
Targeting of Regulators as a Promising Approach in the Search for Novel Antimicrobial Agents




Abstract
1. Introduction
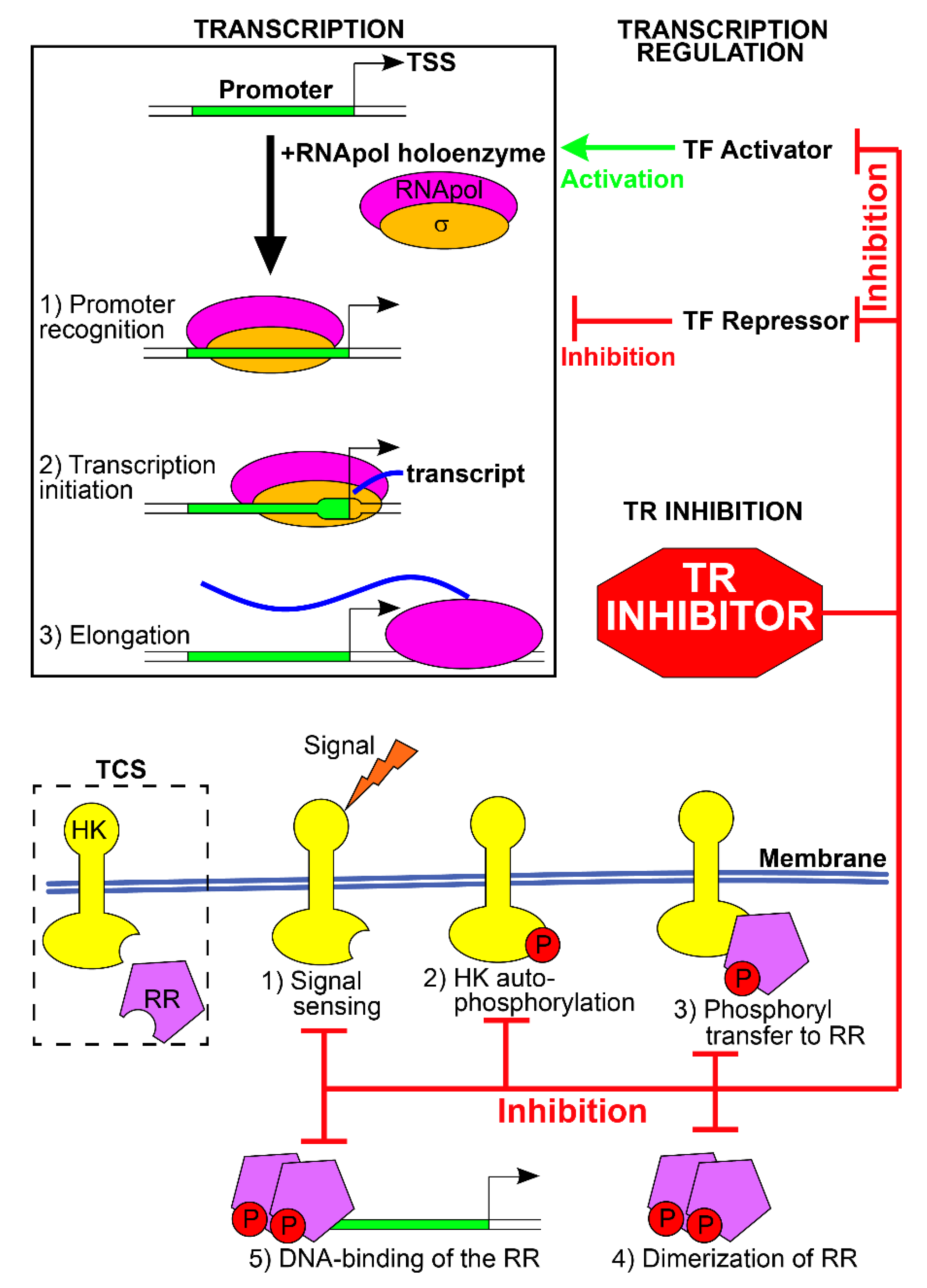
2. Transcriptional Regulators
2.1. TRs as New Targets for Antibacterial Therapies
2.2. Signal Sensing—ComD (Streptococcus pneumoniae)
2.3. Autophosphorylation of HK—QseC (Many Bacteria)
2.4. Phosphoryl Transfer to RR—BasR, CreB (E. coli)
2.5. DNA Binding—HsrA (Also Known as HP1043, Helicobacter pylori)
2.6. Dimerization—WalK/WalR (Multiple Microorganisms)
2.7. Unknown—VirF (Shigella flexneri)
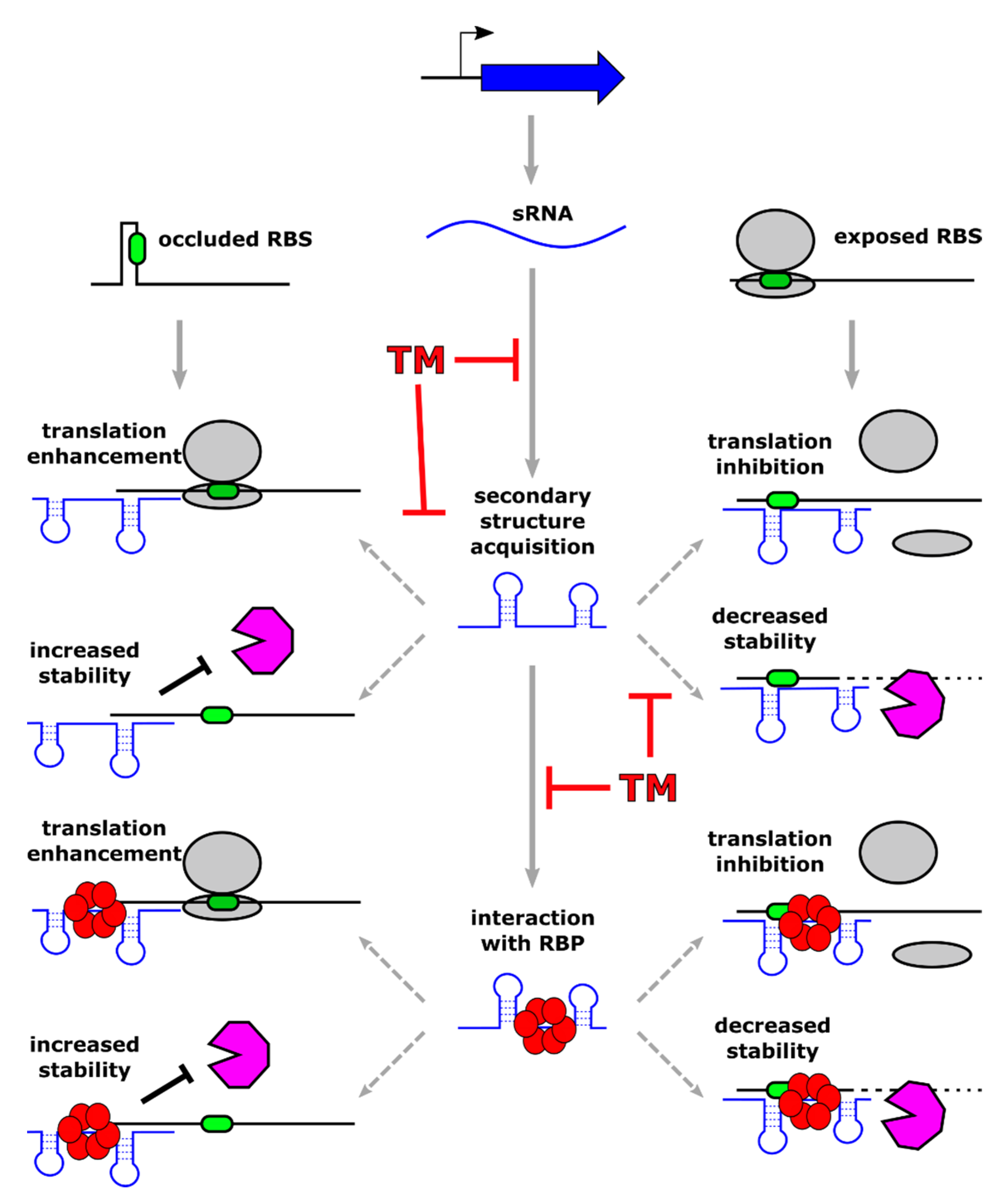
3. Posttranscriptional Regulators (sRNA)
3.1. sRNAs-Mediated Mechanisms of Regulation
3.2. Examples of Regulatory sRNAs Controlling Virulence and Pathogenesis
3.3. Examples of Regulatory sRNAs Involved in Antibiotic Response and Resistance
3.4. Regulatory sRNAs as Targets for Novel Antibacterial Molecules
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roncarati, D.; Scarlato, V. Regulation of heat-shock genes in bacteria: From signal sensing to gene expression output. FEMS Microbiol. Rev. 2017, 41, 549–574. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Fillat, M.F.; Lanas, A. Transcriptional regulators: Valuable targets for novel antibacterial strategies. Future Med. Chem. 2018, 10, 541–560. [Google Scholar] [CrossRef] [PubMed]
- Caldelari, I.; Chao, Y.; Romby, P.; Vogel, J. RNA-Mediated Regulation in Pathogenic Bacteria. Cold Spring Harb. Perspect. Med. 2013, 3, a010298. [Google Scholar] [CrossRef]
- Hirakawa, H.; Kurushima, J.; Hashimoto, Y.; Tomita, H. Progress Overview of Bacterial Two-Component Regulatory Systems as Potential Targets for Antimicrobial Chemotherapy. Antibiotics 2020, 9, 635. [Google Scholar] [CrossRef]
- Mejía-Almonte, C.; Busby, S.J.W.; Wade, J.T.; van Helden, J.; Arkin, A.P.; Stormo, G.D.; Eilbeck, K.; Palsson, B.O.; Galagan, J.E.; Collado-Vides, J. Redefining fundamental concepts of transcription initiation in bacteria. Nat. Rev. Genet. 2020, 21, 699–714. [Google Scholar] [CrossRef]
- Losick, R.; Pero, J. Cascades of sigma factors. Cell 1981, 25, 582–584. [Google Scholar] [CrossRef]
- Stock, A.M.; Robinson, V.L.; Goudreau, P.N. Two-Component Signal Transduction. Annu. Rev. Biochem. 2000, 69, 183–215. [Google Scholar] [CrossRef]
- Danielli, A.; Amore, G.; Scarlato, V. Built shallow to maintain homeostasis and persistent infection: Insight into the transcriptional regulatory network of the gastric human pathogen Helicobacter pylori. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef] [PubMed]
- Mühlen, S.; Dersch, P. Anti-virulence strategies to target bacterial infections. Curr Top Microbiol Immunol. 2016, 398, 147–183. [Google Scholar] [CrossRef]
- Johnson, B.K.; Abramovitch, R.B. Small Molecules That Sabotage Bacterial Virulence. Trends Pharmacol. Sci. 2017, 38, 339–362. [Google Scholar] [CrossRef]
- Good, J.A.D.; Andersson, C.; Hansen, S.; Wall, J.; Krishnan, K.S.; Begum, A.; Grundström, C.; Niemiec, M.S.; Vaitkevicius, K.; Chorell, E.; et al. Attenuating Listeria monocytogenes Virulence by Targeting the Regulatory Protein PrfA. Cell Chem. Biol. 2016, 23, 404–414. [Google Scholar] [CrossRef]
- Bellale, E.; Naik, M.; VB, V.; Ambady, A.; Narayan, A.; Ravishankar, S.; Ramachandran, V.; Kaur, P.; McLaughlin, R.; Whiteaker, J.; et al. Diarylthiazole: An Antimycobacterial Scaffold Potentially Targeting PrrB-PrrA Two-Component System. J. Med. Chem. 2014, 57, 6572–6582. [Google Scholar] [CrossRef]
