



Enzymatic Cleavage of Stx2a in the Gut and Identification of Pancreatic Elastase and Trypsin as Possible Main Cleavers
 , , , ,
, , , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Stx2a Production and Purification
2.2. Sampling and Fractioning of Stool
2.3. Cleavage of Stx2a A Subunit by Trypsin or Furin
2.4. Incubation of Stx2a with Human Stool Supernatant or Its Fractions with or without Protease Inhibitors
2.5. Incubation of Stx2a with Recombinant CELA3B
2.6. Determination of the Structure of Stx2a A Subunit by Immunoblotting
2.7. Identification of the Cleavage Position of the A Subunit of Stx2a by Components Present in Human Stool
2.8. Detection of Chymotrypsin-like Elastase 3B (CELA3B) by Immunoblotting
3. Results
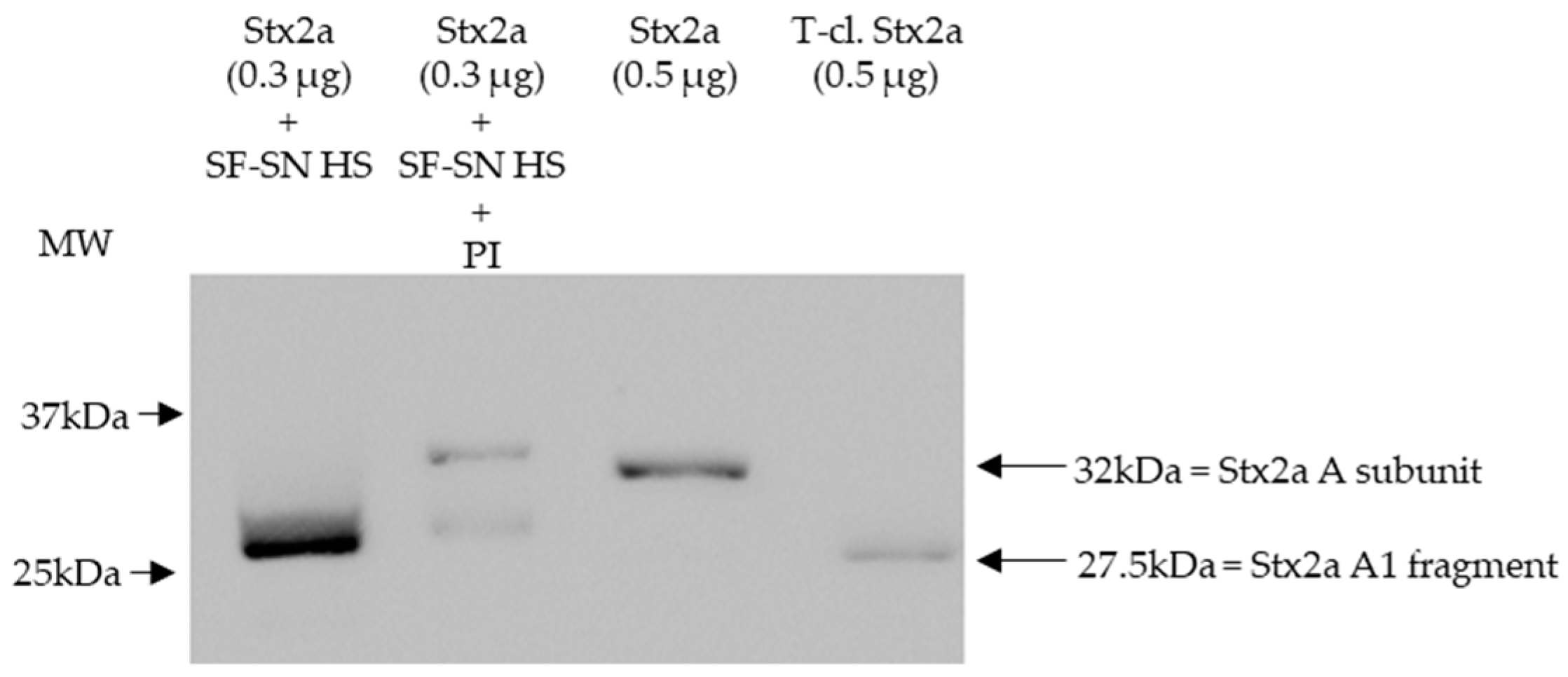
3.1. Enzymes Present in Human Stool Cleave the A Subunit of Stx2a In Vitro
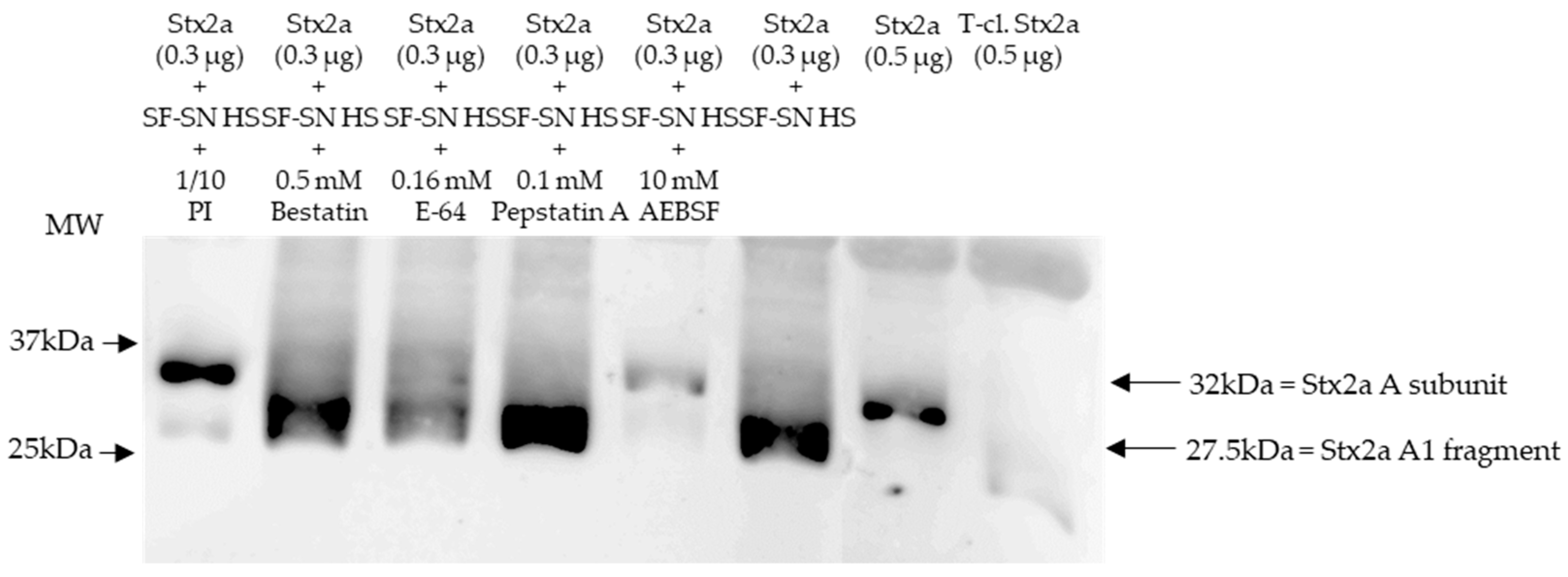
3.2. Serine Protease(s) in Human Stool Are Responsible for the Cleavage of the A Subunit of Stx2a
3.3. Localization of the Cleavage Position of the A subunit of Stx2a Induced by Different Purified Enzymes or Components Present in Stool from Healthy Human Donors
3.4. Chymotrypsin-like Elastase 3B (CELA3B) Induces Partial Cleavage of A Subunit of Stx2a In Vitro
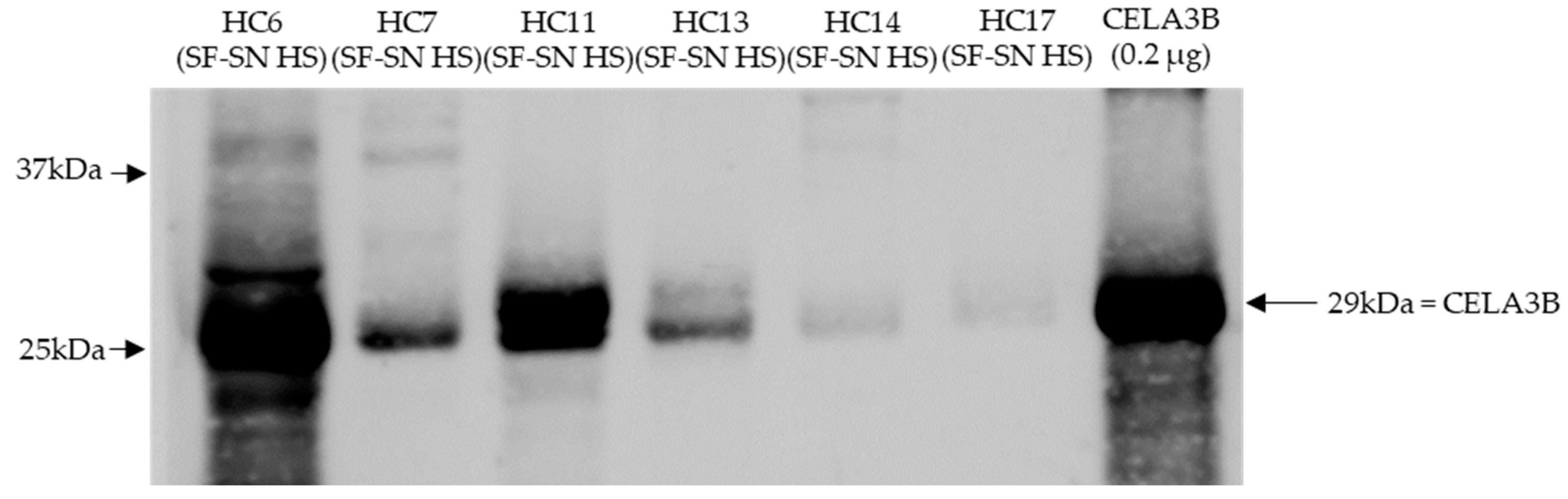
3.5. Stool of Healthy Donors Appear to Contain CELA3B
3.6. Stool from EHEC-Infected Patients Show a Similar Stx2a Cleavage Pattern
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tarr, P.I.; Gordon, C.A.; Chandler, W.L. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 2005, 365, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Rosales, A.; Hofer, J.; Zimmerhackl, L.-B.; Jungraithmayr, T.C.; Riedl, M.; Giner, T.; Strasak, A.; Orth-Höller, D.; Würzner, R.; Karch, H. Need for long-term follow-up in enterohemorrhagic Escherichia coli-associated hemolytic uremic syndrome due to late-emerging sequelae. Clin. Infect. Dis. 2012, 54, 1413–1421. [Google Scholar] [CrossRef] [PubMed]
