
An Experiment in Denmark to Infect Wounded Sitka Spruce with the Rotstop Isolate of Phlebiopsis gigantea, and Its Implications for the Control of Heterobasidion annosum in Britain


Abstract
1. Introduction
2. Methods and Materials
2.1. Site
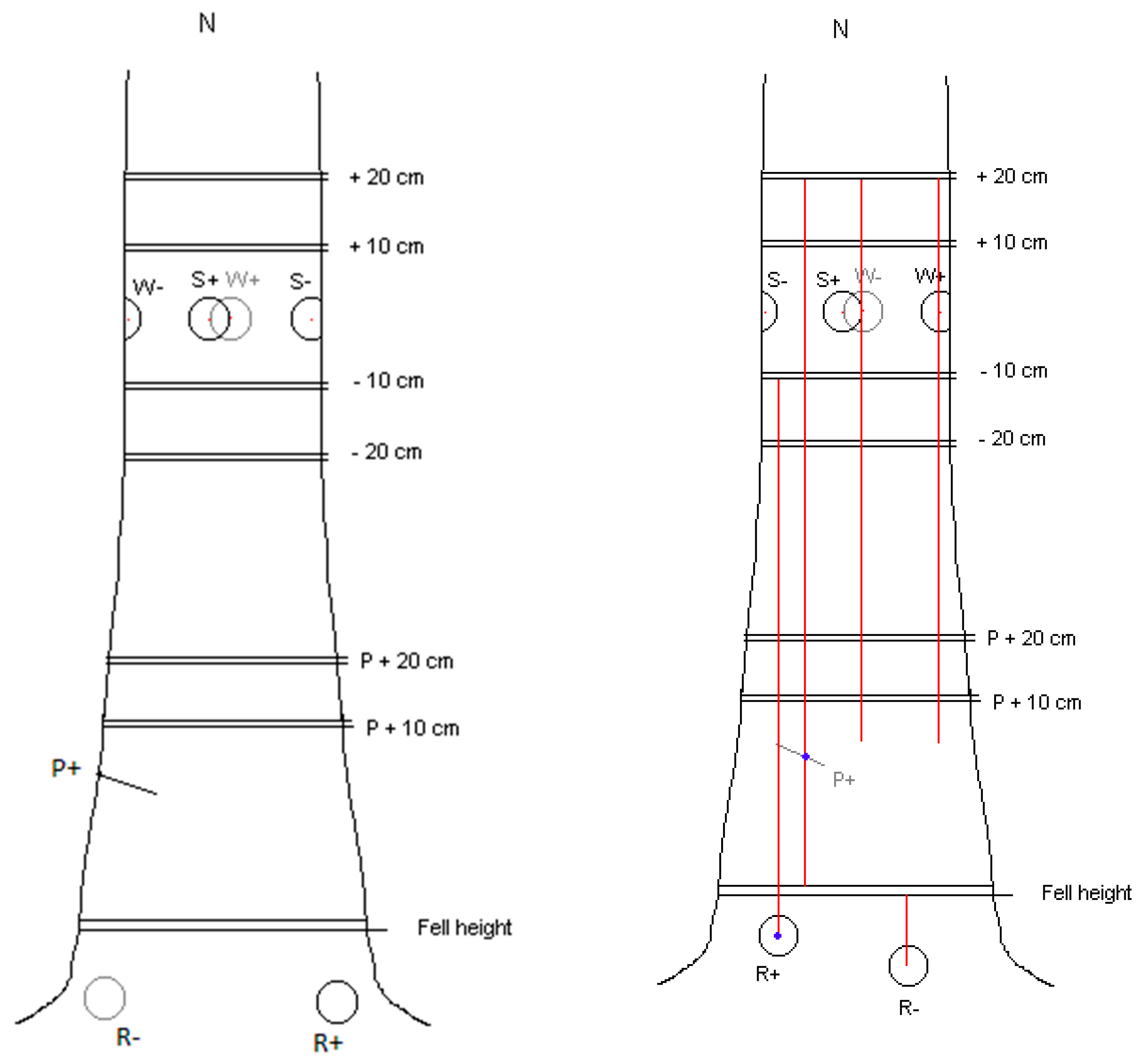
2.2. Wounding
2.3. Preparation and Application of Inocula
2.4. Sampling
3. Results
3.1. Numbers of Trees Sampled
3.2. Physical State of Wounds
3.3. Re-Isolation of P. gigantea (Rotstop Isolate)
3.4. Extent of Stain Associated with Wounding and Inoculation
3.5. SEM and TEM Studies
3.6. Wood Moisture
4. Discussion
4.1. Implications for Wound Colonisation
4.2. Wider Aspects of the Use of BCAs in British Forestry
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pratt, J.E. Fomes annosus butt-rot of Sitka spruce III. Losses in yield and value of timber in diseased trees and stands. Forestry 1979, 52, 113–127. [Google Scholar] [CrossRef]
- Redfern, D.B.; Pratt, J.E.; Hendry, S.J.; Low, J.D. Development of a policy and strategy for controlling infection by Heterobasidion annosum in British forests: A review of supporting research. For. Int. J. For. Res. 2010, 83, 207–218. [Google Scholar] [CrossRef]
- Pratt, J.E. Use of a computer model of a forest disease in long-term strategic planning. In Proceedings of the 11th International Conference on Root and Butt Rots, Poznan, Poland, 16–22 August 2004; Mańka, M., Lakomy, P., Eds.; The August Cieszkowski Agricultural University: Poznan, Poland; pp. 467–474. [Google Scholar]
- Pratt, J.E. Stump treatment against Fomes. In Forest Research Annual Report and Accounts 2001–2002; The Stationary Office: Edinburgh, UK, 2002; pp. 76–85. [Google Scholar]
- Bendz-Hellgren, M.; Lipponen, K.; Solheim, H.; Thomsen, I.M. The Nordic Countries. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Chapter, 17, Woodward, S., Stenlid, J., Karjalainen, R., Huttermann, A., Eds.; CAB International: Wallingford, UK, 1998. [Google Scholar]
- Korhonen, K.; Lipponen, K.; Bendz, M.; Johansson, S.M.; Ryen, I.; Venn, K.; Seiskari, P.; Niemi, M. Control of Hetero-basidion annosum by stump treatment with “Rotstop”, a new commercial formulation of Peniophora gigantea. In Proceedings of the Eighth IUFRO Conference on Root and But Rots, Sweden/Finland, Uppsala, Sweden, 26 August–1 September 1993; Johansson, M., Stenlid, J., Eds.; Swedish University of Agricultural Science: Uppsala, Sweden, 1994; pp. 675–685. [Google Scholar]
