
Plant-Growth-Promoting Effect by Cell Components of Purple Non-Sulfur Photosynthetic Bacteria


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria, Plant and Chemicals
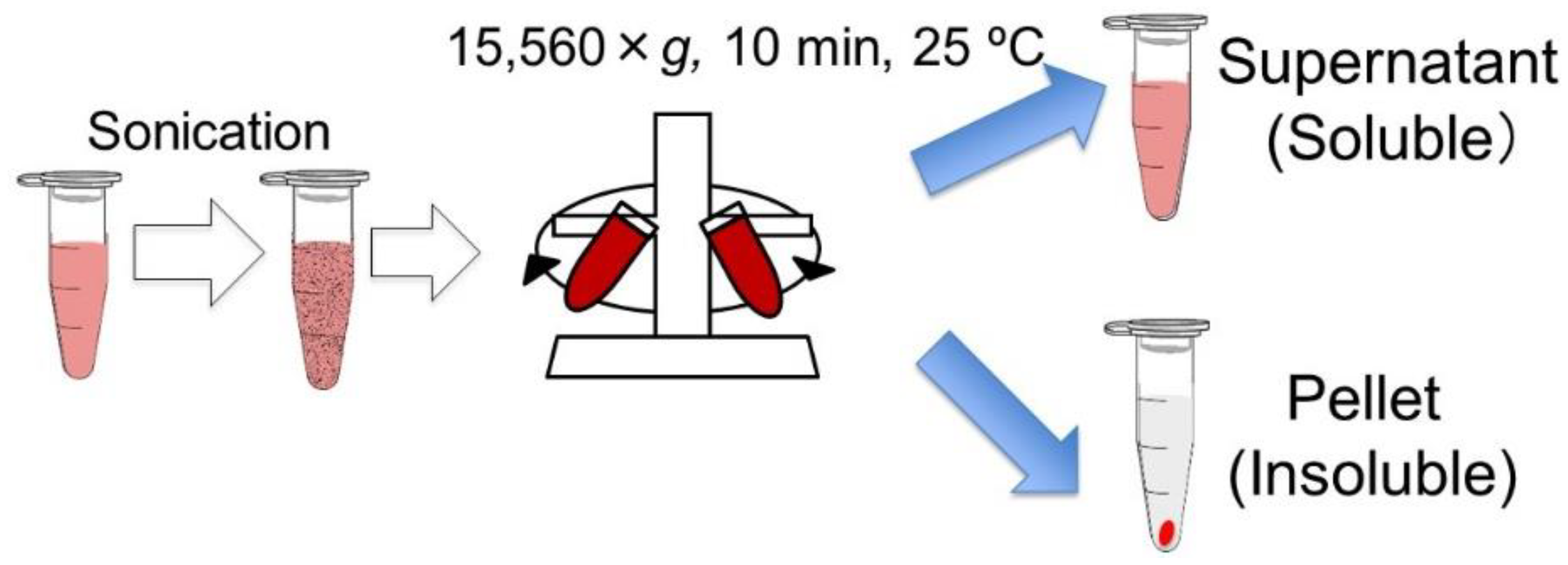
2.2. Preparation of Inactivated Cells and Cell Components
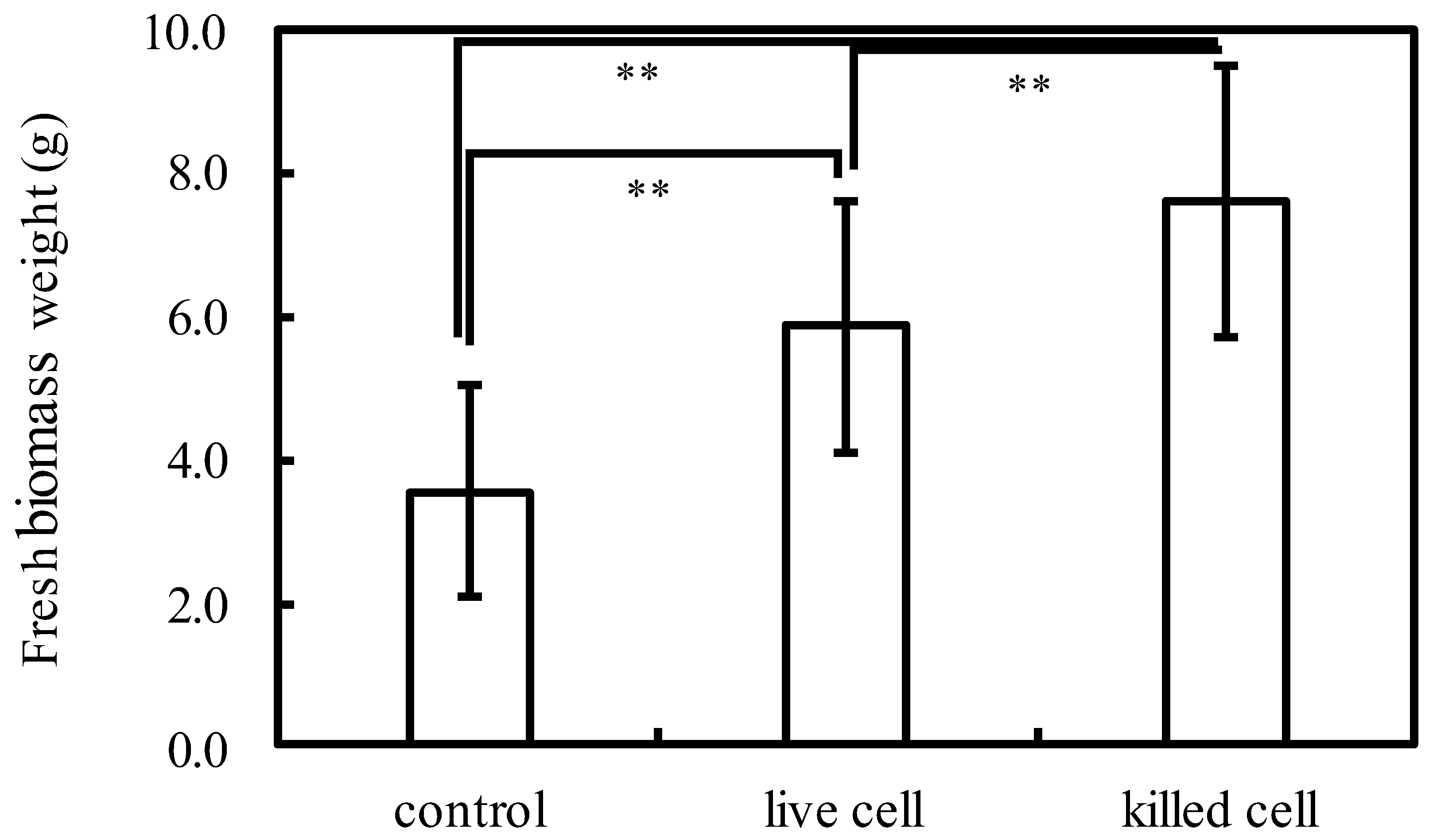
2.3. Live or Inactivated Cells Experimental Design
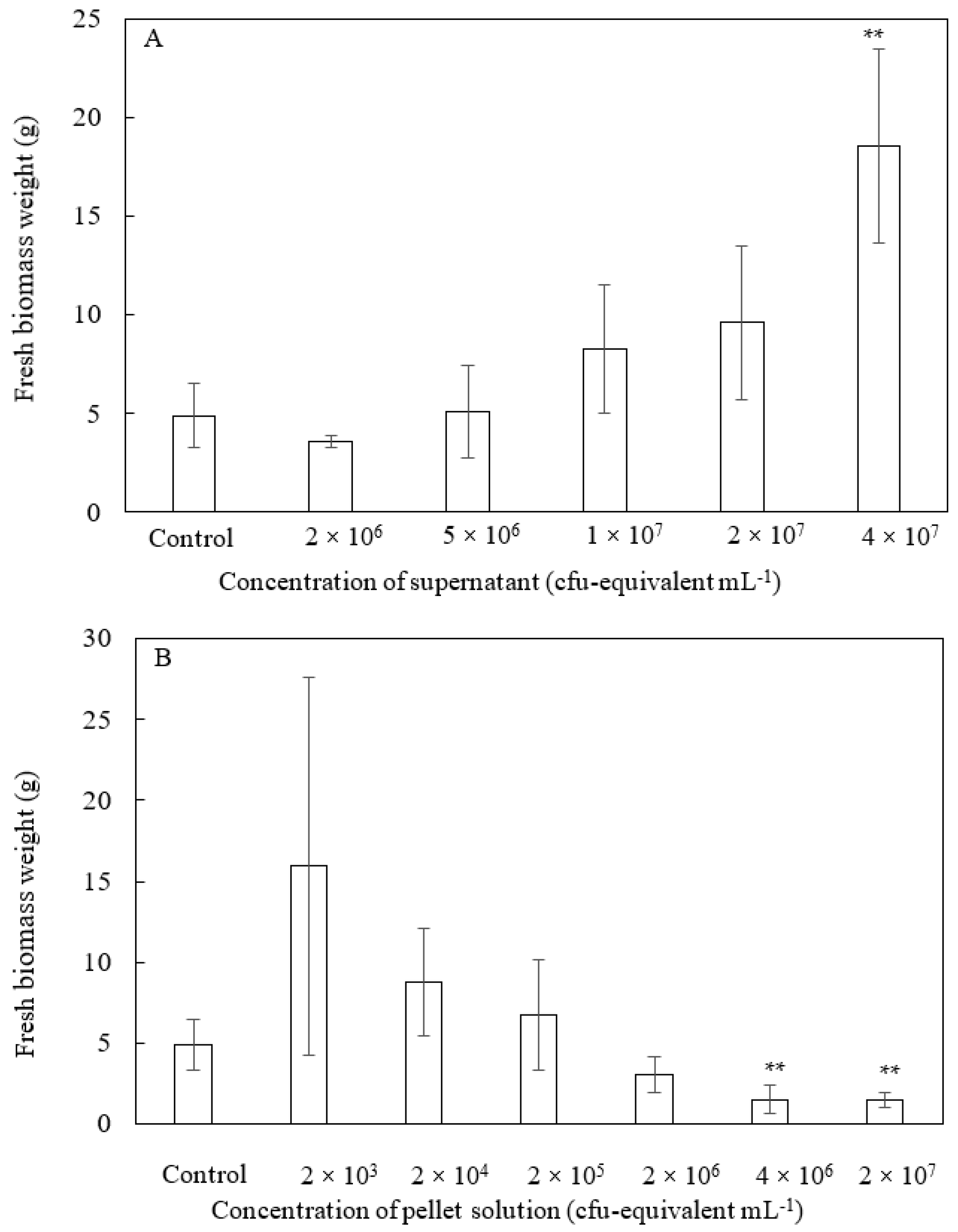
2.4. Cell Supernatant or Pellet Experimental Design
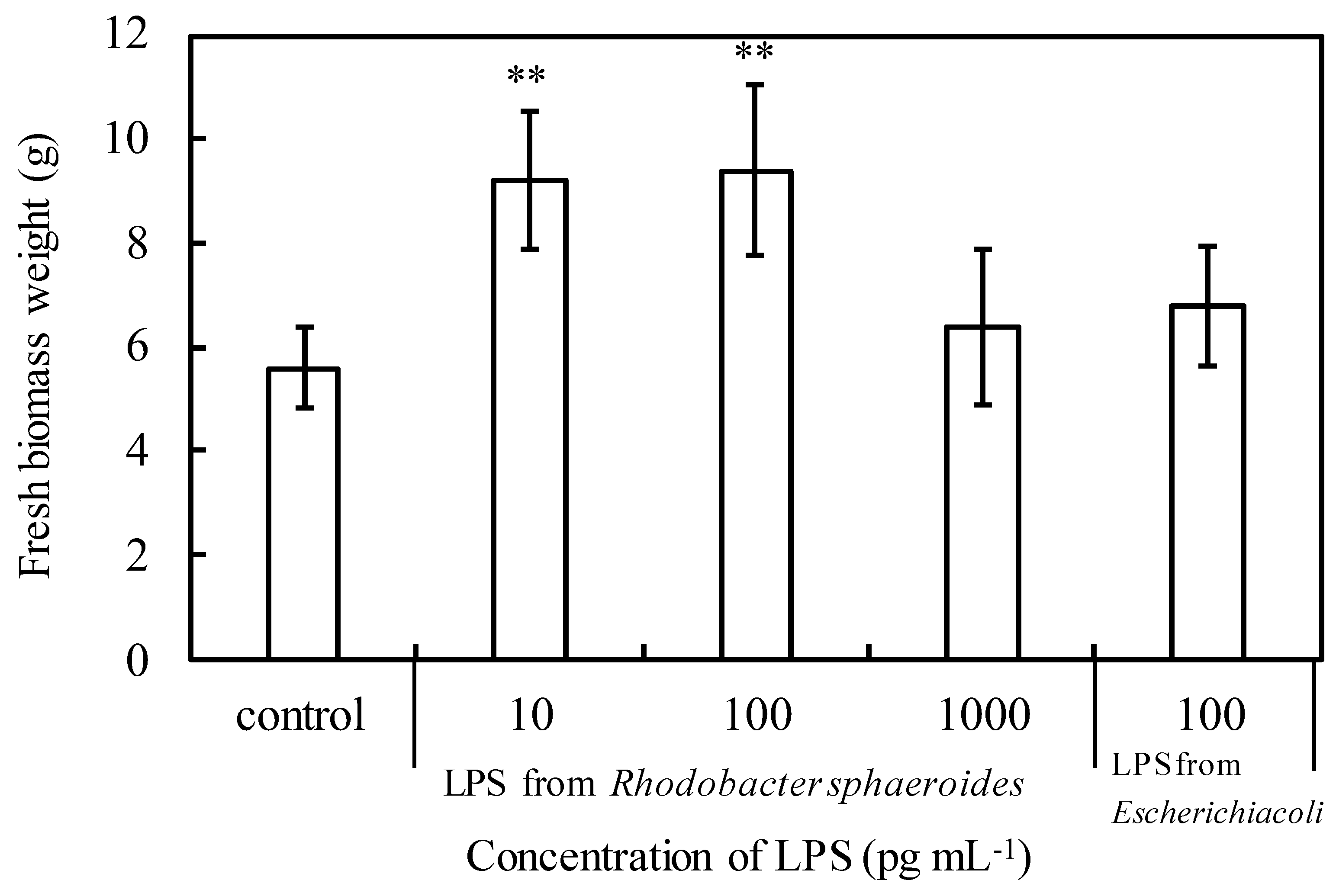
2.5. LPS Experimental Design
2.6. Statistical Analysis
3. Results
3.1. Effects of Soluble and Insoluble Fractions of Inactivated Cells on Plant Growth
3.2. Effects of Lipopolysaccharide (LPS) on Plant Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rana, G.; Meikap, S.; Mondol, M.; Bose, P.P. Green-fertilizer, Rhodospirillum rubrum, for agricultural development on fly-ash without any toxic metal ion release. Basic Res. J. Agric. Sci. Rev. 2016, 5, 109–117. [Google Scholar]
- Coppens, J.; Grunert, O.; Van Den Hende, S.; Vanhoutte, I.; Boon, N.; Haesaert, G.; De Gelder, L. The use of microalgae as a high-value organic slow-release fertilizer results in tomatoes with increased carotenoid and sugar levels. J. Appl. Phycol. 2016, 28, 2367–2377. [Google Scholar] [CrossRef]
