
Combined Effect of Entomopathogens against Thrips tabaci Lindeman (Thysanoptera: Thripidae): Laboratory, Greenhouse and Field Trials




Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Entomopathogenic Nematodes
2.3. Entomopathogenic Fungi
2.4. Small Cup Bioassay in Laboratory
2.5. Potted Soil Bioassay in Laboratory
2.6. Potted Plant Bioassay in Greenhouse
2.7. Field Trials
2.8. Statistical Analysis
3. Results
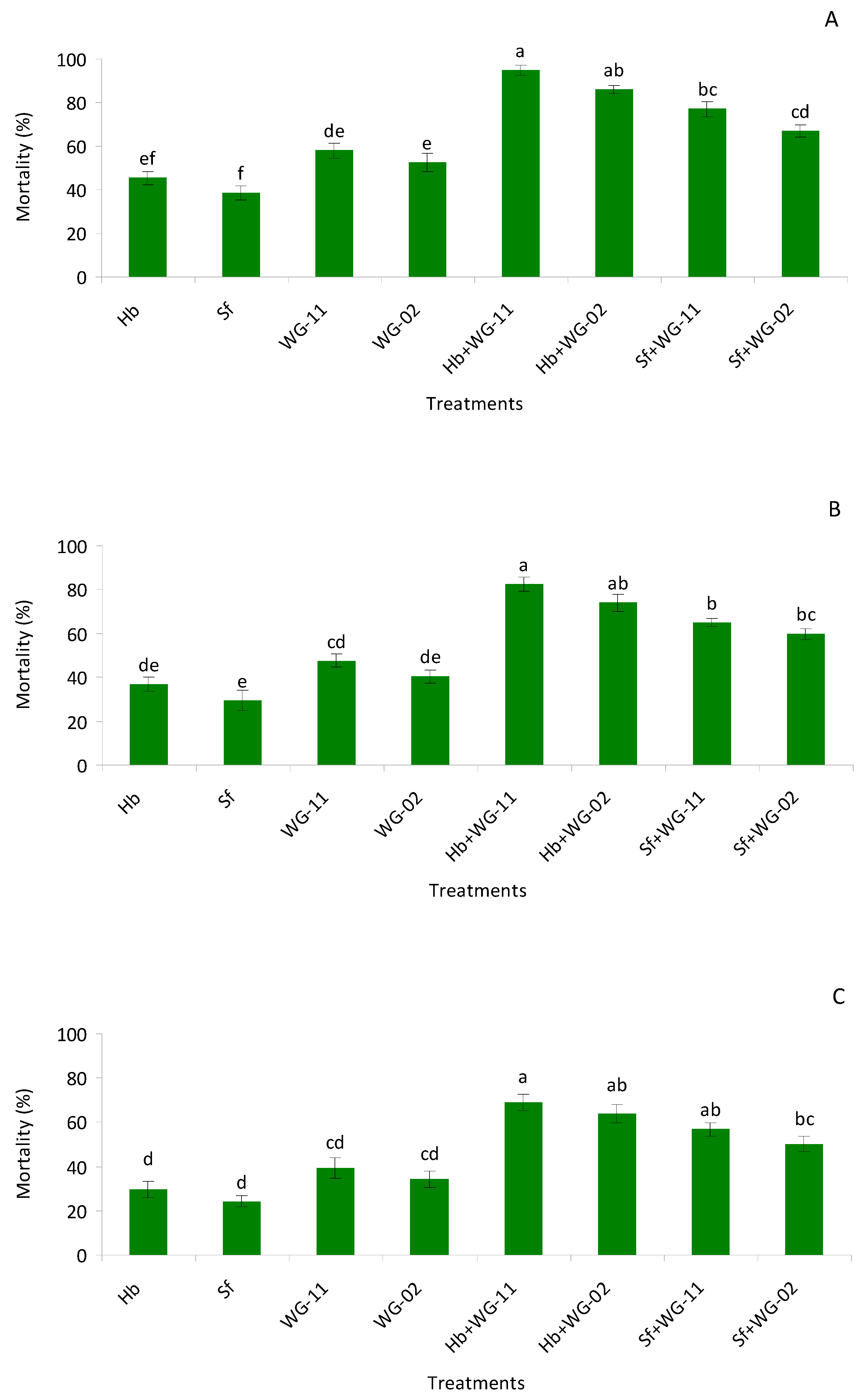
3.1. Small Cup Bioassay in Laboratory
3.2. Potted Soil Bioassay in Laboratory
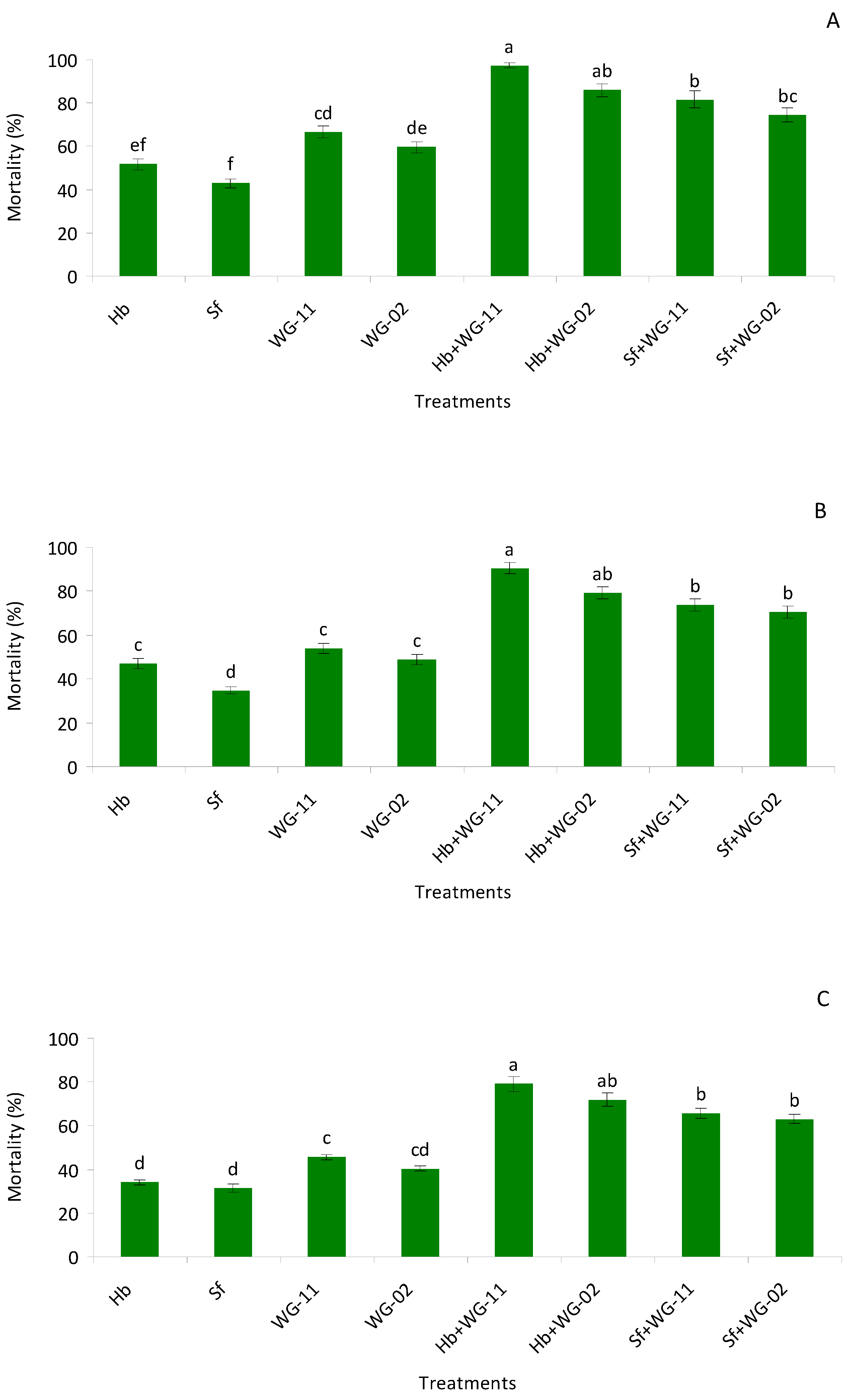
3.3. Plant Potted Bioassay in Geenhouse
3.4. Field Trials
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murai, T. Effect of temperature on development and reproduction of the onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), on pollen and honey solution. Appl. Entomol. Zool. 2000, 35, 499–504. [Google Scholar] [CrossRef]
- McKenzie, C.L.; Miller, M.E.; Edelson, J.V.; Cartwright, B. Injury to Onions by Thrips tabaci (Thysanoptera: Thripidae) and Its Role in the Development of Purple Blotch. Environ. Entomol. 1993, 22, 1266–1277. [Google Scholar] [CrossRef]
- Shelton, A.M.; Wilsey, W.T.; Schmaedick, M.A. Management of Onion Thrips (Thysanoptera: Thripidae) on Cabbage by Using Plant Resistance and Insecticides. J. Econ. Entomol. 1998, 91, 329–333. [Google Scholar] [CrossRef]
- Koschier, E.H.; Sedy, K.A.; Novak, J. Influence of plant volatiles on feeding damage caused by the onion thrips Thrips tabaci. Crop. Prot. 2002, 21, 419–425. [Google Scholar] [CrossRef]
- Birithia, R.; Subramanian, S.; Pappu, H.R.; Sseruwagi, P.; Muthomi, J.W.; Narla, R.D. First Report of Iris yellow spot virus Infecting Onion in Kenya and Uganda. Plant Dis. 2011, 95, 1195. [Google Scholar] [CrossRef]
- Birithia, R.; Subramanian, S.; Muthomi, J.; Narla, R. Resistance to Iris yellow spot virus and onion thrips among onion varieties grown in Kenya. Int. J. Trop. Insect Sci. 2014, 34, 73–79. [Google Scholar] [CrossRef]
- Sdoodee, R.; Teakle, D.S. Transmission of tobacco streak virus by Thrips tabaci: A new method of plant virus transmission. Plant Pathol. 1987, 36, 377–380. [Google Scholar] [CrossRef]
- Hardy, V.G.; Teakle, D.S. Transmission of sowbane mosaic virus by Thrips tabaci in the presence and absence of virus-carrying pollen. Ann. Appl. Biol. 1992, 121, 315–320. [Google Scholar] [CrossRef]
- Kritzman, A.; Gera, A.; Raccah, B.; van Lent, J.W.M.; Peters, D. The route of tomato spotted wild virus inside the thrips body in relation to transmission efficiency. Arch. Virol. 2002, 147, 2143–2156. [Google Scholar] [CrossRef] [PubMed]
- Jenser, G.; Gáborjányi, R.; Szénási, Á.; Almaši, A.; Grasselli, M. Significance of hibernated Thrips tabaci Lindeman (Thysan., Thripidae) adults in the epidemic of tomato spotted wilt virus. J. Appl. Entomol. 2003, 127, 7–11. [Google Scholar] [CrossRef]
- Berndt, O.; Meyhöfer, R.; Poehling, H.-M. The edaphic phase in the ontogenesis of Frankliniella occidentalis and comparison of Hypoaspis miles and Hypoaspis aculeifer as predators of soil-dwelling thrips stages. Biol. Control. 2004, 30, 17–24. [Google Scholar] [CrossRef]
- Ansari, M.A.; Brownbridge, M.; Shah, F.A.; Butt, T.M. Efficacy of entomopathogenic fungi against soil-dwelling life stages of western flower thrips, Frankliniella occidentalis, in plant-growing media. Entomol. Exp. Appl. 2008, 127, 80–87. [Google Scholar] [CrossRef]