- Zheng, H.; Williams, J.T.; Aleiwi, B.; Ellsworth, E.; Abramovitch, R.B. Inhibiting Mycobacterium tuberculosis DosRST Signaling by Targeting Response Regulator DNA Binding and Sensor Kinase Heme. ACS Chem. Biol. 2020, 15, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Taneja, N.K.; Dhingra, S.; Tyagi, J.S. DevR (DosR) mimetic peptides impair transcriptional regulation and survival of Mycobacterium tuberculosis under hypoxia by inhibiting the autokinase activity of DevS sensor kinase. BMC Microbiol. 2014, 14, 195. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, M.; Southall, N.; Zheng, W.; Wang, S. A High-Throughput Assay for Developing Inhibitors of PhoP, a Virulence Factor of Mycobacterium tuberculosis. Comb. Chem. High Throughput Screen. 2016, 19, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.K.; Colvin, C.J.; Needle, D.B.; Mba Medie, F.; Champion, P.A.D.; Abramovitch, R.B. The Carbonic Anhydrase Inhibitor Ethoxzolamide Inhibits the Mycobacterium tuberculosis PhoPR Regulon and Esx-1 Secretion and Attenuates Virulence. Antimicrob. Agents Chemother. 2015, 59, 4436–4445. [Google Scholar] [CrossRef]
- Guarnieri, M.T.; Zhang, L.; Shen, J.; Zhao, R. The Hsp90 Inhibitor Radicicol Interacts with the ATP-Binding Pocket of Bacterial Sensor Kinase PhoQ. J. Mol. Biol. 2008, 379, 82–93. [Google Scholar] [CrossRef]
- Tang, Y.T.; Gao, R.; Havranek, J.J.; Groisman, E.A.; Stock, A.M.; Marshall, G.R. Inhibition of Bacterial Virulence: Drug-Like Molecules Targeting the Salmonella enterica PhoP Response Regulator. Chem. Biol. Drug Des. 2012, 79, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.N.; MacNair, C.R.; Cao, M.P.T.; Perry, J.N.; Magolan, J.; Brown, E.D.; Coombes, B.K. Targeting Two-Component Systems Uncovers a Small-Molecule Inhibitor of Salmonella Virulence. Cell Chem. Biol. 2020, 27, 793–805.e7. [Google Scholar] [CrossRef]
- González, A.; Salillas, S.; Velázquez-Campoy, A.; Espinosa Angarica, V.; Fillat, M.F.; Sancho, J.; Lanas, Á. Identifying potential novel drugs against Helicobacter pylori by targeting the essential response regulator HsrA. Sci. Rep. 2019, 9, 11294. [Google Scholar] [CrossRef]
- González, A.; Casado, J.; Chueca, E.; Salillas, S.; Velázquez-Campoy, A.; Sancho, J.; Lanas, Á. Small Molecule Inhibitors of the Response Regulator ArsR Exhibit Bactericidal Activity against Helicobacter pylori. Microorganisms 2020, 8, 503. [Google Scholar] [CrossRef]
- Emanuele, A.A.; Adams, N.E.; Chen, Y.-C.; Maurelli, A.T.; Garcia, G.A. Potential novel antibiotics from HTS targeting the virulence-regulating transcription factor, VirF, from Shigella flexneri. J. Antibiot. 2014, 67, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Zhang, J.; Chen, M.; Wu, Y.; Wang, X.; Chen, J.; Zhang, J.; Shen, X.; Qu, D.; Jiang, H. The Effect of the Potential PhoQ Histidine Kinase Inhibitors on Shigella flexneri Virulence. PLoS ONE 2011, 6, e23100. [Google Scholar] [CrossRef] [PubMed]
- Hung, D.T.; Shakhnovich, E.A.; Pierson, E.; Mekalanos, J.J. Small-Molecule Inhibitor of Vibrio cholerae Virulence and Intestinal Colonization. Science 2005, 310, 670–674. [Google Scholar] [CrossRef]
- Shakhnovich, E.A.; Hung, D.T.; Pierson, E.; Lee, K.; Mekalanos, J.J. Virstatin inhibits dimerization of the transcriptional activator ToxT. Proc. Natl. Acad. Sci. USA 2007, 104, 2372–2377. [Google Scholar] [CrossRef]
- Anthouard, R.; DiRita, V.J. Small-Molecule Inhibitors of toxT Expression in Vibrio cholerae. mBio 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Ulijasz, A.T.; Weisblum, B. Dissecting the VanRS Signal Transduction Pathway with Specific Inhibitors. J. Bacteriol. 1999, 181, 627–631. [Google Scholar] [CrossRef]