- Pikaar, I.; de Vrieze, J.; Rabaey, K.; Herrero, M.; Smith, P.; Verstraete, W. Carbon emission avoidance and capture by producing in-reactor microbial biomass based food, feed and slow release fertilizer: Potentials and limitations. Sci. Total Environ. 2018, 644, 1525–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakarika, M.; Spanoghe, J.; Sui, Y.; Wambacq, E.; Grunert, O.; Haesaert, G.; Spiller, M.; Vlaeminck, S.E. Purple non-sulphur bacteria and plant production: Benefits for fertilization, stress resistance and the environment. Microb. Biotechnol. 2019, 13, 1336–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kantha, T.; Kantachote, D.; Klongdee, N. Potential of biofertilizers from selected Rhodopseudomonas palustris strains to assist rice (Oryza sativa L. subsp. indica) growth under salt stress and to reduce greenhouse gas emissions. Annu. Microbiol. 2015, 65, 2109–2118. [Google Scholar] [CrossRef]
- Kantachote, D.; Nunkaew, T.; Kantha, T.; Chaiprapat, S. Biofertilizers from Rhodopseudomonas palustris strains to enhance rice yields and reduce methane emissions. Appl. Soil. Ecol. 2016, 100, 154–161. [Google Scholar] [CrossRef]
- Sakpirom, J.; Kantachote, D.; Nunkaew, T.; Khan, E. Characterizations of purple non-sulfur bacteria isolated from paddy fields, and identification of strains with potential for plant growth-promotion, greenhouse gas mitigation and heavy metal bioremediation. Res. Microbiol. 2017, 168, 266–275. [Google Scholar] [CrossRef]
