Abstract
The chemical modification of cellular macromolecules by the transfer of ADP-ribose unit(s), known as ADP-ribosylation, is an ancient homeostatic and stress response control system. Highly conserved across the evolution, ADP-ribosyltransferases and ADP-ribosylhydrolases control ADP-ribosylation signalling and cellular responses. In addition to proteins, both prokaryotic and eukaryotic transferases can covalently link ADP-ribosylation to different conformations of nucleic acids, thus highlighting the evolutionary conservation of archaic stress response mechanisms. Here, we report several structural and functional aspects of DNA ADP-ribosylation modification controlled by the prototype DarT and DarG pair, which show ADP-ribosyltransferase and hydrolase activity, respectively. DarT/DarG is a toxin–antitoxin system conserved in many bacterial pathogens, for example in Mycobacterium tuberculosis, which regulates two clinically important processes for human health, namely, growth control and the anti-phage response. The chemical modulation of the DarT/DarG system by selective inhibitors may thus represent an exciting strategy to tackle resistance to current antimicrobial therapies.
1. Introduction
ADP-ribosylation is a reversible post-translational modification (PTM) found in all three domains of life, as well as in several viruses [1,2,3,4]. It was identified in the 1970s as a key enzymatic activity required for cholera and diphtheria toxin pathogenesis [5,6]. Since then, the understanding of ADP-ribosylation has increased, from bacteria to mammals. Today, it is mainly known in the scientific community for its key role in DNA damage repair [7,8,9,10] and for being the target of tailored cancer therapies [11,12,13,14]. However, the functions of ADP-ribosylation are also vital for controlling many additional physiological processes, such as transcription and translation [15,16,17,18,19,20,21], cell proliferation [22,23], and cell death [24,25,26] along with stress and immune responses [27,28,29,30,31,32,33] and many others [34,35,36]. The control of cell homeostasis in both prokaryotes and eukaryotes by ADP-ribosylation [19,34,35,37,38,39,40,41] makes this a PTM of great interest in many fields of human health.
ADP-ribosylation is characterised by the addition of ADP-ribose unit(s) from nicotinamide adenine dinucleotide (NAD+) onto cellular substrates with the release of nicotinamide [42,43]. Consistent with the number of ADP-ribose moieties attached, single or multiple, the reaction is further differentiated into mono-ADP-ribosylation (MARylation) and poly-ADP-ribosylation (PARylation), respectively [44]. It was originally discovered as a PTM mainly targeting proteins [45], but it can also be covalently attached to additional macromolecules, including DNA [38,46,47,48,49] and RNA [48,50], as well as to small molecules such as antibiotics [51], ATP, and ADP [36].
There is a balanced interplay between specialised enzymes, namely, ADP-ribosyltransferases and ADP-ribosylhydrolases, which are responsible for the attachment and removal of the modification from cellular targets, respectively. This interplay shapes ADP-ribosylation signalling [43,44,52,53,54]. The dysregulation of these enzymatic activities in humans thus has implications in the pathogenicity of several diseases, above all, neurological disorders [55,56], cancer [57,58], and bacterial- and viral-mediated infections, as discussed here [59,60,61,62].
ADP-ribosylation is currently known to be involved in a large number of infectious diseases worldwide [63,64,65,66], including COVID-19 [33,60,61,67,68,69,70,71,72], Legionnaires’ disease [73,74,75,76,77], and the infectious diseases caused by the virulent M. tuberculosis. From a pathogenic point of view, the mechanisms of ADP-ribosylation in M. tuberculosis infection involve the modification of their own/endogenous targets rather than the host proteins, ultimately enabling the cell to adapt within the host and to improve the biological fitness. Among these mechanisms, the activity of the DarT/DarG toxin–antitoxin (TA) ADP-ribosylation system in M. tuberculosis targets bacterial genomic DNA. As a result of DNA modification, ADP-ribosylation slows growth and potentially induces bacterial persistence, a phenotypic state that correlates with antibiotic resistance [38,40,78].
The excessive use of antibiotics to counteract pathogen infections has led to the spread of antibiotic resistant “superbugs”, which currently represent a major public health threat [79,80]. Antibiotic resistance occurs in a wide range of bacterial infections and is determined by the activation of pathogen resistance/defence mechanisms, which also enable the cells to become persistent and tolerant to antibiotics [81,82]. Bacterial TA systems are widespread in Gram-negative and Gram-positive bacteria, and are involved in cell regulatory mechanisms in response to stress stimuli [83,84,85], including antibiotic resistance. Targeting TA modules such as the ADP-ribosylation DarT/DarG system can thus act as a blueprint for designing alternative drugs to current therapeutic treatments of antibiotic-resistant pathogens [86,87].
In this perspective, we discuss the structural and mechanistic aspects of DarT/DarG toxin–antitoxin-mediated control of DNA ADP-ribosylation. In addition, we then address the pathogenic role of the DarT/DarG TA pair as well as the therapeutic perspectives that the modulation of this specific ADP-ribosylation signalling may have.
2. ADP-Ribosylation in Bacteria
2.1. NAD+-Dependent Endotoxins and Exotoxins Involved in ADP-Ribosylation Signalling
ADP-ribosylation sustains prokaryotic cells in both cell metabolic processes and pathogenic mechanisms through the activity of NAD+-dependent enzymes, namely, endotoxins and exotoxins, respectively (Figure 1A–C).
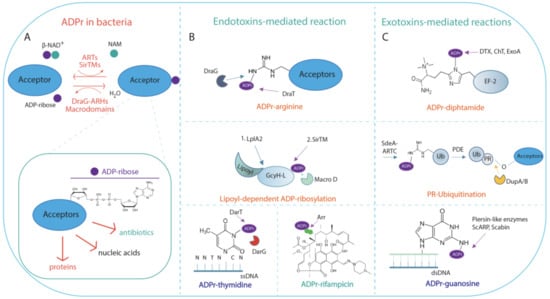
Figure 1.
Schematic representation of ADP-ribosylation reaction in bacterial meta-cell. (A) ADP-ribosylation (ADPr) reaction is catalysed by NAD+-dependent diphtheria toxin-like ARTs ARTDs, the cholera toxin-like ARTs ARTCs, or SirTMs, which transfer a single ADP-ribose unit on acceptors. Macrodomain-containing hydrolases (Macrodomains) or DraG-related ADP-ribosylhydrolases reverse the reaction by generating free ADP-ribose and unmodified acceptor. Inset: ADP-ribose moiety linked to acceptor substrates, which can be proteins, nucleic acids, or antibiotics. (B) Endotoxin-mediated reaction. Endotoxins can modify proteins, nucleic acids, or antibiotics. ARTDs, ARTCs, and SirTM modify endogenous bacterial substrates on different residues as indicated. ADP-ribosylated (ADPr)-arginine: MARylation of arginine residue is performed by the DraT enzyme, and is reversed by the cognate DraG ADP-ribosylhydrolase. Further examples are provided in the text. Lipoyl-dependent MARylation is carried out by SirTM and is dependent on prior lipoylation of the lipoyl-carrier protein GcyH-L, by the lipoate-protein ligase A (LplA2). The modification is reversed by the MacroD hydrolase, which is encoded within the same SirTM operon; ADP-ribosylated (ADPr)-thymidine: the reaction is performed by the endotoxin DarT that modifies thymidine base on ssDNA; the cognate DarG antitoxin reverses the modification; ADP-ribosylated (ADPr)-rifampicin: MARylation of the rifampin antibiotic is catalysed by Arr toxin. (C) Exotoxin-mediated reactions. ARTDs and ARTCs modify host targets on different residues as indicated. ADP-ribosylated (ADPr)-diphthamide: the reaction is catalysed by the toxins DTX, ChT, and ExoA, which irreversibly transfer ADP-ribose on the residue diphthamide on the elongation factor 2; PR-Ubiquitination. SdeA toxin catalyses the ADP-ribosylation (ADPr)-dependent ubiquitination of host proteins in a two-step reaction as detailed in the text. The reaction is reversed by the phosphodiesterases DubA/B; ADP-ribosylated (ADPr)-guanosine. The irreversible ADP-ribosylation on guanosine in dsDNA is performed by the pierisin-like enzymes ScARP and Scabin.
The majority of endotoxins and exotoxins belong to the superfamily of ADP-ribosyltransferase (ART) enzymes, which, despite limited sequence similarity, share a conserved secondary structure and protein fold [2,88]. ARTs fall into three phylogenetically distinct clades according to the catalytic triad composition: (i) the diphtheria toxin-like ARTs (ARTDs), characterised by the catalytic H-Y-[EDQ] triad; (ii) the cholera toxin-like ARTs (ARTCs), harbouring the R-S-E motif in the catalytic domain; and (iii) the tRNA 2’-phosphotransferase (TpT1/KptA) containing the H-H-h motif, with h containing any hydrophobic residues [2,43,52]. In addition to these subgroups, in Staphylococcus aureus and Streptococcus pyogenes, the microbial SirTMs, which belong to the Sirtuin superfamily of enzymes, catalyse the lipoyl-dependent ADP-ribosylation of proteins following a non-canonical deacylation reaction [89].
The substrate selectivity of ARTs is provided by two conserved functional motifs called the ARTT loop (ADP-ribosylating turn-turn), which is also known as the acceptor-loop (A-loop), and the donor-loop (D-loop). The D-loop is exclusive to ARTDs. Both loop structures are evolutionarily highly conserved, although their amino acid sequence and length vary greatly among the ARTDs [43]. In comparison with eukaryotic ARTs, bacterial enzymes show narrow amino acid residues specificity in host targets. Bacterial ARTs are in fact able to MARylate target proteins on several amino acid residues (i.e., arginine, cysteine, threonine, asparagine, and glutamine for ARTCs, diphthamide for ARTDs) (Figure 1B,C). Unlike some mammalian ART homologues (namely, PARP1 and PARP2) [90,91,92], bacterial ARTs do not need specificity factors [43]. In addition, bacterial ADP-ribosyltransferases can also modify nucleic acids (Figure 1B,C) [38,93]. Some bacterial ARTs, such as the ARTD homologue Arr-ms from Mycobacterium smegmatis, can also ADP-ribosylate the hydroxyl group at C23 of rifamycin and derivatives, thus inactivating antibiotics [51,94] (Figure 1B).
ADP-ribosylation is a reversible PTM. Two structurally distinct families of ADP-ribosylhydrolases, namely, DraG-related ADP-ribosylhydrolases (ARHs) and macrodomain-containing ADP-ribosylhydrolases, revert ADP-ribosylation signalling in bacterial cells (Figure 1A) [4].
DraG-related ARHs, from the founder DraG protein found in the nitrogen-fixing bacterial species R. rubrum and A. brasilense [37,95], are compact and globular modules with a typical domain length of 290–360 residues, with a central core motif comprising 13 orthogonal α-helices and a variable number of supplementary helices depending on the organism and type. The divalent metal ions enable the correct positioning of the substrate in the active site [53]. Structural studies on R. rubrum DraG hydrolase show that the de-MARylation of substrates occurs via the opening of the ribose ring and the formation of a protonated Schiff base. This substrate opening leads to a shift in metal coordination, allowing a nucleophilic attack by a water molecule activated by Mn2+ and resulting in a tetrahedral intermediate. The proton transfer via D97 promotes intermediate collapse and the release of arginine [96].
Macrodomain-containing hydrolases, harbouring the ADP-ribose-binding domain known as the macrodomain, share a conserved α/β/α fold consisting of a six-stranded mixed β-sheet surrounded by five α-helices [53]. Substrate binding takes place in a deep cleft on the top of the domain and several conserved interactions contribute to stabilise the ligand–macrodomain complex [53,54]. Based on ADP-ribosylhydrolase activity, macrodomains are further classified into mono-ADP-ribosylhydrolases (including MacroD-type and ALC1-like enzymes) and poly-ADP-ribosylhydrolases (PARG). Several bacterial macrodomains have been characterised as belonging to MacroD-, ALC1-, and PARG-like phylogenetically distinct groups that regulate a variety of cellular processes by deacetylating O-acetyl-ADP-ribose, and by hydrolysing MARylated targets, which include proteins and RNA [50,97,98]. In addition, several enzymes such as the TARG1-type macrodomain enzyme from Fusobacterium mortiferum ATCC 9817 [99] and the bacterial PARG from Thermomonospora curvata [97] have been reported to hydrolyse chains of ADP-ribose in vitro. The finding of an endogenous bacterial PARG-processing enzyme in Deinococcus radiodurans would seem to indicate an active prokaryotic PARylation machinery that may be involved in the stress response, given that PARG disruption leads to PAR enrichment in treated cells and loss of recovery after UV irradiation [100].
In a similar way to what happened for cancer treatment with the discovery of specific inhibitors of PARP1/PARP2 and PARG, the in-depth understanding of the enzymatic reactions and structural features of both bacterial ARTs and hydrolases promises important advances in antimicrobial therapies, which may eventually help to tackle antibiotic resistance.
2.2. Functional Aspects of ADP-Ribosylation in Bacteria
From a functional point of view, endotoxins modify endogenous targets, thus regulating the oxidative stress response [89], morphological differentiation and antibiotic production [101,102], and the maintenance of cell homeostasis in response to environmental stimuli, as exemplified by the Rhodospirillum rubrum and Azospirillum brasilense DraT/DraG system that regulates nitrogen fixation [37,95,96,103]. On the other hand, exotoxins promote pathogenic mechanisms through the transfer of ADP-ribose onto host targets, which alters signal transduction (e.g., CTX from Vibrio. cholerae; ETEC from Escherichia coli), cellular cytoskeleton organisation along with membrane trafficking (e.g., C2 toxin from Clostridium botulinum; ExoS toxin from Pseudomonas. aeruginosa), and protein synthesis (e.g., DTX from Corynebacterium diphtheriae; ChT from Vibrio cholerae) [4,45,62]. Bacterial exotoxins appear to be involved in the aetiology of important diseases [4,62,104]. Of these, SidE family effectors regulate the pathogenicity of Legionella pneumophila by non-canonical phosphoribosyl ubiquitination (Figure 1C), which interferes with the host physiological ubiquitination machinery [73,76,105,106], ultimately leading to the impairment of mitophagy and the secretory pathway [107]. The SdeA toxin, which is one of SidE family effectors released by the pathogenic L. pneumophila, catalyses the ADP-ribosylation-dependent ubiquitination of host proteins in a two-step reaction. Firstly, SdeA transfers the ADP-ribose on arginine 42 of a ubiquitin (Ub) molecule to generate an ADP-ribosylated-Ub intermediate due to the presence of an ARTC domain; the ADP-ribosylated-Ub intermediate is then converted to phosphoribosyl-Ub by the SdeA phosphodiesterase activity and is then conjugated through an ester linkage to a serine residue to target protein. Cognate phosphodiesterases DubA/B revert the reaction (Figure 1C).