- Dovana, F.; Gonthier, P.; Garbelotto, M. Inter- and Intra-Continental Genetic Variation in the Generalist Conifer Wood Saprobic Fungus Phlebiopsis gigantea. Forests 2021, 12, 751. [Google Scholar] [CrossRef]
- Kallio, T. Peniophora gigantea (Fr.) Massee and wounded spruce (Picea abies (l.) Karst. Acta For. Fenn. 1973, 133, 28. [Google Scholar] [CrossRef][Green Version]
- Brandtberg, M.; (Lallemand Inc., Toronto, ON, Canada). Personal communication, 8 December 2021.
- Bier, J.E.; Foster, R.E.; Salisbury, P.J. Studies in forest pathology IV. Decay of Sitka spruce on the Queen Charlotte Islands. Can. Dept. Agric. Technol. Bull. 1946, 56, 35. [Google Scholar]
- Pratt, J.E.; Johansson, M.; Huttermann, A. Chemical control of Heterobasidion annosum. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Chapter 14; Woodward, S., Stenlid, J., Karjalainen, R., Huttermann, A., Eds.; CAB International: Wallingford, UK, 1988. [Google Scholar]
- Pratt, J.E.; Thor, M. Improving mechanised stump treatment against Fomes root rot in Europe. J. For. 2001, 95, 119–127. [Google Scholar]
- Gregory, S.C. The Development of Stain in Wounded Sitka Spruce Stems. For. Int. J. For. Res. 1986, 59, 199–208. [Google Scholar] [CrossRef]
- Pawsey, R.G.; Gladman, R.J. Decay in Standing Conifers Developing from Extraction Damage; Forest Record No 54; H.M.S.O.: London, UK, 1965; 25p. [Google Scholar]
- Tubby, K. Final Report on Use of Biological Control Products Used Against Heterobasidion annosum (Fomes annosus) on pine and spruce in Britain, Internal report of the Research Agency of the Forestry Commission; 2010; 19p, unpublished. [Google Scholar]
- Magnusson, A.; Skaug, H.J.; Nielsen, A.; Berg, C.W.; Kristensen, K.; Maechler, M.; van Bentham, K.J.; Bolker, B.M.; Brooks, M.E. 2017. glmmTMB: Generalized Linear Mixed Models Using Template Model Builder, R package version 0.1.3; The R Core Team: Vienna, Austria, 2022; Available online: https://github.com/glmmTMB (accessed on 1 June 2022).