- Karch, H.; Tarr, P.I.; Bielaszewska, M. Enterohaemorrhagic Escherichia coli in human medicine. Int. J. Med. Microbiol. 2005, 295, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Russo, L.M.; Melton-Celsa, A.R.; Smith, M.J.; O’Brien, A.D. Comparisons of Native Shiga Toxins (Stxs) Type 1 and 2 with Chimeric Toxins Indicate that the Source of the Binding Subunit Dictates Degree of Toxicity. PLoS ONE 2014, 9, e93463. [Google Scholar] [CrossRef]
- Gould, L.H.; Bopp, C.; Strockbine, N.; Atkinson, R.; Baselski, V.; Body, B.; Carey, R.; Crandall, C.; Hurd, S.; Kaplan, R.; et al. Recommendations for diagnosis of shiga toxin--producing Escherichia coli infections by clinical laboratories. MMWR. Recomm. Rep. Morb. Mortal. Wkly. Rep. Recomm. Rep. 2009, 58, 1–14. [Google Scholar] [CrossRef]
- Wijnsma, K.L.; van Bommel, S.A.M.; van der Velden, T.; Volokhina, E.; Schreuder, M.F.; van den Heuvel, L.P.; van de Kar, N.C.A.J. Fecal diagnostics in combination with serology: Best test to establish STEC-HUS. Pediatr. Nephrol. 2016, 31, 2163. [Google Scholar] [CrossRef]
- Tarr, P.I.; Neill, M.A.; Clausen, C.R.; Watkins, S.L.; Christie, D.L.; Hickman, R.O. Escherichia coli 0157:H7 and the Hemolytic Uremic Syndrome: Importance of Early Cultures in Establishing the Etiology. J. Infect. Dis. 1990, 162, 553–556. [Google Scholar] [CrossRef]
- Gerritzen, A.; Wittke, J.-W.; Wolff, D. Rapid and sensitive detection of Shiga toxin-producing Escherichia coli directly from stool samples by real-time PCR in comparison to culture, enzyme immunoassay and Vero cell cytotoxicity assay. Clin. Lab. 2011, 57, 993–998. [Google Scholar]
- Hurley, B.P.; Jacewicz, M.; Thorpe, C.M.; Lincicome, L.L.; King, A.J.; Keusch, G.T.; Acheson, D.W.K. Shiga toxins 1 and 2 translocate differently across polarized intestinal epithelial cells. Infect. Immun. 1999, 67, 6670–6677. [Google Scholar] [CrossRef]
- Müthing, J.; Meisen, I.; Zhang, W.; Bielaszewska, M.; Mormann, M.; Bauerfeind, R.; Schmidt, M.A.; Friedrich, A.W.; Karch, H. Promiscuous Shiga toxin 2e and its intimate relationship to Forssman. Glycobiology 2012, 22, 849–862. [Google Scholar] [CrossRef]
- Ergonul, Z.; Hughes, A.K.; Kohan, D.E. Induction of apoptosis of human brain microvascular endothelial cells by shiga toxin 1. J. Infect. Dis. 2003, 187, 154–158. [Google Scholar] [CrossRef]
