
Effects of Functional Depletion of Doublesex on Male Development in the Sawfly, Athalia rosae

 and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. RNA Extraction and RT-PCR
2.3. Quantitative Real-Time RT-PCR
2.4. Preparation of dsRNAs
2.5. Injection of dsRNA into Insects
2.6. Observation of Internal Reproductive and Genital Organs
2.7. Behavioral Assays
2.8. Statictics
3. Results
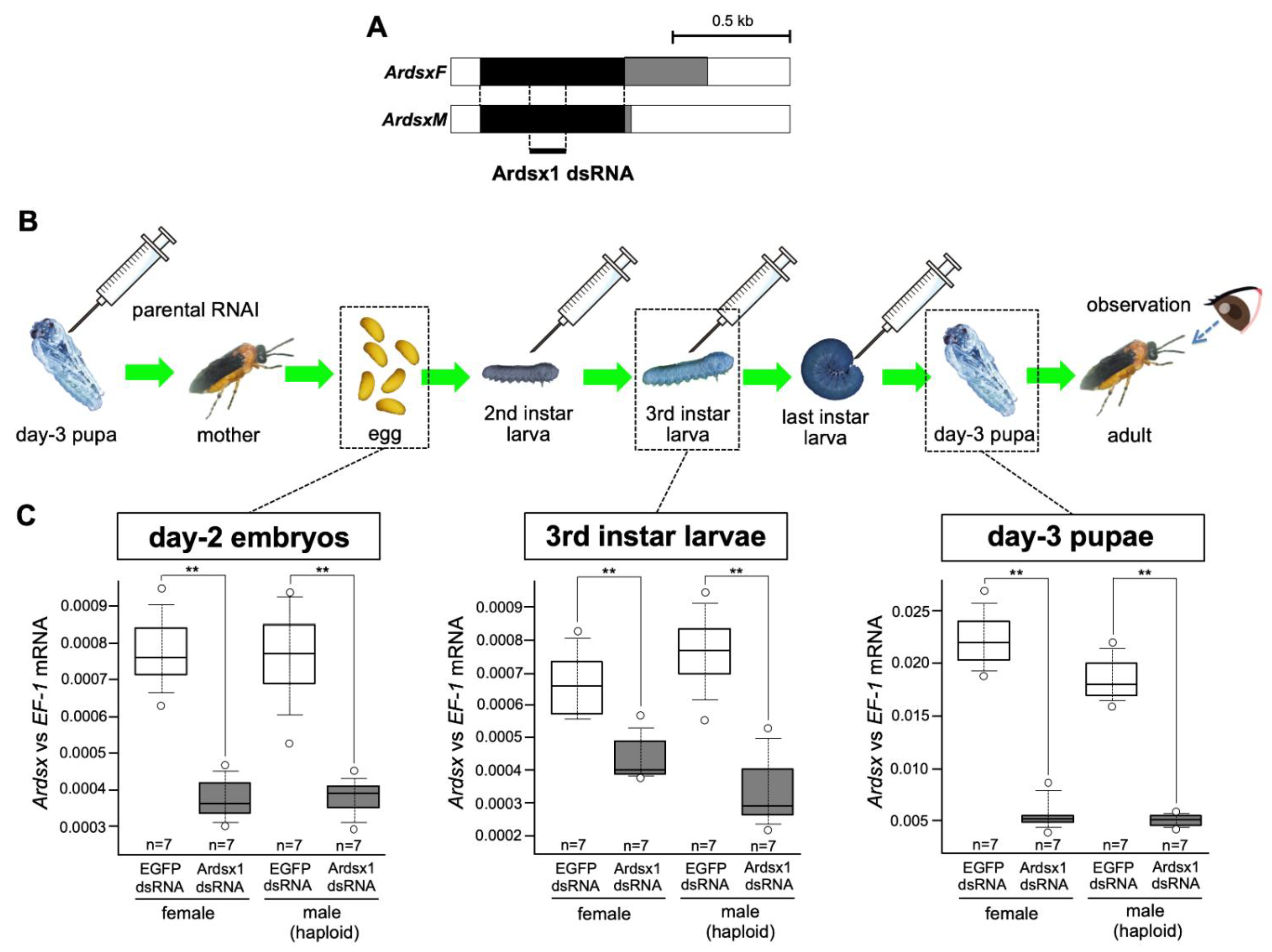
3.1. Effects of Repeated Injections of Ardsx dsRNAs on Ardsx Expression
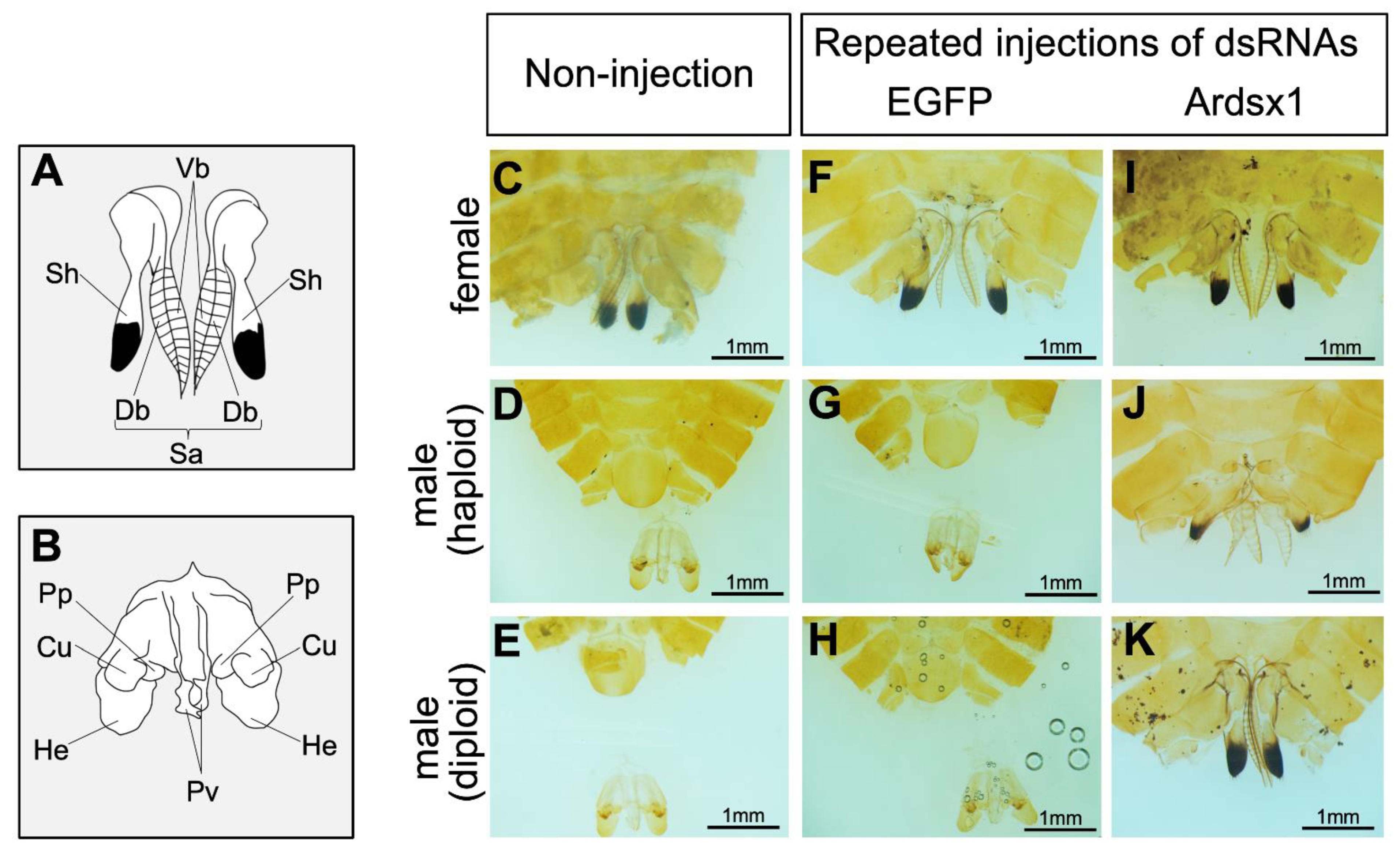
3.2. Effects of Knockdown of Ardsx Expression on Sexual Differentiation of Genitalia
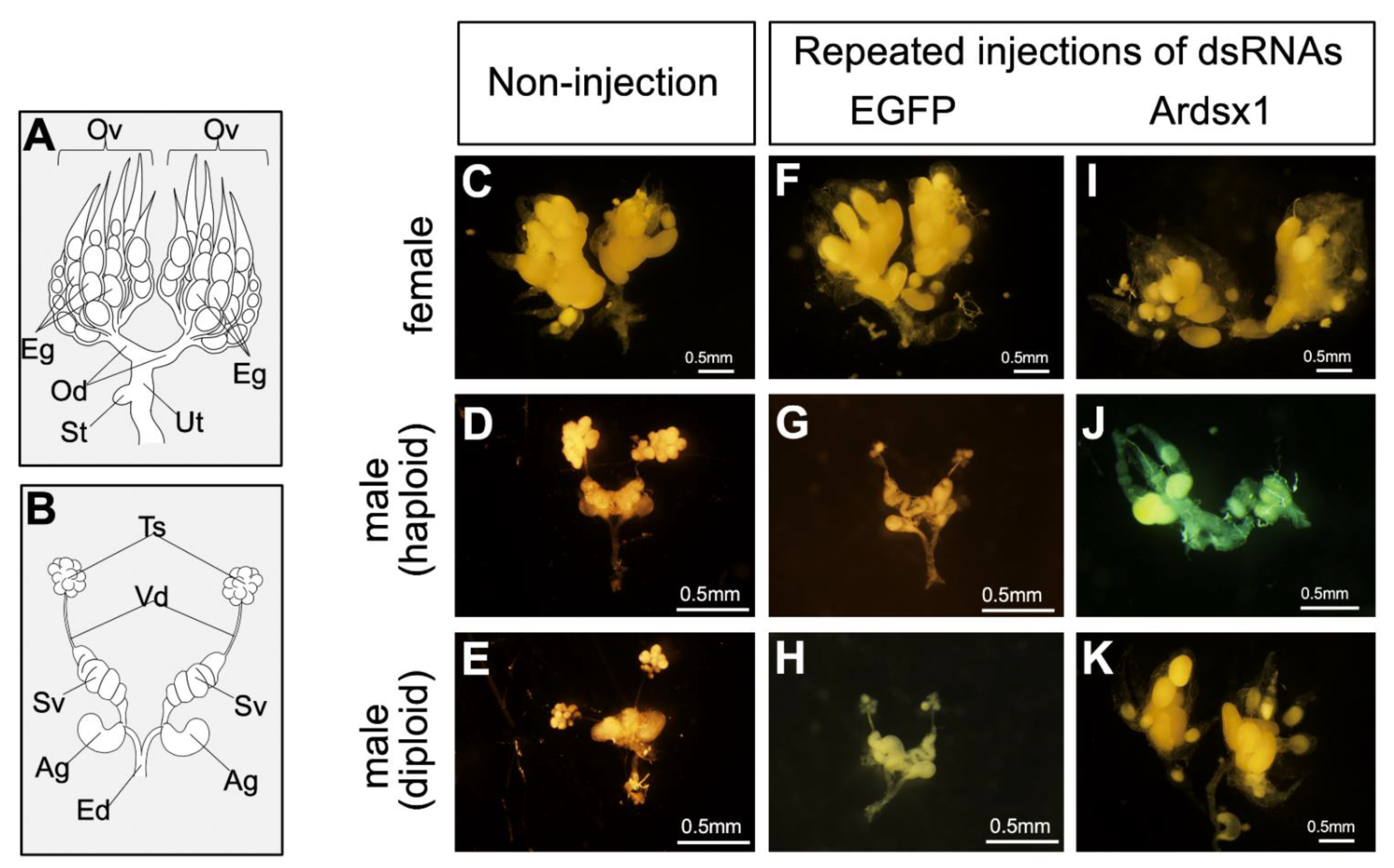
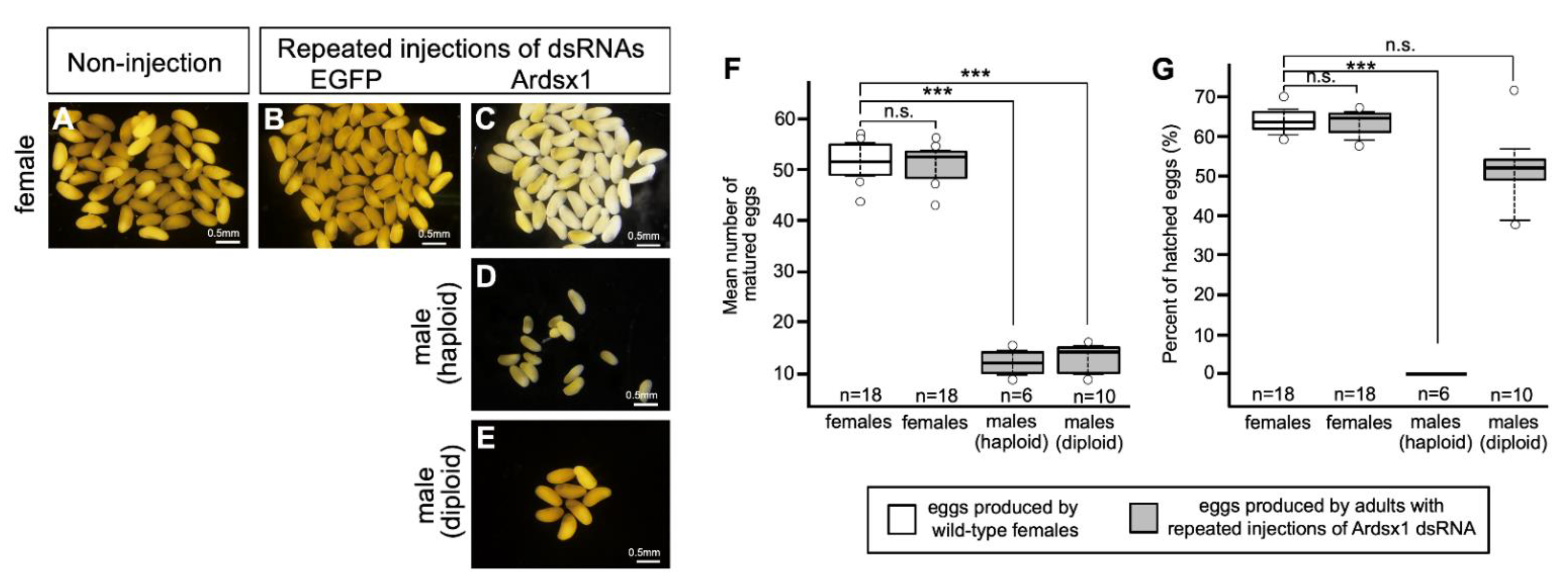
3.3. Effects of Knockdown of Ardsx Expression on Oogenesis
3.4. Effects of Ardsx Knockdown on Sexual Behavior
4. Discussion

5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herpin, A.; Schartl, M. Plasticity of gene-regulatory networks controlling sex determination: Of masters, slaves, usual suspects, newcomers, and usurpators. EMBO Rep. 2015, 16, 1260–1274. [Google Scholar] [CrossRef]
- Verhulst, E.C.; van de Zande, L. Double nexus-Doublesex is the connecting element in sex determination. Brief Funct. Genom. 2015, 14, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.G. Sex determination cascade in insects: A great treasure house of alternative splicing. In Reproductive and Developmental Strategies; Kobayashi, K., Kitano, T., Iwao, Y., Kondo, M., Eds.; Springer: Tokyo, Japan, 2018; pp. 267–288. [Google Scholar]
