | 20221026-5-(11) | T. epicoccoides | OR961800 | OR981243 | OR973286 | AEA |
| 25 | Hainan | CSF25300 | 20221026-5-(12) | T. epicoccoides | OR961801 | OR981244 | OR973287 | AEA |
| 25 | Hainan | CSF25301 | 20221026-5-(13) | T. epicoccoides | OR961802 | OR981245 | OR973288 | AIA |
| 25 | Hainan | CSF25302 c | 20221026-5-(14) | T. epicoccoides | OR961803 | OR981246 | OR973289 | AFA |
| 25 | Hainan | CSF25303 | 20221026-5-(15) | T. epicoccoides | OR961804 | OR981247 | OR973290 | AAA |
| 26 | Hainan | CSF25134 c | 20221026-2-(1) | T. destructans | OR961805 | OR981248 | OR973291 | CCA |
| 26 | Hainan | CSF25135 | 20221026-2-(2) | T. destructans | OR961806 | OR981249 | OR973292 | ACA |
| 26 | Hainan | CSF25136 | 20221026-2-(3) | T. destructans | OR961807 | OR981250 | OR973293 | ACA |
| 26 | Hainan | CSF25286 | 20221026-2-(4) | T. epicoccoides | – | OR981251 | OR973294 | -EA |
| 26 | Hainan | CSF25287 | 20221026-2-(5) | T. epicoccoides | OR961808 | OR981252 | OR973295 | CAA |
| 26 | Hainan | CSF25288 | 20221026-2-(6) | T. epicoccoides | OR961809 | OR981253 | OR973296 | AAA |
| 26 | Hainan | CSF25289 | 20221026-2-(7) | T. epicoccoides | – | OR981254 | OR973297 | -IA |
| 26 | Hainan | CSF25290 | 20221026-2-(8) | T. epicoccoides | OR961810 | OR981255 | – | AE- |
| 26 | Hainan | CSF25291 | 20221026-2-(9) | T. epicoccoides | – | OR981256 | OR973298 | -EA |
| 27 | Hainan | CSF25137 c | 20221026-3-(1) | T. destructans | OR961811 | OR981257 | OR973299 | ACA |
| 27 | Hainan | CSF25138 | 20221026-3-(2) | T. destructans | OR961812 | OR981258 | OR973300 | CAA |
| 27 | Hainan | CSF25139 | 20221026-3-(3) | T. destructans | OR961813 | OR981259 | OR973301 | AAA |
| 27 | Hainan | CSF25140 | 20221026-3-(4) | T. destructans | OR961814 | OR981260 | OR973302 | AAA |
| 27 | Hainan | CSF25141 | 20221026-3-(5) | T. destructans | OR961815 | OR981261 | OR973303 | AAA |
| 27 | Hainan | CSF25142 | 20221026-3-(6) | T. destructans | OR961816 | OR981262 | OR973304 | CAA |
| 27 | Hainan | CSF25143 | 20221026-3-(7) | T. destructans | OR961817 | OR981263 | OR973305 | CAA |
| 28 | Hainan | CSF25276 | 20221026-1-(51) | T. epicoccoides | OR961818 | OR981264 | OR973306 | EAA |
| 28 | Hainan | CSF25277 c | 20221026-1-(52) | T. epicoccoides | OR961819 | OR981265 | OR973307 | DEA |
| 28 | Hainan | CSF25280 | 20221026-1-(55) | T. epicoccoides | OR961820 | OR981266 | OR973308 | BAA |
| 28 | Hainan | CSF25283 | 20221026-1-(58) | T. epicoccoides | OR961821 | OR981267 | OR973309 | AAA |
| 28 | Hainan | CSF25284 | 20221026-1-(59) | T. epicoccoides | OR961822 | OR981268 | OR973310 | CAA |
| 29 | Guangdong | CSF25612 | 20230221-2-(1) | T. destructans | OR961823 | OR981269 | OR973311 | CAA |
| 29 | Guangdong | CSF25613 | 20230221-2-(2) | T. destructans | OR961824 | OR981270 | OR973312 | CAA |
| 29 | Guangdong | CSF25614 | 20230221-2-(3) | T. destructans | OR961825 | OR981271 | OR973313 | ACA |
| 29 | Guangdong | CSF25615 | 20230221-2-(4) | T. destructans | OR961826 | OR981272 | OR973314 | CAA |
| 29 | Guangdong | CSF25616 | 20230221-2-(5) | T. destructans | OR961827 | OR981273 | OR973315 | AAA |
| 29 | Guangdong | CSF25617 | 20230221-2-(6) | T. destructans | OR961828 | OR981274 | OR973316 | AAA |
| 29 | Guangdong | CSF25618 | 20230221-2-(7) | T. destructans | OR961829 | OR981275 | OR973317 | CAA |
| 29 | Guangdong | CSF25619 | 20230221-2-(8) | T. destructans | OR961830 | OR981276 | OR973318 | CAA |
| 29 | Guangdong | CSF25620 | 20230221-2-(9) | T. destructans | OR961831 | OR981277 | OR973319 | CAA |
| 29 | Guangdong | CSF25621 | 20230221-2-(10) | T. destructans | OR961832 | OR981278 | OR973320 | ACA |
| 29 | Guangdong | CSF25622 | 20230221-2-(11) | T. destructans | OR961833 | OR981279 | OR973321 | CAA |
| 29 | Guangdong | CSF25623 | 20230221-2-(12) | T. destructans | OR961834 | OR981280 | OR973322 | AAA |
| 29 | Guangdong | CSF25520 | 20230221-2-(20) | T. epicoccoides | OR961835 | OR981281 | OR973323 | EAA |
| 29 | Guangdong | CSF25521 | 20230221-2-(21) | T. epicoccoides | OR961836 | OR981282 | OR973324 | ACA |
| 29 | Guangdong | CSF25522 | 20230221-2-(22) | T. epicoccoides | OR961837 | OR981283 | OR973325 | AAA |
| 29 | Guangdong | CSF25523 | 20230221-2-(23) | T. epicoccoides | OR961838 | OR981284 | OR973326 | EAA |
| 29 | Guangdong | CSF25524 | 20230221-2-(25) | T. epicoccoides | OR961839 | OR981285 | OR973327 | EAA |
| 29 | Guangdong | CSF25525 | 20230221-2-(26) | T. epicoccoides | OR961840 | OR981286 | OR973328 | EAA |
| 30 | Guangdong | CSF25599 | 20230221-1-(1) | T. destructans | OR961841 | OR981287 | OR973329 | CAA |
| 30 | Guangdong | CSF25600 | 20230221-1-(2) | T. destructans | OR961842 | OR981288 | OR973330 | AAA |
| 30 | Guangdong | CSF25601 c | 20230221-1-(3) | T. destructans | OR961843 | OR981289 | OR973331 | CCA |
| 30 | Guangdong | CSF25602 | 20230221-1-(4) | T. destructans | OR961844 | OR981290 | OR973332 | AAA |
| 30 | Guangdong | CSF25603 | 20230221-1-(5) | T. destructans | OR961845 | OR981291 | OR973333 | AAA |
| 30 | Guangdong | CSF25604 | 20230221-1-(6) | T. destructans | OR961846 | OR981292 | OR973334 | AAA |
| 30 | Guangdong | CSF25605 | 20230221-1-(7) | T. destructans | OR961847 | OR981293 | OR973335 | CAA |
| 30 | Guangdong | CSF25606 | 20230221-1-(8) | T. destructans | OR961848 | OR981294 | OR973336 | AAA |
| 30 | Guangdong | CSF25607 | 20230221-1-(9) | T. destructans | OR961849 | OR981295 | OR973337 | ACA |
| 30 | Guangdong | CSF25608 | 20230221-1-(10) | T. destructans | OR961850 | OR981296 | OR973338 | AAA |
| 30 | Guangdong | CSF25609 | 20230221-1-(11) | T. destructans | OR961851 | OR981297 | OR973339 | CAA |
| 30 | Guangdong | CSF25610 | 20230221-1-(12) | T. destructans | OR961852 | OR981298 | OR973340 | AAA |
| 30 | Guangdong | CSF25611 | 20230221-1-(14) | T. destructans | OR961853 | OR981299 | OR973341 | AAA |
| 30 | Guangdong | CSF25512 | 20230221-1-(24) | T. epicoccoides | OR961854 | OR981300 | OR973342 | ACA |
| 30 | Guangdong | CSF25513 c | 20230221-1-(26) | T. epicoccoides | OR961855 | OR981301 | OR973343 | AIA |
| 30 | Guangdong | CSF25514 | 20230221-1-(27) | T. epicoccoides | OR961856 | OR981302 | OR973344 | ACA |
| 30 | Guangdong | CSF25515 | 20230221-1-(28) | T. epicoccoides | OR961857 | OR981303 | OR973345 | ACA |
| 30 | Guangdong | CSF25516 | 20230221-1-(29) | T. epicoccoides | OR961858 | OR981304 | OR973346 | AAA |
| 30 | Guangdong | CSF25517 | 20230221-1-(30) | T. epicoccoides | OR961859 | OR981305 | OR973347 | ACA |
| 31 | Guangdong | CSF25587 | 20230220-3-(1) | T. destructans | OR961860 | OR981306 | OR973348 | AAA |
| 31 | Guangdong | CSF25588 | 20230220-3-(2) | T. destructans | OR961861 | OR981307 | OR973349 | AAA |
| 31 | Guangdong | CSF25589 | 20230220-3-(3) | T. destructans | OR961862 | OR981308 | OR973350 | AAA |
| 31 | Guangdong | CSF25590 | 20230220-3-(4) | T. destructans | OR961863 | OR981309 | OR973351 | AAA |
| 31 | Guangdong | CSF25591 | 20230220-3-(5) | T. destructans | OR961864 | OR981310 | OR973352 | AAA |
| 31 | Guangdong | CSF25592 | 20230220-3-(6) | T. destructans | OR961865 | OR981311 | OR973353 | AAA |
| 31 | Guangdong | CSF25593 | 20230220-3-(7) | T. destructans | OR961866 | OR981312 | OR973354 | AAA |
| 31 | Guangdong | CSF25594 | 20230220-3-(8) | T. destructans | OR961867 | OR981313 | OR973355 | AAA |
| 31 | Guangdong | CSF25595 | 20230220-3-(10) | T. destructans | OR961868 | OR981314 | OR973356 | AAA |
| 31 | Guangdong | CSF25596 | 20230220-3-(11) | T. destructans | OR961869 | OR981315 | OR973357 | AAA |
| 31 | Guangdong | CSF25597 | 20230220-3-(12) | T. destructans | OR961870 | OR981316 | OR973358 | CAA |
| 31 | Guangdong | CSF25598 | 20230220-3-(13) | T. destructans | OR961871 | OR981317 | OR973359 | CAA |
| 31 | Guangdong | CSF25500 | 20230220-3-(25) | T. epicoccoides | OR961872 | OR981318 | OR973360 | ACA |
| 31 | Guangdong | CSF25501 | 20230220-3-(26) | T. epicoccoides | OR961873 | OR981319 | OR973361 | ACA |
| 31 | Guangdong | CSF25502 | 20230220-3-(27) | T. epicoccoides | OR961874 | OR981320 | OR973362 | AAA |
| 31 | Guangdong | CSF25503 | 20230220-3-(28) | T. epicoccoides | OR961875 | OR981321 | OR973363 | ACA |
| 31 | Guangdong | CSF25504 | 20230220-3-(29) | T. epicoccoides | OR961876 | OR981322 | OR973364 | ACA |
| 31 | Guangdong | CSF25505 | 20230220-3-(30) | T. epicoccoides | – | OR981323 | OR973365 | -EA |