- Wong, W.-T.; Tseng, C.-H.; Hsu, S.-C.S.-H.; Lur, H.-S.; Mo, C.-W.; Huang, C.-N.; Hsu, S.-C.; Lee, K.-T.; Liu, C.-T. Promoting effects of a single Rhodopseudomonas palustris inoculant on plant growth by Brassica rapa chinensis under low fertilizer input. Microbes Environ. 2014, 29, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Koh, R.-H.; Song, H. Effects of application of Rhodopseudomonas sp. on seed germination and growth of tomato under axenic conditions. J. Microbiol. Biotechnol. 2007, 17, 1805–1810. [Google Scholar]
- Lee, K.-H.; Koh, R.-H.; Song, H.-G. Enhancement of growth and yield of tomato by Rhodopseudomonas sp. under greenhouse conditions. J. Microbiol. 2008, 46, 641–646. [Google Scholar] [CrossRef]
- Kantha, T.; Chaiyasut, C.; Kantachote, D.; Sukrong, S.; Muangprom, A. Selection of photosynthetic bacteria producing 5-aminolevulinic acid from soil of organic saline paddy fields from the Northeast region of Thailand. Afr. J. Microbiol. Res. 2010, 4, 1848–1855. [Google Scholar]
- Nunkaew, T.; Kantachote, D.; Kanzaki, H.; Nitoda, T.; Ritchie, R.J. Effects of 5-aminolevulinic acid (ALA)-containing supernatants from selected Rhodopseudomonas palustris strains on rice growth under NaCl stress, with mediating effects on chlorophyll, photosynthetic electron trans. Electron. J. Biotechnol. 2014, 17, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Elbadry, M.; El-Bassel, A.; Elbanna, K. Occurrence and dynamics of phototrophic purple nonsulphur bacteria compared with other asymbiotic nitrogen fixers in ricefields of Egypt. World J. Microbiol. Biotechnol. 1999, 15, 359–362. [Google Scholar] [CrossRef]