- Zumbrun, S.D.; Hanson, L.; Sinclair, J.F.; Freedy, J.; Melton-Celsa, A.R.; Rodriguez-Canales, J.; Hanson, J.C.; O’Brien, A.D. Human intestinal tissue and cultured colonic cells contain globotriaosylceramide synthase mRNA and the alternate Shiga toxin receptor globotetraosylceramide. Infect. Immun. 2010, 78, 4488–4499. [Google Scholar] [CrossRef] [PubMed]
- Legros, N.; Pohlentz, G.; Steil, D.; Müthing, J. Shiga toxin-glycosphingolipid interaction: Status quo of research with focus on primary human brain and kidney endothelial cells. Int. J. Med. Microbiol. 2018, 308, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Lentz, E.K.; Leyva-Illades, D.; Lee, M.S.; Cherla, R.P.; Tesh, V.L. Differential response of the human renal proximal tubular epithelial cell line HK-2 to shiga toxin types 1 and 2. Infect. Immun. 2011, 79, 3527–3540. [Google Scholar] [CrossRef]
- Orth, D.; Grif, K.; Khan, A.B.; Naim, A.; Dierich, M.P.; Würzner, R. The Shiga toxin genotype rather than the amount of Shiga toxin or the cytotoxicity of Shiga toxin in vitro correlates with the appearance of the hemolytic uremic syndrome. Diagn. Microbiol. Infect. Dis. 2007, 59, 235–242. [Google Scholar] [CrossRef]
- Ho, N.K.; Henry, A.C.; Johnson-Henry, K.; Sherman, P.M. Pathogenicity, Host Responses and Implications for Management of Enterohemorrhagic Escherichia coli O157:H7 Infection. Can. J. Gastroenterol. Hepatol. 2013, 27, 281–285. [Google Scholar] [CrossRef]
- O’Brien, A.D.; Holmes, R.K. Shiga and Shiga-like toxins. Microbiol. Rev. 1987, 51, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Olsnes, S.; Reisbig, R.; Eiklid, K. Subunit structure of Shigella cytotoxin. J. Biol. Chem. 1981, 256, 8732–8738. [Google Scholar] [CrossRef] [PubMed]
- Garred, O.; Van Deurs, B.; Sandvig, K. Furin-induced Cleavage and Activation of Shiga Toxin. J. Biol. Chem. 1995, 270, 10817–10821. [Google Scholar] [CrossRef]
- Melton-Celsa, A.R.; Darnell, S.C.; O’Brien, A.D. Activation of Shiga-like toxins by mouse and human intestinal mucus correlates with virulence of enterohemorrhagic Escherichia coli O91:H21 isolates in orally infected, streptomycin-treated mice. Infect. Immun. 1996, 64, 1569–1576. [Google Scholar] [CrossRef]