| 32 | Guangdong | CSF25624 | 20230221-3-(3) | T. destructans | OR961877 | OR981324 | OR973366 | CCA |
| 32 | Guangdong | CSF25625 | 20230221-3-(4) | T. destructans | OR961878 | OR981325 | OR973367 | ACA |
| 32 | Guangdong | CSF25626 | 20230221-3-(5) | T. destructans | OR961879 | OR981326 | OR973368 | AAA |
| 32 | Guangdong | CSF25627 | 20230221-3-(6) | T. destructans | OR961880 | OR981327 | OR973369 | CAA |
| 32 | Guangdong | CSF25628 | 20230221-3-(7) | T. destructans | OR961881 | OR981328 | OR973370 | AAA |
| 32 | Guangdong | CSF25629 c | 20230221-3-(8) | T. destructans | OR961882 | OR981329 | OR973371 | ACA |
| 32 | Guangdong | CSF25630 | 20230221-3-(9) | T. destructans | OR961883 | OR981330 | OR973372 | AAA |
| 32 | Guangdong | CSF25631 | 20230221-3-(10) | T. destructans | OR961884 | OR981331 | OR973373 | AAA |
| 32 | Guangdong | CSF25632 | 20230221-3-(11) | T. destructans | OR961885 | OR981332 | OR973374 | AAA |
| 32 | Guangdong | CSF25633 | 20230221-3-(12) | T. destructans | OR961886 | OR981333 | OR973375 | AAA |
| 32 | Guangdong | CSF25634 | 20230221-3-(13) | T. destructans | OR961887 | OR981334 | OR973376 | CCA |
| 32 | Guangdong | CSF25635 | 20230221-3-(14) | T. destructans | OR961888 | OR981335 | OR973377 | CAA |
| 32 | Guangdong | CSF25636 | 20230221-3-(15) | T. destructans | OR961889 | OR981336 | OR973378 | AAA |
| 32 | Guangdong | CSF25531 | 20230221-3-(18) | T. epicoccoides | OR961890 | OR981337 | OR973379 | ACA |
| 32 | Guangdong | CSF25532 | 20230221-3-(19) | T. epicoccoides | – | OR981338 | OR973380 | -IA |
| 32 | Guangdong | CSF25533 | 20230221-3-(20) | T. epicoccoides | OR961891 | OR981339 | OR973381 | ACA |
| 32 | Guangdong | CSF25534 | 20230221-3-(21) | T. epicoccoides | – | OR981340 | OR973382 | -AA |
| 32 | Guangdong | CSF25535 | 20230221-3-(22) | T. epicoccoides | OR961892 | OR981341 | OR973383 | ACA |
| 32 | Guangdong | CSF25536 | 20230221-3-(23) | T. epicoccoides | – | OR981342 | OR973384 | -AA |
| 33 | Guangdong | CSF25576 | 20230220-2-(1) | T. destructans | OR961893 | OR981343 | OR973385 | AAA |
| 33 | Guangdong | CSF25577 | 20230220-2-(2) | T. destructans | OR961894 | OR981344 | OR973386 | AAA |
| 33 | Guangdong | CSF25578 | 20230220-2-(3) | T. destructans | OR961895 | OR981345 | OR973387 | AAA |
| 33 | Guangdong | CSF25579 | 20230220-2-(4) | T. destructans | OR961896 | OR981346 | OR973388 | AAA |
| 33 | Guangdong | CSF25580 | 20230220-2-(5) | T. destructans | OR961897 | OR981347 | OR973389 | AAA |
| 33 | Guangdong | CSF25581 | 20230220-2-(6) | T. destructans | OR961898 | OR981348 | OR973390 | CCA |
| 33 | Guangdong | CSF25582 | 20230220-2-(7) | T. destructans | OR961899 | OR981349 | OR973391 | AAA |
| 33 | Guangdong | CSF25583 | 20230220-2-(8) | T. destructans | OR961900 | OR981350 | OR973392 | CCA |
| 33 | Guangdong | CSF25584 | 20230220-2-(9) | T. destructans | OR961901 | OR981351 | OR973393 | AAA |
| 33 | Guangdong | CSF25585 | 20230220-2-(11) | T. destructans | OR961902 | OR981352 | OR973394 | AAA |
| 33 | Guangdong | CSF25586 | 20230220-2-(12) | T. destructans | OR961903 | OR981353 | OR973395 | CAA |
| 33 | Guangdong | CSF25497 | 20230220-2-(18) | T. epicoccoides | OR961904 | OR981354 | OR973396 | EAA |
| 33 | Guangdong | CSF25498 | 20230220-2-(19) | T. epicoccoides | OR961905 | OR981355 | OR973397 | ECA |
| 33 | Guangdong | CSF25499 | 20230220-2-(20) | T. epicoccoides | OR961906 | OR981356 | OR973398 | EAA |
| 34 | Guangdong | CSF25568 | 20230220-1-(2) | T. destructans | OR961907 | OR981357 | OR973399 | AAA |
| 34 | Guangdong | CSF25569 | 20230220-1-(5) | T. destructans | OR961908 | OR981358 | OR973400 | CCA |
| 34 | Guangdong | CSF25570 | 20230220-1-(7) | T. destructans | OR961909 | OR981359 | OR973401 | CAA |
| 34 | Guangdong | CSF25571 | 20230220-1-(9) | T. destructans | OR961910 | OR981360 | OR973402 | AAA |
| 34 | Guangdong | CSF25572 | 20230220-1-(10) | T. destructans | OR961911 | OR981361 | OR973403 | CCA |
| 34 | Guangdong | CSF25573 | 20230220-1-(11) | T. destructans | OR961912 | OR981362 | OR973404 | CAA |
| 34 | Guangdong | CSF25574 | 20230220-1-(14) | T. destructans | OR961913 | OR981363 | OR973405 | AAA |
| 34 | Guangdong | CSF25575 c | 20230220-1-(16) | T. destructans | OR961914 | OR981364 | OR973406 | AAA |
| 34 | Guangdong | CSF25495 | 20230220-1-(17) | T. epicoccoides | – | OR981365 | OR973407 | -EA |
| 34 | Guangdong | CSF25496 | 20230220-1-(18) | T. epicoccoides | – | OR981366 | OR973408 | -IA |
| 35 | Guangdong | CSF25371 | 20230216-5-(20) | T. epicoccoides | OR961915 | OR981367 | OR973409 | CAA |
| 35 | Guangdong | CSF25372 c | 20230216-5-(22) | T. epicoccoides | OR961916 | OR981368 | OR973410 | ECA |
| 35 | Guangdong | CSF25373 | 20230216-5-(24) | T. epicoccoides | OR961917 | OR981369 | OR973411 | CAA |
| 35 | Guangdong | CSF25374 c | 20230216-5-(27) | T. epicoccoides | OR961918 | OR981370 | OR973412 | EAA |
| 35 | Guangdong | CSF25375 | 20230216-5-(30) | T. epicoccoides | OR961919 | OR981371 | OR973413 | EAA |
| 35 | Guangdong | CSF25376 | 20230216-5-(32) | T. epicoccoides | OR961920 | OR981372 | OR973414 | ECA |
| 36 | Guangdong | CSF25356 | 20230216-3-(9) | T. epicoccoides | OR961921 | OR981373 | OR973415 | AAA |
| 36 | Guangdong | CSF25357 | 20230216-3-(10) | T. epicoccoides | OR961922 | OR981374 | OR973416 | EAA |
| 36 | Guangdong | CSF25358 | 20230216-3-(12) | T. epicoccoides | OR961923 | OR981375 | OR973417 | ECA |
| 36 | Guangdong | CSF25359 | 20230216-3-(14) | T. epicoccoides | OR961924 | OR981376 | OR973418 | EAA |
| 36 | Guangdong | CSF25360 | 20230216-3-(15) | T. epicoccoides | OR961925 | OR981377 | OR973419 | CAA |
| 36 | Guangdong | CSF25361 | 20230216-3-(16) | T. epicoccoides | OR961926 | OR981378 | OR973420 | ECA |
| 37 | Guangdong | CSF25367 c | 20230216-4-(1) | T. epicoccoides | OR961927 | OR981379 | OR973421 | CCA |
| 37 | Guangdong | CSF25368 c | 20230216-4-(3) | T. epicoccoides | OR961928 | OR981380 | OR973422 | BAA |
| 37 | Guangdong | CSF25369 | 20230216-4-(4) | T. epicoccoides | OR961929 | OR981381 | OR973423 | EAA |
| 37 | Guangdong | CSF25370 | 20230216-4-(5) | T. epicoccoides | OR961930 | OR981382 | OR973424 | EAA |
| 38 | Guangdong | CSF25347 | 20230216-2-(10) | T. epicoccoides | OR961931 | OR981383 | OR973425 | EAA |
| 38 | Guangdong | CSF25348 | 20230216-2-(11) | T. epicoccoides | OR961932 | OR981384 | OR973426 | AAA |
| 38 | Guangdong | CSF25349 | 20230216-2-(12) | T. epicoccoides | OR961933 | OR981385 | OR973427 | CAA |
| 38 | Guangdong | CSF25350 | 20230216-2-(13) | T. epicoccoides | OR961934 | OR981386 | OR973428 | AAA |
| 38 | Guangdong | CSF25351 c | 20230216-2-(14) | T. epicoccoides | OR961935 | OR981387 | OR973429 | HAA |
| 38 | Guangdong | CSF25352 | 20230216-2-(15) | T. epicoccoides | OR961936 | OR981388 | OR973430 | AAA |
| 39 | Guangdong | CSF25325 | 20230215-1-(1) | T. epicoccoides | OR961937 | OR981389 | OR973431 | CCA |
| 39 | Guangdong | CSF25326 c | 20230215-1-(2) | T. epicoccoides | OR961938 | OR981390 | OR973432 | CAA |
| 39 | Guangdong | CSF25327 | 20230215-1-(3) | T. epicoccoides | OR961939 | OR981391 | OR973433 | EAA |
| 39 | Guangdong | CSF25328 | 20230215-1-(6) | T. epicoccoides | OR961940 | OR981392 | OR973434 | ACA |
| 39 | Guangdong | CSF25329 | 20230215-1-(7) | T. epicoccoides | OR961941 | OR981393 | OR973435 | EAA |
| 39 | Guangdong | CSF25330 | 20230215-1-(8) | T. epicoccoides | OR961942 | OR981394 | OR973436 | EAA |
| 40 | Guangdong | CSF25336 | 20230216-1-(11) | T. epicoccoides | OR961943 | OR981395 | OR973437 | ECA |
| 40 | Guangdong | CSF25337 | 20230216-1-(13) | T. epicoccoides | OR961944 | OR981396 | OR973438 | ECA |
| 40 | Guangdong | CSF25338 | 20230216-1-(15) | T. epicoccoides | OR961945 | OR981397 | OR973439 | EAA |
| 40 | Guangdong | CSF25339 | 20230216-1-(16) | T. epicoccoides | OR961946 | OR981398 | OR973440 | AAA |
| 40 | Guangdong | CSF25340 c | 20230216-1-(17) | T. epicoccoides | OR961947 | OR981399 | OR973441 | BCA |
| 40 | Guangdong | CSF25341 | 20230216-1-(18) | T. epicoccoides | OR961948 | OR981400 | OR973442 | EAA |
| 41 | Guangdong | CSF25546 | 20230217-3-(1) | T. destructans | OR961949 | OR981401 | OR973443 | CCA |
| 41 | Guangdong | CSF25547 c | 20230217-3-(2) | T. epicoccoides | OR961950 | OR981402 | OR973444 | XBA |
| 41 | Guangdong | CSF25548 | 20230217-3-(6) | T. destructans | OR961951 | OR981403 | OR973445 | AAA |