- Gamal-Eldin, H.; Elbanna, K. Field evidence for the potential of Rhodobacter capsulatus as biofertilizer for flooded rice. Curr. Microbiol. 2011, 62, 391–395. [Google Scholar] [CrossRef]
- Batool, K.; Zahra, F.; Rehman, Y. Arsenic-redox transformation and plant growth promotion by purple nonsulfur bacteria Rhodopseudomonas palustris CS2 and Rhodopseudomonas faecalis SS5. BioMed Res. Int. 2017, 2017, 6250327. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, N.; Koga, A.; Okuhata, H.; Tanaka, S.; Yamada, N.; Maki, T.; Hayashi, S.; Yamamoto, S.; Miyasaka, H. Isolation of a marine purple non-sulfur photosynthetic bacterium with a high ability of glycerol assimilation. Int. J. Plant Anim. Environ. Sci. 2019, 9, 214–221. [Google Scholar] [CrossRef]
- Erbs, G.; Newman, M.-A. The role of lipopolysaccharide and peptidoglycan, two glycosylated bacterial microbe-associated molecular patterns (MAMPs), in plant innate immunity. Mol. Plant Pathol. 2012, 13, 95–104. [Google Scholar] [CrossRef]
- Darveau, R.P.; Hancock, R.E.W. Procedure for isolation of bacterial lipopolysaccharides from both smooth and rough Pseudomonas aeruginosa and Salmonella typhimurium strains. J. Bacteriol. 1983, 155, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, N.; Honovich, J.P.; Hara, H.; Cotter, R.J.; Takayama, K. Location of fatty acids in lipid A obtained from lipopolysaccharide of Rhodopseudomonas sphaeroides ATCC 17023. J. Biol. Chem. 1988, 263, 5502–5504. [Google Scholar] [CrossRef]