- The R Core Team. R: A Language and Environment for Statistical Computing. 4.2.1; The R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Otjen, L.; Blanchette, R.A. A discussion of microstructural changes in wood decay during decomposition by white rot basidiomycetes. Can. J. Bot. 1986, 64, 905–911. [Google Scholar] [CrossRef]
- Liese, W. Ultrastructural Aspects of Woody Tissue Disintegration. Annu. Rev. Phytopathol. 1970, 8, 231–258. [Google Scholar] [CrossRef]
- Schwarze, F.W.M.R.; Engels, J.; Mattheck, C. Fungal Strategies of Wood Dacay in Trees; Springer: Berlin/Heidelberg, Germany, 2000; ISBN 978-3-642-57302-6. [Google Scholar]
- Bailey, P.J.; Woodward, S.; Pratt, J.E. Colonisation and Degradation of Sitka Spruce sapwood by the Rotstop Strain of Phlebiopsis gigantea; Aberdeen Centre for Environmental Sustainability: Aberdeen, UK, 2002; 5p, Unpublished paper. [Google Scholar]
- Meier, H. Über Zellwandabbau durch Holzvermorschungspilze und die submikroskopische Struktur von Fichentra-cheiden und Birkenholz-fasern. Holz Roh Werkst 1955, 13, 323–338. [Google Scholar] [CrossRef]
- Siebold, M.; Leidich, P.; Bertini, M.; Deflorio, G.; Feldmann, J.; Krupp, E.; Halmschlager, E.; Woodward, S. Distribution of elements in the bark of Sitka spruce following wounding and inoculation with Heterobasidion annosum. In Proceedings of the IUFRO WP 7.02.01 Root and But Rot of Forest Trees, Firenzie, Italy, 4–12 September 2011; pp. 52–54. [Google Scholar]
- Woodward, S.; Pearce, R.B. The role of stilbenes in resistance of Sitka spruce (Picea sitchensis (Bong) Carr.) to entry of fungal pathogens. Physiol. Mol. Plant Pathol. 1988, 33, 127–149. [Google Scholar] [CrossRef]
- Sun, H.; Paulin, L.; Alatalo, E.; Asiegbu, F.O. Response of living tissues of Pinus sylvestris to the saprotrophic biocontrol fungus Phlebiopsis gigantea. Tree Physiol. 2011, 31, 438–451. [Google Scholar] [CrossRef]
- Vainio, E.J. Ecological impacts of Phlebiopsis gigantea biocontrol treatment against Heterobasidion spp. as revealed by fungal community profiling and population analyses. Diss. For. 2008, 63, 80. [Google Scholar] [CrossRef]
- O’Hanlon, R.; Harrington, J. The macrofungal component of biodiversity in Irish Sitka spruce forests. Irish Forest. 2011, 68, 40–53. [Google Scholar]
- Pratt, J.E. Fomes annosus Butt-Rot of Sitka spruce 1. Observations on the development of butt rot in individual trees and in stands. Forestry 1979, 52, 11–29. [Google Scholar] [CrossRef]
- Pratt, J.E.; Redfern, D.B. Infection of Sitka spruce stumps by spores of Heterobasidion annosum: Control by means of urea. Forestry 2001, 74, 73–78. [Google Scholar] [CrossRef]
- Johansson, S.; Pratt, J.; O Asiegbu, F. Treatment of Norway spruce and Scots pine stumps with urea against the root and butt rot fungus Heterobasidion annosum—Possible modes of action. For. Ecol. Manag. 2002, 157, 87–100. [Google Scholar] [CrossRef]
- Gibbs, J.; Greig, B.; Pratt, J. Fomes root rot in Thetford Forest, East Anglia: Past, present and future. For. Int. J. For. Res. 2002, 75, 191–202. [Google Scholar] [CrossRef]
- Redfern, D.B. Melanotus proteus: A newly recorded colonist of Sitka spruce stumps in Britain and a potential competitor for Heterobasidion annosum. Plant Pathol. 1991, 40, 483–486. [Google Scholar] [CrossRef]
- Holdenreider, O.; Greig, B.J.W. Biological Methods of Control. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Chapter, 13, Woodward, S., Stenlid, J., Karjalainen, R., Huttermann, A., Eds.; CAB International: Wallingford, UK, 1998; 589p. [Google Scholar]
- Soutrenon, A.; Levy, A.; Legrand, P.; Lung-Escarpment, B.; Guillamin, J.J.; Delatour, C. Comparison between three stump treatments to control Heterobasidion annosum (urea, disodium octaborate tetrahydrate, Phlebiopsis gigantea). In Proceedings of the Root and Butt Rots of Forest Trees Conference, Carcans-Maubuisson, France, 1–7 September 1997; INRA: Paris, France Les Colloques, No 89. ; pp. 381–389. [Google Scholar]
- Redfern, D.B. The effect of wood moisture on infection of Sitka spruce stumps by basidiospores of Heterobasidion annosum. For. Pathol. 1993, 23, 218–235. [Google Scholar] [CrossRef]
- Yerbury, M.D. The Spatial and Temporal Distribution of Moisture within Sitka Spruce Standing Trees and Roundwood Logs; Abstract only. Ph.D. Thesis, University of Glasgow, Glasgow, UK, 2017. Available online: https://theses.gla.ac.uk/7868/ (accessed on 1 June 2022).