| 41 | Guangdong | CSF25549 | 20230217-3-(8) | T. destructans | OR961952 | OR981404 | OR973446 | CAA |
| 41 | Guangdong | CSF25550 | 20230217-3-(13) | T. destructans | OR961953 | OR981405 | OR973447 | AAA |
| 41 | Guangdong | CSF25551 | 20230217-3-(16) | T. destructans | OR961954 | OR981406 | OR973448 | AAA |
| 41 | Guangdong | CSF25552 | 20230217-3-(17) | T. destructans | OR961955 | OR981407 | OR973449 | AAA |
| 41 | Guangdong | CSF25553 | 20230217-3-(21) | T. destructans | OR961956 | OR981408 | OR973450 | CAA |
| 41 | Guangdong | CSF25554 | 20230217-3-(24) | T. destructans | OR961957 | OR981409 | OR973451 | CCA |
| 41 | Guangdong | CSF25555 | 20230217-3-(25) | T. destructans | OR961958 | OR981410 | OR973452 | CAA |
| 41 | Guangdong | CSF25556 | 20230217-3-(30) | T. destructans | OR961959 | OR981411 | OR973453 | CAA |
| 41 | Guangdong | CSF25557 c | 20230217-3-(37) | T. destructans | OR961960 | OR981412 | OR973454 | CAA |
| 41 | Guangdong | CSF25399 | 20230217-3-(51) | T. epicoccoides | OR961961 | OR981413 | OR973455 | AAA |
| 41 | Guangdong | CSF25400 | 20230217-3-(52) | T. epicoccoides | OR961962 | OR981414 | OR973456 | AAA |
| 41 | Guangdong | CSF25401 | 20230217-3-(53) | T. epicoccoides | OR961963 | OR981415 | OR973457 | CAA |
| 41 | Guangdong | CSF25402 | 20230217-3-(54) | T. epicoccoides | OR961964 | OR981416 | OR973458 | ECA |
| 41 | Guangdong | CSF25403 | 20230217-3-(55) | T. epicoccoides | OR961965 | OR981417 | OR973459 | AAA |
| 41 | Guangdong | CSF25404 | 20230217-3-(56) | T. epicoccoides | OR961966 | OR981418 | OR973460 | ECA |
| 42 | Guangdong | CSF25558 | 20230219-1-(5) | T. destructans | OR961967 | OR981419 | OR973461 | AAA |
| 42 | Guangdong | CSF25559 | 20230219-1-(8) | T. destructans | OR961968 | OR981420 | OR973462 | AAA |
| 42 | Guangdong | CSF25560 | 20230219-1-(9) | T. destructans | OR961969 | OR981421 | OR973463 | CAA |
| 42 | Guangdong | CSF25561 | 20230219-1-(15) | T. destructans | OR961970 | OR981422 | OR973464 | AAA |
| 42 | Guangdong | CSF25562 | 20230219-1-(17) | T. destructans | OR961971 | OR981423 | OR973465 | AAA |
| 42 | Guangdong | CSF25563 | 20230219-1-(23) | T. destructans | OR961972 | OR981424 | OR973466 | CAA |
| 42 | Guangdong | CSF25564 | 20230219-1-(25) | T. destructans | OR961973 | OR981425 | OR973467 | CAA |
| 42 | Guangdong | CSF25565 | 20230219-1-(26) | T. destructans | OR961974 | OR981426 | OR973468 | CAA |
| 42 | Guangdong | CSF25566 | 20230219-1-(27) | T. destructans | OR961975 | OR981427 | OR973469 | AAA |
| 42 | Guangdong | CSF25567 | 20230219-1-(29) | T. destructans | OR961976 | OR981428 | OR973470 | CAA |
| 43 | Guangdong | CSF25390 | 20230217-2-(18) | T. epicoccoides | OR961977 | OR981429 | OR973471 | ECA |
| 43 | Guangdong | CSF25391 | 20230217-2-(20) | T. epicoccoides | OR961978 | OR981430 | OR973472 | AAA |
| 43 | Guangdong | CSF25392 | 20230217-2-(21) | T. epicoccoides | OR961979 | OR981431 | OR973473 | AAA |
| 43 | Guangdong | CSF25393 | 20230217-2-(24) | T. epicoccoides | OR961980 | OR981432 | OR973474 | EAA |
| 43 | Guangdong | CSF25394 | 20230217-2-(25) | T. epicoccoides | OR961981 | OR981433 | OR973475 | EAA |
| 43 | Guangdong | CSF25395 | 20230217-2-(26) | T. epicoccoides | OR961982 | OR981434 | OR973476 | EAA |
| 44 | Guangdong | CSF25378 | 20230217-1-(13) | T. epicoccoides | OR961983 | OR981435 | OR973477 | ECA |
| 44 | Guangdong | CSF25379 | 20230217-1-(14) | T. epicoccoides | OR961984 | OR981436 | OR973478 | CAA |
| 44 | Guangdong | CSF25380 | 20230217-1-(15) | T. epicoccoides | OR961985 | OR981437 | OR973479 | ECA |
| 44 | Guangdong | CSF25381 | 20230217-1-(17) | T. epicoccoides | OR961986 | OR981438 | OR973480 | EAA |
| 44 | Guangdong | CSF25382 | 20230217-1-(18) | T. epicoccoides | OR961987 | OR981439 | OR973481 | EAA |
| 44 | Guangdong | CSF25383 | 20230217-1-(21) | T. epicoccoides | OR961988 | OR981440 | OR973482 | EAA |
| 45 | Guangdong | CSF25436 c | 20230218-1-(6) | T. epicoccoides | OR961989 | OR981441 | OR973483 | ACA |
| 45 | Guangdong | CSF25437 | 20230218-1-(7) | T. epicoccoides | OR961990 | OR981442 | OR973484 | EAA |
| 45 | Guangdong | CSF25438 | 20230218-1-(8) | T. epicoccoides | OR961991 | OR981443 | OR973485 | AAA |
| 45 | Guangdong | CSF25439 | 20230218-1-(9) | T. epicoccoides | OR961992 | OR981444 | OR973486 | EAA |
| 45 | Guangdong | CSF25440 | 20230218-1-(10) | T. epicoccoides | OR961993 | OR981445 | OR973487 | EAA |
| 45 | Guangdong | CSF25441 | 20230218-1-(11) | T. epicoccoides | OR961994 | OR981446 | OR973488 | BAA |
| 46 | Guangdong | CSF25419 | 20230217-4-(3) | T. epicoccoides | OR961995 | OR981447 | OR973489 | EAA |
| 46 | Guangdong | CSF25420 | 20230217-4-(4) | T. epicoccoides | OR961996 | OR981448 | OR973490 | AAA |
| 46 | Guangdong | CSF25421 | 20230217-4-(5) | T. epicoccoides | OR961997 | OR981449 | OR973491 | AAA |
| 46 | Guangdong | CSF25422 | 20230217-4-(6) | T. epicoccoides | OR961998 | OR981450 | OR973492 | ECA |
| 46 | Guangdong | CSF25423 | 20230217-4-(7) | T. epicoccoides | – | OR981451 | OR973493 | -CA |
| 46 | Guangdong | CSF25424 | 20230217-4-(8) | T. epicoccoides | OR961999 | OR981452 | OR973494 | EAA |
| 47 | Guangdong | CSF25451 | 20230218-2-(7) | T. epicoccoides | OR962000 | OR981453 | OR973495 | AAA |
| 47 | Guangdong | CSF25452 | 20230218-2-(8) | T. epicoccoides | OR962001 | OR981454 | OR973496 | EAA |
| 47 | Guangdong | CSF25453 | 20230218-2-(9) | T. epicoccoides | OR962002 | OR981455 | OR973497 | BAA |
| 47 | Guangdong | CSF25454 | 20230218-2-(10) | T. epicoccoides | OR962003 | OR981456 | OR973498 | AAA |
| 47 | Guangdong | CSF25455 | 20230218-2-(11) | T. epicoccoides | OR962004 | OR981457 | OR973499 | ECA |
| 47 | Guangdong | CSF25456 | 20230218-2-(13) | T. epicoccoides | OR962005 | OR981458 | OR973500 | ACA |
| 48 | Guangdong | CSF25468 | 20230218-3-(12) | T. epicoccoides | OR962006 | OR981459 | OR973501 | AAA |
| 48 | Guangdong | CSF25469 | 20230218-3-(13) | T. epicoccoides | OR962007 | OR981460 | OR973502 | ACA |
| 48 | Guangdong | CSF25470 | 20230218-3-(14) | T. epicoccoides | OR962008 | OR981461 | OR973503 | AAA |
| 48 | Guangdong | CSF25471 | 20230218-3-(15) | T. epicoccoides | OR962009 | OR981462 | OR973504 | ACA |
| 48 | Guangdong | CSF25472 | 20230218-3-(16) | T. epicoccoides | OR962010 | OR981463 | OR973505 | EAA |
| 48 | Guangdong | CSF25473 | 20230218-3-(17) | T. epicoccoides | OR962011 | OR981464 | OR973506 | EAA |
| 49 | Guangdong | CSF25482 | 20230218-4-(2) | T. epicoccoides | OR962012 | OR981465 | OR973507 | AAA |
| 49 | Guangdong | CSF25483 | 20230218-4-(4) | T. epicoccoides | OR962013 | OR981466 | OR973508 | EAA |
| 49 | Guangdong | CSF25484 | 20230218-4-(5) | T. epicoccoides | OR962014 | OR981467 | OR973509 | BAA |
| 49 | Guangdong | CSF25485 | 20230218-4-(6) | T. epicoccoides | OR962015 | OR981468 | OR973510 | EAA |
| 49 | Guangdong | CSF25486 | 20230218-4-(7) | T. epicoccoides | OR962016 | OR981469 | OR973511 | ACA |
| 49 | Guangdong | CSF25487 | 20230218-4-(8) | T. epicoccoides | OR962017 | OR981470 | OR973512 | EAA |
| 50 | Fujian | CSF25801 | 20230330-1-(21) | T. epicoccoides | OR962018 | OR981471 | OR973513 | ACA |
| 50 | Fujian | CSF25802 | 20230330-1-(22) | T. epicoccoides | OR962019 | OR981472 | OR973514 | EAA |
| 50 | Fujian | CSF25803 | 20230330-1-(23) | T. epicoccoides | OR962020 | OR981473 | OR973515 | AAA |
| 50 | Fujian | CSF25804 | 20230330-1-(24) | T. epicoccoides | OR962021 | OR981474 | OR973516 | AAA |
| 50 | Fujian | CSF25805 | 20230330-1-(25) | T. epicoccoides | OR962022 | OR981475 | OR973517 | ECA |
| 51 | Fujian | CSF25773 c | 20230328-1-(2) | T. epicoccoides | OR962023 | OR981476 | OR973518 | DIA |
| 51 | Fujian | CSF25774 | 20230328-1-(5) | T. epicoccoides | OR962024 | OR981477 | OR973519 | DIA |
| 51 | Fujian | CSF25775 | 20230328-1-(7) | T. epicoccoides | OR962025 | OR981478 | OR973520 | DIA |
| 51 | Fujian | CSF25776 | 20230328-1-(8) | T. epicoccoides | OR962026 | OR981479 | OR973521 | DIA |
| 51 | Fujian | CSF25777 | 20230328-1-(9) | T. epicoccoides | OR962027 | OR981480 | OR973522 | ECA |
| 51 | Fujian | CSF25778 | 20230328-1-(11) | T. epicoccoides | OR962028 | OR981481 | OR973523 | AAA |
| 52 | Fujian | CSF25782 | 20230329-1-(1) | T. epicoccoides | – | OR981482 | OR973524 | -EA |