Of particular interest from this perspective is that recent discoveries have established nucleic acids, such as genomic DNA and RNAs, as novel ADP-ribosylation targets [48,49,50], which, although involved in crucial physiological processes, are not yet fully understood in either mammals or prokaryotes [47,48,50,108]. To date, the ADP-ribosylation of DNA has only been characterised in a few bacterial systems including pierisin-like members and the DarT/DarG system. Pierisin and the pierisin-like ARTs represent a small group of ARTC toxins that prevalently ADP-ribosylate DNA. Pierisin, which is the founder of the family, has been identified in the cabbage butterfly species, Pieris rapae, where it counteracts the non-habitual parasitoids [109]. Extracts from P. rapae are highly cytotoxic toward insect and mammalian cells. In fact, pierisin induces irreversible host apoptosis by ADP-ribosylating N2 amino groups of 2′-deoxyguanosine into double-stranded DNA (dsDNA) in a non-conserved sequence manner, and as such, pierisin likely plays a role in the defence mechanism [110]. Similarly, the bacterial pierisin-like Scabin from Streptomyces scabies [111] and ScARP from Streptomyces coelicolor [112] are able to modify DNA on the exocyclic amino group on guanine bases and most of its derivatives in either single-stranded (ssDNA) or dsDNA. The disruption of ScARP affects S. coelicolor morphological differentiation, sporulation, and increased antibiotic production [101].
Unlike pierisin-like ARTCs, the DarT toxin from Thermus aquaticus and M. tuberculosis can specifically modify genomic ssDNA on thymidine in the conserved nucleotide sequence NNTNTCN, which can strongly hinder bacterial cell growth and, in turn, can have implications in antimicrobial susceptibility. Importantly, the cellular effects on the bacterial growth induced by DarT can be neutralised by DarG hydrolase, which, by reverting DNA-ADP-ribosylation, acts as an antitoxin [38]. Section 3 details the DarT/DarG system.
3. The DarT/DarG ADP-Ribosylation-Dependent System
3.1. DarT Is a New PARP-Like Toxin and a Potential Molecular Target for Antimicrobial Therapy
ADP-ribosylation catalysed by DarT specifically targets the thymidine bases present in conserved ssDNA sequence NNTNTCN in T. aquaticus and TTTT/A in M. tuberculosis, respectively, thereby showing no activity on dsDNA and RNA or protein targets. Substrate specificity toward a thymidine base also takes place for DarT toxin homologues, as highlighted in enteropathogenic E. coli DarT, which shows a preference for TCT/TTT sequences by modifying the third thymidine base of these motifs [40]. From a structural point of view, DarT can be considered a divergent ART enzyme given that it lacks the canonical catalytic triad residues found in ARTD and ARTC bacterial endotoxins (Figure 2A). Compared to bacterial ARTDs and ARTCs, DarT is very variable in terms of primary structure and motifs in comparison with bacterial ARTDs and ARTCs (Figure 2A). Nevertheless, the overall ART fold is maintained, as revealed by 3D resolution structural insights into Thermus sp. 2.9 DarT [78]. In fact, DarT is a PARP-like enzyme [78], as also predicted by phylogenetically analyses, as it is closer to human ARTDs than bacterial ART counterparts (Figure 2B).
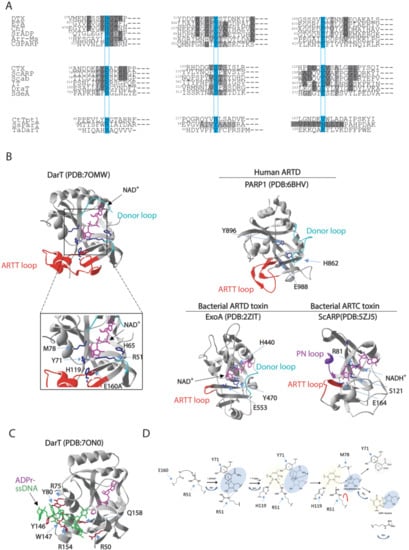
Figure 2.
Comparison of amino acid sequences and 3D structures of representative ARTs. (A) Alignment of the partial sequences of the bacterial ARTs. ARTD members, which harbour the H-Y-E catalytic residues, include: DTX, diphtheria toxin from C. diphtheriae; ETA, exotoxin A from P. aeruginosa; Ch toxin, cholix toxin from V. cholerae; SrADP, toxin from Streptomyces roseifaciens; Arr-Ms, rifamycin ADP-ribosylation toxin from Mycobacterium smegmatis; Cd-PARP, toxin from Clostridium perfringens CD 160. ARTC members, which enclose R-S-E catalytic residues, include: CTX, cholera toxin from V. cholerae; ScARP, toxin from S. coelicolor; Scabin from S. scabies; IT, iota toxin from C. perfringens; DraT, dinitrogenase reductase ADP-ribosyltransferase from R. rubrum; SdeA, ADP-ribosylation-dependent ubiquitination toxin from L. pneumophila. Divergent enzymes include: CtTpt1, Tpt1 RNA-phosphotransferase enzyme from Clostridium thermocellum; ParT, ADP-ribosylating toxin of ParT/ParS TA system from Sphingobium sp. YBL2; TaDarT, DNA ADP-ribosylating toxin of DarT/DarG TA system from T. aquaticus. The residues involved in catalysis are boxed on a light blue background. Identities or accepted amino acid substitutions are indicated in dark and light grey, respectively. (B) Cartoon–stick model of Thermus sp. 2.9 DarT(E160A) (PDB:7OMW, [78]) showing the NAD+ binding site in complex with NAD+, the amino acid residues involved in the catalytic activity (blue), the regulatory ARTT-loop (red) and the donor-loop (light blue). Inset: the catalytic site residues R51, H65, Y71, M78, H119, and E160A localise in proximity of nicotinamide in the active site. Cartoon–stick models of the 3D structure of the human ARTD PARP1 (PDB:6BHV, [113]), the bacterial ARTD-toxin ExoA (PDB:2ZIT, [114]), and the bacterial ARTC-toxin ScARP (PDB:5ZJ5, [115]) are shown as exemplars. (C) Cartoon–stick model of Thermus sp. 2.9 DarT(E160A) (PDB:7ON0, [78]) in complex with ADP-ribosylated ssDNA showing the residues (dark red) involved in the interaction with ssDNA (green). (D) Catalytic mechanism proposed for DarT-mediated ADP-ribosylation reaction of DNA.
Secondary structure elements are found conserved such as the fold of the central six-stranded b-sheet core and the helices between strands β1 and β2 (β1–2) and β2 and β3 (β2–3). Target DNA binds to a solvent-accessible channel placed orthogonally to the NAD+ molecule (Figure 2B,C) and is stabilised by the ARTT loop, which is known to affect substrate specificity in ARTDs as mentioned before. The length of the ARTT loop in DarT exceeds the ARTT loop of bacterial ARTDs and is instead comparable to loops found in human ARTDs, including PARP1, PARP2, and PARP3, thus forming a stable scaffold for the DNA target.
Given that the ARTT loop is found conserved in human ARTDs and that several ARTDs also ADP-ribosylate DNA [47,116,117,118], it is tempting to speculate that the ARTT loop plays a role in the interaction with the DNA target. The DNA binding site is located in a groove enriched with basic amino acid residues that enable the formation of a series of interactions that serve the sequence-specific ADP-ribosylation by DarT, with thymidine targeted for ADP-ribosylation pointing orthogonally to the DNA backbone deep inside the active site of DarT. Additional interactions between the DNA fragment and DarT side chains and main chains, in addition to structural waters, stabilise the phosphate–ribose backbone. DarT does not exhibit any NADase activity or auto-ADP-ribosylation activity and shows a distinct catalytic mechanism in comparison with other ARTDs. DarT binds the NAD+ substrate within a large pocket with key interactions resulting in a binding mode of constrained conformation. The adenine moiety is stabilised by hydrogen bonding to the K28 and L30 backbone amides, the adenine–proximal ribose bonds with its 2′ and 3′ hydroxyl groups to T15/H13 and N19, respectively. On the other hand, the NAM moiety is permanently maintained in position by π–π interaction with Y71 and hydrogen bonding of its primary amide to I14 and intra-molecularly to the beta-phosphate [78]. DarT shows a diverse arrangement of the catalytic site, wherein a key role is played by R51 residue, which expands the canonical ART catalytic motifs’ repertoire. ADP-ribosylation reaction occurs in several steps including: (1) locking of the thymidine base in plane for ADP-ribosylation by H119; (2) polarisation of the NAD+ molecule for cleavage sustained by Y71 and R51; (3) stabilisation of the oxocarbenium ion resulting from NAD+ cleavage by M78; and (4) proton abstraction from N3 of the thymidine base by R51. The latter step represents a new mechanism of ADP-ribosylation that has not been reported for other ARTs (Figure 3C). In fact, DarT-mediated ADP-ribosylation requires the presence of both R51 and E160 residues to perform the reaction, as NAD+ polarisation for cleavage is promoted by R51, which, when mutated, results in a loss of DarT cytotoxicity and enzymatic activity. This mechanism is different from the canonical NAD+ polarisation generally found in ARTs, where it is mediated by the interaction of the 2′′ hydroxyl group of the NAM ribose with the conserved catalytic glutamate residue. These data show that a new motif also supports the ADP-ribosylation reaction, which prompts the question as to whether DarT is an early version of ARTDs or a more evolved form that specialised in ADP-ribosylation of DNA. The advance in the understanding of such peculiar DarT enzymatic catalytic mechanisms will surely help in designing specific small molecules able to modulate DarT activity. This would represent an interesting molecular target for designing future antimicrobial strategies (please see Section 4).
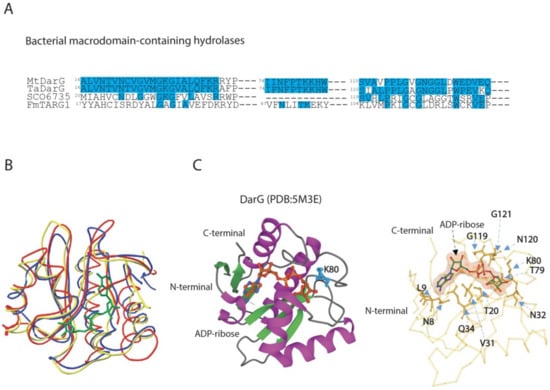
Figure 3.
Comparison of amino acid sequences and 3D structures of macrodomain-containing hydrolases belonging to ALC1-like group. (A) Alignment of partial sequences of ALC1-like hydrolases from bacteria. MtDarG, DarG from M. tuberculosis; TaDarG, DarG from T. aquaticus; SCO6735, macrodomain-containing hydrolase from S. coelicolor; FmTARG1, TARG1 homologue from F. mortiferum. Identities are indicated in light blue. (B) Structural comparison between DarG from T. aquaticus in complex with ADP-ribose (yellow line, PDB: 5M3E, [38]), human TARG1 in complex with ADP-ribose (blue line, PDB:4J5S, [55]) and SCO6735 (red line, PDB:5E3B, [119]). (C) DarG from T. aquaticus (cartoon) in complex with ADP-ribose (ball and stick). The catalytic K80 is shown in light blue (left panel). Close up of the T. aquaticus DarG active site showing the residues involved in ADP-ribose binding (right panel).
3.2. DarG Macrodomain-Containing Hydrolase Counteracts DarT Toxicity
The macrodomain-containing hydrolase DarG from T. aquaticus reverts DNA-ADP-ribosylation adduct on the thymidine base as observed in overexpressing E. coli cells with the consequent rescue of cell growth [38]. DarG antitoxins, which were characterised in T. aquaticus and M. tuberculosis, show a 56.4% sequence identity and a low sequence similarity to other bacterial macrodomain-containing hydrolases (Figure 3A).
DarG antitoxins share a resemblance to human terminal ADP-ribose glycohydrolase 1 (TARG1), and thus belong to the phylogenetically distinct ALC1-like sub-class of macrodomains. The ALC1-like macrodomain-containing enzymes bear similarity to the macrodomain fold found in the human chromatin-remodelling enzyme, ALC1 (Amplified Liver in Cancer1), which does not possess enzymatic activity, but interacts with PAR and catalyses PARP1-dependent nucleosome remodelling upon DNA damage [120]. From a functional point of view, members of the ALC1-like class display mono-ADP-ribosylhydrolase activity by hydrolysing the acyl-ADP-ribose ester bond by lysine residue, also exemplified by TARG1. The K84 nucleophilically attacks the C1’ atom of the distal ribose, leading to the formation of a lysyl-ADP-ribose intermediate with the release of the de-ADP-ribosylated E/D residue. The lysyl-intermediate is then resolved by residue D125, which frees the ADP-ribose, and restores the K84 residue [55].
The DarG macrodomain adopts a typical macrodomain fold composed of a six-stranded mixed β sheet arranged between four α helices and one 310-helical element. The ligand-binding pocket of the DarG macrodomain is made up of four surface loops where the bound ADP-ribose is located, and it interacts with neighbouring amino acid residues by forming hydrogen bonds (Figure 3B). The finding of W83 at the entrance of the cleft to stack with thymidine base for a correct position and K80 mostly involved in catalysis reflects the corresponding residues found in TARG1. The ligand-binding pocket is stabilised by the formation of hydrogen bonds. Key residues are found conserved, including K80, which is free to access the thymidine–ribose bond, and which is located in an equivalent position of K84 in TARG1. In fact, the mutation of K80A results in inactive DarG with a loss of hydrolase activity, similarly to the corresponding mutation on catalytic lysine in TARG1 [55]. However, the DarG catalytic mechanism remains unclear and needs further investigations.
The great similarity in the structural fold encountered between bacterial DarG and human TARG1 (Figure 3B) suggests that TARG1 plays a role in reversing ADP-ribosylation from DNA. Experimental evidence shows that the overexpression of DarT in human TARG1 knockout cell lines induces a strong DNA damage response due to replication fork progression arrest and cell death, and that the reintroduction of TARG1 activity is required for the reversal of DarT genotoxic effects. This indicates that TARG1 is the main macrodomain enzyme in human cells that acts as a DNA repair factor analogously to DarG [121].