- Coats, S.R.; Pham, T.-T.T.; Bainbridge, B.W.; Reife, R.A.; Darveau, R.P. MD-2 mediates the ability of tetra-acylated and penta-acylated lipopolysaccharides to antagonize Escherichia coli lipopolysaccharide at the TLR4 signaling complex. J. Immunol. 2005, 175, 4490–4498. [Google Scholar] [CrossRef] [Green Version]
- Teghanemt, A.; Zhang, D.; Levis, E.N.; Weiss, J.P.; Gioannini, T.L. Molecular basis of reduced potency of underacylated endotoxins. J. Immunol. 2005, 175, 4669–4676. [Google Scholar] [CrossRef] [PubMed]
- Visintin, A.; Halmen, K.A.; Latz, E.; Monks, B.G.; Golenbock, D.T. Pharmacological inhibition of endotoxin responses is achieved by targeting the TLR4 coreceptor, MD-2. J. Immunol. 2005, 175, 6465–6472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitoh, S.; Akashi, S.; Tamada, T.; Tanimura, N.; Kobayashi, M.; Konno, K.; Matsumoto, F.; Fukase, K.; Kusumoto, S.; Nagai, Y.; et al. Lipid A antagonist, lipid IVa, is distinct from lipid A in interaction with Toll-like receptor 4 (TLR4)-MD-2 and ligand-induced TLR4 oligomerization. Int. Immunol. 