- Steiner, M.Y.; Spohr, L.J.; Goodwin, S. Relative humidity controls pupation success and dropping behaviour of western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Aust. J. Entomol. 2010, 50, 179–186. [Google Scholar] [CrossRef]
- Holmes, N.D.; Bennison, J.A.; Maulden, K.A.; Kirk, W.D.J. The pupation behaviour of the western flower thrips, Frankliniella occidentalis (Pergande). Acta Phytopathol. Entomol. Hung. 2012, 47, 87–96. [Google Scholar] [CrossRef]
- MacIntyre Allen, J.K.; Scott-Dupree, C.D.; Tolman, J.H.; Harris, C.R. Resistance of Thrips tabaci to pyrethroid and organo-phosphorous insecticides in Ontario, Canada. Pest. Manag. Sci. 2005, 61, 809–815. [Google Scholar] [CrossRef]
- Shelton, A.M.; Zhao, J.Z.; Nault, B.A.; Plate, J.; Musser, F.R.; Larentzaki, E. Patterns of insecticide resistance in onion thrips (Thysanoptera: Thripidae) in onion fields in New York. J. Econ. Entomol. 2006, 99, 1798–1804. [Google Scholar] [CrossRef]
- Bielza, P. Insecticide resistance management strategies against the western flower thrips, Frankliniella occidentalis. Pest. Manag. Sci. 2008, 64, 1131–1138. [Google Scholar] [CrossRef]
- Herron, G.A.; James, T.M.; Rophail, J.; Mo, J. Australian populations of onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), are resistant to some insecticides used for their control. Aust. J. Entomol. 2008, 47, 361–364. [Google Scholar] [CrossRef]
- Gao, Y.; Lei, Z.; Reitz, S.R. Western flower thrips resistance to insecticides: Detection, mechanisms and management strategies. Pest. Manag. Sci. 2012, 68, 1111–1121. [Google Scholar] [CrossRef]
- Loomans, A.J.M.; Murai, T. Culturing thrips and parasitoids. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International Harpenden: Herts, UK, 1997; pp. 477–503. [Google Scholar]
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Fail, J.; Shelton, A.M. Onion thrips (Thysanoptera: Thripidae): A global pest of in-creasing concern in onion. J. Econ. Entomol. 2011, 104, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Berndt, O. Entomopathogenic nematodes and soil dwelling predatory mites: Suitable antagonists for enhanced biological control of Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Ph.D. Thesis, University of Hannover, Hannover, Germany, 20 December 2002; p. 140. [Google Scholar]
- Shapiro-Ilan, D.I.; Hazir, S.; Glazer, I. Advances in use of entomopathogenic nematodes in IPM, In Integrated Management of Insect Pests: Current and Future Developments; Kogan, M., Heinrichs, E.A., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2020; pp. 649–678. [Google Scholar]
- Shapiro-Ilan, D.I.; Hiltpold, I.; Lewis, E.E. Ecology of invertebrate pathogens: Nematodes. In Ecology of Invertebrate Diseases; Hajek, A.E., Shapiro-Ilan, D.I., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2018; pp. 415–440. [Google Scholar]
- Boemare, N.; Laumond, C.; Mauleon, H. The entomopathogenic nematode–bacterium complex: Biology, life cycle, and verte-brate safety. Biocontrol Sci. Technol. 1996, 6, 333–345. [Google Scholar] [CrossRef]
- Adams, B.J.; Nguyen, K.B. Taxonomy and Systematics. In Entomopathogenic Nematology; Gaugler, R., Ed.; CABI Publishing: New Brunswick, NJ, USA, 2002; pp. 1–34. [Google Scholar]
- Dowds, B.C.A.; Peters, A. Virulence mechanisms. In Entomopathogenic Nematology; Gaugler, R., Ed.; CABI Publishing: New Brunswick, NJ, USA, 2003; pp. 79–90. [Google Scholar]
- Shapiro-Ilan, D.I.; Han, R.; Dolinksi, C. Entomopathogenic Nematode Production and Application Technology. J. Nematol. 2012, 44, 206–217. [Google Scholar]