- Ulijasz, A.T.; Kay, B.K.; Weisblum, B. Peptide Analogues of the VanS Catalytic Center Inhibit VanR Binding to Its Cognate Promoter. Biochemistry 2000, 39, 11417–11424. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, Y.; Doi, A.; Furuta, E.; Dubrac, S.; Ishizaki, Y.; Okada, M.; Igarashi, M.; Misawa, N.; Yoshikawa, H.; Okajima, T.; et al. Novel antibacterial compounds specifically targeting the essential WalR response regulator. J. Antibiot. 2010, 63, 127–134. [Google Scholar] [CrossRef]
- Watanabe, T.; Igarashi, M.; Okajima, T.; Ishii, E.; Kino, H.; Hatano, M.; Sawa, R.; Umekita, M.; Kimura, T.; Okamoto, S.; et al. Isolation and Characterization of Signermycin B, an Antibiotic That Targets the Dimerization Domain of Histidine Kinase WalK. Antimicrob. Agents Chemother. 2012, 56, 3657–3663. [Google Scholar] [CrossRef]
- Furuta, E.; Yamamoto, K.; Tatebe, D.; Watabe, K.; Kitayama, T.; Utsumi, R. Targeting protein homodimerization: A novel drug discovery system. FEBS Lett. 2005, 579, 2065–2070. [Google Scholar] [CrossRef]
- Kitayama, T.; Iwabuchi, R.; Minagawa, S.; Shiomi, F.; Cappiello, J.; Sawada, S.; Utsumi, R.; Okamoto, T. Unprecedented olefin-dependent histidine-kinase inhibitory of zerumbone ring-opening material. Bioorg. Med. Chem. Lett. 2004, 14, 5943–5946. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, Y.; Kubo, N.; Matsunaga, H.; Igarashi, M.; Utsumi, R. Development of an Antivirulence Drug against Streptococcus mutans: Repression of Biofilm Formation, Acid Tolerance, and Competence by a Histidine Kinase Inhibitor, Walkmycin C. Antimicrob. Agents Chemother. 2011, 55, 1475–1484. [Google Scholar] [CrossRef]
- Gilmour, R.; Foster, J.E.; Sheng, Q.; McClain, J.R.; Riley, A.; Sun, P.-M.; Ng, W.-L.; Yan, D.; Nicas, T.I.; Henry, K.; et al. New Class of Competitive Inhibitor of Bacterial Histidine Kinases. J. Bacteriol. 2005, 187, 8196–8200. [Google Scholar] [CrossRef]
- Abed, N.; Bickle, M.; Mari, B.; Schapira, M.; Sanjuan-España, R.; Sermesant, K.R.; Moncorgé, O.; Mouradian-Garcia, S.; Barbry, P.; Rudkin, B.B.; et al. A Comparative Analysis of Perturbations Caused by a Gene Knock-out, a Dominant Negative Allele, and a Set of Peptide Aptamers. Mol. Cell. Proteom. 2007, 6, 2110–2121. [Google Scholar] [CrossRef] [PubMed]
- Roychoudhury, S.; Zielinski, N.A.; Ninfa, A.J.; Allen, N.E.; Jungheim, L.N.; Nicas, T.I.; Chakrabarty, A.M. Inhibitors of two-component signal transduction systems: Inhibition of alginate gene activation in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1993, 90, 965–969. [Google Scholar] [CrossRef]
- Ho, Y.-H.; Sung, T.-C.; Chen, C.-S. Lactoferricin B Inhibits the Phosphorylation of the Two-Component System Response Regulators BasR and CreB. Mol. Cell. Proteom. 2012, 11, M111.014720. [Google Scholar] [CrossRef] [PubMed]
- Strauch, M.A.; Mendoza, D.; Hoch, J.A. cis-Unsaturated fatty acids specifically inhibit a signal-transducing protein kinase required for initiation of sporulation in Bacillus subtilis. Mol. Microbiol. 1992, 6, 2909–2917. [Google Scholar] [CrossRef] [PubMed]
- Grossman, S.; Soukarieh, F.; Richardson, W.; Liu, R.; Mashabi, A.; Emsley, J.; Williams, P.; Cámara, M.; Stocks, M.J. Novel quinazolinone inhibitors of the Pseudomonas aeruginosa quorum sensing transcriptional regulator PqsR. Eur. J. Med. Chem. 2020, 208, 112778. [Google Scholar] [CrossRef]