- Kato, Y.; Kobayashi, K.; Watanabe, H.; Iguchi, T. Environmental sex determination in the branchiopod crustacean Daphnia magna: Deep conservation of a Doublesex gene in the sex-determining pathway. PLoS Genet. 2011, 7, e1001345. [Google Scholar] [CrossRef] [PubMed]
- Toyota, K.; Kato, Y.; Sato, M.; Sugiura, N.; Miyagawa, S.; Miyakawa, H.; Watanabe, H.; Oda, S.; Ogino, Y.; Hiruta, C.; et al. Molecular cloning of doublesex genes of four Cladocera (water flea) species. BMC Genom. 2013, 14, 239. [Google Scholar] [CrossRef]
- Schütt, C.; Nöthiger, R. Structure, function and evolution of sex-determining systems in Dipteran insects. Development 2000, 127, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.S.; Wolfner, M.F. A molecular analysis of doublesex, a bifunctional gene that controls both male and female sexual differentiation in Drosophila melanogaster. Genes Dev. 1988, 2, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Burtis, K.C.; Baker, B.S. Drosophila doublesex gene controls somatic sexual differentiation by producing alternatively spliced mRNAs encoding related sex-specific polypeptides. Cell 1989, 56, 997–1010. [Google Scholar] [CrossRef]
- Cline, T.W.; Meyer, B.J. Vive la difference: Males vs. females in flies vs. worms. Annu. Rev. Genet. 1996, 30, 637–702. [Google Scholar] [CrossRef]
- Hildreth, P.E. Doublesex, recessive gene that transforms both males and females of Drosophila into intersexes. Genetics 1965, 51, 659–678. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, Y.; Li, Z.; Ling, L.; Zeng, B.; James, A.A.; Tan, A.; Huang, Y. Transcription activator-like effector nuclease (TALEN)-mediated female-specific sterility in the silkworm, Bombyx mori. Insect Mol. Biol. 2014, 23, 800–807. [Google Scholar] [CrossRef]