Similar reversal activity has been described for the macrodomain hydrolase SCO6735, known for its regulatory role in antibiotic production in S. coelicolor [102]. SCO6735 has been identified as a functional homologue of DarG as it neutralises DarT activity by hydrolysing the thymidine-linked DNA-ADP-ribosylation [119]. Structural studies have shown that SCO6735 has a notable structural similarity to T. aquaticus DarG and human TARG1, even though TARG1 and SCO6735 also de-MARylate protein targets while DarG does not [119]. The overall macrodomain fold in SCO6735 is conserved (Figure 3B); the superimposition of the SCO6735 crystal structure with TARG1 and DarG in complex with ADP-ribose revealed a putative active site confined by three loops. The diphosphate and distal ribose are located between two loops, namely, the phosphate-binding (PB) and substrate-binding (SB) loop. The central loop in SCO6735 is five amino acids longer than DarG and TARG1 and provides Streptomyces hydrolase the ability to reverse ADP-ribose from thymidine-linked ADP-ribosylation and from aspartate/glutamate-linked proteins. The catalytic mechanism relies on the correct arrangement of the V25 and Q85 residues and a catalytic water molecule within the active site that sits between these residues and the diphosphate of the ADP-ribose. The mutation of Q85, located in an equivalent position to the catalytic lysine in DarG and TARG1 (K80 and K84, respectively), leads to a complete loss of activity. These observations suggest a diversification of catalytic reaction in this sub-class of macrodomain hydrolases, providing the rationale for the design of selective inhibitors or even agonists [119].
3.3. DarT/DarG TA System Molecular Mechanisms and Biological Functions
Bacterial genes encoding toxin and cognate antitoxin are frequently organised into operons, whose gene expression is regulated at a transcriptional and translational level. Under certain physiological conditions, the antitoxin protects the cell from the harmful effects of the toxin through a blockade or neutralising toxin activity [83,122,123]. Under stress conditions, the toxin is released and free to specifically impair one or more of several different cell events including DNA replication, translation, cytoskeleton formation, and membrane integrity [85,124,125]. More than 10,000 putative TA systems have been predicted by bioinformatic analyses [126,127], which can be classified into different types based on the nature of the antitoxin (non-coding RNA or protein) and on the interaction mode between the toxin and antitoxin components (Table 1) [85,125].

Table 1.
Classification of TA systems with the related targets and affected cellular functions.
In type II TA systems, the toxin effects are mainly counteracted by the direct binding of antitoxin to cognate toxin through protein–protein interaction, forming a stable toxin–antitoxin complex [83,87,137]. As summarised in Table 2, type II toxins include endoribonucleases that target mRNA, rRNA, and tRNA; ribosome-poisoning protein acetyltransferases that target tRNAs; topoisomerase inhibitors; cell wall inhibitors; and enzymes generating PTMs that target a diverse array of cellular targets, with the majority of them involved in the downregulation of cell metabolism [138,139].
Table 2.
Bacterial toxins displaying post-translational modification activity in type II TA systems.
ADP-ribosylation is a new player in TA biology. DarT/DarG TA was initially ascribed to type II, but it is now recognised as a type II/IV hybrid system, as DarT toxicity is mainly abrogated by DarG enzymatic activity by removing the DNA-ADP-ribose adduct rather than by TA complex assembly, as detailed below.
DarT catalyses the MARylation of NNTNTCN and TTTT/A motifs in ssDNA genomic sequences in T. aquaticus and M. tuberculosis, respectively. This enzymatic activity results in the formation of a thymidine ADP-ribose adduct that is recognised by the DNA damage repair system as a DNA lesion [38,78] (Figure 4A). It seems that M. tuberculosis uses this system to introduce adducts at the origin of DNA replication (OriC), which controls replication and cell growth. DarT highly ADP-ribosylates genomic DNA in DarG-depleted M. tuberculosis cells, leading to the activation of DNA damage response (Figure 4A). As a final outcome, the recruitment of DNAB, the replicative helicase interacting with ssDNA at the OriC and driving DNA branch migration during replication, may be impaired at cell division [38,78].
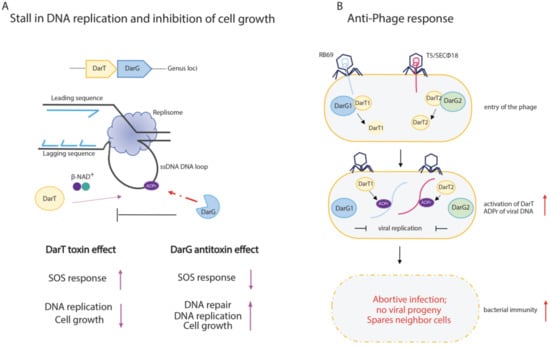
Figure 4.
Schematic representation of DarT/DarG TA system biological functions. (A) DarT/DarG system in the regulation of bacterial cell growth. DarT-mediated ADP-ribosylation of ssDNA on thymidine found in consensus sequences causes a stall of DNA replication and concomitant arrest of cell growth. The activity of the DarG antitoxin counteracts DarT activity through the removal of ADP-ribose from the marked thymidine on ssDNA: DarG-mediated removal of ADP-ribose enables the replication to proceed and cell growth to re-establish. (B) The DarT/DarG system and the anti-phage response. Upon entry of the phages, the DarT1 and DarT2 endotoxins ADP-ribosylate viral DNA, which is unable to replicate. The overall downregulation of cell metabolic processes triggers the abortive infection programme, which leads to the host cell death and prevents viral progeny spreading in order to protect the bacterial cell population.
The ADP-ribosylation of genomic DNA can be counteracted by the DarG antitoxin, which reverts DNA-ADP-ribose adducts [38] (Figure 4A), thus acting as a non-canonical DNA repair factor specific for ADP-ribosyl-thymidine adducts and re-establishing bacterial cell replication. The exquisite specificity of DarG reversal has been confirmed by in vitro experiments, where human macrodomain-containing hydrolases such as MacroD1 or PARG and DraG-related ADP-ribosylhydrolase ARH3 were unable to remove ADP-ribose from genomic DNA [78]. Further investigations support the protective role of DarG against DarT-mediated toxic effects, as the activation of the DNA damage response leads to cell death in M. tuberculosis DarG-depleted cells [153]. The same molecular mechanisms are shared by DarT/DarG TA systems from other species, such as in the enteropathogenic E. coli, where the ADP-ribosylation of genomic TCT or TTT DNA sequences can affect bacterial growth and viability [40]. Notably, complementation studies show that T. aquaticus DarG bears mutations in the hydrolase domain, namely, in the catalytic K80 residue, yet negatively affects DarT activity, thus suggesting that the antitoxin effect of DarG can also pass through additional mechanisms [38]. Consequently, the DarT/DarG TA pair can be considered a novel hybrid TA system.
In addition to the DarG antitoxin, the DarT-mediated DNA adducts can be also repaired by the sequential action of RecF-mediated homologous recombination, which leads to the conversion of a single-strand lesion into a double-strand lesion, which is then repaired by the nucleotide excision repair (NER) pathway [40].
The finding that the DarT/DarG system is also present in other bacterial species including the pathogenic P. aeruginosa, Acinetobacter baumannii, and K. pneumoniae [40] suggests a conserved role in pathogenic bacteria. However, the triggers that specifically induce DarT toxin activity remain unknown.
3.4. Functional Outcomes of DarT/DarG System in Prokaryotic Immunity
TA systems regulate several physiological processes including plasmid stabilisation and cell viability [154], persister cell formation [82,155], stress response [156], and biofilms [157] as well as multidrug resistance [86], pathogenicity [158], and defence from bacteriophages [139,159]. Overall, TA systems behave like versatile modulators of bacterial physiology that exploit the same biochemical mechanism to regulate a wide range of different activities.
With regard to phage defence, several bacterial strains harness diverse anti-phage defence systems relying on: (i) the degradation of phage nucleic acids by acting through restriction endonucleases and the CRISPR-Cas system; (ii) abortive infection-activating mechanisms that kill the bacterial population before phage replication; and (iii) the inhibition of DNA and RNA synthesis through the production of small molecules with inhibitory activity, which in turn sustains bacterial immunity [160,161].
The DarT/DarG TA system modulates the anti-phage response through the ADP-ribosylation of viral DNA and the consequent induction of the abortive infection mechanism [146] (Figure 4B). As already mentioned above, abortive infection is a well-conserved mechanism within bacterial immunity, and is widespread in Gram-positive and Gram-negative bacteria, where cell death takes place prior to the maturation of the phage progeny, thus preventing the spread of phages to neighbouring cells and protecting the bacterial population.
TA systems are known to have a pivotal role in the immunity of bacteria against phages, as their activation upon phage infection leads to cell death or growth arrest [131,162,163,164,165]. The role of ADP-ribosylation in bacterial immunity is exemplified by the DarT1/DarG1 and DarT2/DarG2 TA systems, which are found in the defence islands of the E. coli MG1655 bacterial genome, which are homologues of T. aquaticus DarT/DarG [146]. DarT toxin is conserved in both systems, whereas DarG1 and DarG2 belong to two different subfamilies; DarG1 encloses a putative YbiA-like fold, while DarG2 harbours a highly conserved macrodomain. Both DarT1/DarG1 and DarT2/DarG2 protect E. coli cells from natural bacteriophage infections, given that DarT1 and DarT2 are involved in ADP-ribosylation of viral DNA, with the consequent hindering of the phage’s genome replication, RNA synthesis, and assembly of mature/infective virions (Figure 4B).
DarT1/DarG1 and DarT2/DarG2 appear to target different phages (i.e., RB69 and T5/SEC 18, respectively) and are active under different growing conditions (DarTG1 during fast growth, DarTG2 during slow growth). These findings may suggest that a different regulatory mechanism activates the two DarT/DarG systems [146]. The molecular mechanism that unleashes the DarT toxin remains elusive; yet, though a still-unknown phage-derived trigger that frees the DarT toxin to exert an anti-phage response may perhaps explain it. The finding of phage mutants that exhibit spontaneous resistance to this immunity system by interfering with DarT/DarG activity adds another layer of complexity to the bacteria–phage conflicts [146].
A more comprehensive understanding of DarT/DarG biology may also result in the rational design of selective phage-based therapies as an alternative to antibiotics for treating resistant pathogens by manipulating endogenous anti-phage responses [166,167]. As such, small molecules inhibiting DarT may be exploited to counteract bacterial defence mechanisms against phages, which, to date, represent the most real alternative to antibiotics.
4. Exploitation of DarT/DarG Biology for a Rational Design of Antimicrobial Agents
Antibiotic resistance and the recurrence of bacterial infections are two of the most urgent threats to future global public health, with implications for all areas of medicine [168]. Antibiotic treatment misuse in humans and animals has accelerated the generation of antibiotic-resistant bacterial strains. In addition, the lack of novel effective antibacterial compounds, also due to poor investment in antimicrobial research, has increased this concern [168,169]. In fact, in the past 15 years, only one new class of antibiotics against Gram-positive bacteria has been introduced into clinical practice. The majority of antibiotics on the market are based on existing drugs selected to overcome the resistance acquired by bacteria against their related compounds [170]. Therefore, in order to tackle antibiotic resistance and recurring infections, it is imperative to search for antibacterial agents that rely on innovative modes of action.
Current antibiotics mostly target bacterial enzymes, ribosomal RNA, cell wall construction, and cell membrane function. Despite being widely used for the treatment of diverse infectious diseases, antibiotic treatments are not effective enough to eradicate highly resistant pathogens such as those referred as to ESKAPE. These resistant pathogens include E. coli, S. aureus, K. pneumoniae, A. baumanii, P. aeruginosa, and Enterobacter species, which are considered a priority by the World Health Organization for the urgent need of alternative therapeutics to antibiotic treatments. Therefore, alternative approaches to eradicate bacterial infection have been explored to deliver new therapies with clinical utility [171].
Mono-ADP-ribosyltransferase toxins are produced by pathogenic bacteria to infect the host cell with the impairment of vital molecular pathways [62]. These exotoxins exploit the host intracellular NAD+ to accomplish bacterial infection, which, in turn, causes a decrease in the level of NAD+ in the host, resulting in energy store depletion, immune evasion, or cell death [172]. In addition, some pathogens can also modulate NAD+ metabolism to support their fitness through the activity of pathogenic-specific enzymes such as NADases, or by the modulation of the activity of host NAD+-dependent enzymes (i.e., Sirtuins, PARPs, and CD38) [172,173]. Very recently, the pharmacological inhibition of PARPs in patients affected by diabetes mellitus has been reported to decrease intracellular M. tuberculosis (Mtb) in human macrophages, identifying PARP targeting as a potential novel strategy for host-directed therapy against M. tuberculosis and possibly against other infectious diseases [174].
With regard to NAD+-targeting toxins, the therapeutic inhibition of NAD+-dependent reactions in bacteria is still in its infancy and mainly relies on the chemical modulation of the NAD+ interaction pocket within the ART domain in order to block enzymatic activity. In the last two decades, antimicrobial strategies against ADP-ribosylating toxins have been proposed given that they are expected to provide new drug targets to disarm antibiotic-resistant bacteria. Different strategies, starting from the combination of PARP inhibitors, have been tested on P. aeruginosa Exotoxin A, V. cholerae Cholix toxin, V. splendidus Vis toxin, S. scabies Scabin toxin, Bacillus cereus Certhrax toxin, Paenibacillus larvae C3larvin, and Plx2A ([175] and the references therein). Such strategies have been searched for using ARTD non-specific inhibitors such as PJ34 [176], largely known for targeting human ARTDs (i.e., PARPs), polyphenolic extracts [177], and small molecules from the screening of synthetic libraries [178,179,180]. These attempts have led to the identification of lead compounds that can be further modified and explored in drug design. A promising approach relies on the use of natural compounds from plant origin that can hinder bacterial infections [181,182].
Protein–protein interactions (PPIs) have emerged as promising drug targets [183,184,185,186] and intensive efforts have led to the clinical use of PPI modulators as next-generation therapeutics in cancer treatments [187]. With regard to infectious diseases, the treatment of HIV/AIDS with the antiretroviral drugs enfuvirtide and maraviroc, which target host–pathogen interactions, provides a successful example of PPI-based drugs [188]. Since PPIs naturally occur in bacteria and regulate a multitude of cellular processes, bacterial protein–protein interactions are considered to be good candidates as a target for antibiotic drug discovery; however, to date, they are still underexplored [189,190].