- Maura, D.; Rahme, L.G. Pharmacological Inhibition of the Pseudomonas aeruginosa MvfR Quorum-Sensing System Interferes with Biofilm Formation and Potentiates Antibiotic-Mediated Biofilm Disruption. Antimicrob. Agents Chemother. 2017, 61, e01362-17. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Kirsch, B.; Maurer, C.K.; de Jong, J.C.; Braunshausen, A.; Steinbach, A.; Hartmann, R.W. Optimization of anti-virulence PqsR antagonists regarding aqueous solubility and biological properties resulting in new insights in structure—Activity relationships. Eur. J. Med. Chem. 2014, 79, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Ilangovan, A.; Fletcher, M.; Rampioni, G.; Pustelny, C.; Rumbaugh, K.; Heeb, S.; Cámara, M.; Truman, A.; Chhabra, S.R.; Emsley, J.; et al. Structural Basis for Native Agonist and Synthetic Inhibitor Recognition by the Pseudomonas aeruginosa Quorum Sensing Regulator PqsR (MvfR). PLoS Pathog. 2013, 9, e1003508. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, F.; Baldelli, V.; Halliday, N.; Pantalone, P.; Polticelli, F.; Fiscarelli, E.; Williams, P.; Visca, P.; Leoni, L.; Rampioni, G. Identification of FDA-Approved Drugs as Antivirulence Agents Targeting the pqs Quorum-Sensing System of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Moreira, C.G.; Li, D.R.; Reading, N.C.; Ritchie, J.M.; Waldor, M.K.; Williams, N.; Taussig, R.; Wei, S.; Roth, M.; et al. Targeting QseC Signaling and Virulence for Antibiotic Development. Science 2008, 321, 1078–1080. [Google Scholar] [CrossRef] [PubMed]
- Curtis, M.M.; Russell, R.; Moreira, C.G.; Adebesin, A.M.; Wang, C.; Williams, N.S.; Taussig, R.; Stewart, D.; Zimmern, P.; Lu, B.; et al. QseC Inhibitors as an Antivirulence Approach for Gram-Negative Pathogens. mBio 2014, 5, e02165-14. [Google Scholar] [CrossRef] [PubMed]
- Lyon, G.J.; Mayville, P.; Muir, T.W.; Novick, R.P. Rational design of a global inhibitor of the virulence response in Staphylococcus aureus, based in part on localization of the site of inhibition to the receptor-histidine kinase, AgrC. Proc. Natl. Acad. Sci. USA 2000, 97, 13330–13335. [Google Scholar] [CrossRef]
- Sully, E.K.; Malachowa, N.; Elmore, B.O.; Alexander, S.M.; Femling, J.K.; Gray, B.M.; DeLeo, F.R.; Otto, M.; Cheung, A.L.; Edwards, B.S.; et al. Selective Chemical Inhibition of agr Quorum Sensing in Staphylococcus aureus Promotes Host Defense with Minimal Impact on Resistance. PLoS Pathog. 2014, 10, e1004174. [Google Scholar] [CrossRef]
- Yang, T.; Tal-Gan, Y.; Paharik, A.E.; Horswill, A.R.; Blackwell, H.E. Structure—Function Analyses of a Staphylococcus epidermidis Autoinducing Peptide Reveals Motifs Critical for AgrC-type Receptor Modulation. ACS Chem. Biol. 2016, 11, 1982–1991. [Google Scholar] [CrossRef]
- Zhu, L.; Lau, G.W. Inhibition of Competence Development, Horizontal Gene Transfer and Virulence in Streptococcus pneumoniae by a Modified Competence Stimulating Peptide. PLoS Pathog. 2011, 7, e1002241. [Google Scholar] [CrossRef]
- Yang, Y.; Tal-Gan, Y. Exploring the competence stimulating peptide (CSP) N-terminal requirements for effective ComD receptor activation in group1 Streptococcus pneumoniae. Bioorg. Chem. 2019, 89, 102987. [Google Scholar] [CrossRef]
- Nakayama, J.; Yokohata, R.; Sato, M.; Suzuki, T.; Matsufuji, T.; Nishiguchi, K.; Kawai, T.; Yamanaka, Y.; Nagata, K.; Tanokura, M.; et al. Development of a Peptide Antagonist against fsr Quorum Sensing of Enterococcus faecalis. ACS Chem. Biol. 2013, 8, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.J.; Bobay, B.G.; Stowe, S.D.; Olson, A.L.; Peng, L.; Su, Z.; Actis, L.A.; Melander, C.; Cavanagh, J. Identification of BfmR, a Response Regulator Involved in Biofilm Development, as a Target for a 2-Aminoimidazole-Based Antibiofilm Agent. Biochemistry 2012, 51, 9776–9778. [Google Scholar] [CrossRef] [PubMed]