- Xu, J.; Zhan, S.; Chen, S.; Zeng, B.; Li, Z.; James, A.A.; Tan, A.; Huang, Y. Sexually dimorphic traits in the silkworm, Bombyx mori, are regulated by doublesex. Insect Biochem. Mol. Biol. 2017, 80, 42–51. [Google Scholar] [CrossRef]
- Ito, Y.; Harigai, A.; Nakata, M.; Hosoya, T.; Araya, K.; Oba, Y.; Ito, A.; Ohde, T.; Yaginuma, T.; Niimi, T. The role of doublesex in the evolution of exaggerated horns in the Japanese rhinoceros beetle. EMBO Rep. 2013, 14, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.C.; Werren, J.H.; Verhulst, E.C.; Giebel, J.D.; Kamping, A.; Beukeboom, L.W.; van de Zande, L. Identification and characterization of the doublesex gene of Nasonia. Insect. Mol. Biol. 2009, 18, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Shukla, J.N.; Nagaraju, J. Two female-specific DSX proteins are encoded by the sex- specific transcripts of dsx, and are required for female sexual differentiation in two wild silkmoth species, Antheraea assama and Antheraea mylitta (Lepidoptera, Saturniidae). Insect Biochem. Mol. Biol. 2010, 40, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Shukla, J.N.; Palli, S.R. Doublesex target genes in the red flour beetle, Tribolium castaneum. Sci. Rep. 2012, 2, 948. [Google Scholar] [CrossRef]
- Mysore, K.; Sun, L.; Tomchaney, M.; Sullivan, G.; Adams, H.; Piscoya, A.S.; Severson, D.W.; Syed, Z.; Duman-Scheel, M. siRNA-mediated silencing of doublesex during female development of the Dengue vector mosquito Aedes aegypti. PLoS Negl. Trop. Dis. 2015, 9, e0004213. [Google Scholar] [CrossRef]
- Kyrou, K.; Hammond, A.M.; Galizi, R.; Kranjc, N.; Burt, A.; Beaghton, A.K.; Nolan, T.; Crisanti, A. A CRISPR-Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nat. Biotechnol. 2018, 36, 1062–1066. [Google Scholar] [CrossRef]