| 52 | Fujian | CSF25783 | 20230329-1-(2) | T. epicoccoides | OR962029 | OR981483 | OR973525 | EAA |
| 52 | Fujian | CSF25784 | 20230329-1-(3) | T. epicoccoides | OR962030 | OR981484 | OR973526 | EAA |
| 52 | Fujian | CSF25785 | 20230329-1-(4) | T. epicoccoides | OR962031 | OR981485 | OR973527 | ACA |
| 52 | Fujian | CSF25786 | 20230329-1-(5) | T. epicoccoides | – | OR981486 | OR973528 | -CA |
| 52 | Fujian | CSF25787 | 20230329-1-(6) | T. epicoccoides | OR962032 | OR981487 | OR973529 | EAA |
| 53 | Fujian | CSF25790 | 20230329-2-(1) | T. epicoccoides | OR962033 | OR981488 | OR973530 | DIA |
| 53 | Fujian | CSF25791 | 20230329-2-(4) | T. epicoccoides | OR962034 | OR981489 | OR973531 | DIA |
| 53 | Fujian | CSF25792 | 20230329-2-(5) | T. epicoccoides | OR962035 | OR981490 | OR973532 | DIA |
| 53 | Fujian | CSF25793 | 20230329-2-(6) | T. epicoccoides | OR962036 | OR981491 | OR973533 | DIA |
| 53 | Fujian | CSF25794 | 20230329-2-(7) | T. epicoccoides | – | OR981492 | OR973534 | -IA |
| 53 | Fujian | CSF25795 | 20230329-2-(8) | T. epicoccoides | OR962037 | OR981493 | OR973535 | DIA |
| 54 | Fujian | CSF25850 | 20230331-3-(1) | T. epicoccoides | OR962038 | OR981494 | OR973536 | CAA |
| 54 | Fujian | CSF25851 | 20230331-3-(2) | T. epicoccoides | OR962039 | OR981495 | OR973537 | ECA |
| 54 | Fujian | CSF25852 | 20230331-3-(4) | T. epicoccoides | OR962040 | OR981496 | OR973538 | EAA |
| 54 | Fujian | CSF25853 | 20230331-3-(5) | T. epicoccoides | OR962041 | OR981497 | OR973539 | CAA |
| 54 | Fujian | CSF25854 | 20230331-3-(6) | T. epicoccoides | OR962042 | OR981498 | OR973540 | CAA |
| 54 | Fujian | CSF25855 | 20230331-3-(7) | T. epicoccoides | OR962043 | OR981499 | OR973541 | EAA |
| 55 | Fujian | CSF25840 c | 20230331-2-(2) | T. epicoccoides | OR962044 | OR981500 | OR973542 | BAA |
| 55 | Fujian | CSF25841 | 20230331-2-(3) | T. epicoccoides | OR962045 | OR981501 | OR973543 | ECA |
| 55 | Fujian | CSF25842 | 20230331-2-(4) | T. epicoccoides | OR962046 | OR981502 | OR973544 | EAA |
| 55 | Fujian | CSF25843 | 20230331-2-(5) | T. epicoccoides | OR962047 | OR981503 | OR973545 | AAA |
| 55 | Fujian | CSF25844 | 20230331-2-(6) | T. epicoccoides | OR962048 | OR981504 | OR973546 | AAA |
| 55 | Fujian | CSF25845 | 20230331-2-(9) | T. epicoccoides | OR962049 | OR981505 | OR973547 | EAA |
| 56 | Fujian | CSF25830 | 20230331-1-(1) | T. epicoccoides | OR962050 | OR981506 | OR973548 | ACA |
| 56 | Fujian | CSF25831 | 20230331-1-(2) | T. epicoccoides | OR962051 | OR981507 | OR973549 | EAA |
| 56 | Fujian | CSF25832 | 20230331-1-(3) | T. epicoccoides | OR962052 | OR981508 | OR973550 | BAA |
| 56 | Fujian | CSF25833 | 20230331-1-(5) | T. epicoccoides | OR962053 | OR981509 | OR973551 | CCA |
| 56 | Fujian | CSF25834 | 20230331-1-(6) | T. epicoccoides | OR962054 | OR981510 | OR973552 | EAA |
| 56 | Fujian | CSF25835 | 20230331-1-(9) | T. epicoccoides | OR962055 | OR981511 | OR973553 | AAA |
| 57 | Fujian | CSF25820 | 20230330-3-(1) | T. epicoccoides | OR962056 | OR981512 | OR973554 | EAA |
| 57 | Fujian | CSF25821 | 20230330-3-(2) | T. epicoccoides | OR962057 | OR981513 | OR973555 | ECA |
| 57 | Fujian | CSF25822 | 20230330-3-(3) | T. epicoccoides | OR962058 | OR981514 | OR973556 | CAA |
| 57 | Fujian | CSF25823 | 20230330-3-(4) | T. epicoccoides | OR962059 | OR981515 | OR973557 | ECA |
| 57 | Fujian | CSF25824 | 20230330-3-(5) | T. epicoccoides | OR962060 | OR981516 | OR973558 | EAA |
| 57 | Fujian | CSF25825 | 20230330-3-(6) | T. epicoccoides | OR962061 | OR981517 | OR973559 | ECA |
| 58 | Fujian | CSF25810 | 20230330-2-(1) | T. epicoccoides | OR962062 | OR981518 | OR973560 | AAA |
| 58 | Fujian | CSF25811 | 20230330-2-(3) | T. epicoccoides | OR962063 | OR981519 | OR973561 | CCA |
| 58 | Fujian | CSF25812 | 20230330-2-(4) | T. epicoccoides | OR962064 | OR981520 | OR973562 | CAA |
| 58 | Fujian | CSF25813 | 20230330-2-(5) | T. epicoccoides | OR962065 | OR981521 | OR973563 | CCA |
| 58 | Fujian | CSF25814 | 20230330-2-(6) | T. epicoccoides | OR962066 | OR981522 | OR973564 | EAA |
| 58 | Fujian | CSF25815 | 20230330-2-(8) | T. epicoccoides | OR962067 | OR981523 | OR973565 | ACA |
| 59 | Fujian | CSF25860 | 20230401-1-(1) | T. epicoccoides | OR962068 | OR981524 | OR973566 | AAA |
| 59 | Fujian | CSF25861 | 20230401-1-(2) | T. epicoccoides | OR962069 | OR981525 | OR973567 | EAA |
| 59 | Fujian | CSF25862 | 20230401-1-(3) | T. epicoccoides | OR962070 | OR981526 | OR973568 | AAA |
| 59 | Fujian | CSF25863 | 20230401-1-(4) | T. epicoccoides | OR962071 | OR981527 | OR973569 | EAA |
| 59 | Fujian | CSF25864 | 20230401-1-(5) | T. epicoccoides | OR962072 | OR981528 | OR973570 | EAA |
| 59 | Fujian | CSF25865 | 20230401-1-(7) | T. epicoccoides | OR962073 | OR981529 | OR973571 | EAA |
3. Results
3.1. Fungal Isolations
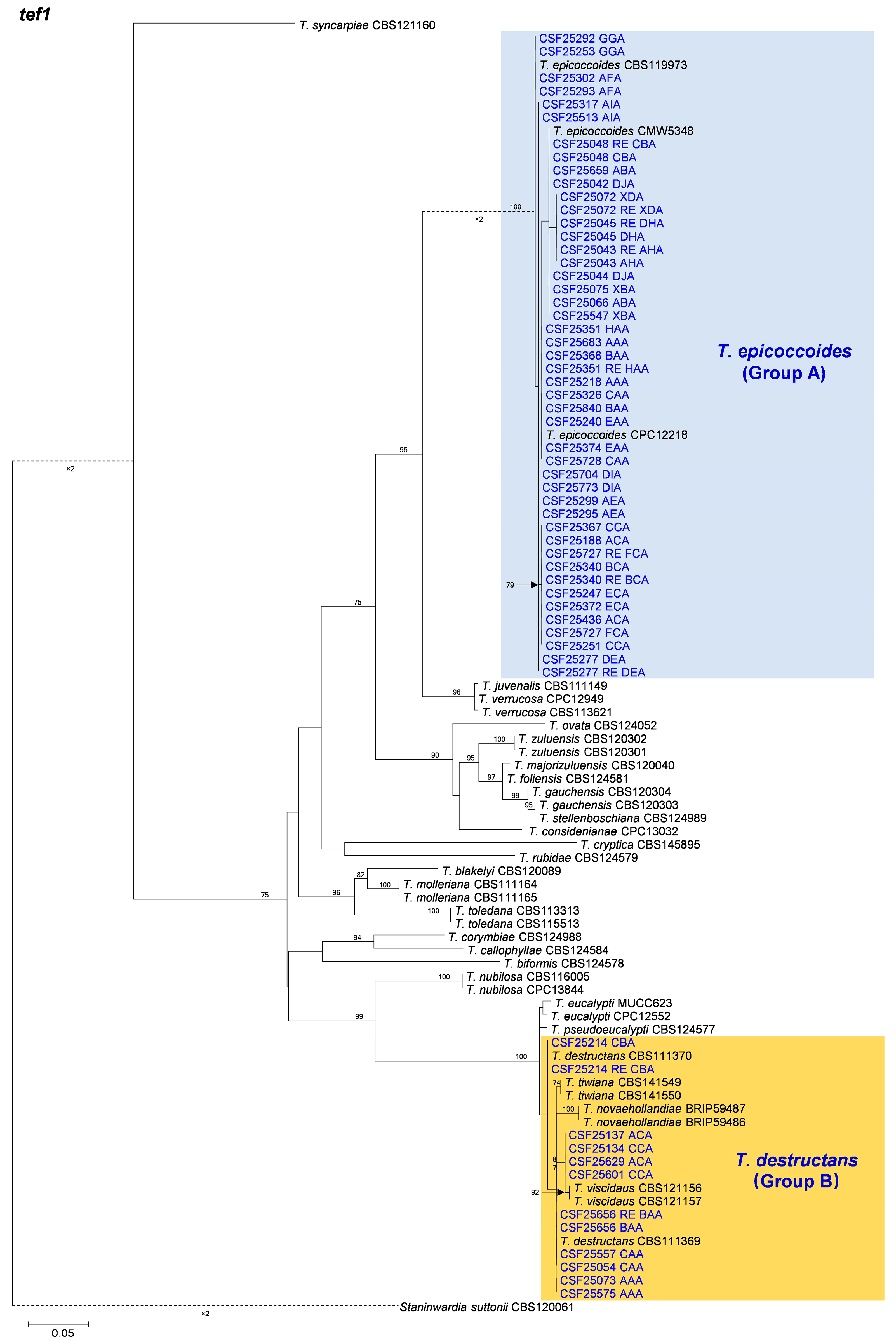
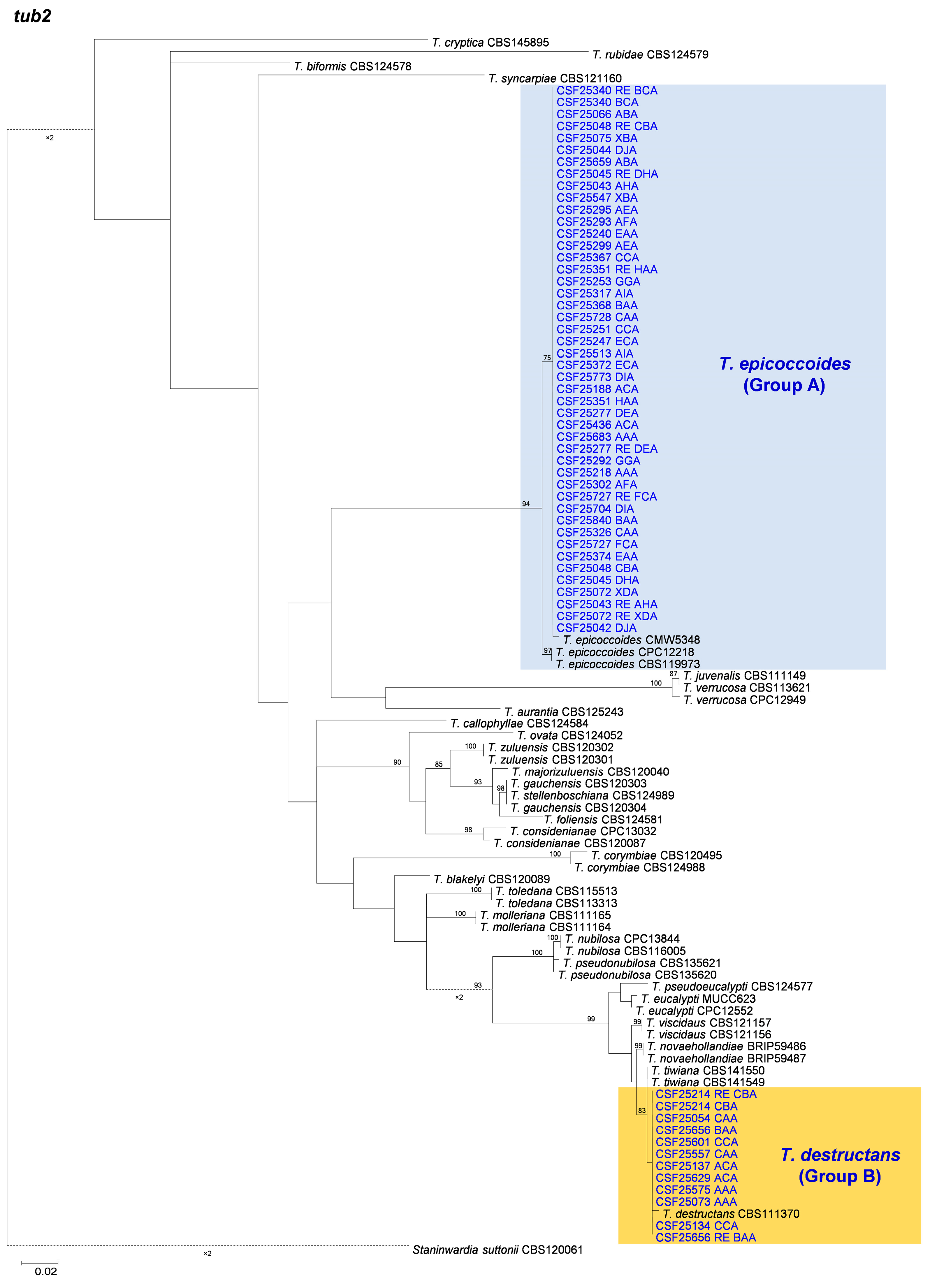
3.2. Multi-Gene Phylogenetic Analyses and Species Identification
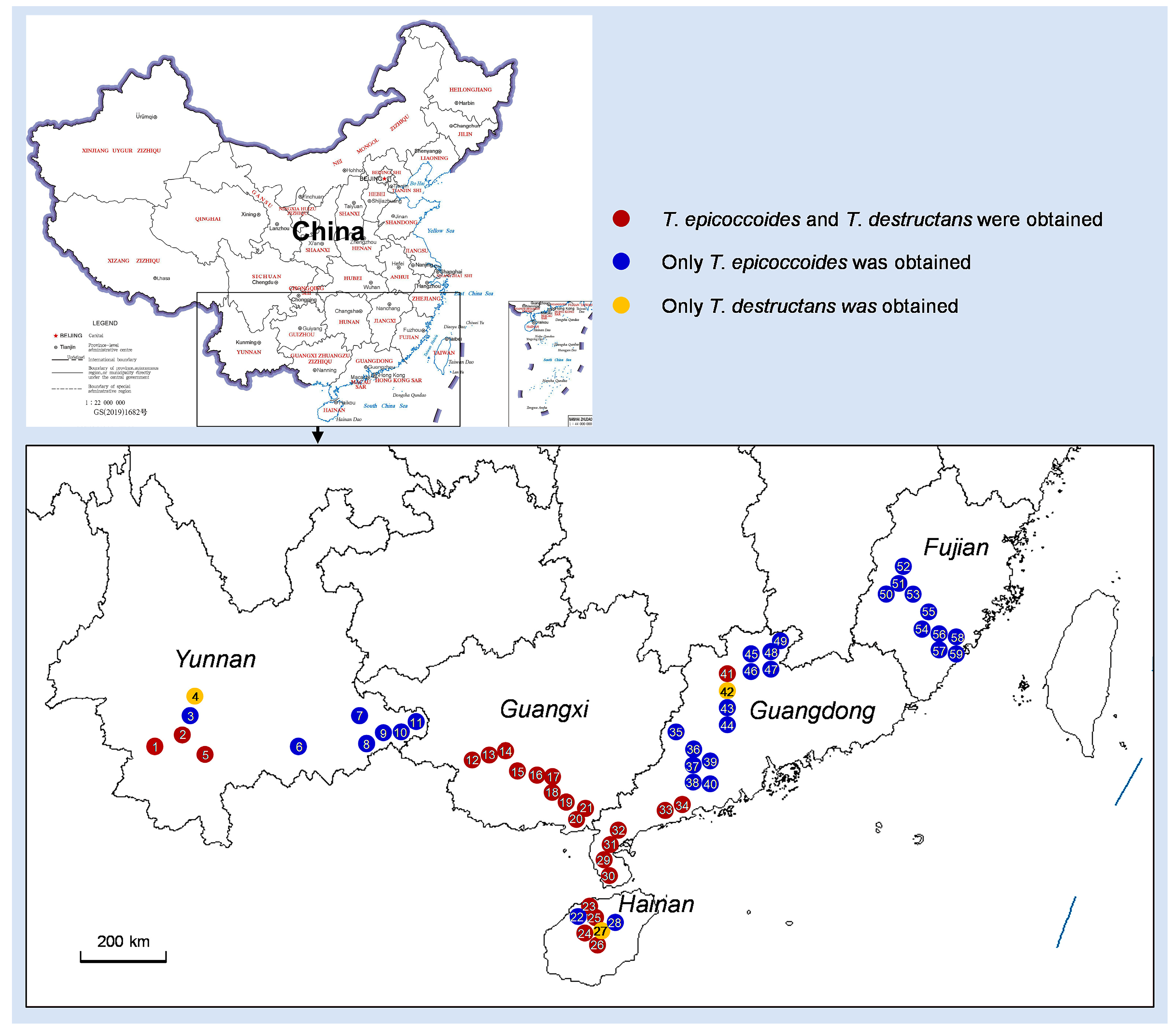
3.3. Distribution of Teratosphaeria Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xie, Y.J. Scientific innovation leads to fast development of eucalypt research and industry in China. Eucalypt Sci. Technol. 2022, 39, 35–42. [Google Scholar]
- Xie, Y.J.; Du, A.P. Eucalypts in Southern China; CCP Central Party School Press: Beijing, China, 2019. [Google Scholar]
- Xie, Y.J.; Arnold, R.J.; Wu, Z.H.; Chen, S.F.; Du, A.P.; Luo, J.Z. Advances in eucalypt research in China. Front. Agric. Sci. Eng. 2017, 4, 380–390. [Google Scholar] [CrossRef]
- Chen, S.F. Summary of fungal pathogens from eucalypts in China: 2006–2013. Eucalypt Sci. Technol. 2014, 31, 37–65. [Google Scholar]
- Turnbull, J.W. Development of Sustainable Forestry Plantations in China: A Review; Australian Centre for International Agricultural Research: Canberra, Australia, 2007.
- Zhou, X.D.; Wingfield, M.J. Eucalypt diseases and their management in China. Australas. Plant Pathol. 2011, 40, 339–345. [Google Scholar] [CrossRef]
- Old, K.M.; Wingfield, M.J.; Yuan, Z.Q. A Manual of Diseases of Eucalypts in South–East Asia; Center for International Forestry Research: Jakarta, Indonesia, 2003. [Google Scholar]
- Carstensen, G.D.; Venter, S.N.; Wingfield, M.J.; Coutinho, T.A. Two Ralstonia species associated with bacterial wilt of Eucalyptus. Plant Pathol. 2017, 66, 393–403. [Google Scholar] [CrossRef]
- Chen, S.F.; Gryzenhout, M.; Roux, J.; Xie, Y.J.; Wingfield, M.J.; Zhou, X.D. Identification and pathogenicity of Chrysoporthe cubensis on Eucalyptus and Syzygium spp. in South China. Plant Dis. 2010, 94, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.F.; Barnes, I.; Chungu, D.; Roux, J.; Wingfield, M.J.; Xie, Y.J.; Zhou, X.D. High population diversity and increasing importance of the Eucalyptus stem canker pathogen, Teratosphaeria zuluensis, in South China. Australas. Plant Pathol. 2011, 40, 407–415. [Google Scholar] [CrossRef]
- Li, G.Q.; Liu, F.F.; Li, J.Q.; Liu, Q.L. Botryosphaeriaceae from Eucalyptus plantations and adjacent plants in China. Persoonia 2018, 40, 63–95. [Google Scholar] [CrossRef]
- Wang, W.; Li, G.Q.; Liu, Q.L.; Chen, S.F. Cryphonectriaceae on Myrtales in China: Phylogeny, host range, and pathogenicity. Persoonia 2020, 45, 101–131. [Google Scholar] [CrossRef]
- Burgess, T.I.; Andjic, V.; Hardy, G.E.S.; Dell, B.; Xu, D. First report of Phaeophleospora destructans in China. J. Trop. For. Sci. 2006, 18, 144–146. [Google Scholar]
- Chen, S.F.; Lombard, L.; Roux, J.; Xie, Y.J.; Wingfield, M.J.; Zhou, X.D. Novel species of Calonectria associated with Eucalyptus leaf blight in Southeast China. Persoonia 2011, 26, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.C.; Chen, S.F. Calonectria pentaseptata causes severe leaf disease of cultivated Eucalyptus on the Leizhou Peninsula of southern China. Plant Dis. 2020, 104, 493–509. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.X.; Chen, S.F. Species diversity, mating strategy and pathogenicity of Calonectria species from diseased leaves and soils in the Eucalyptus plantation in southern China. J. Fungi 2021, 7, 73. [Google Scholar] [CrossRef]
- Liang, X.Y.; Wang, Q.C.; Chen, S.F. Phylogeny, morphology, distribution, and pathogenicity of seven Calonectria species from leaf blighted Eucalyptus in HaiNai Island, China. Plant Dis. 2023, 107, 2579–2605. [Google Scholar] [CrossRef]