- Pelliciari, S.; Pinatel, E.; Vannini, A.; Peano, C.; Puccio, S.; De Bellis, G.; Danielli, A.; Scarlato, V.; Roncarati, D. Insight into the essential role of the Helicobacter pylori HP1043 orphan response regulator: Genome-wide identification and characterization of the DNA-binding sites. Sci. Rep. 2017, 7, 41063. [Google Scholar] [CrossRef]
- Zannoni, A.; Pelliciari, S.; Musiani, F.; Chiappori, F.; Roncarati, D.; Scarlato, V. Definition of the Binding Architecture to a Target Promoter of HP1043, the Essential Master Regulator of Helicobacter pylori. Int. J. Mol. Sci. 2021, 22, 7848. [Google Scholar] [CrossRef]
- Di Martino, M.L.; Falconi, M.; Micheli, G.; Colonna, B.; Prosseda, G. The Multifaceted Activity of the VirF Regulatory Protein in the Shigella Lifestyle. Front. Mol. Biosci. 2016, 3, 61. [Google Scholar] [CrossRef]
- Dutta, T.; Srivastava, S. Small RNA-mediated regulation in bacteria: A growing palette of diverse mechanisms. Gene 2018, 656, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Ikemura, T.; Dahlberg, J.E. Small ribonucleic acids of Escherichia coli. I. Characterization by polyacrylamide gel electrophoresis and fingerprint analysis. J. Biol. Chem. 1973, 248, 5024–5032. [Google Scholar] [CrossRef]
- Mizuno, T.; Chou, M.Y.; Inouye, M. A unique mechanism regulating gene expression: Translational inhibition by a complementary RNA transcript (micRNA). Proc. Natl. Acad. Sci. USA 1984, 81, 1966–1970. [Google Scholar] [CrossRef]
- Novick, R.P.; Iordanescu, S.; Projan, S.J.; Kornblum, J.; Edelman, I. pT181 plasmid replication is regulated by a countertranscript-driven transcriptional attenuator. Cell 1989, 59, 395–404. [Google Scholar] [CrossRef]
- Novick, R.P.; Ross, H.F.; Projan, S.J.; Kornblum, J.; Kreiswirth, B.; Moghazeh, S. Synthesis of staphylococcal virulence factors is controlled by a regulatory RNA molecule. EMBO J. 1993, 12, 3967–3975. [Google Scholar] [CrossRef]
- Bossi, L.; Figueroa-Bossi, N. Competing endogenous RNAs: A target-centric view of small RNA regulation in bacteria. Nat. Rev. Microbiol. 2016, 14, 775–784. [Google Scholar] [CrossRef]
- Potts, A.H.; Vakulskas, C.A.; Pannuri, A.; Yakhnin, H.; Babitzke, P.; Romeo, T. Global role of the bacterial post-transcriptional regulator CsrA revealed by integrated transcriptomics. Nat. Commun. 2017, 8, 1596. [Google Scholar] [CrossRef]
- Kavita, K.; de Mets, F.; Gottesman, S. New aspects of RNA-based regulation by Hfq and its partner sRNAs. Curr. Opin. Microbiol. 2018, 42, 53–61. [Google Scholar] [CrossRef]
- Holmqvist, E.; Berggren, S.; Rizvanovic, A. RNA-binding activity and regulatory functions of the emerging sRNA-binding protein ProQ. Biochim. Biophys. Acta Gene Regul. Mech. 2020, 1863, 194596. [Google Scholar] [CrossRef] [PubMed]
- Bossi, L.; Figueroa-Bossi, N.; Bouloc, P.; Boudvillain, M. Regulatory interplay between small RNAs and transcription termination factor Rho. Biochim. Biophys. Acta-Gene Regul. Mech. 2020, 1863, 194546. [Google Scholar] [CrossRef] [PubMed]
- Felden, B.; Augagneur, Y. Diversity and Versatility in Small RNA-Mediated Regulation in Bacterial Pathogens. Front. Microbiol. 2021, 12, 719977. [Google Scholar] [CrossRef]
- Wagner, E.G.H.; Romby, P. Small RNAs in Bacteria and Archaea: Who they are, what they do, and how they do it. In Advances in Genetics; Academic Press: Cambridge, MA, USA, 2015; pp. 133–208. [Google Scholar]