- Al-Siyabi, A.A.; Kinawy, M.M.; Al-Ansri, M.; Mahar, A.N.; Gowen, S.R.; Hague, N.G.N. The susceptibility of onion thrips, Thrips tabaci to Heterorhabditis indicus. Commun. Agric. Appl. Biol. Sci. 2006, 71, 239–243. [Google Scholar]
- Kashkouli, M.; Khajehali, J.; Poorjavad, N. Impact of entomopathogenic nematodes on Thrips tabaci Lindeman (Thysanoptera: Thripidae) life stages in the laboratory and under semi field conditions. J. Biopest. 2014, 7, 77–84. [Google Scholar]
- Ebssa, L.; Borgemeister, C.; Poehling, H.-M. Effectiveness of different species/strains of entomopathogenic nematodes for control of western flower thrips (Frankliniella occidentalis) at various concentrations, host densities, and temperatures. Biol. Control. 2004, 29, 145–154. [Google Scholar] [CrossRef]
- Ebssa, L.; Borgemeister, C.; Berndt, O.; Poehling, H.-M. Impact of Entomopathogenic Nematodes on Different Soil-dwelling Stages of Western Flower Thrips, Frankliniella occidentalis (Thysanoptera: Thripidae), in the Laboratory and Under Semi-field Conditions. Biocontrol Sci. Technol. 2001, 11, 515–525. [Google Scholar] [CrossRef]
- Ekesi, S.; Maniania, N.K. Use of Entomopathogenic Fungi in Biological Pest. Management; Research Signpost: Kerala, India, 2007; p. 333. [Google Scholar]
- Feng, M.G.; Poprawski, T.J.; Khachatourians, G.G. Production, formulation and application of the entomopathogenic fungus Beauveria bassiana for insect control: Current status. Biocontrol Sci. Technol. 1994, 4, 3–34. [Google Scholar] [CrossRef]
- Vega, F.E.; Meyling, N.V.; Luangsa-Ard, J.J.; Blackwell, M. Fungal Entomopathogens. In Insect Pathology; Elsevier BV: Amsterdam, The Netherlands, 2012; pp. 171–220. [Google Scholar]
- Samšiňáková, A.; Mišíková, S.; Leopold, J. Action of enzymatic systems of Beauveria bassiana on the cuticle of the greater wax moth larvae (Galleria mellonella). J. Invertebr. Pathol. 1971, 18, 322–330. [Google Scholar] [CrossRef]
- Maniania, N.K.; Sithanantham, S.; Ekesi, S.; Ampong-Nyarko, K.; Baumgartner, J.; Lohr, B.; Matoka, C.M. A field trial of the entomogenous fungus Metarhizium anisopliae for control of onion thrips, Thrips tabaci. Crop. Prot. 2003, 22, 553–559. [Google Scholar] [CrossRef]
- Al-mazra’awi, M.S.; Al-Abbadi, A.; Shatnawi, M.A.; Ateyyat, M. Effect of application method on the interaction between Beauveria bassiana and neem tree extract when combined for Thrips tabaci (Thysanoptera: Thripidae) control. J. Food Agricul. Environ. 2009, 7, 869–873. [Google Scholar]
- Wu, S.; Gao, Y.; Xu, X.; Zhang, Y.; Wang, J.; Lei, Z.; Smagghe, G. Laboratory and greenhouse evaluation of a new ento-mopathogenic strain of Beauveria bassiana for control of the onion thrips Thrips tabaci. Biocontrol Sci. Technol. 2013, 23, 794–802. [Google Scholar] [CrossRef]
- Williams, M.E.D.C. Biological Control of Thrips on Ornamental Crops: Interactions Between the Predatory Mite Neoseiulus cucumeris (Acari: Phytoseiidae) and Western Flower Thrips, Frankliniella occidentalis (Thysanoptera: Thripidae), on Cyclamen. Biocontrol Sci. Technol. 2001, 11, 41–55. [Google Scholar] [CrossRef]
- Herrin, B.; Warnock, D. Resistance of Impatiens Germplasm to Western Flower Thrips Feeding Damage. HortScience 2002, 37, 802–804. [Google Scholar] [CrossRef]
- Arthurs, S.; Heinz, K.M. Evaluation of the nematodes Steinernema feltiae and Thripinema nicklewoodi as biological control agents of western flower thrips Frankliniella occidentalis infesting chrysanthemum. Biocontrol Sci. Technol. 2006, 16, 141–155. [Google Scholar] [CrossRef]