Toxin–antitoxin systems represent a substantial pool of interactions within bacteria [83,191] that can be exploited for the development of advanced antibiotics [189,190,191]. Diverse PPI-based approaches have led to the discovery of peptides and small-molecule compounds that interfere with PPIs in TA systems, with the impairment of translation, cell wall synthesis, and lipase activity. However, no inhibitors are currently used in clinics. Given that DarG counteracts DarT activity even through the formation of a DarT–DarG complex, a PPI-based approach may be also considered in order to interfere with DarT function.
From this perspective, we have discussed the recent advances in the regulatory role exerted by the DarT/DarG hybrid TA system in the control of cell growth and abortive infection, strictly relying on ADP-ribosylation signalling. Within this framework, targeting DarT activity may represent a valuable alternative strategy for the therapeutic treatment of highly resistant pathogenic bacteria, such as M. tuberculosis, by preventing persistence activation. In addition, the availability of high-resolution-structure DarT provides the rationale for designing selective drugs to use as antimicrobial agents with less daunting side effects for the host.
PARP inhibitors for the therapeutic manipulation of ADP-ribosylation have been proposed for a wide range of disorders both in human and animal models, including cardiovascular, inflammatory, autoimmune, and neurological disorders [57]. In contrast, targeting ADP-ribosylation as a therapeutic intervention to counteract infectious diseases and related antibiotic resistant bacterial strains has been much less explored, with the exception of viral-mediated disease, where ADP-ribosylation is emerging as a crucial process in host–pathogen conflicts [69,71]. The growing body of evidence for the critical role of NAD+ as a co-factor of enzymes involved in bacterial physiology and the pathogenic mechanism as well as in host–pathogen interactions, also including viral-mediated diseases, highlights the importance of investigating these molecular pathways in order to find novel therapeutic strategies.
5. Conclusions
Recent discoveries have established DNA and RNA as the novel ADP-ribosylated substrates [48,49,50]. In mammals, the reversible ADP-ribosylation of DNA is mediated by PARP1, PARP2, and PARP3, which can ADP-ribosylate phosphorylated DNA termini on ds-DNA in vitro following a phosphorylation-dependent ADP-ribosylation mechanism; however, the functional outcomes remain unknown [116,117,118,192].
Recently, the reversible PARP1-mediated PARylation of ssDNA that targets adenosine residues has also been identified both in vitro and in vivo, where it does not seem to be activated by DNA strand breaks [193]. Other human PARPs, such as TRPT1, PARP10, PARP11, PARP12, and PARP15, appear to target the 5’-phosphorylated end of single-stranded RNA in vitro [47,48,49,50], giving rise to a novel RNA capping mechanism.
Several PARPs also have a role in the antiviral response through the inhibition of the virus life cycle at different stages, from transcription to translation, as exemplified by PARP7 and PARP13, which are involved in the exosome degradation of viral RNAs, and PARP12, which is responsible for the impairment of viral translation through the direct binding of viral RNA, and by the downregulation of cellular processes such as translation ([194] and the references therein). The importance of ADP-ribosylation-dependent systems in the antiviral response is highlighted by the fact that several viruses, such as alphaviruses and coronaviruses, have evolved macrodomains to counter hosts’ defensive processes controlled by ADP-ribosylation [33,53,60,195,196,197,198]. Viral macrodomains represent potential targets of antiviral drugs [69,70,71,199,200,201]. The role of ADP-ribosylation in antiviral and stress response, for instance, involving the ADP-ribosylation of viral genetic material, are reminiscent of the DNA modifications observed in lower organisms, where the transfer of ADP-ribose on nucleic acids results in the defence mechanism’s response. DarT/DarG represents one of the most ancient ADP-ribosylation-dependent systems with a role in bacterial immunity (e.g., against viral infections) as well as in the stress response. The modulation of the DarT/DarG system may also help the design of new effective antimicrobial agents in this context.
Anti-phage defence mechanisms have been extensively studied. However, many aspects still need clarification [202]. Several molecular processes underlie the anti-phage defence, which is mostly based on the degradation of the viral genome (e.g., restriction/modification enzymes, CRISPR-Cas systems, Argonaute proteins), the inhibition of DNA and RNA synthesis (e.g., chemical defence, secondary metabolite, nucleotide depletion), and abortive infection [160]. Abortive infection takes place through several molecular mechanisms, which include CRISPR-Cas and TA systems, among others [160]. A new group of retrons, belonging to prokaryotic reverse transcriptases, have been characterised to confer resistance to a wide range of phages [203,204]. Intriguingly, they share a tripartite module organisation reminiscent of TA systems, and are composed of reverse transcriptase, a multicopy single-stranded DNA (msDNA) and RcaT, an additional element protein [205]. Retrons can also be potentially used in genome editing, as they catalyse the polymerisation of DNA from an RNA template [204].
The systems mentioned above are just a few examples of the great diversity of defence systems found or predicted in bacterial cells to resist phage attack. In fact, a large number of genes encoding for different defence systems are found on the bacterial genome within chromosome regions known as “defence islands”, some of which are estimated to contain more than 100 defence genes [202]. Overall, such co-localisation of different defence genes suggests a functional link between the defence systems, including a possible coregulation mechanism.
More than 10,000 TA systems have been found on bacterial genomes, with many bacterial species encoding dozens of TA modules. For instance, E. coli K12 and M. tuberculosis encode more than 30, and at least 80, different TA systems [146], thus suggesting that different molecular activities support TA systems in their functional outcomes. DarT/DarG represents the first TA system that induces the stress response by growth control and abortive infection by ADP-ribosylating host genomic DNA and viral DNA with the concomitant inhibition of host DNA replication and cell growth. More recently, a ParT/ParS TA system from Sphingobium sp. YBL2 was found to hinder nucleotide metabolism with the induction of a persistence state by ADP-ribosylating target protein [39], highlighting NAD+ as a key component for triggering the prokaryotic immune response [173].
The wide distribution of ADP-ribosylation systems across all domains of life highlights the importance of this modification throughout evolution [1,2,3,4,62,206,207]. Overall, we believe that our review highlights the emergence of a new and exciting research area in the ADP-ribosylation field with implications in the regulation of cellular functions still to be discovered.
Author Contributions
G.C. and L.P. conceived and co-wrote the manuscript. R.C. performed supporting studies and contributed to the preparation of the figures. All authors have read and agreed to the published version of the manuscript.
Funding
Work in the L.P. group is funded by the Ovarian Cancer Research Alliance (OCRA, Collaborative Research Development Grant, n.813369), the POR Campania FESR 2014/2020 (projects SATIN and RECOVER-COVID19), and by NUTRIAGE-CNR. G.C.’s research activity is supported by “One Health Basic and Translational Research Actions addressing Unmet Needs on Emerging Infectious Diseases—INFACT”—CUP B53C20040570005.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Ivan Ahel, William Dunn School of Pathology, University of Oxford; Giuseppe Manco, Institute of Biochemistry and Cell Biology, National Research Council of Italy (CNR); and Rosa Marina Melillo, Department of Molecular Medicine and Medical Biotechnology, University of Naples “Federico II” for the critical reading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Perina, D.; Mikoč, A.; Ahel, J.; Ćetković, H.; Žaja, R.; Ahel, I. Distribution of Protein Poly(ADP-Ribosyl)Ation Systems across All Domains of Life. DNA Repair 2014, 23, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Aravind, L.; Zhang, D.; de Souza, R.F.; Anand, S.; Iyer, L.M. The Natural History of ADP-Ribosyltransferases and the ADP-Ribosylation System. Curr. Top. Microbiol. Immunol. 2015, 384, 3–32. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, L.; Mikoč, A.; Ahel, I. ADP-Ribosylation: New Facets of an Ancient Modification. FEBS J. 2017, 284, 2932–2946. [Google Scholar] [CrossRef] [PubMed]
- Mikolčević, P.; Hloušek-Kasun, A.; Ahel, I.; Mikoč, A. ADP-Ribosylation Systems in Bacteria and Viruses. Comput. Struct. Biotechnol. J. 2021, 19, 2366–2383. [Google Scholar] [CrossRef]
- Jørgensen, R.; Purdy, A.E.; Fieldhouse, R.J.; Kimber, M.S.; Bartlett, D.H.; Merrill, A.R. Cholix Toxin, a Novel ADP-Ribosylating Factor from Vibrio Cholerae. J. Biol. Chem. 2008, 283, 10671–10678. [Google Scholar] [CrossRef]
- Collier, R.J.; Pappenheimer, A.M. Studies on the mode of action of diphtheria toxin. I. Phosphorylated intermediates in normal and intoxicated hela cells. J. Exp. Med. 1964, 120, 1007–1018. [Google Scholar] [CrossRef]
- D’Amours, D.; Desnoyers, S.; D’Silva, I.; Poirier, G.G. Poly(ADP-Ribosyl)Ation Reactions in the Regulation of Nuclear Functions. Biochem. J. 1999, 342, 249–268. [Google Scholar] [CrossRef]
- De Vos, M.; Schreiber, V.; Dantzer, F. The Diverse Roles and Clinical Relevance of PARPs in DNA Damage Repair: Current State of the Art. Biochem. Pharmacol. 2012, 84, 137–146. [Google Scholar] [CrossRef]
- Gupte, R.; Liu, Z.; Kraus, W.L. PARPs and ADP-Ribosylation: Recent Advances Linking Molecular Functions to Biological Outcomes. Genes Dev. 2017, 31, 101–126. [Google Scholar] [CrossRef]
- Schützenhofer, K.; Rack, J.G.M.; Ahel, I. The Making and Breaking of Serine-ADP-Ribosylation in the DNA Damage Response. Front. Cell Dev. Biol. 2021, 9, 745922. [Google Scholar] [CrossRef]
- Lord, C.J.; Ashworth, A. PARP Inhibitors: Synthetic Lethality in the Clinic. Science 2017, 355, 1152–1158. [Google Scholar] [CrossRef]
- Palazzo, L.; Ahel, I. PARPs in Genome Stability and Signal Transduction: Implications for Cancer Therapy. Biochem. Soc. Trans. 2018, 46, 1681–1695. [Google Scholar] [CrossRef]
- Slade, D. PARP and PARG Inhibitors in Cancer Treatment. Genes Dev. 2020, 34, 360–394. [Google Scholar] [CrossRef]
- Curtin, N.J.; Szabo, C. Poly(ADP-Ribose) Polymerase Inhibition: Past, Present and Future. Nat. Rev. Drug Discov. 2020, 19, 711–736. [Google Scholar] [CrossRef]
- Krishnakumar, R.; Kraus, W.L. PARP-1 Regulates Chromatin Structure and Transcription through a KDM5B-Dependent Pathway. Mol. Cell 2010, 39, 736–749. [Google Scholar] [CrossRef]