- Melamed, S.; Peer, A.; Faigenbaum-Romm, R.; Gatt, Y.E.; Reiss, N.; Bar, A.; Altuvia, Y.; Argaman, L.; Margalit, H. Global Mapping of Small RNA-Target Interactions in Bacteria. Mol. Cell 2016, 63, 884–897. [Google Scholar] [CrossRef] [PubMed]
- Bronesky, D.; Wu, Z.; Marzi, S.; Walter, P.; Geissmann, T.; Moreau, K.; Vandenesch, F.; Caldelari, I.; Romby, P. Staphylococcus aureus RNAIII and Its Regulon Link Quorum Sensing, Stress Responses, Metabolic Adaptation, and Regulation of Virulence Gene Expression. Annu. Rev. Microbiol. 2016, 70, 299–316. [Google Scholar] [CrossRef]
- Gupta, R.K.; Luong, T.T.; Lee, C.Y. RNAIII of the Staphylococcus aureus agr system activates global regulator MgrA by stabilizing mRNA. Proc. Natl. Acad. Sci. USA 2015, 112, 14036–14041. [Google Scholar] [CrossRef]
- Vannini, A.; Roncarati, D.; Spinsanti, M.; Scarlato, V.; Danielli, A. In depth analysis of the Helicobacter pylori cag pathogenicity island transcriptional responses. PLoS ONE 2014, 9, e98416. [Google Scholar] [CrossRef]
- Vannini, A.; Roncarati, D.; Danielli, A. The cag-pathogenicity island encoded CncR1 sRNA oppositely modulates Helicobacter pylori motility and adhesion to host cells. Cell. Mol. Life Sci. 2016, 73, 3151–3168. [Google Scholar] [CrossRef]
- Vannini, A.; Pinatel, E.; Costantini, P.E.; Pelliciari, S.; Roncarati, D.; Puccio, S.; De Bellis, G.; Peano, C.; Danielli, A. Comprehensive mapping of the Helicobacter pylori NikR regulon provides new insights in bacterial nickel responses. Sci. Rep. 2017, 7, 45458. [Google Scholar] [CrossRef]
- Eisenbart, S.K.; Alzheimer, M.; Pernitzsch, S.R.; Dietrich, S.; Stahl, S.; Sharma, C.M. A Repeat-Associated Small RNA Controls the Major Virulence Factors of Helicobacter pylori. Mol. Cell 2020, 80, 210–226.e7. [Google Scholar] [CrossRef]
- Kinoshita-Daitoku, R.; Kiga, K.; Miyakoshi, M.; Otsubo, R.; Ogura, Y.; Sanada, T.; Bo, Z.; Phuoc, T.V.; Okano, T.; Iida, T.; et al. A bacterial small RNA regulates the adaptation of Helicobacter pylori to the host environment. Nat. Commun. 2021, 12, 2085. [Google Scholar] [CrossRef]
- Joshi, B.; Singh, B.; Nadeem, A.; Askarian, F.; Wai, S.N.; Johannessen, M.; Hegstad, K. Transcriptome Profiling of Staphylococcus aureus Associated Extracellular Vesicles Reveals Presence of Small RNA-Cargo. Front. Mol. Biosci. 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, Y.; Song, Z.; Li, R.; Ruan, H.; Liu, Q.; Huang, X. sncRNAs packaged by Helicobacter pylori outer membrane vesicles attenuate IL-8 secretion in human cells. Int. J. Med. Microbiol. 2020, 310, 151356. [Google Scholar] [CrossRef]
- Koeppen, K.; Hampton, T.H.; Jarek, M.; Scharfe, M.; Gerber, S.A.; Mielcarz, D.W.; Demers, E.G.; Dolben, E.L.; Hammond, J.H.; Hogan, D.A.; et al. A Novel Mechanism of Host-Pathogen Interaction through sRNA in Bacterial Outer Membrane Vesicles. PLoS Pathog. 2016, 12, e1005672. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi Badi, S.; Bruno, S.P.; Moshiri, A.; Tarashi, S.; Siadat, S.D.; Masotti, A. Small RNAs in Outer Membrane Vesicles and Their Function in Host-Microbe Interactions. Front. Microbiol. 2020, 11, 1209. [Google Scholar] [CrossRef]
- Chareyre, S.; Mandin, P. Bacterial Iron Homeostasis Regulation by sRNAs. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Mediati, D.G.; Wu, S.; Wu, W.; Tree, J.J. Networks of Resistance: Small RNA Control of Antibiotic Resistance. Trends Genet. 2021, 37, 35–45. [Google Scholar] [CrossRef]
- Pinel-Marie, M.-L.; Brielle, R.; Felden, B. Dual Toxic-Peptide-Coding Staphylococcus aureus RNA under Antisense Regulation Targets Host Cells and Bacterial Rivals Unequally. Cell Rep. 2014, 7, 424–435. [Google Scholar] [CrossRef]