- Brownbridge, M.; Buitenhuis, R.; Murphy, G.; Waite, M.; Scott-Dupree, C. Banker plants, trap crops and other bio-protection developments in Canadian greenhouse floriculture. In Proceedings of the 4th International Symposium on Biological Control of Arthropods, Pucon, Chile, 4–8 March 2013; pp. 133–136. [Google Scholar]
- Ansari, M.; Tirry, L.; Moens, M. Interaction between Metarhizium anisopliae CLO 53 and entomopathogenic nematodes for the control of Hoplia philanthus. Biol. Control. 2004, 31, 172–180. [Google Scholar] [CrossRef]
- Ansari, M.A.; Shah, F.A.; Butt, T.M. Combined use of entomopathogenic nematodes and Metarhizium anisopliae as a new ap-proach for black vine weevil, Otiorhynchus sulcatus, control. Entomol. Experim. Appl. 2008, 129, 340–347. [Google Scholar] [CrossRef]
- Ansari, M.A.; Shah, F.A.; Butt, T.M. The entomopathogenic nematode Steinernema kraussei and Metarhizium anisopliae work synergistically in controlling overwintering larvae of the black vine weevil, Otiorhynchus sulcatus, in strawberry growbags. Biocontrol Sci. Technol. 2010, 20, 99–105. [Google Scholar] [CrossRef]
- Koppenhöfer, A.; Grewal, P. Nematodes as biocontrol agents. Entomology and nematology. In Compatibility and Interactions with Agrochemicals and Other Biocontrol Agents; Grewal, P.S., Ehlers, R., Shapiro-Ilan, D.I., Eds.; CABI Publishing: Oxfordshire, UK, 2005; pp. 363–381. [Google Scholar]
- Tarasco, E.; Santiago-Álvarez, C.; Triggian, O.; Quesada-Moraga, E. Laboratory studies on the competition for insect haemocoel between Beauveria bassiana and Steinernema ichnusae recovered in the same ecological niche. Biocontrol Sci. Technol. 2011, 21, 693–704. [Google Scholar] [CrossRef]
- Usman, M.; Gulzar, S.; Wakil, W.; Wu, S.; Piñero, J.C.; Leskey, T.C.; Nixon, L.J.; Oliveira-Hofman, C.; Toews, M.D.; Shapiro-Ilan, D. Virulence of Entomopathogenic Fungi to Rhagoletis pomonella (Diptera: Tephritidae) and Interactions with Entomopathogenic Nematodes. J. Econ. Entomol. 2020, 113, 2627–2633. [Google Scholar] [CrossRef] [PubMed]
- Otieno, J.A.; Pallmann, P.; Poehling, H.-M. The combined effect of soil-applied azadirachtin with entomopathogens for inte-grated management of western flower thrips. J. Appl. Entomol. 2016, 140, 174–186. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Morales-Ramos, J.A.; Rojas, M.G. In Vivo Production of Entomopathogenic Nematodes. In Advanced Structural Safety Studies; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2016; Volume 1477, pp. 137–158. [Google Scholar]
- Inglis, G.D.; Enkerli, J.; Goettel, M.S. Laboratory techniques used for entomopathogenic fungi: Hypocreales. In Manual of Tech-niques in Invertebrate Pathology; Lacey, L.A., Ed.; Academic Press: London, UK, 2012; pp. 189–253. [Google Scholar]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Mead, R.; Sokal, R.R.; Rohlf, F.J. Biometry. Biometry 1982, 38, 863. [Google Scholar] [CrossRef]
- Minitab. MINITAB Release 14 for Windows; Minitab Inc.: State College, PA, USA, 2003. [Google Scholar]
- Shapiro-Ilan, D.I.; Jackson, M.; Reilly, C.C.; Hotchkiss, M.W. Effects of combining an entomopathogenic fungi or bacterium with entomopathogenic nematodes on mortality of Curculio caryae (Coleoptera: Curculionidae). Biol. Control. 2004, 30, 119–126. [Google Scholar] [CrossRef]
- Azazy, A.M.; AbdelAll, M.F.M.; El-Sappagh, I.A.; Khalil, A.E.H. Biological control of the onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), in open fields using Egyptian entomopathogenic nematode isolates. Egypt. J. Biol. Pest. Control. 2018, 28, 27. [Google Scholar] [CrossRef]