- Zhou, X.D.; de Beer, Z.W.; Xie, Y.J.; Pegg, G.S.; Wingfield, M.J. DNA based identification of Quambalaria pitereka causing severe leaf blight of Corymbia citriodora in China. Fungal Divers. 2007, 25, 245–254. [Google Scholar]
- Chen, S.F.; Liu, Q.L.; Li, G.Q.; Wingfield, M.J. Quambalaria species associated with eucalypt diseases in southern China. Front. Agric. Sci. Eng. 2017, 4, 433–447. [Google Scholar] [CrossRef]
- Hunter, G.C.; Wingfield, B.D.; Crous, P.W.; Wingfield, M.J. A multi–gene phylogeny for species of Mycosphaerella occurring on Eucalyptus leaves. Stud. Mycol. 2006, 55, 147–161. [Google Scholar] [CrossRef]
- Pérez, C.A.; Wingfield, M.J.; Altier, N.A.; Blanchette, R.A. Mycosphaerellaceae and Teratosphaeriaceae associated with Eucalyptus leaf diseases and stem cankers in Uruguay. For. Pathol. 2009, 39, 349–360. [Google Scholar] [CrossRef]
- Carnegie, A.J.; Pegg, G.S.; White, D.; Burgess, T.I. Species within Mycosphaerellaceae and Teratosphaeriaceae from eucalypts in eastern Australia. Australas. Plant Pathol. 2011, 40, 366–384. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Cheewangkoon, R.; Carnegie, A.J.; Burgess, T.I.; Summerell, B.A.; Edwards, J.; Taylor, P.W.J.; Groenewald, J.Z. Foliar pathogens of eucalypts. Stud. Mycol. 2019, 94, 125–298. [Google Scholar] [CrossRef]
- Pérez, G.; Burgess, T.I.; Slippers, B.; Carnegie, A.J.; Wingfield, B.D.; Wingfield, M.J. Teratosphaeria pseudonubilosa sp. nov., a serious Eucalyptus leaf pathogen in the Teratosphaeria nubilosa species complex. Australas. Plant Pathol. 2014, 43, 67–77. [Google Scholar] [CrossRef]
- Andjic, V.; Maxwell, A.; Hardy, G.E.S.; Burgess, T.I. New cryptic species of Teratosphaeria on Eucalyptus in Australia. IMA Fungus 2016, 7, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Andjic, V.; Hardy, G.E.S.; Cortinas, M.N.; Wingfield, M.J.; Burgess, T.I. Multiple gene genealogies reveal important relationships between species of Phaeophleospora infecting Eucalyptus leaves. FEMS Microbiol. Lett. 2007, 268, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Burgess, T.I.; Barber, P.A.; Sufaati, S.; Xu, D.P.; Hardy, G.E.S.; Dell, B. Mycosphaerella spp. on Eucalyptus in Asia: New species, new hosts and new records. Fungal Divers. 2007, 24, 135–157. [Google Scholar]
- Huang, C.L.; Chen, W.W.; Luo, J.T.; Tan, X.F.; Yang, X.H.; Wei, J.G. Identification of fungal pathogens of fast–growing Eucalyptus leaf disease in Guangxi. For. Pest Dis. 2012, 31, 1–6. [Google Scholar]
- Crous, P.W.; Braun, U.; Hunter, G.C.; Wingfield, M.J.; Verkley, G.J.M.; Shin, H.D.; Nakashima, C.; Groenewald, J.Z. Phylogenetic lineages in Pseudocercospora. Stud. Mycol. 2013, 75, 37–114. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the Consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1–40. [Google Scholar] [CrossRef]
- Cheewangkoon, R.; Crous, P.W.; Hyde, K.D.; Groenewald, J.Z.; To-anan, C. Species of Mycosphaerell and related anamorphs on Eucalyptus leaves from Thailand. Persoonia 2008, 21, 77–91. [Google Scholar] [CrossRef]
- Liang, Z.C. Forest Diseases in China; China Forestry Publishing House: Beijing, China, 1984. [Google Scholar]
- Zhou, T.S. A preliminary study on pathogenic fungi of the leaf purple spot blotch of Eucalyptus spp. J. Southwest For. Coll. 1990, 10, 180–183. [Google Scholar]
- Wu, X.Y. Occurrence and control of forest diseases in Hainan Island. Trop. For. 2001, 29, 162–166. [Google Scholar]
- Luo, J.T.; Xue, Z.N.; Liao, W.J.; Yuan, C.L.; Ouyang, J.Y.; Shan, Y. Investigation on the diseases in fast growing Eucalyptus spp. in Guangxi. For. Pest Dis. 2012, 31, 21–24. [Google Scholar]
- Zhang, G.H.; Song, S.Y.; Liu, D.B.; Fan, C.M.; Hu, Z.M. Pathogen identification for brown spot disease and anthracnose of Eucalyptus robusta and its analysis. China Plant Prot. 2014, 34, 10–13. [Google Scholar]
- Yang, Y.J. Occurrence and control of main diseases and insect pests of Eucalyptus plantation in Hainan. Mod. Agric. Sci. Technol. 2015, 24, 134–138. [Google Scholar]
- Brooker, M.I.H.; Kleinig, D.A. Field Guide to Eucalypts Volume 2: South-Western and Southern Australia; Inkata Press: Melbourne, Australia, 1990. [Google Scholar]
- Potts, B.M.; Pederick, L.A. Morphology, phylogeny, origin, distribution and genetic diversity of eucalypts. In Diseases and Pathogens of Eucalypts; Keane, P.J., Kile, G.A., Podger, F.D., Brown, B.N., Eds.; CSIRO Publishing: Melbourne, Australia, 2000. [Google Scholar]
- Sankaran, K.V.; Sutton, B.C.; Minter, D. A Checklist of Fungi Recorded on Eucalyptus; CAB International: Oxon, UK, 1995. [Google Scholar]
- Andjic, V.; Carnegie, A.J.; Pegg, G.S.; Hardy, G.E.S.J.; Maxwell, A.; Crous, P.W.; Pérez, C.; Wingfield, M.J.; Burgess, T.I. 23 years of research on Teratosphaeria leaf blight of Eucalyptus. For. Ecol. Manag. 2019, 443, 19–27. [Google Scholar] [CrossRef]
- van Burik, J.A.H.; Schreckhise, R.W.; White, T.C.; Bowden, R.A.; Myerson, D. Comparison of six extraction techniques for isolation of DNA from filamentous fungi. Med. Mycol. 1998, 36, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Taole, M.M.; Burgess, T.I.; Gryzenhout, M.; Wingfield, B.D.; Wingfield, M.J. DNA sequence incongruence and inconsistent morphology obscure species boundaries in the Teratosphaeria suttonii species complex. Mycoscience 2012, 53, 270–283. [Google Scholar] [CrossRef]
- Solís, M.; Wingfield, M.J.; Hammerbacher, A.; Naidoo, S. Comparison of the infection biology of Teratosphaeria destructans and Teratosphaeria epicoccoides on Eucalyptus. Plant Dis. 2022, 106, 1944–1951. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Michael, I., Thomas, W.G., John, J.S., Eds.; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 31, 1323–1330. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetic analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Crous, P.W.; Boden, D. Kirramyces destructans sp. nov., a serious leaf pathogen of Eucalyptus in Indonesia. S. Afr. J. Bot. 1996, 62, 325–327. [Google Scholar] [CrossRef]
- Walker, J.; Sutton, B.C.; Pascoe, I.G. Phaeoseptoria eucalypti and similar fungi on Eucalyptus, with description of Kirramyces gen. nov. (Coelomycetes). Mycol. Res. 1992, 96, 911–924. [Google Scholar] [CrossRef]
- Greyling, I.; Wingfield, M.J.; Coetzee, M.P.; Marincowitz, S.; Roux, J. The Eucalyptus shoot and leaf pathogen Teratosphaeria destructans recorded in South Africa. South. For. 2016, 78, 123–129. [Google Scholar] [CrossRef]
- Park, R.F.; Keane, P.J. Three Mycosphaerella species from leaf diseases of Eucalyptus. Trans. Br. Mycol. Soc. 1982, 79, 95–100. [Google Scholar] [CrossRef]
- Park, R.F.; Keane, P.J. Leaf diseases of Eucalyptus associated with Mycosphaerella species. Trans. Br. Mycol. Soc. 1982, 79, 101–115. [Google Scholar] [CrossRef]
- Aptroot, A. Mycosphaerella and its anamorphs: 2. Conspectus of Mycosphaerella. CBS Biodivers. Ser. 2006, 5, 1–231. [Google Scholar]
- Crous, P.W.; Aptroot, A.; Kang, J.; Braun, U. The genus Mycosphaerella and its anamorphs. Stud. Mycol. 2000, 45, 107–121. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Mansilla, J.P.; Alfenas, A.C.; Groenewald, J.Z. Phylogenetic reassessment of Mycosphaerella spp. and their anamorphs occurring on Eucalyptus. II. Stud. Mycol. 2006, 55, 99–131. [Google Scholar] [CrossRef] [PubMed]
- Verkley, G.J.M.; Quaedvlieg, W.; Shin, H.; Crous, P.W. A new approach to species delimitation in Septoria. Stud. Mycol. 2013, 75, 213–305. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Braun, U.; Groenewald, J.Z. Mycosphaerella is polyphyletic. Stud. Mycol. 2007, 58, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Hunter, G.C.; Crous, P.W.; Wingfield, B.D.; Pongpanich, K.; Wingfield, M.J. Pseudocercospora flavomarginata sp. nov., from Eucalyptus leaves in Thailand. Fungal Divers. 2006, 22, 71–90. [Google Scholar]
- Heather, W.A. Some Aspects of the Ecology and Pathology of Phaeoseptoria eucalypti Hans. Emend. Walker on Some Species of the Genus Eucalyptus. Ph.D. Thesis, The Australian National University, Canberra, Australia, 1965. [Google Scholar]
- Wang, Q.C.; Liu, F.F.; Liu, Q.L.; Wu, W.X.; Wingfield, M.J.; Chen, S.F. Comparison of hyphal fragments and spores to evaluate the pathogenicity of the Eucalyptus leaf and shoot pathogen Calonectria pseudoreteaudii. Plant Dis. 2022, 106, 3145–3153. [Google Scholar] [CrossRef]