- Pinel-Marie, M.-L.; Brielle, R.; Riffaud, C.; Germain-Amiot, N.; Polacek, N.; Felden, B. RNA antitoxin SprF1 binds ribosomes to attenuate translation and promote persister cell formation in Staphylococcus aureus. Nat. Microbiol. 2021, 6, 209–220. [Google Scholar] [CrossRef]
- Jayaraman, K.; McParland, K.; Miller, P.; Ts’o, P.O.P. Selective inhibition of Escherichia coli protein synthesis and growth by nonionic oligonucleotides complementary to the 3′ end of 16S rRNA. Proc. Natl. Acad. Sci. USA 1981, 78, 1537–1541. [Google Scholar] [CrossRef]
- Sully, E.K.; Geller, B.L. Antisense antimicrobial therapeutics. Curr. Opin. Microbiol. 2016, 33, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Geller, B.L.; Li, L.; Martinez, F.; Sully, E.; Sturge, C.R.; Daly, S.M.; Pybus, C.; Greenberg, D.E. Morpholino oligomers tested in vitro, in biofilm and in vivo against multidrug-resistant Klebsiella pneumoniae. J. Antimicrob. Chemother. 2018, 73, 1611–1619. [Google Scholar] [CrossRef]
- Vogel, J. An RNA biology perspective on species-specific programmable RNA antibiotics. Mol. Microbiol. 2020, 113, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.E. Peptide Nucleic Acids (PNA) in Chemical Biology and Drug Discovery. Chem. Biodivers. 2010, 7, 786–804. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Target TR | Category of Targeted TR | Microorganism | Inhibitory Effects of the Compounds on the TR | Effects of the Compounds on the Bacterium (Type of Test, Amount of Compound) | Refs. |
|---|---|---|---|---|---|
| PrfA | Activator | L. monocytogenes | DNA-binding | Reduce virulence (DNA-binding IC50 6–7 μM) | [11] |
| PrrB/PrrA | HK | M. tuberculosis | Unknown | Bactericidal (MIC 0.06–8 μg/mL) | [12] |
| DosT/DosS/DosR | HKs, RR | M. tuberculosis | Sensor of the HKs, autophosphorylation of HK, or DNA-binding of the RR | Decrease the survival in specific growth conditions (EC50 0.6–9.8 μM) | [13,14] |
| PhoR/PhoP | RR | M. tuberculosis | DNA-binding of the RR | Decrease virulence (IC50 5 μM, 100 μM in macrophages, 100 mg/Kg in mice) | [15,16] |
| PhoQ/PhoP | HK, RR | S. enterica | Autophosphorylation of HK or DNA-binding of the RR | Unknown (DNA-binding IC50 3.6–285 μM) | [17,18] |
| SsrA/SsrB | RR | S. enterica | Unknown | Decrease virulence and sensitize the pathogen to other antibiotics (2.5 mg/Kg in mice) | [19] |
| PmrB/PmrA | RR | S. enterica | Unknown | Decrease virulence and sensitize the pathogen to other antibiotics (2.5 mg/Kg in mice) | [19] |
| HsrA (HP1043) | Orphan RR | H. pylori | DNA-binding | Bactericidal (MIC/MBC 4–128 mg/L) | [20] |
| ArsS/ArsR | RR | H. pylori | DNA-binding | Bactericidal (MIC/MBC 32–128 mg/L) | [21] |
| VirF | Activator | S. flexneri | Unknown | Decrease virulence and host invasion (IC50 14–66 μM) | [22] |
| PhoQ/PhoP | HK | S. flexneri | Autophosphorylation of HK | Decrease virulence and host invasion (enzymatic inhibition IC50 8–70 μM) | [23] |
| ToxT | Activator | V. cholerae | Dimerization or ToxT expression | Decrease host colonization (IC50 20–30 μM, EC50 2.7–25 μM, 100–200 mg/mouse) | [24,25,26] |
| VanS/VanR | RR | E. faecium | Phosphoryl transfer to RR or DNA-binding of the RR | Unknown (DNA-binding IC50 3 μM) | [27,28] |
| WalK/WalR | HK, RR | S. pneumoniae S. pyogenes S. epidermidis S. mutans E. faecalis B. subtilis S. aureus | Dimerization of HK, autophosphorylation of HK, phosphoryl transfer to RR or dimerization of the RR | Reduce bacterial growth (enzymatic inhibition IC50 37–62 μM, MIC 0.39–128 μg/mL or 8–16 μM) | [29,30,31,32] |