- Gibson, B.A.; Zhang, Y.; Jiang, H.; Hussey, K.M.; Shrimp, J.H.; Lin, H.; Schwede, F.; Yu, Y.; Kraus, W.L. Chemical Genetic Discovery of PARP Targets Reveals a Role for PARP-1 in Transcription Elongation. Science 2016, 353, 45–50. [Google Scholar] [CrossRef]
- Kim, D.-S.; Camacho, C.V.; Nagari, A.; Malladi, V.S.; Challa, S.; Kraus, W.L. Activation of PARP-1 by SnoRNAs Controls Ribosome Biogenesis and Cell Growth via the RNA Helicase DDX21. Mol. Cell 2019, 75, 1270–1285.e14. [Google Scholar] [CrossRef]
- Gupte, R.; Nandu, T.; Kraus, W.L. Nuclear ADP-Ribosylation Drives IFNγ-Dependent STAT1α Enhancer Formation in Macrophages. Nat. Commun. 2021, 12, 3931. [Google Scholar] [CrossRef]
- Challa, S.; Khulpateea, B.R.; Nandu, T.; Camacho, C.V.; Ryu, K.W.; Chen, H.; Peng, Y.; Lea, J.S.; Kraus, W.L. Ribosome ADP-Ribosylation Inhibits Translation and Maintains Proteostasis in Cancers. Cell 2021, 184, 4531–4546.e26. [Google Scholar] [CrossRef]
- Jones, A.; Kraus, W.L. Multiomics Analysis of the NAD+-PARP1 Axis Reveals a Role for Site-Specific ADP-Ribosylation in Splicing in Embryonic Stem Cells. Genes Dev. 2022, 36, 601–617. [Google Scholar] [CrossRef]
- Manco, G.; Lacerra, G.; Porzio, E.; Catara, G. ADP-Ribosylation Post-Translational Modification: An Overview with a Focus on RNA Biology and New Pharmacological Perspectives. Biomolecules 2022, 12, 443. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.; Coughlin, M.; Mitchison, T.J. Tankyrase-1 Polymerization of Poly(ADP-Ribose) Is Required for Spindle Structure and Function. Nat. Cell Biol. 2005, 7, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Boehler, C.; Gauthier, L.R.; Mortusewicz, O.; Biard, D.S.; Saliou, J.-M.; Bresson, A.; Sanglier-Cianferani, S.; Smith, S.; Schreiber, V.; Boussin, F.; et al. Poly(ADP-Ribose) Polymerase 3 (PARP3), a Newcomer in Cellular Response to DNA Damage and Mitotic Progression. Proc. Natl. Acad. Sci. USA 2011, 108, 2783–2788. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Schreek, S.; Cerni, C.; Schamberger, C.; Lesniewicz, K.; Poreba, E.; Vervoorts, J.; Walsemann, G.; Grötzinger, J.; Kremmer, E.; et al. PARP-10, a Novel Myc-Interacting Protein with Poly(ADP-Ribose) Polymerase Activity, Inhibits Transformation. Oncogene 2005, 24, 1982–1993. [Google Scholar] [CrossRef] [PubMed]
- Andrabi, S.A.; Kang, H.C.; Haince, J.-F.; Lee, Y.-I.; Zhang, J.; Chi, Z.; West, A.B.; Koehler, R.C.; Poirier, G.G.; Dawson, T.M.; et al. Iduna Protects the Brain from Glutamate Excitotoxicity and Stroke by Interfering with Poly(ADP-Ribose) Polymer-Induced Cell Death. Nat. Med. 2011, 17, 692–699. [Google Scholar] [CrossRef]
- Bachmann, S.B.; Frommel, S.C.; Camicia, R.; Winkler, H.C.; Santoro, R.; Hassa, P.O. DTX3L and ARTD9 Inhibit IRF1 Expression and Mediate in Cooperation with ARTD8 Survival and Proliferation of Metastatic Prostate Cancer Cells. Mol. Cancer 2014, 13, 125. [Google Scholar] [CrossRef]
- Leung, A.K.L.; Vyas, S.; Rood, J.E.; Bhutkar, A.; Sharp, P.A.; Chang, P. Poly(ADP-Ribose) Regulates Stress Responses and MicroRNA Activity in the Cytoplasm. Mol. Cell 2011, 42, 489–499. [Google Scholar] [CrossRef]
- Catara, G.; Grimaldi, G.; Schembri, L.; Spano, D.; Turacchio, G.; Lo Monte, M.; Beccari, A.R.; Valente, C.; Corda, D. PARP1-Produced Poly-ADP-Ribose Causes the PARP12 Translocation to Stress Granules and Impairment of Golgi Complex Functions. Sci. Rep. 2017, 7, 14035. [Google Scholar] [CrossRef]
- Guo, X.; Ma, J.; Sun, J.; Gao, G. The Zinc-Finger Antiviral Protein Recruits the RNA Processing Exosome to Degrade the Target MRNA. Proc. Natl. Acad. Sci. USA 2007, 104, 151–156. [Google Scholar] [CrossRef]
- Atasheva, S.; Frolova, E.I.; Frolov, I. Interferon-Stimulated Poly(ADP-Ribose) Polymerases Are Potent Inhibitors of Cellular Translation and Virus Replication. J. Virol. 2014, 88, 2116–2130. [Google Scholar] [CrossRef]
- Zhang, Y.; Mao, D.; Roswit, W.T.; Jin, X.; Patel, A.C.; Patel, D.A.; Agapov, E.; Wang, Z.; Tidwell, R.M.; Atkinson, J.J.; et al. PARP9-DTX3L Ubiquitin Ligase Targets Host Histone H2BJ and Viral 3C Protease to Enhance Interferon Signaling and Control Viral Infection. Nat. Immunol. 2015, 16, 1215–1227. [Google Scholar] [CrossRef]
- Kim, C.; Wang, X.-D.; Yu, Y. PARP1 Inhibitors Trigger Innate Immunity via PARP1 Trapping-Induced DNA Damage Response. eLife 2020, 9, e60637. [Google Scholar] [CrossRef]
- Alhammad, Y.M.O.; Fehr, A.R. The Viral Macrodomain Counters Host Antiviral ADP-Ribosylation. Viruses 2020, 12, 384. [Google Scholar] [CrossRef]
- Abd Elmageed, Z.Y.; Naura, A.S.; Errami, Y.; Zerfaoui, M. The Poly(ADP-Ribose) Polymerases (PARPs): New Roles in Intracellular Transport. Cell. Signal. 2012, 24, 1–8. [Google Scholar] [CrossRef]
- Cardamone, M.D.; Gao, Y.; Kwan, J.; Hayashi, V.; Sheeran, M.; Xu, J.; English, J.; Orofino, J.; Emili, A.; Perissi, V. Neuralized-like Protein 4 (NEURL4) Mediates ADP-Ribosylation of Mitochondrial Proteins. J. Cell Biol. 2022, 221, e202101021. [Google Scholar] [CrossRef]
- Jia, A.; Huang, S.; Song, W.; Wang, J.; Meng, Y.; Sun, Y.; Xu, L.; Laessle, H.; Jirschitzka, J.; Hou, J.; et al. TIR-Catalyzed ADP-Ribosylation Reactions Produce Signaling Molecules for Plant Immunity. Science 2022, 377, eabq8180. [Google Scholar] [CrossRef]
- Nordlund, S.; Högbom, M. ADP-Ribosylation, a Mechanism Regulating Nitrogenase Activity. FEBS J. 2013, 280, 3484–3490. [Google Scholar] [CrossRef]
- Jankevicius, G.; Ariza, A.; Ahel, M.; Ahel, I. The Toxin-Antitoxin System DarTG Catalyzes Reversible ADP-Ribosylation of DNA. Mol. Cell 2016, 64, 1109–1116. [Google Scholar] [CrossRef]
- Piscotta, F.J.; Jeffrey, P.D.; Link, A.J. ParST Is a Widespread Toxin-Antitoxin Module That Targets Nucleotide Metabolism. Proc. Natl. Acad. Sci. USA 2019, 116, 826–834. [Google Scholar] [CrossRef]
- Lawarée, E.; Jankevicius, G.; Cooper, C.; Ahel, I.; Uphoff, S.; Tang, C.M. DNA ADP-Ribosylation Stalls Replication and Is Reversed by RecF-Mediated Homologous Recombination and Nucleotide Excision Repair. Cell Rep. 2020, 30, 1373–1384.e4. [Google Scholar] [CrossRef]
- Kong, L.; Feng, B.; Yan, Y.; Zhang, C.; Kim, J.H.; Xu, L.; Rack, J.G.M.; Wang, Y.; Jang, J.-C.; Ahel, I.; et al. Noncanonical Mono(ADP-Ribosyl)Ation of Zinc Finger SZF Proteins Counteracts Ubiquitination for Protein Homeostasis in Plant Immunity. Mol. Cell 2021, 81, 4591–4604.e8. [Google Scholar] [CrossRef]
- Ueda, K.; Hayaishi, O. ADP-Ribosylation. Annu. Rev. Biochem. 1985, 54, 73–100. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.S.; Chang, P. Insights into the Biogenesis, Function, and Regulation of ADP-Ribosylation. Nat. Chem. Biol. 2018, 14, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.; Matic, I.; Uchima, L.; Rood, J.; Zaja, R.; Hay, R.T.; Ahel, I.; Chang, P. Family-Wide Analysis of Poly(ADP-Ribose) Polymerase Activity. Nat. Commun. 2014, 5, 4426. [Google Scholar] [CrossRef] [PubMed]
- Krueger, K.M.; Barbieri, J.T. The Family of Bacterial ADP-Ribosylating Exotoxins. Clin. Microbiol. Rev. 1995, 8, 34–47. [Google Scholar] [CrossRef]
- Nakano, T.; Takahashi-Nakaguchi, A.; Yamamoto, M.; Watanabe, M. Pierisins and CARP-1: ADP-Ribosylation of DNA by ARTCs in Butterflies and Shellfish. Curr. Top. Microbiol. Immunol. 2015, 384, 127–149. [Google Scholar] [CrossRef]
- Munnur, D.; Ahel, I. Reversible Mono-ADP-Ribosylation of DNA Breaks. FEBS J. 2017, 284, 4002–4016. [Google Scholar] [CrossRef]
- Weixler, L.; Schäringer, K.; Momoh, J.; Lüscher, B.; Feijs, K.L.H.; Žaja, R. ADP-Ribosylation of RNA and DNA: From in Vitro Characterization to in Vivo Function. Nucleic Acids Res. 2021, 49, 3634–3650. [Google Scholar] [CrossRef]
- Groslambert, J.; Prokhorova, E.; Ahel, I. ADP-Ribosylation of DNA and RNA. DNA Repair 2021, 105, 103144. [Google Scholar] [CrossRef]
- Munnur, D.; Bartlett, E.; Mikolčević, P.; Kirby, I.T.; Rack, J.G.M.; Mikoč, A.; Cohen, M.S.; Ahel, I. Reversible ADP-Ribosylation of RNA. Nucleic Acids Res. 2019, 47, 5658–5669. [Google Scholar] [CrossRef]
- Baysarowich, J.; Koteva, K.; Hughes, D.W.; Ejim, L.; Griffiths, E.; Zhang, K.; Junop, M.; Wright, G.D. Rifamycin Antibiotic Resistance by ADP-Ribosylation: Structure and Diversity of Arr. Proc. Natl. Acad. Sci. USA 2008, 105, 4886–4891. [Google Scholar] [CrossRef]
- Hottiger, M.O.; Hassa, P.O.; Lüscher, B.; Schüler, H.; Koch-Nolte, F. Toward a Unified Nomenclature for Mammalian ADP-Ribosyltransferases. Trends Biochem. Sci. 2010, 35, 208–219. [Google Scholar] [CrossRef]
- Rack, J.G.M.; Perina, D.; Ahel, I. Macrodomains: Structure, Function, Evolution, and Catalytic Activities. Annu. Rev. Biochem. 2016, 85, 431–454. [Google Scholar] [CrossRef]
- Rack, J.G.M.; Palazzo, L.; Ahel, I. (ADP-Ribosyl)Hydrolases: Structure, Function, and Biology. Genes Dev. 2020, 34, 263–284. [Google Scholar] [CrossRef]
- Sharifi, R.; Morra, R.; Appel, C.D.; Tallis, M.; Chioza, B.; Jankevicius, G.; Simpson, M.A.; Matic, I.; Ozkan, E.; Golia, B.; et al. Deficiency of Terminal ADP-Ribose Protein Glycohydrolase TARG1/C6orf130 in Neurodegenerative Disease. EMBO J. 2013, 32, 1225–1237. [Google Scholar] [CrossRef]
- Liu, C.; Fang, Y. New Insights of Poly(ADP-Ribosylation) in Neurodegenerative Diseases: A Focus on Protein Phase Separation and Pathologic Aggregation. Biochem. Pharmacol. 2019, 167, 58–63. [Google Scholar] [CrossRef]
- Palazzo, L.; Mikolčević, P.; Mikoč, A.; Ahel, I. ADP-Ribosylation Signalling and Human Disease. Open Biol. 2019, 9, 190041. [Google Scholar] [CrossRef]
- Demény, M.A.; Virág, L. The PARP Enzyme Family and the Hallmarks of Cancer Part 1. Cell Intrinsic Hallmarks. Cancers 2021, 13, 2042. [Google Scholar] [CrossRef]
- Simon, N.C.; Aktories, K.; Barbieri, J.T. Novel Bacterial ADP-Ribosylating Toxins: Structure and Function. Nat. Rev. Microbiol. 2014, 12, 599–611. [Google Scholar] [CrossRef]
- Fehr, A.R.; Channappanavar, R.; Jankevicius, G.; Fett, C.; Zhao, J.; Athmer, J.; Meyerholz, D.K.; Ahel, I.; Perlman, S. The Conserved Coronavirus Macrodomain Promotes Virulence and Suppresses the Innate Immune Response during Severe Acute Respiratory Syndrome Coronavirus Infection. mBio 2016, 7, e01721-16. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.K.L.; McPherson, R.L.; Griffin, D.E. Macrodomain ADP-Ribosylhydrolase and the Pathogenesis of Infectious Diseases. PLoS Pathog. 2018, 14, e1006864. [Google Scholar] [CrossRef] [PubMed]
- Catara, G.; Corteggio, A.; Valente, C.; Grimaldi, G.; Palazzo, L. Targeting ADP-Ribosylation as an Antimicrobial Strategy. Biochem. Pharmacol. 2019, 167, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Fieldhouse, R.J.; Turgeon, Z.; White, D.; Merrill, A.R. Cholera- and Anthrax-like Toxins Are among Several New ADP-Ribosyltransferases. PLoS Comput. Biol. 2010, 6, e1001029. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Liu, C.; Shan, L.; He, P. Protein ADP-Ribosylation Takes Control in Plant-Bacterium Interactions. PLoS Pathog. 2016, 12, e1005941. [Google Scholar] [CrossRef] [PubMed]