- Garred, O.; Dubinina, E.; Holm, P.K.; Olsnes, S.; van Deurs, B.; Kozlov, J.V.; Sandvig, K. Role of processing and intracellular transport for optimal toxicity of Shiga toxin and toxin mutants. Exp. Cell Res. 1995, 218, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Kokai-Kun, J.F.; Melton-Celsa, A.R.; O’Brien, A.D. Elastase in Intestinal Mucus Enhances the Cytotoxicity of Shiga Toxin Type 2d. J. Biol. Chem. 2000, 275, 3713–3721. [Google Scholar] [CrossRef] [PubMed]
- Melton-Celsa, A.R.; Kokai-Kun, J.F.; O’Brien, A.D. Activation of Shiga toxin type 2d (Stx2d) by elastase involves cleavage of the C-terminal two amino acids of the A2 peptide in the context of the appropriate B pentamer. Mol. Microbiol. 2002, 43, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Brigotti, M.; Orth-Höller, D.; Carnicelli, D.; Porcellini, E.; Galassi, E.; Tazzari, P.L.; Ricci, F.; Manoli, F.; Manet, I.; Talasz, H.; et al. The structure of the Shiga toxin 2a A-subunit dictates the interactions of the toxin with blood components. Cell. Microbiol. 2019, 21, e13000. [Google Scholar] [CrossRef] [PubMed]
- Orth, D.; Khan, A.B.; Naim, A.; Grif, K.; Brockmeyer, J.; Karch, H.; Joannidis, M.; Clark, S.J.; Day, A.J.; Fidanzi, S.; et al. Shiga toxin activates complement and binds factor H: Evidence for an active role of complement in hemolytic uremic syndrome. J. Immunol. 2009, 182, 6394–6400. [Google Scholar] [CrossRef]
- Faserl, K.; Chetwynd, A.J.; Lynch, I.; Thorn, J.A.; Lindner, H.H. Corona Isolation Method Matters: Capillary Electrophoresis Mass Spectrometry Based Comparison of Protein Corona Compositions Following On-Particle versus In-Solution or In-Gel Digestion. Nanomaterials 2019, 9, 898. [Google Scholar] [CrossRef]
- Schoberleitner, I.; Faserl, K.; Sarg, B.; Egle, D.; Brunner, C.; Wolfram, D. Quantitative Proteomic Characterization of Foreign Body Response towards Silicone Breast Implants Identifies Chronological Disease-Relevant Biomarker Dynamics. Biomolecules 2023, 13, 305. [Google Scholar] [CrossRef]
- Lea, N.; Lord, J.M.; Roberts, L.M. Proteolytic cleavage of the A subunit is essential for maximal cytotoxicity of Escherichia coli O157:H7 Shiga-like toxin-1. Microbiology 1999, 145 Pt 5, 999–1004. [Google Scholar] [CrossRef]