- Dlamini, T.; Allsopp, E.; Malan, A. Management of Frankliniella occidentalis (Western Flower Thrips), and the Potential use of Entomopathogenic Nematodes: A South African Perspective. Afr. Entomol. 2019, 27, 265–278. [Google Scholar] [CrossRef]
- Barbercheck, M.E.; Kaya, H.K. Interactions between Beauveria bassiana and the entomopathogenic nematodes, Steinernema feltiae and Heterorhabditis heliothidis. J. Invertebra. Pathol. 1990, 55, 225–234. [Google Scholar] [CrossRef]
- Sánchez, L. Heterorhabditis bacteriophora HCEstrategia de desarrollo como agente de control biológico de plagas in-sectiles. Ph.D. Thesis, Universidad Agraria de la Habana, Centro Nacional de Sanidad Agropecuaria, San José de las Lajas, Cuba, 2002. [Google Scholar]
- Zayed, A.; Shamseldean, M.; Abd El Aleem, K.M.; Fergany, Y.A. Interactions between Beauveria bassiana and the ento-mopathogenic nematodes Heterorhabditis bacteriophora and Steinernema sp. In vivo effects on host preference of the nematode and on pathogen progeny production in the greater wax moth Galleria mellonella L. Efflatonia 2003, 3, 15–24. [Google Scholar]
- Correa-Cuadros, J.P.; Sáenz-Aponte, A.; Rodríguez-Bocanegra, M.X. In vitro interaction of Metarhizium anisopliae Ma9236 and Beauveria bassiana Bb9205 with Heterorhabditis bacteriophora HNI0100 for the control of Plutella xylostella. SpringerPlus 2016, 5, 1–8. [Google Scholar] [CrossRef]
- Ansari, M.A.; Shah, F.A.; Tirry, L.; Moens, M. Field trials against Hoplia philanthus (Coleoptera: Scarabaeidae) with a combi-nation of an entomopathogenic nematode and the fungus Metarhizium anisopliae CLO. Biol. Control. 2006, 39, 453–459. [Google Scholar] [CrossRef]
- Ecology of Invertebrate Diseases; Hajek, A.E., Shapiro-Ilan, D.I., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2018; p. 657. [Google Scholar]



{kind=link}
{kind=link}
| Stage | Treatments | Observed Mortality (%) | Expected Mortality (%) | Chi Square | Interaction |
|---|---|---|---|---|---|
| Pre-pupae | Hb + WG-11 | 94.73 | 77.10 | 4.03 | Synergism |
| Hb + WG-02 | 85.96 | 74.23 | 1.85 | Additive | |
| Sf + WG-11 | 77.19 | 74.15 | 0.12 | Additive | |
| Sf + WG-02 | 66.66 | 70.91 | 0.25 | Additive | |
| Pupae | Hb + WG-11 | 82.45 | 66.75 | 3.69 | Additive |
| Hb + WG-02 | 73.68 | 62.32 | 2.06 | Additive | |
| Sf + WG-11 | 64.91 | 63.06 | 0.05 | Additive | |
| Sf + WG-02 | 59.64 | 58.14 | 0.03 | Additive | |
| Late second instar larvae | Hb + WG-11 | 68.42 | 55.83 | 2.83 | Additive |
| Hb + WG-02 | 63.15 | 52.04 | 2.37 | Additive | |
| Sf + WG-11 | 56.14 | 52.60 | 0.23 | Additive | |
| Sf + WG-02 | 49.12 | 48.53 | 0.007 | Additive |
| Stage | Treatments | Observed Mortality (%) | Expected Mortality (%) | Chi Square | Interaction |
|---|---|---|---|---|---|
| Pre-pupae | Hb + WG-11 | 97.36 | 83.91 | 2.15 | Additive |
| Hb + WG-02 | 85.96 | 80.53 | 0.36 | Additive | |
| Sf + WG-11 | 81.57 | 80.99 | 0.004 | Additive | |
| Sf + WG-02 | 74.56 | 76.99 | 0.07 | Additive | |
| Pupae | Hb + WG-11 | 90.43 | 75.55 | 2.93 | Additive |
| Hb + WG-02 | 79.13 | 72.78 | 0.55 | Additive | |
| Sf + WG-11 | 73.91 | 69.94 | 0.22 | Additive | |
| Sf + WG-02 | 70.43 | 66.54 | 0.22 | Additive | |
| Late second instar larvae | Hb + WG-11 | 78.94 | 64.21 | 3.37 | Additive |
| Hb + WG-02 | 71.92 | 60.75 | 2.05 | Additive | |
| Sf + WG-11 | 65.78 | 62.77 | 0.14 | Additive | |