- Toniti, W.; Yoshida, T.; Tsurumura, T.; Irikura, D.; Monma, C.; Kamata, Y.; Tsuge, H. Crystal Structure and Structure-Based Mutagenesis of Actin-Specific ADP-Ribosylating Toxin CPILE-a as Novel Enterotoxin. PLoS ONE 2017, 12, e0171278. [Google Scholar] [CrossRef]
- Belyy, A.; Lindemann, F.; Roderer, D.; Funk, J.; Bardiaux, B.; Protze, J.; Bieling, P.; Oschkinat, H.; Raunser, S. Mechanism of Threonine ADP-Ribosylation of F-Actin by a Tc Toxin. Nat. Commun. 2022, 13, 4202. [Google Scholar] [CrossRef]
- Rack, J.G.M.; Zorzini, V.; Zhu, Z.; Schuller, M.; Ahel, D.; Ahel, I. Viral Macrodomains: A Structural and Evolutionary Assessment of the Pharmacological Potential. Open Biol. 2020, 10, 200237. [Google Scholar] [CrossRef]
- Russo, L.C.; Tomasin, R.; Matos, I.A.; Manucci, A.C.; Sowa, S.T.; Dale, K.; Caldecott, K.W.; Lehtiö, L.; Schechtman, D.; Meotti, F.C.; et al. The SARS-CoV-2 Nsp3 Macrodomain Reverses PARP9/DTX3L-Dependent ADP-Ribosylation Induced by Interferon Signaling. J. Biol. Chem. 2021, 297, 101041. [Google Scholar] [CrossRef]
- Dasovich, M.; Zhuo, J.; Goodman, J.A.; Thomas, A.; McPherson, R.L.; Jayabalan, A.K.; Busa, V.F.; Cheng, S.-J.; Murphy, B.A.; Redinger, K.R.; et al. High-Throughput Activity Assay for Screening Inhibitors of the SARS-CoV-2 Mac1 Macrodomain. ACS Chem. Biol. 2022, 17, 17–23. [Google Scholar] [CrossRef]
- Leung, A.K.L.; Griffin, D.E.; Bosch, J.; Fehr, A.R. The Conserved Macrodomain Is a Potential Therapeutic Target for Coronaviruses and Alphaviruses. Pathog. Basel Switz. 2022, 11, 94. [Google Scholar] [CrossRef]
- Roy, A.; Alhammad, Y.M.; McDonald, P.; Johnson, D.K.; Zhuo, J.; Wazir, S.; Ferraris, D.; Lehtiö, L.; Leung, A.K.L.; Fehr, A.R. Discovery of Compounds That Inhibit SARS-CoV-2 Mac1-ADP-Ribose Binding by High-Throughput Screening. Antiviral Res. 2022, 203, 105344. [Google Scholar] [CrossRef]
- Zheng, M.; Schultz, M.B.; Sinclair, D.A. NAD+ in COVID-19 and Viral Infections. Trends Immunol. 2022, 43, 283–295. [Google Scholar] [CrossRef]
- Maculins, T.; Fiskin, E.; Bhogaraju, S.; Dikic, I. Bacteria-Host Relationship: Ubiquitin Ligases as Weapons of Invasion. Cell Res. 2016, 26, 499–510. [Google Scholar] [CrossRef]
- Bhogaraju, S.; Dikic, I. Cell Biology: Ubiquitination without E1 and E2 Enzymes. Nature 2016, 533, 43–44. [Google Scholar] [CrossRef]
- Akturk, A.; Wasilko, D.J.; Wu, X.; Liu, Y.; Zhang, Y.; Qiu, J.; Luo, Z.-Q.; Reiter, K.H.; Brzovic, P.S.; Klevit, R.E.; et al. Mechanism of Phosphoribosyl-Ubiquitination Mediated by a Single Legionella Effector. Nature 2018, 557, 729–733. [Google Scholar] [CrossRef]
- Kalayil, S.; Bhogaraju, S.; Bonn, F.; Shin, D.; Liu, Y.; Gan, N.; Basquin, J.; Grumati, P.; Luo, Z.-Q.; Dikic, I. Insights into Catalysis and Function of Phosphoribosyl-Linked Serine Ubiquitination. Nature 2018, 557, 734–738. [Google Scholar] [CrossRef]
- Dikic, I.; Schulman, B.A. An Expanded Lexicon for the Ubiquitin Code. Nat. Rev. Mol. Cell Biol. 2022, 25, 1–15. [Google Scholar] [CrossRef]
- Schuller, M.; Butler, R.E.; Ariza, A.; Tromans-Coia, C.; Jankevicius, G.; Claridge, T.D.W.; Kendall, S.L.; Goh, S.; Stewart, G.R.; Ahel, I. Molecular Basis for DarT ADP-Ribosylation of a DNA Base. Nature 2021, 596, 597–602. [Google Scholar] [CrossRef]
- Rice, L.B. The Clinical Consequences of Antimicrobial Resistance. Curr. Opin. Microbiol. 2009, 12, 476–481. [Google Scholar] [CrossRef]
- Aslam, B.; Khurshid, M.; Arshad, M.I.; Muzammil, S.; Rasool, M.; Yasmeen, N.; Shah, T.; Chaudhry, T.H.; Rasool, M.H.; Shahid, A.; et al. Antibiotic Resistance: One Health One World Outlook. Front. Cell. Infect. Microbiol. 2021, 11, 771510. [Google Scholar] [CrossRef]
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial Resistance in Bacteria: Mechanisms, Evolution, and Persistence. J. Mol. Evol. 2020, 88, 26–40. [Google Scholar] [CrossRef]
- Page, R.; Peti, W. Toxin-Antitoxin Systems in Bacterial Growth Arrest and Persistence. Nat. Chem. Biol. 2016, 12, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, Targets, and Triggers: An Overview of Toxin-Antitoxin Biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Wood, T.K. A Primary Physiological Role of Toxin/Antitoxin Systems Is Phage Inhibition. Front. Microbiol. 2020, 11, 1895. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Yadav, M.; Ghosh, C.; Rathore, J.S. Bacterial Toxin-Antitoxin Modules: Classification, Functions, and Association with Persistence. Curr. Res. Microb. Sci. 2021, 2, 100047. [Google Scholar] [CrossRef]
- Yang, Q.E.; Walsh, T.R. Toxin-Antitoxin Systems and Their Role in Disseminating and Maintaining Antimicrobial Resistance. FEMS Microbiol. Rev. 2017, 41, 343–353. [Google Scholar] [CrossRef]
- Równicki, M.; Lasek, R.; Trylska, J.; Bartosik, D. Targeting Type II Toxin–Antitoxin Systems as Antibacterial Strategies. Toxins 2020, 12, 568. [Google Scholar] [CrossRef]
- Holbourn, K.P.; Shone, C.C.; Acharya, K.R. A Family of Killer Toxins. Exploring the Mechanism of ADP-Ribosylating Toxins. FEBS J. 2006, 273, 4579–4593. [Google Scholar] [CrossRef]
- Rack, J.G.M.; Morra, R.; Barkauskaite, E.; Kraehenbuehl, R.; Ariza, A.; Qu, Y.; Ortmayer, M.; Leidecker, O.; Cameron, D.R.; Matic, I.; et al. Identification of a Class of Protein ADP-Ribosylating Sirtuins in Microbial Pathogens. Mol. Cell 2015, 59, 309–320. [Google Scholar] [CrossRef]
- Bonfiglio, J.J.; Fontana, P.; Zhang, Q.; Colby, T.; Gibbs-Seymour, I.; Atanassov, I.; Bartlett, E.; Zaja, R.; Ahel, I.; Matic, I. Serine ADP-Ribosylation Depends on HPF1. Mol. Cell 2017, 65, 932–940.e6. [Google Scholar] [CrossRef]
- Palazzo, L.; Leidecker, O.; Prokhorova, E.; Dauben, H.; Matic, I.; Ahel, I. Serine Is the Major Residue for ADP-Ribosylation upon DNA Damage. eLife 2018, 7, e34334. [Google Scholar] [CrossRef]
- Suskiewicz, M.J.; Zobel, F.; Ogden, T.E.H.; Fontana, P.; Ariza, A.; Yang, J.-C.; Zhu, K.; Bracken, L.; Hawthorne, W.J.; Ahel, D.; et al. HPF1 Completes the PARP Active Site for DNA Damage-Induced ADP-Ribosylation. Nature 2020, 579, 598–602. [Google Scholar] [CrossRef]
- Carpusca, I.; Jank, T.; Aktories, K. Bacillus Sphaericus Mosquitocidal Toxin (MTX) and Pierisin: The Enigmatic Offspring from the Family of ADP-Ribosyltransferases. Mol. Microbiol. 2006, 62, 621–630. [Google Scholar] [CrossRef]
- Quan, S.; Venter, H.; Dabbs, E.R. Ribosylative Inactivation of Rifampin by Mycobacterium Smegmatis Is a Principal Contributor to Its Low Susceptibility to This Antibiotic. Antimicrob. Agents Chemother. 1997, 41, 2456–2460. [Google Scholar] [CrossRef]
- Ma, Y.; Ludden, P.W. Role of the Dinitrogenase Reductase Arginine 101 Residue in Dinitrogenase Reductase ADP-Ribosyltransferase Binding, NAD Binding, and Cleavage. J. Bacteriol. 2001, 183, 250–256. [Google Scholar] [CrossRef]
- Berthold, C.L.; Wang, H.; Nordlund, S.; Högbom, M. Mechanism of ADP-Ribosylation Removal Revealed by the Structure and Ligand Complexes of the Dimanganese Mono-ADP-Ribosylhydrolase DraG. Proc. Natl. Acad. Sci. USA 2009, 106, 14247–14252. [Google Scholar] [CrossRef]
- Chen, D.; Vollmar, M.; Rossi, M.N.; Phillips, C.; Kraehenbuehl, R.; Slade, D.; Mehrotra, P.V.; von Delft, F.; Crosthwaite, S.K.; Gileadi, O.; et al. Identification of Macrodomain Proteins as Novel O-Acetyl-ADP-Ribose Deacetylases. J. Biol. Chem. 2011, 286, 13261–13271. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, C.; Song, Y.; Shao, C.; Zhang, X.; Zang, J. Structural Insights into the Mechanism of Escherichia Coli YmdB: A 2′-O-Acetyl-ADP-Ribose Deacetylase. J. Struct. Biol. 2015, 192, 478–486. [Google Scholar] [CrossRef]
- García-Saura, A.G.; Zapata-Pérez, R.; Hidalgo, J.F.; Sánchez-Ferrer, Á. Comparative Inhibitory Profile and Distribution of Bacterial PARPs, Using Clostridioides Difficile CD160 PARP as a Model. Sci. Rep. 2018, 8, 8056. [Google Scholar] [CrossRef]
- Cho, C.-C.; Chien, C.-Y.; Chiu, Y.-C.; Lin, M.-H.; Hsu, C.-H. Structural and Biochemical Evidence Supporting Poly ADP-Ribosylation in the Bacterium Deinococcus Radiodurans. Nat. Commun. 2019, 10, 1491. [Google Scholar] [CrossRef]
- Szirák, K.; Keserű, J.; Biró, S.; Schmelczer, I.; Barabás, G.; Penyige, A. Disruption of SCO5461 Gene Coding for a Mono-ADP-Ribosyltransferase Enzyme Produces a Conditional Pleiotropic Phenotype Affecting Morphological Differentiation and Antibiotic Production in Streptomyces Coelicolor. J. Microbiol. 2012, 50, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Lalić, J.; Posavec Marjanović, M.; Palazzo, L.; Perina, D.; Sabljić, I.; Žaja, R.; Colby, T.; Pleše, B.; Halasz, M.; Jankevicius, G.; et al. Disruption of Macrodomain Protein SCO6735 Increases Antibiotic Production in Streptomyces Coelicolor*. J. Biol. Chem. 2016, 291, 23175–23187. [Google Scholar] [CrossRef] [PubMed]
- Moure, V.R.; Costa, F.F.; Cruz, L.M.; Pedrosa, F.O.; Souza, E.M.; Li, X.-D.; Winkler, F.; Huergo, L.F. Regulation of Nitrogenase by Reversible Mono-ADP-Ribosylation. Curr. Top. Microbiol. Immunol. 2015, 384, 89–106. [Google Scholar] [CrossRef] [PubMed]
- Prygiel, M.; Polak, M.; Mosiej, E.; Wdowiak, K.; Formińska, K.; Zasada, A.A. New Corynebacterium Species with the Potential to Produce Diphtheria Toxin. Pathog. Basel Switz. 2022, 11, 1264. [Google Scholar] [CrossRef]
- Ashida, H.; Kim, M.; Sasakawa, C. Exploitation of the Host Ubiquitin System by Human Bacterial Pathogens. Nat. Rev. Microbiol. 2014, 12, 399–413. [Google Scholar] [CrossRef]
- Qiu, J.; Luo, Z.-Q. Hijacking of the Host Ubiquitin Network by Legionella Pneumophila. Front. Cell. Infect. Microbiol. 2017, 7, 487. [Google Scholar] [CrossRef]
- Liu, Y.; Mukherjee, R.; Bonn, F.; Colby, T.; Matic, I.; Glogger, M.; Heilemann, M.; Dikic, I. Serine-Ubiquitination Regulates Golgi Morphology and the Secretory Pathway upon Legionella Infection. Cell Death Differ. 2021, 28, 2957–2969. [Google Scholar] [CrossRef]
- Schuller, M.; Ahel, I. Beyond Protein Modification: The Rise of Non-Canonical ADP-Ribosylation. Biochem. J. 2022, 479, 463–477. [Google Scholar] [CrossRef]
- Watanabe, M.; Takamura-Enya, T.; Kanazawa, T.; Totsuka, Y.; Matsushima-Hibiya, Y.; Koyama, K.; Sugimura, T.; Wakabayashi, K. Mono(ADP-Ribosyl)Ation of DNA by Apoptosis-Inducing Protein, Pierisin. Nucleic Acids Res. Suppl. 2002, 2, 243–244. [Google Scholar] [CrossRef]
- Yamamoto, M.; Nakano, T.; Matsushima-Hibiya, Y.; Totsuka, Y.; Takahashi-Nakaguchi, A.; Matsumoto, Y.; Sugimura, T.; Wakabayashi, K. Molecular Cloning of Apoptosis-Inducing Pierisin-like Proteins, from Two Species of White Butterfly, Pieris Melete and Aporia Crataegi. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 154, 326–333. [Google Scholar] [CrossRef]
- Lyons, B.; Ravulapalli, R.; Lanoue, J.; Lugo, M.R.; Dutta, D.; Carlin, S.; Merrill, A.R. Scabin, a Novel DNA-Acting ADP-Ribosyltransferase from Streptomyces Scabies*. J. Biol. Chem. 2016, 291, 11198–11215. [Google Scholar] [CrossRef]
- Nakano, T.; Matsushima-Hibiya, Y.; Yamamoto, M.; Takahashi-Nakaguchi, A.; Fukuda, H.; Ono, M.; Takamura-Enya, T.; Kinashi, H.; Totsuka, Y. ADP-Ribosylation of Guanosine by SCO5461 Protein Secreted from Streptomyces Coelicolor. Toxicon 2013, 63, 55–63. [Google Scholar] [CrossRef]
- Langelier, M.-F.; Zandarashvili, L.; Aguiar, P.M.; Black, B.E.; Pascal, J.M. NAD+ Analog Reveals PARP-1 Substrate-Blocking Mechanism and Allosteric Communication from Catalytic Center to DNA-Binding Domains. Nat. Commun. 2018, 9, 844. [Google Scholar] [CrossRef]