- Zhuo, J.C.; Hu, Q.L.; Zhang, H.H.; Zhang, M.Q.; Jo, S.B.; Zhang, C.X. Identification and functional analysis of the doublesex gene in the sexual development of a hemimetabolous insect, the brown planthopper. Insect Biochem. Mol. Biol. 2018, 102, 31–42. [Google Scholar] [CrossRef]
- McAfee, A.; Pettis, J.S.; Tarpy, D.R.; Foster, L.J. Feminizer and doublesex knock-outs cause honey bees to switch sexes. PLoS Biol. 2019, 17, e3000256. [Google Scholar] [CrossRef]
- Whiting, P.W. Selective fertilization and sex determination in Hymenoptera. Science 1933, 78, 537–538. [Google Scholar] [CrossRef]
- Naito, T.; Suzuki, H. Sex determination in the sawfly, Athalia rosae ruficornis (Hymenoptera): Occurrence of triploid males. J. Hered. 1991, 82, 101–104. [Google Scholar] [CrossRef]
- Mine, S.; Sumitani, M.; Aoki, F.; Hatakeyama, M.; Suzuki, M.G. Identification and functional characterization of the sex-determining gene doublesex in the sawfly, Athalia rosae. Appl. Entomol. Zool. 2017, 52, 497–509. [Google Scholar] [CrossRef]
- Yoshiyama, N.; Tojo, K.; Hatakeyama, M. A survey of the effectiveness of non-cell autonomous RNAi throughout development in the sawfly, Athalia rosae (Hymenoptera). J. Insect Physiol. 2013, 59, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Sawa, M.; Oishi, K. Studies on the sawfly, Athalia rosae (Insecta, Hymenoptera, Tenthredinidae). III. Fertilization by sperm injection. Zool. Sci. 1989, 6, 557–563. [Google Scholar]