2004, 16, 961–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallejo-Ochoa, J.; López-Marmolejo, M.; Hernández-Esquivel, A.A.; Méndez-Gómez, M.; Suárez-Soria, L.N.; Castro-Mercado, E. Early plant growth and biochemical responses induced by Azospirillum brasilense Sp245 lipopolysaccharides in wheat (Triticum aestivum L.) seedlings are attenuated by procyanidin B2. Protoplasma 2018, 255, 685–694. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayashi, S.; Iwamoto, Y.; Hirakawa, Y.; Mori, K.; Yamada, N.; Maki, T.; Yamamoto, S.; Miyasaka, H. Plant-Growth-Promoting Effect by Cell Components of Purple Non-Sulfur Photosynthetic Bacteria. Microorganisms 2022, 10, 771. https://doi.org/10.3390/microorganisms10040771
Hayashi S, Iwamoto Y, Hirakawa Y, Mori K, Yamada N, Maki T, Yamamoto S, Miyasaka H. Plant-Growth-Promoting Effect by Cell Components of Purple Non-Sulfur Photosynthetic Bacteria. Microorganisms. 2022; 10(4):771. https://doi.org/10.3390/microorganisms10040771
Chicago/Turabian StyleHayashi, Shuhei, Yasunari Iwamoto, Yuki Hirakawa, Koichi Mori, Naoki Yamada, Takaaki Maki, Shinjiro Yamamoto, and Hitoshi Miyasaka. 2022. "Plant-Growth-Promoting Effect by Cell Components of Purple Non-Sulfur Photosynthetic Bacteria" Microorganisms 10, no. 4: 771. https://doi.org/10.3390/microorganisms10040771
APA StyleHayashi, S., Iwamoto, Y., Hirakawa, Y., Mori, K., Yamada, N., Maki, T., Yamamoto, S., & Miyasaka, H. (2022). Plant-Growth-Promoting Effect by Cell Components of Purple Non-Sulfur Photosynthetic Bacteria. Microorganisms, 10(4), 771. https://doi.org/10.3390/microorganisms10040771