- Jørgensen, R.; Wang, Y.; Visschedyk, D.; Merrill, A.R. The nature and character of the transition state for the ADP-ribosyltransferase reaction. EMBO Rep. 2008, 9, 802–809. [Google Scholar] [CrossRef]
- Yoshida, T.; Tsuge, H. Substrate N2 Atom Recognition Mechanism in Pierisin Family DNA-Targeting, Guanine-Specific ADP-Ribosyltransferase ScARP. J. Biol. Chem. 2018, 293, 13768–13774. [Google Scholar] [CrossRef]
- Talhaoui, I.; Lebedeva, N.A.; Zarkovic, G.; Saint-Pierre, C.; Kutuzov, M.M.; Sukhanova, M.V.; Matkarimov, B.T.; Gasparutto, D.; Saparbaev, M.K.; Lavrik, O.I.; et al. Poly(ADP-Ribose) Polymerases Covalently Modify Strand Break Termini in DNA Fragments in Vitro. Nucleic Acids Res. 2016, 44, 9279–9295. [Google Scholar] [CrossRef]
- Zarkovic, G.; Belousova, E.A.; Talhaoui, I.; Saint-Pierre, C.; Kutuzov, M.M.; Matkarimov, B.T.; Biard, D.; Gasparutto, D.; Lavrik, O.I.; Ishchenko, A.A. Characterization of DNA ADP-Ribosyltransferase Activities of PARP2 and PARP3: New Insights into DNA ADP-Ribosylation. Nucleic Acids Res. 2018, 46, 2417–2431. [Google Scholar] [CrossRef]
- Matta, E.; Kiribayeva, A.; Khassenov, B.; Matkarimov, B.T.; Ishchenko, A.A. Insight into DNA Substrate Specificity of PARP1-Catalysed DNA Poly(ADP-Ribosyl)Ation. Sci. Rep. 2020, 10, 3699. [Google Scholar] [CrossRef]
- Hloušek-Kasun, A.; Mikolčević, P.; Rack, J.G.M.; Tromans-Coia, C.; Schuller, M.; Jankevicius, G.; Matković, M.; Bertoša, B.; Ahel, I.; Mikoč, A. Streptomyces Coelicolor Macrodomain Hydrolase SCO6735 Cleaves Thymidine-Linked ADP-Ribosylation of DNA. Comput. Struct. Biotechnol. J. 2022, 20, 4337–4350. [Google Scholar] [CrossRef]
- Ahel, D.; Horejs, Ã.-Z.; Wiechens, N.; Polo, S.; Garcia-Wilson, E.; Ahel, I.; Flynn, H.; Skehel, M.; West, S.; Jackson, S.; et al. Poly(ADP-Ribose)-Dependent Regulation of DNA Repair by the Chromatin Remodeling Enzyme ALC1. Science 2009, 325, 1240–1243. [Google Scholar] [CrossRef]
- Tromans-Coia, C.; Sanchi, A.; Moeller, G.K.; Timinszky, G.; Lopes, M.; Ahel, I. TARG1 Protects against Toxic DNA ADP-Ribosylation. Nucleic Acids Res. 2021, 49, 10477–10492. [Google Scholar] [CrossRef] [PubMed]
- Leplae, R.; Geeraerts, D.; Hallez, R.; Guglielmini, J.; Drèze, P.; Van Melderen, L. Diversity of Bacterial Type II Toxin-Antitoxin Systems: A Comprehensive Search and Functional Analysis of Novel Families. Nucleic Acids Res. 2011, 39, 5513–5525. [Google Scholar] [CrossRef] [PubMed]
- Unterholzner, S.J.; Poppenberger, B.; Rozhon, W. Toxin-Antitoxin Systems: Biology, Identification, and Application. Mob. Genet. Elem. 2013, 3, e26219. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Pati, S.; Kaushik, H.; Singh, S.; Garg, L.C. Toxin-Antitoxin Systems and Their Medical Applications: Current Status and Future Perspective. Appl. Microbiol. Biotechnol. 2021, 105, 1803–1821. [Google Scholar] [CrossRef] [PubMed]
- Jurėnas, D.; Fraikin, N.; Goormaghtigh, F.; Van Melderen, L. Biology and Evolution of Bacterial Toxin-Antitoxin Systems. Nat. Rev. Microbiol. 2022, 20, 335–350. [Google Scholar] [CrossRef]
- Shao, Y.; Harrison, E.M.; Bi, D.; Tai, C.; He, X.; Ou, H.-Y.; Rajakumar, K.; Deng, Z. TADB: A Web-Based Resource for Type 2 Toxin-Antitoxin Loci in Bacteria and Archaea. Nucleic Acids Res. 2011, 39, D606–D611. [Google Scholar] [CrossRef]
- Akarsu, H.; Bordes, P.; Mansour, M.; Bigot, D.-J.; Genevaux, P.; Falquet, L. TASmania: A Bacterial Toxin-Antitoxin Systems Database. PLoS Comput. Biol. 2019, 15, e1006946. [Google Scholar] [CrossRef]
- Gerdes, K.; Bech, F.W.; Jørgensen, S.T.; Løbner-Olesen, A.; Rasmussen, P.B.; Atlung, T.; Boe, L.; Karlstrom, O.; Molin, S.; von Meyenburg, K. Mechanism of Postsegregational Killing by the Hok Gene Product of the ParB System of Plasmid R1 and Its Homology with the RelF Gene Product of the E. Coli RelB Operon. EMBO J. 1986, 5, 2023–2029. [Google Scholar] [CrossRef]
- Bernard, P.; Couturier, M. Cell Killing by the F Plasmid CcdB Protein Involves Poisoning of DNA-Topoisomerase II Complexes. J. Mol. Biol. 1992, 226, 735–745. [Google Scholar] [CrossRef]
- Castro-Roa, D.; Garcia-Pino, A.; De Gieter, S.; van Nuland, N.A.J.; Loris, R.; Zenkin, N. The Fic Protein Doc Uses an Inverted Substrate to Phosphorylate and Inactivate EF-Tu. Nat. Chem. Biol. 2013, 9, 811–817. [Google Scholar] [CrossRef]
- Fineran, P.C.; Blower, T.R.; Foulds, I.J.; Humphreys, D.P.; Lilley, K.S.; Salmond, G.P.C. The Phage Abortive Infection System, ToxIN, Functions as a Protein-RNA Toxin-Antitoxin Pair. Proc. Natl. Acad. Sci. USA 2009, 106, 894–899. [Google Scholar] [CrossRef]
- Masuda, H.; Tan, Q.; Awano, N.; Wu, K.-P.; Inouye, M. YeeU Enhances the Bundling of Cytoskeletal Polymers of MreB and FtsZ, Antagonizing the CbtA (YeeV) Toxicity in Escherichia Coli. Mol. Microbiol. 2012, 84, 979–989. [Google Scholar] [CrossRef]
- Wang, X.; Lord, D.M.; Cheng, H.-Y.; Osbourne, D.O.; Hong, S.H.; Sanchez-Torres, V.; Quiroga, C.; Zheng, K.; Herrmann, T.; Peti, W.; et al. A New Type V Toxin-Antitoxin System Where MRNA for Toxin GhoT Is Cleaved by Antitoxin GhoS. Nat. Chem. Biol. 2012, 8, 855–861. [Google Scholar] [CrossRef]
- Markovski, M.; Wickner, S. Preventing Bacterial Suicide: A Novel Toxin-Antitoxin Strategy. Mol. Cell 2013, 52, 611–612. [Google Scholar] [CrossRef]
- Marimon, O.; Teixeira, J.M.C.; Cordeiro, T.N.; Soo, V.W.C.; Wood, T.L.; Mayzel, M.; Amata, I.; García, J.; Morera, A.; Gay, M.; et al. An Oxygen-Sensitive Toxin-Antitoxin System. Nat. Commun. 2016, 7, 13634. [Google Scholar] [CrossRef]
- Choi, J.S.; Kim, W.; Suk, S.; Park, H.; Bak, G.; Yoon, J.; Lee, Y. The Small RNA, SdsR, Acts as a Novel Type of Toxin in Escherichia Coli. RNA Biol. 2018, 15, 1319–1335. [Google Scholar] [CrossRef]
- Fraikin, N.; Goormaghtigh, F.; Van Melderen, L. Type II Toxin-Antitoxin Systems: Evolution and Revolutions. J. Bacteriol. 2020, 202, e00763-19. [Google Scholar] [CrossRef]
- Zhang, S.-P.; Feng, H.-Z.; Wang, Q.; Kempher, M.L.; Quan, S.-W.; Tao, X.; Niu, S.; Wang, Y.; Feng, H.-Y.; He, Y.-X. Bacterial Type II Toxin-Antitoxin Systems Acting through Post-Translational Modifications. Comput. Struct. Biotechnol. J. 2021, 19, 86–93. [Google Scholar] [CrossRef]
- LeRoux, M.; Laub, M.T. Toxin-Antitoxin Systems as Phage Defense Elements. Annu. Rev. Microbiol. 2022, 76, 21–43. [Google Scholar] [CrossRef]
- Kaspy, I.; Rotem, E.; Weiss, N.; Ronin, I.; Balaban, N.Q.; Glaser, G. HipA-Mediated Antibiotic Persistence via Phosphorylation of the Glutamyl-TRNA-Synthetase. Nat. Commun. 2013, 4, 3001. [Google Scholar] [CrossRef]
- Germain, E.; Castro-Roa, D.; Zenkin, N.; Gerdes, K. Molecular Mechanism of Bacterial Persistence by HipA. Mol. Cell 2013, 52, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Vang Nielsen, S.; Turnbull, K.J.; Roghanian, M.; Bærentsen, R.; Semanjski, M.; Brodersen, D.E.; Macek, B.; Gerdes, K. Serine-Threonine Kinases Encoded by Split HipA Homologs Inhibit Tryptophanyl-TRNA Synthetase. mBio 2019, 10, e01138-19. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.; Stanger, F.V.; Scheu, P.D.; de Jong, I.G.; Goepfert, A.; Glatter, T.; Gerdes, K.; Schirmer, T.; Dehio, C. Adenylylation of Gyrase and Topo IV by FicT Toxins Disrupts Bacterial DNA Topology. Cell Rep. 2015, 12, 1497–1507. [Google Scholar] [CrossRef]
- Lu, C.; Nakayasu, E.S.; Zhang, L.-Q.; Luo, Z.-Q. Identification of Fic-1 as an Enzyme That Inhibits Bacterial DNA Replication by AMPylating GyrB, Promoting Filament Formation. Sci. Signal. 2016, 9, ra11. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.; Liesch, M.; Körner, J.; Québatte, M.; Engel, P.; Dehio, C. A Bacterial Toxin-Antitoxin Module Is the Origin of Inter-Bacterial and Inter-Kingdom Effectors of Bartonella. PLoS Genet. 2017, 13, e1007077. [Google Scholar] [CrossRef]
- LeRoux, M.; Srikant, S.; Teodoro, G.I.C.; Zhang, T.; Littlehale, M.L.; Doron, S.; Badiee, M.; Leung, A.K.L.; Sorek, R.; Laub, M.T. The DarTG Toxin-Antitoxin System Provides Phage Defence by ADP-Ribosylating Viral DNA. Nat. Microbiol. 2022, 7, 1028–1040. [Google Scholar] [CrossRef]
- Ting, S.-Y.; Bosch, D.E.; Mangiameli, S.M.; Radey, M.C.; Huang, S.; Park, Y.-J.; Kelly, K.A.; Filip, S.K.; Goo, Y.A.; Eng, J.K.; et al. Bifunctional Immunity Proteins Protect Bacteria against FtsZ-Targeting ADP-Ribosylating Toxins. Cell 2018, 175, 1380–1392.e14. [Google Scholar] [CrossRef]
- Freire, D.M.; Gutierrez, C.; Garza-Garcia, A.; Grabowska, A.D.; Sala, A.J.; Ariyachaokun, K.; Panikova, T.; Beckham, K.S.H.; Colom, A.; Pogenberg, V.; et al. An NAD+ Phosphorylase Toxin Triggers Mycobacterium Tuberculosis Cell Death. Mol. Cell 2019, 73, 1282–1291.e8. [Google Scholar] [CrossRef]
- Cheverton, A.M.; Gollan, B.; Przydacz, M.; Wong, C.T.; Mylona, A.; Hare, S.A.; Helaine, S. A Salmonella Toxin Promotes Persister Formation through Acetylation of TRNA. Mol. Cell 2016, 63, 86–96. [Google Scholar] [CrossRef]
- Jurėnas, D.; Garcia-Pino, A.; Van Melderen, L. Novel Toxins from Type II Toxin-Antitoxin Systems with Acetyltransferase Activity. Plasmid 2017, 93, 30–35. [Google Scholar] [CrossRef]
- Qian, H.; Yu, H.; Li, P.; Zhu, E.; Yao, Q.; Tai, C.; Deng, Z.; Gerdes, K.; He, X.; Gan, J.; et al. Toxin-Antitoxin Operon KacAT of Klebsiella Pneumoniae Is Regulated by Conditional Cooperativity via a W-Shaped KacA-KacT Complex. Nucleic Acids Res. 2019, 47, 7690–7702. [Google Scholar] [CrossRef]
- Wilcox, B.; Osterman, I.; Serebryakova, M.; Lukyanov, D.; Komarova, E.; Gollan, B.; Morozova, N.; Wolf, Y.I.; Makarova, K.S.; Helaine, S.; et al. Escherichia Coli ItaT Is a Type II Toxin That Inhibits Translation by Acetylating Isoleucyl-TRNAIle. Nucleic Acids Res. 2018, 46, 7873–7885. [Google Scholar] [CrossRef]
- Zaveri, A.; Wang, R.; Botella, L.; Sharma, R.; Zhu, L.; Wallach, J.B.; Song, N.; Jansen, R.S.; Rhee, K.Y.; Ehrt, S.; et al. Depletion of the DarG Antitoxin in Mycobacterium Tuberculosis Triggers the DNA-Damage Response and Leads to Cell Death. Mol. Microbiol. 2020, 114, 641–652. [Google Scholar] [CrossRef]
- Magnuson, R.D. Hypothetical Functions of Toxin-Antitoxin Systems. J. Bacteriol. 2007, 189, 6089–6092. [Google Scholar] [CrossRef]
- Korch, S.B.; Hill, T.M. Ectopic Overexpression of Wild-Type and Mutant HipA Genes in Escherichia Coli: Effects on Macromolecular Synthesis and Persister Formation. J. Bacteriol. 2006, 188, 3826–3836. [Google Scholar] [CrossRef]
- Gerdes, K.; Christensen, S.K.; Løbner-Olesen, A. Prokaryotic Toxin-Antitoxin Stress Response Loci. Nat. Rev. Microbiol. 2005, 3, 371–382. [Google Scholar] [CrossRef]
- Wang, X.; Wood, T.K. Toxin-Antitoxin Systems Influence Biofilm and Persister Cell Formation and the General Stress Response. Appl. Environ. Microbiol. 2011, 77, 5577–5583. [Google Scholar] [CrossRef]
- Wen, Y.; Behiels, E.; Devreese, B. Toxin-Antitoxin Systems: Their Role in Persistence, Biofilm Formation, and Pathogenicity. Pathog. Dis. 2014, 70, 240–249. [Google Scholar] [CrossRef]
- Qiu, J.; Zhai, Y.; Wei, M.; Zheng, C.; Jiao, X. Toxin-Antitoxin Systems: Classification, Biological Roles, and Applications. Microbiol. Res. 2022, 264, 127159. [Google Scholar] [CrossRef]