- Murray, A.C.; Woodward, S. Temporal changes in functional diversity of culturable bacteria populations in Sitka spruce stumps. For. Pathol. 2007, 37, 217–235. [Google Scholar] [CrossRef]
- Pratt, J.E.; Wang, L. New ways of assessing H. annosum root inoculum. In Proceedings of the XIII International Conference on Root and Butt Rot of Forest Trees, Firenze, Italy, 4–10 September 2012; Capretti, P., Comparini, C., Garbelotto, M., La Porta, N., Santini, A., Eds.; Firenze University Press: Firenze, Italy, 2012; pp. 192–196. [Google Scholar]
- Tubby, K.V.; Scott, D.; Webber, J.F. Relationship between stump treatment coverage using the biological control product PG Suspension, and control of Heterobasidion annosum on Corsican pine, Pinus nigra ssp. laricio. For. Pathol. 2008, 38, 37–46. [Google Scholar] [CrossRef]
- Sun, H.; Korhonen, K.T.; Hantula, J.; Asiegbu, F.O.; Kasanen, R.A.O. Use of a breeding approach for improving biocontrol efficacy of Phlebiopsis gigantea strains against Heterobasidion infection of Norway spruce stumps. FEMS Microbiol. Ecol. 2009, 69, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.-C.; Shen, T.-T.; Jin, W.-T.; Mao, H.-J.; Ran, J.-H.; Wang, X.-Q. Phylotranscriptomics resolves interspecific relationships and indicates multiple historical out-of-North America dispersals through the Bering Land Bridge for the genus Picea (Pinaceae). Mol. Phylogenetics Evol. 2019, 141, 106610. [Google Scholar] [CrossRef] [PubMed]
- Pratt, J.E.; Greig, B.J.W. Heterobasidion annosum: Development of Butt Rot following Thinning in two young First Rotation Stands of Norway Spruce. For. Int. J. For. Res. 1988, 61, 339–347. [Google Scholar] [CrossRef]
- Evans, H.C. Biological Control of Weeds with Fungi. In Agricultural Applications, 2nd ed.; Kempken, F., Ed.; The Mycota XI; Springer: Berlin/Heidelberg, Germany, 2013; pp. 145–172. [Google Scholar]
- Pratt, J.E. Planning and management of stump treatment trials. In Proceedings of the 12th International Conference on Root and Butt Rots of Forest Trees, Berkley, CA, USA, Medford, OR, USA, 12–19 August 2007; Garbelotto, M., Gonthier, P., Eds.; The University of California: Berkeley, CA, USA, 2008; pp. 212–215. [Google Scholar]
- Heslin, M.C.; Blasius, D.; McElhinney, C.; Mitchell, D.T. Michorrhizal and associated fungi of Sitka spruce in Irish forest mixed stands. Eur. J. For. Path. 1992, 22, 46–57. [Google Scholar] [CrossRef]
- Humphrey, J.; Newton, A.; Peace, A.; Holden, E. The importance of conifer plantations in northern Britain as a habitat for native fungi. Biol. Conserv. 2000, 96, 241–252. [Google Scholar] [CrossRef]
- Humphrey, J. Benefits to biodiversity from developing old-growth conditions in British upland spruce plantations: A review and recommendations. For. Int. J. For. Res. 2005, 78, 33–53. [Google Scholar] [CrossRef]
- Wilson, E.; Wirz, R.; Byrne, L. Transformation of Sitka spruce plantations to continuous cover forestry at Dunranhill Forest, County Wicklow, Ireland. Scott. For. 2021, 75, 32–39. [Google Scholar]