- Suzuki, M.G.; Suzuki, K.; Aoki, F.; Ajimura, M. Effect of RNAi-mediated knockdown of the Bombyx mori transformer-2 gene on the sex-specific splicing of Bmdsx pre-mRNA. Int. J. Dev. Biol. 2012, 56, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Amano, T.; Nishida, R.; Kuwahara, Y.; Fukami, H. Pharmacophagous acquisition of clerodendrins by the turnip sawfly (Athalia rosae ruficornis) and their role in the mating behaviour. Chemoecology 1999, 9, 145–150. [Google Scholar] [CrossRef]
- Awane, K.; Kitano, H. Studies on the effect of Clerodendron trichotomum Thunb. on the mating behavior of Athalia infumata (Marlatt) (Hymenoptera: Tenthredinidae). Jpn. J. Appl. Entomol. Zool. 1992, 36, 13–16. [Google Scholar] [CrossRef][Green Version]
- Kitano, H. Experimental studies on the mating behavior of Athalia lugens infumata. Kontyu 1988, 56, 180–188. [Google Scholar]
- Sekine, K.; Furusawa, T.; Hatakeyama, M. The boule gene is essential for spermatogenesis of haploid insect male. Dev. Biol. 2015, 399, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.G.; Ohbayashi, F.; Mita, K.; Shimada, T. The mechanism of sex-specific splicing at the doublesex gene is different between Drosophila melanogaster and Bombyx mori. Insect Biochem. Mol. Biol. 2001, 31, 1201–1211. [Google Scholar] [CrossRef]
- Suzuki, M.G.; Funaguma, S.; Kanda, T.; Tamura, T.; Shimada, T. Analysis of the biological functions of a doublesex homologue in Bombyx mori. Dev. Genes Evol. 2003, 213, 345–354. [Google Scholar] [CrossRef]
- Suzuki, M.G.; Funaguma, S.; Kanda, T.; Tamura, T.; Shimada, T. Role of the male BmDSX protein in the sexual differentiation of Bombyx mori. Evol. Dev. 2005, 7, 58–68. [Google Scholar] [CrossRef]