| Sf + WG-02 | 63.15 | 59.18 | 0.26 | Additive |
| Treatments | 1 Day | 2 Day | 3 Day | 4 Day | 5 Day | 6 Day | 7 Day |
|---|---|---|---|---|---|---|---|
| Hb | 28.33 ± 1.28 bc | 20.50 ± 1.56 bc | 13.16 ± 1.19 c | 10.16 ± 0.70 c | 7.16 ± 0.79 c | 3.83 ± 0.60 c | 1.66 ± 0.33 b |
| Sf | 32.50 ± 1.11 ab | 25.16 ± 1.07 ab | 17.50 ± 1.43 b | 14.50 ± 1.25 b | 10.33 ± 0.71 b | 6.50 ± 0.76 b | 0.00 ± 0.00 c |
| WG-11 | 19.66 ± 1.45 de | 12.50 ± 1.68 de | 8.33 ± 0.55 de | 6.66 ± 0.88 cde | 3.66 ± 0.49 d | 1.66 ± 0.33 cd | 0.00 ± 0.00 c |
| WG-22 | 24.83 ± 1.92 cd | 16.33 ± 1.80 cd | 10.66 ± 0.88 cd | 7.83 ± 0.87 cd | 4.16 ± 0.30 cd | 1.33 ± 0.42 d | 0.00 ± 0.00 c |
| Hb + WG-11 | 8.66 ± 0.55 g | 5.00 ± 0.73 f | 2.50 ± 0.42 f | 1.66 ± 0.33 f | 0.00 ± 0.00 e | 0.00 ± 0.00 d | 0.00 ± 0.00 c |
| Hb + WG-22 | 10.33 ± 0.49 fg | 5.33 ± 0.71 f | 2.66 ± 0.33 f | 2.50 ± 0.42 ef | 0.00 ± 0.00 e | 0.00 ± 0.00 d | 0.00 ± 0.00 c |
| Sf + WG-11 | 14.83 ± 1.10 ef | 7.16 ± 0.65 ef | 5.83 ± 0.60 ef | 3.66 ± 0.33 def | 0.00 ± 0.00 e | 0.00 ± 0.00 d | 0.00 ± 0.00 c |
| Sf + WG-22 | 17.83 ± 1.24 e | 9.16 ± 0.83 ef | 8.16 ± 0.60 de | 4.16 ± 0.40 def | 0.00 ± 0.00 e | 0.00 ± 0.00 d | 0.00 ± 0.00 c |
| Control | 37.33 ± 1.11 a | 29.16 ± 1.16 a | 31.00 ± 1.06 a | 25.50 ± 1.64 a | 18.83 ± 1.57 a | 11.66 ± 0.98 a | 4.83 ± 0.74 a |
| Df | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 |
| F | 63.35 | 54.87 | 117.35 | 68.86 | 89.36 | 59.97 | 36.19 |
| P | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Stage | Treatments | 1 DBA | 3 DPA | 7 DPA | 11 DPA | 15 DPA |
|---|---|---|---|---|---|---|
| Larvae | Hb | 48.85 ± 1.30 ab | 44.53 ± 1.75 ab | 38.51 ± 2.00 bc | 32.75 ± 1.03 b | 25.03 ± 0.99 b |
| Sf | 49.13 ± 1.72 a | 45.60 ± 1.69 a | 40.11 ± 1.85 b | 34.05 ± 1.18 b | 27.21 ± 1.67 b | |
| WG-11 | 48.38 ± 1.34 ab | 44.48 ± 1.91 ab | 37.51 ± 1.74 bc | 30.31 ± 1.36 bc | 22.61 ± 1.24 bc | |
| WG-02 | 46.46 ± 1.37 ab | 42.05 ± 1.46 abc | 36.51 ± 1.16 bc | 27.16 ± 0.98 cd | 19.41 ± 1.13 cd | |
| Hb + WG-11 | 43.06 ± 1.42 ab | 33.10 ± 1.29 d | 24.58 ± 1.33 e | 11.86 ± 0.88 g | 2.86 ± 0.42 g | |
| Hb + WG-02 | 49.00 ± 1.58 a | 36.35 ± 1.31 cd | 28.86 ± 0.96 de | 17.75 ± 0.80 f | 7.13 ± 0.77 fg | |
| Sf + WG-11 | 42.05 ± 0.81 b | 38.16 ± 1.57 bcd | 32.11 ± 1.13 cd | 20.56 ± 0.87 ef | 11.65 ± 0.78 ef | |
| Sf + WG-02 | 47.45 ± 1.39 ab | 41.25 ± 1.88 abc | 35.66 ± 1.63 bcd | 23.03 ± 0.92 de | 14.28 ± 0.93 de | |
| Control | 46.38 ± 1.71 ab | 47.10 ± 1.41 a | 47.80 ± 1.33 a | 48.70 ± 1.42 a | 49.01 ± 1.64 a | |
| df | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | |
| F | 3.12 | 10.70 | 20.76 | 93.35 | 135.17 | |
| p | 0.0078 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Adult | Hb | 33.03 ± 0.87 ab | 31.06 ± 0.70 ab | 26.50 ± 0.95 b | 21.70 ± 0.75 b | 15.33 ± 1.31 bc |
| Sf | 32.30 ± 1.01 ab | 31.01 ± 0.27 ab | 27.98 ± 0.86 b | 22.01 ± 0.87 b | 17.90 ± 1.42 b | |
| WG-11 | 32.71 ± 1.23 ab | 29.61 ± 1.23 bc | 23.85 ± 1.13 bc | 16.13 ± 1.04 cd | 12.63 ± 1.33 cd | |
| WG-02 | 34.26 ± 0.90 ab | 30.76 ± 0.76 ab | 25.90 ± 0.97 b | 18.15 ± 1.07 bc | 13.43 ± 1.30 bcd | |
| Hb + WG-11 | 33.48 ± 0.75 ab | 22.38 ± 0.81 e | 13.46 ± 0.96 f | 5.51 ± 0.63 f | 1.68 ± 0.19 f | |
| Hb + WG-02 | 35.01 ± 0.75 a | 24.11 ± 1.17 de | 16.08 ± 1.07 ef | 8.18 ± 0.59 ef | 3.05 ± 0.35 f | |
| Sf + WG-11 | 31.41 ± 0.77 ab | 25.30 ± 1.09 cde | 19.08 ± 1.87 de | 12.15 ± 0.92 de | 7.96 ± 0.98 ef | |
| Sf + WG-02 | 34.33 ± 1.23 ab | 27.35 ± 1.33 bcd | 21.23 ± 0.91 cd | 14.21 ± 1.19 cd | 9.46 ± 0.68 de | |