- Vasiliauskas, R.; Larsson, E.; Larsson, K.-H.; Stenlid, J. Persistence and long-term impact of Rotstop biological control agent on mycodiversity in Picea abies stumps. Biol. Control 2005, 32, 295–304. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P+ or P− | S+ and S− | W+ and W− | R+ and R− |
|---|---|---|---|
| 4.5 mm Pressler boring colonised by Rotstop (P+) or sterile (P−) inoculated into radial hole drilled to tree centres. | Oidial spore suspension (S+) or sterile water (S−) on filter paper placed onto exposed xylem in the centre of an 80 mm circular wound. | 20 mm × 20 mm wood dowel colonised by Rotstop (W+) or sterile (W−) inserted into 19 mm diameter radial hole at the centre of 80 mm diameter stem wounds. | 20 mm × 20 mm wood dowel colonised by Rotstop (R+) or sterile (R−) inserted into 19 mm diameter radial hole at the centre of an 80 mm diameter wound cut into the surface of a lateral root. |
| Inoculation Number | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Inoculation date | 20.11.97 | 13.4.98 | 7.7.98 | 29.9.98 |
| Felling and sampling date | 21.10.98 | 20.5.99 | 20.5.99 | 6.3.2000 |
| Months incubated | 11 | 13 | 10 | 17 |
| Inoculum Type | Treatment | Inoculum Position | +Rotstop | −Rotstop | Number Inoculated |
|---|---|---|---|---|---|
| Oidial spores | S+, S− | Stem | 39 | 39 | 40 |
| 20 mm pine dowel | W+, W− | Stem | 39 | 39 | 40 |
| 20 mm pine dowel | R+, R− | Root | 39 | 39 | 40 |
| 4.5 mm pine core | P+, P− | Stem | 19 | 20 | 20 * |
| Inoculation | W+ | W− | R+ | R− | P+ | P− | S+ | S− |
|---|---|---|---|---|---|---|---|---|
| Total | 15/39 | 2/39 | 7/39 | 0/39 | 11/19 | 0/20 | 1/39 | 0/39 |
| Percent | 39% a | 5% b | 18% b | 0% b | 58% a | 0% b | 3% b | 0% b |
| Max extent, cm | (+20/−51) | (0) | (+63/−0) | (0) | (+10/−27) | (0) | (0) | (0) |
| Treatment | S+ | S− | W+ | W− | R+ | R− | P+ | P− |
|---|---|---|---|---|---|---|---|---|
| Mean axial extent (cm) | 8 | 14 | 85 | 52 | 45 | 18 | 80 | 52 |
| No. columns of stain | 9 | 14 | 39 | 38 | 39 | 36 | 19 | 19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pratt, J.; Thomsen, I.M. An Experiment in Denmark to Infect Wounded Sitka Spruce with the Rotstop Isolate of Phlebiopsis gigantea, and Its Implications for the Control of Heterobasidion annosum in Britain. Pathogens 2022, 11, 937. https://doi.org/10.3390/pathogens11080937
Pratt J, Thomsen IM. An Experiment in Denmark to Infect Wounded Sitka Spruce with the Rotstop Isolate of Phlebiopsis gigantea, and Its Implications for the Control of Heterobasidion annosum in Britain. Pathogens. 2022; 11(8):937. https://doi.org/10.3390/pathogens11080937
Chicago/Turabian StylePratt, Jim, and Iben M. Thomsen. 2022. "An Experiment in Denmark to Infect Wounded Sitka Spruce with the Rotstop Isolate of Phlebiopsis gigantea, and Its Implications for the Control of Heterobasidion annosum in Britain" Pathogens 11, no. 8: 937. https://doi.org/10.3390/pathogens11080937
APA StylePratt, J., & Thomsen, I. M. (2022). An Experiment in Denmark to Infect Wounded Sitka Spruce with the Rotstop Isolate of Phlebiopsis gigantea, and Its Implications for the Control of Heterobasidion annosum in Britain. Pathogens, 11(8), 937. https://doi.org/10.3390/pathogens11080937