- Tal, N.; Sorek, R. SnapShot: Bacterial Immunity. Cell 2022, 185, 578–578.e1. [Google Scholar] [CrossRef]
- Tesson, F.; Hervé, A.; Mordret, E.; Touchon, M.; d’Humières, C.; Cury, J.; Bernheim, A. Systematic and Quantitative View of the Antiviral Arsenal of Prokaryotes. Nat. Commun. 2022, 13, 2561. [Google Scholar] [CrossRef] [PubMed]
- Koga, M.; Otsuka, Y.; Lemire, S.; Yonesaki, T. Escherichia Coli RnlA and RnlB Compose a Novel Toxin-Antitoxin System. Genetics 2011, 187, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Samson, J.E.; Spinelli, S.; Cambillau, C.; Moineau, S. Structure and Activity of AbiQ, a Lactococcal Endoribonuclease Belonging to the Type III Toxin-Antitoxin System. Mol. Microbiol. 2013, 87, 756–768. [Google Scholar] [CrossRef] [PubMed]
- Dy, R.L.; Przybilski, R.; Semeijn, K.; Salmond, G.P.C.; Fineran, P.C. A Widespread Bacteriophage Abortive Infection System Functions through a Type IV Toxin-Antitoxin Mechanism. Nucleic Acids Res. 2014, 42, 4590–4605. [Google Scholar] [CrossRef]
- Lopatina, A.; Tal, N.; Sorek, R. Abortive Infection: Bacterial Suicide as an Antiviral Immune Strategy. Annu. Rev. Virol. 2020, 7, 371–384. [Google Scholar] [CrossRef]
- Saha, D.; Mukherjee, R. Ameliorating the Antimicrobial Resistance Crisis: Phage Therapy. IUBMB Life 2019, 71, 781–790. [Google Scholar] [CrossRef]
- Xu, H.-M.; Xu, W.-M.; Zhang, L. Current Status of Phage Therapy against Infectious Diseases and Potential Application beyond Infectious Diseases. Int. J. Clin. Pract. 2022, 2022, 4913146. [Google Scholar] [CrossRef]
- Årdal, C.; Balasegaram, M.; Laxminarayan, R.; McAdams, D.; Outterson, K.; Rex, J.H.; Sumpradit, N. Antibiotic Development—Economic, Regulatory and Societal Challenges. Nat. Rev. Microbiol. 2020, 18, 267–274. [Google Scholar] [CrossRef]
- Machowska, A.; Stålsby Lundborg, C. Drivers of Irrational Use of Antibiotics in Europe. Int. J. Environ. Res. Public. Health 2018, 16, 27. [Google Scholar] [CrossRef]
- Lewis, K. The Science of Antibiotic Discovery. Cell 2020, 181, 29–45. [Google Scholar] [CrossRef]
- Czaplewski, L.; Bax, R.; Clokie, M.; Dawson, M.; Fairhead, H.; Fischetti, V.A.; Foster, S.; Gilmore, B.F.; Hancock, R.E.W.; Harper, D.; et al. Alternatives to Antibiotics—A Pipeline Portfolio Review. Lancet Infect. Dis. 2016, 16, 239–251. [Google Scholar] [CrossRef]
- Roussin, M.; Salcedo, S.P. NAD+-Targeting by Bacteria: An Emerging Weapon in Pathogenesis. FEMS Microbiol. Rev. 2021, 45, fuab037. [Google Scholar] [CrossRef]
- Mesquita, I.; Varela, P.; Belinha, A.; Gaifem, J.; Laforge, M.; Vergnes, B.; Estaquier, J.; Silvestre, R. Exploring NAD+ Metabolism in Host-Pathogen Interactions. Cell. Mol. Life Sci. CMLS 2016, 73, 1225–1236. [Google Scholar] [CrossRef]
- van Doorn, C.L.R.; Steenbergen, S.A.M.; Walburg, K.V.; Ottenhoff, T.H.M. Pharmacological Poly (ADP-Ribose) Polymerase Inhibitors Decrease Mycobacterium Tuberculosis Survival in Human Macrophages. Front. Immunol. 2021, 12, 712021. [Google Scholar] [CrossRef]
- Lugo, M.R.; Merrill, A.R. Development of Anti-Virulence Therapeutics against Mono-ADP-Ribosyltransferase Toxins. Toxins 2020, 13, 16. [Google Scholar] [CrossRef]
- Turgeon, Z.; Jørgensen, R.; Visschedyk, D.; Edwards, P.R.; Legree, S.; McGregor, C.; Fieldhouse, R.J.; Mangroo, D.; Schapira, M.; Merrill, A.R. Newly Discovered and Characterized Antivirulence Compounds Inhibit Bacterial Mono-ADP-Ribosyltransferase Toxins. Antimicrob. Agents Chemother. 2011, 55, 983–991. [Google Scholar] [CrossRef]
- Cherubin, P.; Garcia, M.C.; Curtis, D.; Britt, C.B.T.; Craft, J.W.; Burress, H.; Berndt, C.; Reddy, S.; Guyette, J.; Zheng, T.; et al. Inhibition of Cholera Toxin and Other AB Toxins by Polyphenolic Compounds. PLoS ONE 2016, 11, e0166477. [Google Scholar] [CrossRef]
- Yates, S.P.; Taylor, P.L.; Jørgensen, R.; Ferraris, D.; Zhang, J.; Andersen, G.R.; Merrill, A.R. Structure–Function Analysis of Water-Soluble Inhibitors of the Catalytic Domain of Exotoxin A from Pseudomonas Aeruginosa. Biochem. J. 2005, 385, 667–675. [Google Scholar] [CrossRef]
- Pinto, A.F.; Ebrahimi, M.; Saleeb, M.; Forsberg, Å.; Elofsson, M.; Schüler, H. Identification of Inhibitors of Pseudomonas Aeruginosa Exotoxin-S ADP-Ribosyltransferase Activity. SLAS Discov. 2016, 21, 590–595. [Google Scholar] [CrossRef]
- Saleeb, M.; Sundin, C.; Aglar, Ö.; Pinto, A.F.; Ebrahimi, M.; Forsberg, Å.; Schüler, H.; Elofsson, M. Structure–Activity Relationships for Inhibitors of Pseudomonas Aeruginosa Exoenzyme S ADP-Ribosyltransferase Activity. Eur. J. Med. Chem. 2018, 143, 568–576. [Google Scholar] [CrossRef]
- Wu, S.-C.; Liu, F.; Zhu, K.; Shen, J.-Z. Natural Products That Target Virulence Factors in Antibiotic-Resistant Staphylococcus Aureus. J. Agric. Food Chem. 2019, 67, 13195–13211. [Google Scholar] [CrossRef]
- Abdelhamid, A.G.; El-Dougdoug, N.K. Controlling Foodborne Pathogens with Natural Antimicrobials by Biological Control and Antivirulence Strategies. Heliyon 2020, 6, e05020. [Google Scholar] [CrossRef] [PubMed]
- Bogan, A.A.; Thorn, K.S. Anatomy of Hot Spots in Protein Interfaces. J. Mol. Biol. 1998, 280, 1–9. [Google Scholar] [CrossRef]
- Modell, A.E.; Blosser, S.L.; Arora, P.S. Systematic Targeting of Protein-Protein Interactions. Trends Pharmacol. Sci. 2016, 37, 702–713. [Google Scholar] [CrossRef]
- Scott, D.E.; Bayly, A.R.; Abell, C.; Skidmore, J. Small Molecules, Big Targets: Drug Discovery Faces the Protein-Protein Interaction Challenge. Nat. Rev. Drug Discov. 2016, 15, 533–550. [Google Scholar] [CrossRef]
- Cossar, P.J.; Lewis, P.J.; McCluskey, A. Protein-Protein Interactions as Antibiotic Targets: A Medicinal Chemistry Perspective. Med. Res. Rev. 2020, 40, 469–494. [Google Scholar] [CrossRef]
- Carro, L. Protein-Protein Interactions in Bacteria: A Promising and Challenging Avenue towards the Discovery of New Antibiotics. Beilstein J. Org. Chem. 2018, 14, 2881–2896. [Google Scholar] [CrossRef]
- Kramer, V.G.; Schader, S.M.; Oliveira, M.; Colby-Germinario, S.P.; Donahue, D.A.; Singhroy, D.N.; Tressler, R.; Sloan, R.D.; Wainberg, M.A. Maraviroc and Other HIV-1 Entry Inhibitors Exhibit a Class-Specific Redistribution Effect That Results in Increased Extracellular Viral Load. Antimicrob. Agents Chemother. 2012, 56, 4154–4160. [Google Scholar] [CrossRef]
- Kahan, R.; Worm, D.J.; de Castro, G.V.; Ng, S.; Barnard, A. Modulators of Protein-Protein Interactions as Antimicrobial Agents. RSC Chem. Biol. 2021, 2, 387–409. [Google Scholar] [CrossRef]
- Gómez Borrego, J.; Torrent Burgas, M. Analysis of Host–Bacteria Protein Interactions Reveals Conserved Domains and Motifs That Mediate Fundamental Infection Pathways. Int. J. Mol. Sci. 2022, 23, 11489. [Google Scholar] [CrossRef]
- Park, S.J.; Son, W.S.; Lee, B.-J. Structural Overview of Toxin-Antitoxin Systems in Infectious Bacteria: A Target for Developing Antimicrobial Agents. Biochim. Biophys. Acta 2013, 1834, 1155–1167. [Google Scholar] [CrossRef]
- Belousova, E.A.; Ishchenko, A.A.; Lavrik, O.I. Dna Is a New Target of Parp3. Sci. Rep. 2018, 8, 4176. [Google Scholar] [CrossRef]
- Musheev, M.U.; Schomacher, L.; Basu, A.; Han, D.; Krebs, L.; Scholz, C.; Niehrs, C. Mammalian N1-Adenosine PARylation Is a Reversible DNA Modification. Nat. Commun. 2022, 13, 6138. [Google Scholar] [CrossRef]
- Fehr, A.R.; Singh, S.A.; Kerr, C.M.; Mukai, S.; Higashi, H.; Aikawa, M. The Impact of PARPs and ADP-Ribosylation on Inflammation and Host–Pathogen Interactions. Genes Dev. 2020, 34, 341–359. [Google Scholar] [CrossRef]
- Eckei, L.; Krieg, S.; Bütepage, M.; Lehmann, A.; Gross, A.; Lippok, B.; Grimm, A.R.; Kümmerer, B.M.; Rossetti, G.; Lüscher, B.; et al. The Conserved Macrodomains of the Non-Structural Proteins of Chikungunya Virus and Other Pathogenic Positive Strand RNA Viruses Function as Mono-ADP-Ribosylhydrolases. Sci. Rep. 2017, 7, 41746. [Google Scholar] [CrossRef]
- Fehr, A.R.; Jankevicius, G.; Ahel, I.; Perlman, S. Viral Macrodomains: Unique Mediators of Viral Replication and Pathogenesis. Trends Microbiol. 2018, 26, 598–610. [Google Scholar] [CrossRef]
- Abraham, R.; Hauer, D.; McPherson, R.L.; Utt, A.; Kirby, I.T.; Cohen, M.S.; Merits, A.; Leung, A.K.L.; Griffin, D.E. ADP-Ribosyl-Binding and Hydrolase Activities of the Alphavirus NsP3 Macrodomain Are Critical for Initiation of Virus Replication. Proc. Natl. Acad. Sci. USA 2018, 115, E10457–E10466. [Google Scholar] [CrossRef]
- Grunewald, M.E.; Chen, Y.; Kuny, C.; Maejima, T.; Lease, R.; Ferraris, D.; Aikawa, M.; Sullivan, C.S.; Perlman, S.; Fehr, A.R. The Coronavirus Macrodomain Is Required to Prevent PARP-Mediated Inhibition of Virus Replication and Enhancement of IFN Expression. PLoS Pathog. 2019, 15, e1007756. [Google Scholar] [CrossRef]
- Schuller, M.; Correy, G.J.; Gahbauer, S.; Fearon, D.; Wu, T.; Díaz, R.E.; Young, I.D.; Carvalho Martins, L.; Smith, D.H.; Schulze-Gahmen, U.; et al. Fragment Binding to the Nsp3 Macrodomain of SARS-CoV-2 Identified through Crystallographic Screening and Computational Docking. Sci. Adv. 2021, 7, eabf8711. [Google Scholar] [CrossRef]
- Gahbauer, S.; Correy, G.J.; Schuller, M.; Ferla, M.P.; Doruk, Y.U.; Rachman, M.; Wu, T.; Diolaiti, M.; Wang, S.; Neitz, R.J.; et al. Iterative Computational Design and Crystallographic Screening Identifies Potent Inhibitors Targeting the Nsp3 Macrodomain of SARS-CoV-2. BioRxiv Prepr. Serv. Biol. 2022, 120, e2212931120. [Google Scholar] [CrossRef] [PubMed]
- Sherrill, L.M.; Joya, E.E.; Walker, A.; Roy, A.; Alhammad, Y.M.; Atobatele, M.; Wazir, S.; Abbas, G.; Keane, P.; Zhuo, J.; et al. Design, Synthesis and Evaluation of Inhibitors of the SARS-CoV-2 Nsp3 Macrodomain. Bioorg. Med. Chem. 2022, 67, 116788. [Google Scholar] [CrossRef] [PubMed]
- Bernheim, A.; Sorek, R. The Pan-Immune System of Bacteria: Antiviral Defence as a Community Resource. Nat. Rev. Microbiol. 2020, 18, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Millman, A.; Bernheim, A.; Stokar-Avihail, A.; Fedorenko, T.; Voichek, M.; Leavitt, A.; Oppenheimer-Shaanan, Y.; Sorek, R. Bacterial Retrons Function In Anti-Phage Defense. Cell 2020, 183, 1551–1561.e12. [Google Scholar] [CrossRef]
- González-Delgado, A.; Mestre, M.R.; Martínez-Abarca, F.; Toro, N. Prokaryotic Reverse Transcriptases: From Retroelements to Specialized Defense Systems. FEMS Microbiol. Rev. 2021, 45, fuab025. [Google Scholar] [CrossRef]
- Bobonis, J.; Mitosch, K.; Mateus, A.; Karcher, N.; Kritikos, G.; Selkrig, J.; Zietek, M.; Monzon, V.; Pfalz, B.; Garcia-Santamarina, S.; et al. Bacterial Retrons Encode Phage-Defending Tripartite Toxin–Antitoxin Systems. Nature 2022, 609, 144–150. [Google Scholar] [CrossRef]
- Lüscher, B.; Bütepage, M.; Eckei, L.; Krieg, S.; Verheugd, P.; Shilton, B.H. ADP-Ribosylation, a Multifaceted Posttranslational Modification Involved in the Control of Cell Physiology in Health and Disease. Chem. Rev. 2018, 118, 1092–1136. [Google Scholar] [CrossRef]
- Iyer, L.M.; Burroughs, A.M.; Anantharaman, V.; Aravind, L. Apprehending the NAD+-ADPr-Dependent Systems in the Virus World. Viruses 2022, 14, 1977. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).