- Taole, M.; Bihon, W.; Wingfield, B.D.; Wingfield, M.J.; Burgess, T.I. Multiple introductions from multiple sources: Invasion patterns for an important Eucalyptus leaf pathogen. Ecol. Evol. 2015, 5, 4210–4220. [Google Scholar] [CrossRef]
- Havenga, M.; Wingfield, B.D.; Wingfield, M.J.; Dreyer, L.L.; Roets, F.; Aylward, J. Low genetic diversity and strong geographic structure in six introduced populations of the Eucalyptus foliar pathogen Teratosphaeria destructans. Plant Pathol. 2020, 69, 1540–1550. [Google Scholar] [CrossRef]
- Havenga, M.; Wingfield, B.D.; Wingfield, M.J.; Roets, F.; Dreyer, L.L.; Tatham, C.T.; Duong, D.A.; Wilken, P.M.; Chen, S.; Aylward, J. Mating strategy and mating type distribution in six global populations of the Eucalyptus foliar pathogen Teratosphaeria destructans. Fungal Genet. Biol. 2020, 137, 103350. [Google Scholar] [CrossRef]
- Havenga, M.; Wingfield, B.D.; Wingfield, M.J.; Marincowitz, S.; Dreyer, L.L.; Roets, F.; Japarudin, Y.; Aylward, J. Genetic recombination in Teratosphaeria destructans causing a new disease outbreak in Malaysia. For. Pathol. 2021, 51, e12683. [Google Scholar] [CrossRef]
- Andjic, V.; Dell, B.; Barber, P.; Hardy, G.E.S.; Wingfield, M.J.; Burgess, T.I. Plants for planting; indirect evidence for the movement of a serious forest pathogen, Teratosphaeria destructans, in Asia. Eur. J. Plant Pathol. 2011, 131, 49–58. [Google Scholar] [CrossRef]
- Solís, M.; Hammerbacher, A.; Wingfield, M.J.; Naidoo, S. A Robust Disease Scoring Method to Screen Eucalyptus for Resistance Against the Aggressive Leaf Pathogen Teratosphaeria destructans. Plant Dis. 2023, 107, 1418–1424. [Google Scholar] [CrossRef] [PubMed]



| Sampling Site No. a | Sample and Isolate Information b | Province | Location Details | GPS Information | Hosts | Collector | Identified Teratosphaeria spp. | Sequenced Isolate Number | Identified Isolate Number of T. epicoccoides | Identified Isolate Number of T. destructans |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 20230315-2 | Yunnan | Xin Village, Yongping Town, Jinggu County, Puer Region, Yunnan | 23°19′44.6052″ N, 100°21′40.014″ E | 2-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 14 | 7 | 7 |
| 2 | 20230315-1 | Yunnan | Hedong Village, Weiyuan Town, Jinggu County, Puer Region, Yunnan | 23°30′52.0056″ N, 100°40′52.4568″ E | 1-year-old E. urophylla | B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 16 | 2 | 14 |
| 3 | 20230314-2 | Yunnan | Baomu Village, Fengshan Town, Jinggu County, Puer Region, Yunnan | 23°43′47.6904″ N, 100°50′40.3008″ E | 2-year-old E. urophylla × E. tereticomis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 4 | 20230314-1 | Yunnan | Xin Village, Anban Town, Zhenyuan County, Puer Region, Yunnan | 23°53′15.0918″ N, 100°55′25.1508″ E | 3-year-old E. urophylla × E. pellita | B.Y. Chen and L.Q. Lu | T. destructans | 3 | 0 | 3 |
| 5 | 20230315-3 | Yunnan | Dawangtian Village, Ninger Town, Ninger County, Puer Region, Yunnan | 23°9′51.0228″ N, 100°59′48.1596″ E | 1-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 14 | 5 | 9 |
| 6 | 20230316-2 | Yunnan | Longjing Village, Gejiu County, Honghe Region, Yunnan | 23°25′4.7496″ N, 103°14′21.9012″ E | 2-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 7 | 20230317-1 | Yunnan | Chachang Village, Yangliujing Town, Guangnan County, Wenshan Region, Yunnan | 23°56′26.412″ N, 105°15′38.9916″ E | 2-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 8 | 20230318-1 | Yunnan | Administration center of Xinhua Town, Funing County, Wenshan Region, Yunnan | 23°36′43.11″ N, 105°37′7.1256″ E | 2-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 9 | 20230318-2 | Yunnan | Podi Village, Xinhua Town, Funing County, Wenshan Region, Yunnan | 23°34′57.3564″ N, 105°34′34.7016″ E | 1-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 10 | 20230318-3 | Yunnan | Nahong Village, Zhesang Town, Funing County, Wenshan Region, Yunnan | 23°44′40.1316″ N, 105°57′12.204″ E | 1-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 11 | 20230319-1 | Yunnan | Zhening Village, Boai Town, Funing County, Wenshan Region, Yunnan | 23°49′53.4504″ N, 106°1′1.4196″ E | 2-year-old E. urophylla × E. tereticomis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 5 | 5 | 0 |
| 12 | 20220703-1 | Guangxi | Xichang Village, Liuqiao Town, Fusui County, Chongzuo Region, Guangxi | 22°17′59.0496″ N, 107°43′39.936″ E | 2-year-old E. urophylla × E. grandis | S.F. Chen, W.X. Wu, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 12 | 7 | 5 |
| 13 | 20220703-3 | Guangxi | Jiucheng Village, Dongmen Town, Fusui County, Chongzuo Region, Guangxi | 22°19′18.4152″ N, 107°47′24.9792″ E | 0.5-year-old E. urophylla × E. grandis | S.F. Chen, W.X. Wu, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 15 | 7 | 8 |
| 14 | 20220703-2 | Guangxi | Namengxin Village, Dongmen Town, Fusui County, Chongzuo Region, Guangxi | 22°20′39.4476″ N, 107°52′20.9568″ E | 2-year-old E. urophylla × E. grandis | S.F. Chen, W.X. Wu, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 13 | 6 | 7 |
| 15 | 20220703-4 | Guangxi | Chongwen Village, Dazhi Town, Qinbei District, Qinzhou Region, Guangxi | 22°1′52.8816″ N, 108°21′27.9576″ E | 0.5-year-old E. urophylla × E. grandis | S.F. Chen, W.X. Wu, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 10 | 2 | 8 |
| 16 | 20220702-3 | Guangxi | Youxing Village, Dongchang Town, Qinnan District, Qinzhou Region, Guangxi | 21°53′7.4832″ N, 108°46′15.3516″ E | 0.5-year-old E. urophylla × E. grandis | S.F. Chen, W.X. Wu, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 14 | 3 | 11 |
| 17 | 20220702-2 | Guangxi | Bengtang Village, Nali Town, Qinnan District, Qinzhou Region, Guangxi | 21°51′37.368″ N, 108°55′57.648″ E | 1-year-old E. urophylla × E. grandis | S.F. Chen, W.X. Wu, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 14 | 5 | 9 |
| 18 | 20220702-1 | Guangxi | Dama Village, Nali Town, Qinnan District, Qinzhou Region, Guangxi | 21°50′39.5124″ N, 108°55′55.3404″ E | 0.5-year-old E. urophylla × E. pellita | S.F. Chen, W.X. Wu, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 14 | 8 | 6 |
| 19 | 20220704-1 | Guangxi | Chezouling Village, Fucheng Town, Yinhai District, Beihai Region, Guangxi | 21°38′8.7252″ N, 109°17′51.4644″ E | 0.5-year-old E. urophylla × E. grandis | S.F. Chen, W.X. Wu, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 15 | 5 | 10 |
| 20 | 20220704-2 | Guangxi | Wuliu Village, Shankou Town, Hepu County, Beihai Region, Guangxi | 21°33′10.3248″ N, 109°42′43.2324″ E | 1-year-old E. urophylla × E. pellita | S.F. Chen, W.X. Wu, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 20 | 6 | 14 |
| 21 | 20220704-3 | Guangxi | Gaopo Village, Shankou Town, Hepu County, Beihai Region, Guangxi | 21°34′3.9792″ N, 109°43′8.6304″ E | 2-year-old E. urophylla | S.F. Chen, W.X. Wu, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 16 | 6 | 10 |
| 22 | 20221026-6 | Hainan | Administration center of Dongying Town, Lingao County, Hainan | 19°57′58.788″ N, 109°38′43.206″ E | 2-year-old E. urophylla × E. grandis | S.F. Chen, Y. Liu, X.Y. Liang, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 3 | 3 | 0 |
| 23 | 20221026-7 | Hainan | Wenshan Village, Dongying Town, Lingao County, Hainan | 19°58′36.4656″ N, 109°38′53.6352″ E | 1-year-old E. urophylla × E. grandis | S.F. Chen, Y. Liu, X.Y. Liang, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 17 | 6 | 11 |
| 24 | 20221026-4 | Hainan | Meisheng Village, Lincheng Town, Lingao County, Hainan | 19°46′16.644″ N, 109°41′31.74″ E | 2-year-old E. urophylla × E. grandis | S.F. Chen, Y. Liu, X.Y. Liang, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 10 | 6 | 4 |
| 25 | 20221026-5 | Hainan | Lanluo Village, Lincheng Town, Lingao County, Hainan | 19°47′49.0452″ N, 109°41′8.7324″ E | 2 to 3-year-old E. urophylla × E. tereticomis | S.F. Chen, Y. Liu, X.Y. Liang, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 15 | 5 | 10 |
| 26 | 20221026-2 | Hainan | Dunxiang Village, Jialai Town, Lingao County, Hainan | 19°43′34.8276″ N, 109°42′52.1712″ E | 2-year-old E. urophylla × E. grandis | S.F. Chen, Y. Liu, X.Y. Liang, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 9 | 6 | 3 |