| Control | 30.08 ± 0.74 b | 34.10 ± 0.61 a | 34.60 ± 0.50 a | 36.05 ± 0.69 a | 36.96 ± 0.82 a | |
| df | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | |
| F | 2.48 | 16.48 | 41.71 | 97.66 | 112.08 | |
| p | 0.0274 | <0.01 | <0.01 | <0.01 | <0.01 |
| Stage | Treatments | 1 DBA | 3 DPA | 7 DPA | 11 DPA | 15 DPA |
|---|---|---|---|---|---|---|
| Larvae | Hb | 53.61 ± 1.85 abc | 50.08 ± 2.71 a | 43.53 ± 1.32 bc | 37.01 ± 1.74 b | 26.63 ± 1.49 bc |
| Sf | 55.21 ± 1.88 ab | 51.10 ± 3.15 a | 45.36 ± 1.82 b | 40.06 ± 2.12 b | 31.41 ± 1.94 b | |
| WG-11 | 57.25 ± 2.06 a | 48.40 ± 2.55 ab | 41.21 ± 1.08 bcd | 32.81 ± 1.89 bc | 19.95 ± 1.10 de | |
| WG-02 | 52.06 ± 1.48 abc | 49.56 ± 2.01 a | 43.18 ± 1.39 bc | 35.90 ± 2.10 b | 24.06 ± 1.18 cd | |
| Hb + WG-11 | 48.91 ± 1.18 bc | 40.76 ± 1.59 b | 26.91 ± 2.13 f | 15.25 ± 1.14 e | 6.11 ± 0.90 h | |
| Hb + WG-02 | 47.11 ± 1.50 c | 43.53 ± 1.62 ab | 29.11 ± 1.00 ef | 19.71 ± 1.68 de | 9.58 ± 0.30 gh | |
| Sf + WG-11 | 51.10 ± 1.72 abc | 45.65 ± 1.04 ab | 35.78 ± 1.63 de | 23.10 ± 1.12 d | 13.11 ± 0.84 fg | |
| Sf + WG-02 | 50.13 ± 0.58 abc | 46.73 ± 1.47 ab | 38.21 ± 1.58 cd | 26.23 ± 1.54 cd | 17.36 ± 1.73 ef | |
| Control | 48.61 ± 1.15 bc | 50.45 ± 1.55 a | 52.35 ± 1.32 a | 53.98 ± 1.41 a | 54.58 ± 1.37 a | |
| df | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | |
| F | 4.54 | 3.42 | 29.92 | 50.30 | 137.84 | |
| p | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Adult | Hb | 36.61 ± 1.84 a | 34.70 ± 1.74 ab | 28.46 ± 2.73 abc | 25.30 ± 2.10 bc | 14.23 ± 1.48 bc |
| Sf | 34.08 ± 1.84 ab | 32.51 ± 1.78 ab | 29.65 ± 2.53 ab | 26.48 ± 2.47 b | 17.08 ± 1.40 b | |
| WG-11 | 35.06 ± 1.89 ab | 30.06 ± 2.17 ab | 25.48 ± 1.62 bcd | 23.30 ± 1.89 bcd | 11.08 ± 0.84 cd | |
| WG-02 | 32.78 ± 2.06 ab | 31.13 ± 2.24 ab | 27.43 ± 2.00 abc | 24.45 ± 1.97 bc | 13.70 ± 0.70 bc | |
| Hb + WG-11 | 30.68 ± 2.62 ab | 25.83 ± 2.12 b | 17.86 ± 1.52 d | 12.78 ± 1.35 e | 3.80 ± 0.42 e | |
| Hb + WG-02 | 32.13 ± 1.87 ab | 28.61 ± 2.00 ab | 20.93 ± 1.56 cd | 15.31 ± 1.19 de | 7.73 ± 0.54 de | |
| Sf + WG-11 | 33.31 ± 2.08 ab | 29.51 ± 1.64 ab | 22.68 ± 0.85 bcd | 17.63 ± 1.73 cde | 8.78 ± 0.53 d | |
| Sf + WG-02 | 27.55 ± 2.37 b | 30.93 ± 1.52 ab | 24.91 ± 1.88 bcd | 20.08 ± 1.96 bcde | 11.68 ± 0.93 cd | |
| Control | 35.33 ± 1.95 ab | 35.83 ± 2.23 a | 35.96 ± 2.26 a | 36.13 ± 1.90 a | 36.90 ± 1.91 a | |
| df | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | |
| F | 2.23 | 2.32 | 8.27 | 15.34 | 80.27 | |
| p | 0.04 | 0.03 | <0.01 | <0.01 | <0.01 |
| Season | Treatment | Leaves Length (cm) | Leaf Weight (g) | No. of Leaves | Neck Diameter (cm) | Bulb Diameter (cm) | No. of Rings/Bulb | Dry Matter (%) | Yield/Plant (g) |
|---|---|---|---|---|---|---|---|---|---|
| 2017–2018 | Hb | 40.00 ± 0.52 c | 15.45 ± 0.45 de | 12.33 ± 0.42 cd | 0.75 ± 0.05 a | 5.78 ± 0.34 bc | 65.01 ± 0.41 cd | 13.00 ± 0.48 c | 75.11 ± 0.59 ef |
| Sf | 39.35 ± 0.39 c | 15.41 ± 0.37 de | 11.21 ± 0.40 d | 0.73 ± 0.03 a | 5.45 ± 0.37 c | 65.48 ± 0.44 cd | 12.43 ± 0.43 c | 74.15 ± 0.46 ef | |
| WG-11 | 41.16 ± 0.44 bc | 16.43 ± 0.48 bcde | 13.30 ± 0.50 bc | 0.78 ± 0.04 a | 6.13 ± 0.47 abc | 66.25 ± 0.40 bcd | 14.06 ± 0.35 abc | 76.01 ± 0.47 de | |
| WG-02 | 41.28 ± 0.51 bc | 16.00 ± 0.45 cde | 12.01 ± 0.45 cd | 0.75 ± 0.04 a | 6.03 ± 0.55 abc | 65.01 ± 0.45 cd | 13.18 ± 0.45 bc | 75.05 ± 0.49 ef | |
| Hb + WG-11 | 44.31 ± 0.53 a | 19.25 ± 0.42 a | 15.01 ± 0.46 ab | 0.88 ± 0.04 a | 7.95 ± 0.45 a | 69.55 ± 0.48 a | 16.01 ± 0.51 a | 82.13 ± 0.41 a | |