| CiaH/CiaR | HK | S. mutans | Autophosphorylation of HK | Decrease virulence (enzymatic inhibition IC50 4.9 μM, in vivo 0.63 μg/mL) | [33] |
| VicK/VicR | HK | S. mutans | Autophosphorylation of HK | Decrease virulence (enzymatic inhibition IC50 2.9 μM, in vivo 0.63 μg/mL) | [33] |
| LiaS/LiaR | HK | S. mutans | Autophosphorylation of HK | Decrease virulence (enzymatic inhibition IC50 5.6 μM, in vivo 0.63 μg/mL) | [33] |
| HpkA/DrrA | HK | T. maritima | Autophosphorylation of HK | Unknown (enzymatic inhibition IC50 0.4–2.3 μM) | [34] |
| Fur | Activator and repressor | E. coli | Dimerization and/or DNA-binding | Decrease virulence (ND) | [35] |
| EnvZ/OmpR | HK | E. coli | Autophosphorylation of HK | Unknown (enzymatic inhibition IC50 1.2 μM) | [33] |
| PhoP/PhoQ | HK | E. coli | Autophosphorylation of HK | Unknown (enzymatic inhibition IC50 1.2 μM) | [33] |
| CheA/CheY | HK | E. coli | Autophosphorylation of HK | Unknown (enzymatic inhibition at 50 μg/mL) | [36] |
| NtrB/NtrC | HK | E. coli | Autophosphorylation of HK | Unknown (enzymatic inhibition at 50 μg/mL) | [36] |
| BasS/BasR | RR | E. coli | Phosphoryl transfer to RR | Reduce growth in specific conditions (growth inhibition at 50 μg/mL) | [37] |
| CreC/CreB | RR | E. coli | Phosphoryl transfer to RR | Reduce growth in specific conditions (growth inhibition at 50 μg/mL) | [37] |
| KinA/SpoOF | HK | B. subtilis | Autophosphorylation of HK | Unknown (enzymatic inhibition at 50 μg/mL) | [36,38] |
| AlgR2/AlgR1 | HK, RR | P. aeruginosa | Autophosphorylation and dephosphorylation of HK, phosphoryl transfer to RR, or DNA-binding | Unknown (enzymatic inhibition at 50 μg/mL) | [36] |
| PqsR | Activator | P. aeruginosa | Signal sensing | Reduce virulence factor production and biofilm formation (IC50 0.2–39 μM) | [39,40,41,42,43] |
| QseC/QseB QseC/QseF QseC/KdpE | HK | Many Gram-negative bacteria | Autophosphorylation of HK | Decrease virulence (20 mg/Kg in mice) | [44,45] |
| AgrC/AgrA | HK, RR | S. aureus | Signal sensing by HK or DNA-binding | Decrease virulence (IC50 10–90 nM, DNA-binding IC50 83 μM) | [46,47] |
| AgrC/AgrA | HK | S. epidermidis | Signal sensing by HK | Decrease virulence (IC50 2–50 nM) | [48] |
| ComD/ComE | HK | S. pneumoniae | Signal sensing by HK | Decrease virulence and horizontal gene transfer (IC50 86–670 nM, EC50 6–83 nM) | [49,50] |
| FsrC/FsrA | HK | E. faecalis | Signal sensing by HK | Reduce host invasion and virulence (IC50 0.026–5 μM) | [51] |
| BfmS/BfmR | RR | A. baumannii | Unknown | Reduce biofilm formation (IC50 10 μM) | [52] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roncarati, D.; Scarlato, V.; Vannini, A. Targeting of Regulators as a Promising Approach in the Search for Novel Antimicrobial Agents. Microorganisms 2022, 10, 185. https://doi.org/10.3390/microorganisms10010185
Roncarati D, Scarlato V, Vannini A. Targeting of Regulators as a Promising Approach in the Search for Novel Antimicrobial Agents. Microorganisms. 2022; 10(1):185. https://doi.org/10.3390/microorganisms10010185
Chicago/Turabian StyleRoncarati, Davide, Vincenzo Scarlato, and Andrea Vannini. 2022. "Targeting of Regulators as a Promising Approach in the Search for Novel Antimicrobial Agents" Microorganisms 10, no. 1: 185. https://doi.org/10.3390/microorganisms10010185
APA StyleRoncarati, D., Scarlato, V., & Vannini, A. (2022). Targeting of Regulators as a Promising Approach in the Search for Novel Antimicrobial Agents. Microorganisms, 10(1), 185. https://doi.org/10.3390/microorganisms10010185