| 27 | 20221026-3 | Hainan | Duolang Village, Duowen Town, Lingao County, Hainan | 19°45′45.1548″ N, 109°44′28.518″ E | 2-year-old E. urophylla × E. tereticomis | S.F. Chen, Y. Liu, X.Y. Liang, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. destructans | 7 | 0 | 7 |
| 28 | 20221026-1 | Hainan | Songbai Village, Huangtong Town, Lingao County, Hainan | 19°47′41.46″ N, 109°49′52.7196″ E | 1-year-old E. urophylla × E. grandis | S.F. Chen, Y. Liu, X.Y. Liang, S.M. Xiang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 5 | 5 | 0 |
| 29 | 20230221-2 | Guangdong | Fanchang Village, Longmen Town, Leizhou County, Zhanjiang Region, Guangdong | 20°40′3.018″ N, 110°1′50.358″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 18 | 6 | 12 |
| 30 | 20230221-1 | Guangdong | Naqi Village, Chengbei Town, Xuwen County, Zhanjiang Region, Guangdong | 20°20′49.56″ N, 110°9′58.5864″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 19 | 6 | 13 |
| 31 | 20230220-3 | Guangdong | Houkeng Village, Chengyue Town, Leizhou County, Zhanjiang Region, Guangdong | 21°7′53.886″ N, 110°6′4.932″ E | 2-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 18 | 6 | 12 |
| 32 | 20230221-3 | Guangdong | Lizhikeng Village, Shicheng Town, Lianjiang County, Zhanjiang Region, Guangdong | 21°31′37.7292″ N, 110°16′12.648″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 19 | 6 | 13 |
| 33 | 20230220-2 | Guangdong | Xiachen Village, Rudong Town, Yangxi County, Yangjiang Region, Guangdong | 21°36′43.29″ N, 111°26′23.118″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 14 | 3 | 11 |
| 34 | 20230220-1 | Guangdong | Li Village, Chengcun Town, Yangxi County, Yangjiang Region, Guangdong | 21°49′47.5284″ N, 111°43′3.8964″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 10 | 2 | 8 |
| 35 | 20230216-5 | Guangdong | Xiadaidong Village, Huilong Town, Deqing County, Zhaoqing Region, Guangdong | 23°11′35.2032″ N, 111°41′21.1596″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 36 | 20230216-3 | Guangdong | Daxintang Village, Helang Town, Yangchun County, Yangjiang Region, Guangdong | 22°34′14.4336″ N, 111°51′53.928″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 37 | 20230216-4 | Guangdong | Xin Village, Helang Town, Yangchun County, Yangjiang Region, Guangdong | 22°33′32.3748″ N, 111°51′22.4568″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 4 | 4 | 0 |
| 38 | 20230216-2 | Guangdong | Tangxia Village, Songbai Town, Yangchun County, Yangjiang Region, Guangdong | 22°27′23.958″ N, 111°53′30.9192″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 39 | 20230215-1 | Guangdong | Xin Village, Shiwang Town, Yangchun County, Yangjiang Region, Guangdong | 22°33′2.6928″ N, 111°57′34.7616″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 40 | 20230216-1 | Guangdong | Shuizhai Village, Chunwan Town, Yangchun County, Yangjiang Region, Guangdong | 22°25′8.1876″ N, 111°56′20.4252″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 41 | 20230217-3 | Guangdong | Fengji Village, Lixi Town, Yingde County, Qingyuan Region, Guangdong | 23°55′21.4824″ N, 113°10′20.4816″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides and T. destructans | 18 | 7 | 11 |
| 42 | 20230219-1 | Guangdong | Kengwei Village, Feilaixia Town, Qingcheng District, Qingyuan Region, Guangdong | 23°53′2.004″ N, 113°10′4.2456″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. destructans | 10 | 0 | 10 |
| 43 | 20230217-2 | Guangdong | Xikeng Village, Feilaixia Town, Qingcheng District, Qingyuan Region, Guangdong | 23°50′40.7112″ N, 113°9′38.2932″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 44 | 20230217-1 | Guangdong | Donger Village, Dongcheng Subdistrict, Qingcheng District, Qingyuan Region, Guangdong | 23°45′27.1476″ N, 113°8′16.656″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 45 | 20230218-1 | Guangdong | Huangwuxin Village, Maba Town, Qujiang District, Shaoguan Region, Guangdong | 24°37′49.3284″ N, 113°35′10.8384″ E | 2-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 46 | 20230217-4 | Guangdong | Heping Village, Wangbu Town, Yingde County, Qingyuan Region, Guangdong | 24°11′49.0884″ N, 113°28′19.1424″ E | 2-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 47 | 20230218-2 | Guangdong | Xinzouwu Village, Wushi Town, Qujiang District, Shaoguan Region, Guangdong | 24°36′12.618″ N, 113°38′22.0128″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 48 | 20230218-3 | Guangdong | Yiwu Village, Datang Town, Qujiang District, Shaoguan Region, Guangdong | 24°43′37.956″ N, 113°42′49.9536″ E | 1-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 49 | 20230218-4 | Guangdong | Xin Village, Daqiao Town, Renhua County, Shaoguan Region, Guangdong | 24°55′28.0596″ N, 113°46′30.2232″ E | 2-year-old E. urophylla × E. grandis | W.X. Wu, X.Y. Liang, B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 50 | 20230330-1 | Fujian | Qing Village, Ansha Town, Yongan County, Sanming Region, Fujian | 25°59′56.4864″ N, 117°6′47.772″ E | 2-year-old E. urophylla × E. pellita | B.Y. Chen and L.Q. Lu | T. epicoccoides | 5 | 5 | 0 |
| 51 | 20230328-1 | Fujian | Zhanglin Village, Caoyuan Town, Yongan County, Sanming Region, Fujian | 26°1′34.122″ N, 117°18′6.5556″ E | 2-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 52 | 20230329-1 | Fujian | Linggan Village, Dahu Town, Yongan County, Sanming Region, Fujian | 26°4′45.6024″ N, 117°19′15.186″ E | 7-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 53 | 20230329-2 | Fujian | Wenbeixin Village, Caoyuan Town, Yongan County, Sanming Region, Fujian | 26°0′45.684″ N, 117°18′44.964″ E | 1-year-old E. urophylla × E. pellita | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 54 | 20230331-3 | Fujian | Hekeng Village, Guantian Town, Zhangping County, Longyan Region, Fujian | 25°1′7.4604″ N, 117°29′14.9388″ E | 2-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 55 | 20230331-2 | Fujian | Xipo Village, Hulin Town, Huaan County, Zhangzhou Region, Fujian | 25°7′1.6356″ N, 117°32′59.4276″ E | 1-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 56 | 20230331-1 | Fujian | Shangxue Village, Huafeng Town, Huaan County, Zhangzhou Region, Fujian | 24°57′58.5504″ N, 117°34′0.5592″ E | 3-year-old E. urophylla × E. pellita | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 57 | 20230330-3 | Fujian | Dakeng Village, Shajian Town, Huaan County, Zhangzhou Region, Fujian | 24°45′37.134″ N, 117°35′51.2052″ E | 1-year-old E. urophylla × E. grandis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 58 | 20230330-2 | Fujian | Xiawenkeng Village, Yanxi Town, Changtai District, Zhangzhou Region, Fujian | 24°45′43.3476″ N, 117°48′4.986″ E | 2-year-old E. urophylla × E. tereticomis | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| 59 | 20230401-1 | Fujian | Dudong Village, Changtai District, Zhangzhou Region, Fujian | 24°37′52.3956″ N, 117°42′12.3624″ E | 2-year-old E. urophylla × E. pellita | B.Y. Chen and L.Q. Lu | T. epicoccoides | 6 | 6 | 0 |
| Species | Isolate No. a | Type | Hosts | Location | Collector | GenBank Accession No. b | References or Source of Data | ||
|---|---|---|---|---|---|---|---|---|---|
| ITS | tef1 | tub2 | |||||||
| Teratosphaeria aurantia | CBS 125243 = MUCC 668 | ex-type | Eucalyptus grandis | Queensland, Australia | G. Whyte | KF901561 | N/A c | KF902984 | [30] |
| T. biformis | CBS 124578 = MUCC 693 | ex-type | Eucalyptus globulus | Queensland, Australia | G. Whyte | KF901564 | KF903287 | KF902987 | [30] |
| T. blakelyi | CBS 120089 = CPC 12837 | ex-type | Eucalyptus blakelyi | New South Wales, Australia | B.A. Summerell | KF901565 | KF903288 | KF902988 | [30] |
| T. callophyllae | CBS 124584 = MUCC 700 | ex-type | Corymbia calophylla | Western Australia, Australia | K. Taylor | KF901566 | KF903289 | FJ532003 | [23,30] |
| T. considenianae | CBS 120087 = CPC 12940 | ex-type | Eucalyptus consideniana | New South Wales, Australia | B.A. Summerell | DQ923527 | N/A | FJ952510 | [23,30] |
| CPC 13032 | unknown | Eucalyptus sp. | New South Wales, Australia | B.A. Summerell | KF901567 | KF903291 | KF902990 | [30] | |
| T. corymbiae | CBS 120495 = DAR 77445 | ex-type | Corymbia maculata | New South Wales, Australia | A.J. Carnegie | EF011657 | N/A | FJ532005 | [23] |