- Kurmanova, A.; Llorente, A.; Polesskaya, A.; Garred, O.; Olsnes, S.; Kozlov, J.; Sandvig, K. Structural requirements for furin-induced cleavage and activation of Shiga toxin. Biochem. Biophys. Res. Commun. 2007, 357, 144–149. [Google Scholar] [CrossRef]
- Nichols, R.G.; Davenport, E.R. The relationship between the gut microbiome and host gene expression: A review. Hum. Genet. 2021, 140, 747–760. [Google Scholar] [CrossRef]
- Braun, E.; Sauter, D. Furin-mediated protein processing in infectious diseases and cancer. Clin. Transl. Immunol. 2019, 8, e1073. [Google Scholar] [CrossRef] [PubMed]
- De Martino, G.N. Calcium-dependent proteolytic activity in rat liver: Identification of two proteases with different calcium requirements. Arch. Biochem. Biophys. 1981, 211, 253–257. [Google Scholar] [CrossRef]
- McCall, K.A.; Huang, C.C.; Fierke, C.A. Function and mechanism of zinc metalloenzymes. J. Nutr. 2000, 130, 1437S–1446S. [Google Scholar] [CrossRef] [PubMed]
- Kangrga, R.N.; Ignjatović, S.D.; Dragaševic, M.M.; Jovičić, S.; Majkić-Singh, N.T. Pancreatic Elastase Levels in Feces As A Marker of Exocrine Pancreatic Function in Patients With Diabetes Mellitus. Lab. Med. 2016, 47, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Tóth, A.Z.; Szabó, A.; Hegyi, E.; Hegyi, P.; Sahin-Tóth, M. Detection of human elastase isoforms by the ScheBo Pancreatic Elastase 1 Test. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G606–G614. [Google Scholar] [CrossRef] [PubMed]
- Schüller, S. Shiga Toxin Interaction with Human Intestinal Epithelium. Toxins 2011, 3, 626–639. [Google Scholar] [CrossRef] [PubMed]
- Acheson, D.W.; Moore, R.; De Breucker, S.; Lincicome, L.; Jacewicz, M.; Skutelsky, E.; Keusch, G.T. Translocation of Shiga toxin across polarized intestinal cells in tissue culture. Infect. Immun. 1996, 64, 3294–3300. [Google Scholar] [CrossRef]
- Malyukova, I.; Murray, K.F.; Zhu, C.; Boedeker, E.; Kane, A.; Patterson, K.; Peterson, J.R.; Donowitz, M.; Kovbasnjuk, O. Macropinocytosis in Shiga toxin 1 uptake by human intestinal epithelial cells and transcellular transcytosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G78–G92. [Google Scholar] [CrossRef]