| Hb + WG-02 | 43.30 ± 0.47 ab | 18.05 ± 0.51 abc | 15.53 ± 0.46 a | 0.85 ± 0.04 a | 7.63 ± 0.43 ab | 68.00 ± 0.40 ab | 15.20 ± 0.40 ab | 80.01 ± 0.48 ab | |
| Sf + WG-11 | 42.23 ± 0.51 ab | 18.46 ± 0.49 ab | 14.01 ± 0.48 abc | 0.83 ± 0.04 a | 6.93 ± 0.50 abc | 67.00 ± 0.49 bc | 15.56 ± 0.49 a | 78.40 ± 0.41 bc | |
| Sf + WG-02 | 42.40 ± 0.47 ab | 17.01 ± 0.41 bcd | 13.10 ± 0.41 bcd | 0.80 ± 0.04 a | 6.33 ± 0.51 abc | 66.38 ± 0.46 bcd | 14.15 ± 0.35 abc | 77.40 ± 0.42 cd | |
| Control | 39.28 ± 0.39 c | 14.36 ± 0.41 e | 11.06 ± 0.37 d | 0.71 ± 0.04 a | 5.06 ± 0.40 c | 64.60 ± 0.51 d | 12.25 ± 0.44 c | 73.36 ± 0.41 f | |
| df | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | |
| F | 13.4 | 12.7 | 12.6 | 1.58 | 4.50 | 12.5 | 9.75 | 38.3 | |
| p | <0.01 | <0.01 | <0.01 | 0.15 | <0.01 | <0.01 | <0.01 | <0.01 | |
| 2018–2019 | Hb | 38.05 ± 0.62 de | 14.53 ± 0.42 cd | 11.13 ± 0.37 bc | 0.71 ± 0.04 a | 5.60 ± 0.40 abc | 63.65 ± 0.55 def | 13.26 ± 0.44 cde | 73.21 ± 0.47 ef |
| Sf | 37.08 ± 0.39 e | 14.25 ± 0.45 cd | 10.48 ± 0.42 c | 0.70 ± 0.03 a | 5.26 ± 0.38 bc | 62.93 ± 0.73 ef | 12.45 ± 0.43 de | 71.25 ± 0.39 fg | |
| WG-11 | 41.38 ± 0.43 bc | 16.01 ± 0.36 bc | 11.25 ± 0.47 bc | 0.75 ± 0.05 a | 5.98 ± 0.34 abc | 65.41 ± 0.45 bcd | 13.80 ± 0.46 abcde | 75.00 ± 0.51 cde | |
| WG-02 | 40.05 ± 0.38 cd | 15.03 ± 0.42 c | 12.20 ± 0.52 bc | 0.73 ± 0.05 a | 5.80 ± 0.40 abc | 64.81 ± 0.42 cde | 13.61 ± 0.47 bcde | 74.15 ± 0.36 de | |
| Hb + WG-11 | 44.38 ± 0.46 a | 19.01 ± 0.37 a | 15.10 ± 0.52 a | 0.83 ± 0.04 a | 7.45 ± 0.45 a | 68.76 ± 0.52 a | 15.75 ± 0.41 a | 80.26 ± 0.42 a | |
| Hb + WG-02 | 43.15 ± 0.38 ab | 18.20 ± 0.44 a | 14.38 ± 0.41 a | 0.80 ± 0.03 a | 7.28 ± 0.44 ab | 68.11 ± 0.46 a | 15.33 ± 0.35 ab | 78.10 ± 0.40 b | |
| Sf + WG-11 | 43.53 ± 0.40 a | 18.51 ± 0.47 a | 14.55 ± 0.36 a | 0.78 ± 0.04 a | 6.73 ± 0.48 abc | 67.36 ± 0.43 ab | 14.85 ± 0.44 abc | 77.03 ± 0.48 bc | |
| Sf + WG-02 | 42.46 ± 0.45 ab | 17.38 ± 0.37 ab | 13.01 ± 0.49 ab | 0.76 ± 0.04 a | 6.45 ± 0.46 abc | 66.88 ± 0.43 abc | 14.16 ± 0.40 abcd | 76.18 ± 0.41 bcd | |
| Control | 37.26 ± 0.38 e | 13.05 ± 0.42 d | 10.45 ± 0.44 c | 0.68 ± 0.03 a | 5.03 ± 0.51 c | 62.40 ± 0.47 f | 12.15 ± 0.43 e | 71.03 ± 0.49 g | |
| df | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | 8, 53 | |
| F | 40.3 | 26.0 | 16.2 | 1.11 | 3.83 | 20.8 | 8.14 | 48.1 | |
| p | <0.01 | <0.01 | <0.01 | 0.37 | <0.01 | <0.01 | <0.01 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gulzar, S.; Wakil, W.; Shapiro-Ilan, D.I. Combined Effect of Entomopathogens against Thrips tabaci Lindeman (Thysanoptera: Thripidae): Laboratory, Greenhouse and Field Trials. Insects 2021, 12, 456. https://doi.org/10.3390/insects12050456
Gulzar S, Wakil W, Shapiro-Ilan DI. Combined Effect of Entomopathogens against Thrips tabaci Lindeman (Thysanoptera: Thripidae): Laboratory, Greenhouse and Field Trials. Insects. 2021; 12(5):456. https://doi.org/10.3390/insects12050456
Chicago/Turabian StyleGulzar, Sehrish, Waqas Wakil, and David I. Shapiro-Ilan. 2021. "Combined Effect of Entomopathogens against Thrips tabaci Lindeman (Thysanoptera: Thripidae): Laboratory, Greenhouse and Field Trials" Insects 12, no. 5: 456. https://doi.org/10.3390/insects12050456
APA StyleGulzar, S., Wakil, W., & Shapiro-Ilan, D. I. (2021). Combined Effect of Entomopathogens against Thrips tabaci Lindeman (Thysanoptera: Thripidae): Laboratory, Greenhouse and Field Trials. Insects, 12(5), 456. https://doi.org/10.3390/insects12050456