| CBS 124988 = CPC 13125 | unknown | Corymbia henryi | New South Wales, Australia | A.J. Carnegie | KF901569 | KF903293 | KF902992 | [30] | |
| T. crispata | CBS 130523 | ex-type | Eucalyptus bridgesiana | New South Wales, Australia | A.J. Carnegie | GQ890348 | N/A | N/A | [23,30] |
| T. cryptica | CBS 145895 = CPC 13839 | ex-epitype | Eucalyptus globulus | Australia | I.W. Smith | KF901572 | KF903298 | KF902997 | [23,30] |
| T. destructans | CBS 111369 = CMW 5219 = CPC 1366 | ex-type | Eucalyptus grandis | Indonesia | M.J. Wingfield | DQ267595 | DQ235113 | N/A | [23] |
| CBS 111370 = CPC 1368 | ex-type | Eucalyptus grandis | Indonesia | M.J. Wingfield | KF901574 | KF903301 | KF903000 | [23,30] | |
| T. epicoccoides | CMW 5348 = CPC 1346 | ex-type | Eucalyptus sp. | Indonesia | M.J. Wingfield | DQ239972 | DQ240170 | DQ240117 | [23] |
| CBS 119973 | unknown | Eucalyptus pellita | Vietnam | T.I. Burgess | KF901784 | KF903359 | KF903055 | [30] | |
| CPC 12218 | unknown | Eucalyptus sp. | Indonesia | M.J. Wingfield | KF901664 | KF903357 | KF903054 | [30] | |
| T. eucalypti | CPC 12552 | not type | Eucalyptus nitens | Tasmania, Australia | C. Mohammed | KF901576 | KF903303 | KF903002 | [30] |
| MUCC 623 | not type | Eucalyptus nitens | New South Wales, Australia | A.J. Carnegie | FJ793255 | EU101623 | EU101566 | [25] | |
| T. foliensis | CBS 124581 = MUCC 670 | ex-type | Eucalyptus globulus | New South Wales, Australia | S. Collins | KF901580 | KF903311 | KF903009 | [30] |
| T. gauchensis | CBS 120303 = CMW 17331 | ex-type | Eucalyptus grandis | Uruguay | M.J. Wingfield | KF901790 | KF903315 | KF903013 | [30] |
| CBS 120304 = CMW 17332 | ex-type | Eucalyptus grandis | Uruguay | M.J. Wingfield | KF901789 | KF903314 | KF903012 | [30] | |
| T. juvenalis | CBS 110906 = CMW 13347 | ex-type | Eucalyptus cladocalyx | South Africa | P.W. Crous | KF901730 | N/A | N/A | [30] |
| CBS 111149 | unknown | Eucalyptus cladocalyx | South Africa | P.W. Crous | KF901670 | KF903317 | KF903015 | [30] | |
| T. majorizuluensis | CBS 120040 = CPC 12712 | ex-type | Eucalyptus botryoides | New South Wales, Australia | B.A. Summerell | KF901581 | KF903319 | KF903017 | [30] |
| T. molleriana | CBS 111164 = CMW 4940 = CPC 1214 | ex-type | Eucalyptus globulus | Portugal | S. McCrae | KF901692 | KF903324 | KF903021 | [30] |
| CBS 111165 = CPC 1215 | ex-type | Eucalyptus globulus | Portugal | S. McCrae | KF901693 | KF903325 | KF903022 | [30] | |
| T. novaehollandiae | BRIP 59486 | ex-type | Eucalyptus camaldulensis | Western Australia, Australia | A. Maxwell | KT972281 | KT972345 | KT972313 | [25] |
| BRIP 59487 | not type | Eucalyptus camaldulensis | Western Australia, Australia | A. Maxwell | KT972282 | KT972346 | KT972314 | [25] | |
| T. nubilosa | CBS 116005 = CMW 3282 = CPC 937 | ex-epitype | Eucalyptus globulus | Victoria, Australia | A.J. Carnegie | KF901686 | KF903336 | KF903033 | [23,30] |
| CPC 13844 | not type | Eucalyptus globulus | Australia | I.W. Smith | KF901590 | KF903344 | KF903041 | [30] | |
| T. ovata | CBS 124052 = CPC 14632 | unknown | Eucalyptus phoenicea | New South Wales, Australia | B.A. Summerell | KF901591 | KF903345 | KF903042 | [30] |
| T. pseudoeucalypti | CBS 124577 = MUCC 607 | ex-type | Eucalyptus grandis × E. camaldulensis | Queensland, Australia | G.S. Pegg | KF901593 | KF903349 | KF903046 | [30] |
| T. pseudonubilosa | CBS 135621 = CMW 30745 | ex-type | Eucalyptus globulus | Victoria, Australia | G. Pérez | HQ130818 | N/A | HQ131318 | [23] |
| CBS 135620 = CMW 30723 | link to paratype | Eucalyptus globulus | Western Australia, Australia | G. Pérez | HQ130817 | N/A | HQ131317 | [23] | |
| T. rubidae | CBS 124579 = MUCC 658 | ex-type | Corymbia calophylla | Western Australia, Australia | P.A. Barber | KF901596 | KF903352 | KF903049 | [30] |
| T. stellenboschiana | CBS 124989 = CPC 13767 | unknown | Eucalyptus punctata | South Africa | P.W. Crous and G. Bills | KF901732 | KF903355 | KF903052 | [30] |
| T. syncarpiae | CBS 121160 = DAR 77433 | ex-type | Syncarpia glomulifera | New South Wales, Australia | A.J. Carnegie and M.J. Wingfield | KF901598 | KF903360 | KF903056 | [30] |
| T. tiwiana | CBS 141549 = BRIP 63496 | ex-type | Eucalyptus urophylla hybrids | Tiwi Island, Australia | T.I. Burgess | KT972297 | KT972362 | KT972330 | [25] |
| CBS 141550 = BRIP 63497 | not type | Eucalyptus urophylla hybrids | Tiwi Island, Australia | T.I. Burgess | KT972305 | KT972369 | KT972337 | [25] | |
| T. toledana | CBS 113313 = CMW 14457 | ex-type | Eucalyptus sp. | Spain | P.W. Crous | KF901734 | KF903361 | KF903058 | [30] |
| CBS 115513 = CPC 10840 | unknown | Eucalyptus sp. | Spain | P.W. Crous and G. Bills | KF901600 | KF903362 | KF903059 | [30] | |
| T. verrucosa | CBS 113621 = CPC 42 | ex-type | Eucalyptus cladocalyx | South Africa | P.W. Crous | KF901645 | KF903365 | KF903062 | [30] |
| CPC 12949 | unknown | Eucalyptus sp. | South Africa | P.W. Crous | KF901768 | KF903364 | KF903061 | [30] | |
| T. viscida | CBS 121157 = MUCC 453 | ex-type | Eucalyptus grandis | Mareeba, Australia | T.I. Burgess | EF031472 | EF031496 | EF031484 | [25] |
| CBS 121156 = MUCC 452 | not type | Eucalyptus grandis | Mareeba, Australia | T.I. Burgess | EF031471 | EF031495 | EF031483 | [25] | |
| T. zuluensis | CBS 120301 = CMW 17321 | ex-epitype | Eucalyptus grandis | South Africa | M.J. Wingfield | KF901735 | KF903368 | KF903064 | [30] |
| CBS 120302 = CMW 17322 | ex-epitype | Eucalyptus grandis | South Africa | M.J. Wingfield | KF901736 | KF903369 | KF903065 | [30] | |
| Staninwardia suttonii | CBS 120061 = CPC 13055 | ex-type | Eucalyptus robusta | Australia | B.A. Summerell | KF901552 | KF903270 | KF902974 | [30] |
| Genotype a | Yunnan | Guangxi | Guangdong | Hainan | Fujian | Five Provinces |
|---|---|---|---|---|---|---|
| AAA | 19 | 5 | 21 | 3 | 9 | 57 |
| ABA | 1 | 2 | 0 | 0 | 0 | 3 |
| ACA | 5 | 13 | 18 | 4 | 4 | 44 |
| AEA | 0 | 1 | 0 | 3 | 0 | 4 |
| AFA | 0 | 1 | 0 | 2 | 0 | 3 |
| AHA | 0 | 1 | 0 | 0 | 0 | 1 |
| AIA | 0 | 0 | 1 | 2 | 0 | 3 |
| BAA | 0 | 1 | 4 | 1 | 2 | 8 |
| BCA | 0 | 0 | 1 | 0 | 0 | 1 |
| CAA | 3 | 2 | 7 | 2 | 5 | 19 |
| CBA | 0 | 1 | 0 | 0 | 0 | 1 |
| CCA | 0 | 1 | 2 | 0 | 3 | 6 |
| DEA | 0 | 0 | 0 | 1 | 0 | 1 |
| DHA | 0 | 1 | 0 | 0 | 0 | 1 |
| DIA | 6 | 1 | 0 | 2 | 9 | 18 |
| DJA | 0 | 2 | 0 | 0 | 0 | 2 |
| EAA | 16 | 11 | 35 | 3 | 17 | 82 |
| ECA | 4 | 6 | 14 | 2 | 7 | 33 |
| FCA | 1 | 0 | 0 | 0 | 0 | 1 |
| GGA | 0 | 1 | 0 | 1 | 0 | 2 |
| HAA | 0 | 0 | 1 | 0 | 0 | 1 |
| Number of Genotype | 8 | 16 | 10 | 12 | 8 | 21 |
| Number of Isolate | 55 | 50 | 104 | 26 | 56 | 291 |
| Ratio of Number of Genotype to Number of Isolate | 0.15 | 0.32 | 0.096 | 0.46 | 0.14 | 0.072 |
| Genotype a | Yunnan | Guangxi | Guangdong | Hainan | Fujian | Five Provinces |
|---|---|---|---|---|---|---|
| AAA | 32 | 37 | 49 | 14 | 0 | 132 |
| ACA | 0 | 1 | 5 | 4 | 0 | 10 |
| BAA | 1 | 0 | 0 | 0 | 0 | 1 |
| CAA | 0 | 45 | 27 | 15 | 0 | 87 |
| CBA | 0 | 1 | 0 | 0 | 0 | 1 |
| CCA | 0 | 1 | 9 | 1 | 0 | 11 |
| Number of Genotype | 2 | 5 | 4 | 4 | 0 | 6 |
| Number of Isolate | 33 | 85 | 90 | 34 | 0 | 242 |
| Ratio of Number of Genotype to Number of Isolate | 0.061 | 0.059 | 0.044 | 0.12 | N/A b | 0.025 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, B.; Wu, W.; Chen, S. Wide Distribution of Teratosphaeria epicoccoides and T. destructans Associated with Diseased Eucalyptus Leaves in Plantations in Southern China. Microorganisms 2024, 12, 129. https://doi.org/10.3390/microorganisms12010129
Chen B, Wu W, Chen S. Wide Distribution of Teratosphaeria epicoccoides and T. destructans Associated with Diseased Eucalyptus Leaves in Plantations in Southern China. Microorganisms. 2024; 12(1):129. https://doi.org/10.3390/microorganisms12010129
Chicago/Turabian StyleChen, Bingyin, Wenxia Wu, and Shuaifei Chen. 2024. "Wide Distribution of Teratosphaeria epicoccoides and T. destructans Associated with Diseased Eucalyptus Leaves in Plantations in Southern China" Microorganisms 12, no. 1: 129. https://doi.org/10.3390/microorganisms12010129
APA StyleChen, B., Wu, W., & Chen, S. (2024). Wide Distribution of Teratosphaeria epicoccoides and T. destructans Associated with Diseased Eucalyptus Leaves in Plantations in Southern China. Microorganisms, 12(1), 129. https://doi.org/10.3390/microorganisms12010129