- Jablaoui, A.; Kriaa, A.; Mkaouar, H.; Akermi, N.; Soussou, S.; Wysocka, M.; Wołoszyn, D.; Amouri, A.; Gargouri, A.; Maguin, E.; et al. Fecal Serine Protease Profiling in Inflammatory Bowel Diseases. Front. Cell. Infect. Microbiol. 2020, 10, 21. [Google Scholar] [CrossRef]
- Pradhan, S.; Karve, S.S.; Weiss, A.A.; Hawkins, J.; Poling, H.M.; Helmrath, M.A.; Wells, J.M.; McCauley, H.A. Tissue Responses to Shiga Toxin in Human Intestinal Organoids. Cell. Mol. Gastroenterol. Hepatol. 2020, 10, 171–190. [Google Scholar] [CrossRef]
- Henrique, I.d.M.; Sacerdoti, F.; Ferreira, R.L.; Henrique, C.; Amaral, M.M.; Piazza, R.M.F.; Luz, D. Therapeutic Antibodies Against Shiga Toxins: Trends and Perspectives. Front. Cell. Infect. Microbiol. 2022, 12, 35. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kellnerová, S.; Huber, S.; Massri, M.; Fleischer, V.; Losso, K.; Sarg, B.; Kremser, L.; Talasz, H.; He, X.; Varrone, E.; et al. Enzymatic Cleavage of Stx2a in the Gut and Identification of Pancreatic Elastase and Trypsin as Possible Main Cleavers. Microorganisms 2023, 11, 2487. https://doi.org/10.3390/microorganisms11102487
Kellnerová S, Huber S, Massri M, Fleischer V, Losso K, Sarg B, Kremser L, Talasz H, He X, Varrone E, et al. Enzymatic Cleavage of Stx2a in the Gut and Identification of Pancreatic Elastase and Trypsin as Possible Main Cleavers. Microorganisms. 2023; 11(10):2487. https://doi.org/10.3390/microorganisms11102487
Chicago/Turabian StyleKellnerová, Sára, Silke Huber, Mariam Massri, Verena Fleischer, Klemens Losso, Bettina Sarg, Leopold Kremser, Heribert Talasz, Xiaohua He, Elisa Varrone, and et al. 2023. "Enzymatic Cleavage of Stx2a in the Gut and Identification of Pancreatic Elastase and Trypsin as Possible Main Cleavers" Microorganisms 11, no. 10: 2487. https://doi.org/10.3390/microorganisms11102487
APA StyleKellnerová, S., Huber, S., Massri, M., Fleischer, V., Losso, K., Sarg, B., Kremser, L., Talasz, H., He, X., Varrone, E., Brigotti, M., Ardissino, G., Orth-Höller, D., & Würzner, R. (2023). Enzymatic Cleavage of Stx2a in the Gut and Identification of Pancreatic Elastase and Trypsin as Possible Main Cleavers. Microorganisms, 11(10), 2487. https://doi.org/10.3390/microorganisms11102487