- Scali, C.; Catteruccia, F.; Li, Q.; Crisanti, A. Identification of sex-specific transcripts of the Anopheles gambiae doublesex gene. J. Exp. Biol. 2005, 208, 3701–3709. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Huang, Z.Y.; Zhang, J. Sex-specific splicing of the honeybee doublesex gene reveals 300 million years of evolution at the bottom of the insect sex-determination pathway. Genetics 2007, 177, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Price, D.C.; Egizi, A.; Fonseca, D.M. The ubiquity and ancestry of insect doublesex. Sci. Rep. 2015, 5, 13068. [Google Scholar] [CrossRef] [PubMed]
- Matson, C.K.; Zarkower, D. Sex and the singular DM domain: Insights into sexual regulation, evolution and plasticity. Nat. Rev. Genet. 2012, 13, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Ledón-Rettig, C.C.; Zattara, E.E.; Moczek, A.P. Asymmetric interactions between doublesex and tissue- and sex-specific target genes mediate sexual dimorphism in beetles. Nat. Commun. 2017, 8, 14593. [Google Scholar] [CrossRef]
- Wexler, J.; Delaney, E.K.; Belles, X.; Schal, C.; Wada-Katsumata, A.; Amicucci, M.J.; Kopp, A. Hemimetabolous insects elucidate the origin of sexual development via alternative splicing. eLife 2019, 8, e47490. [Google Scholar] [CrossRef]
- Savard, J.; Tautz, D.; Richards, S.; Weinstock, G.M.; Gibbs, R.A.; Werren, J.H.; Tettelin, H.; Lercher, M.J. Phylogenomic analysis reveals bees and wasps (Hymenoptera) at the base of the radiation of Holometabolous insects. Genome Res. 2006, 16, 1334–1338. [Google Scholar] [CrossRef]
- Trautwein, M.D.; Wiegmann, B.M.; Beutel, R.; Kjer, K.M.; Yeates, D.K. Advances in insect phylogeny at the dawn of the post-genomic era. Annu. Rev. Entomol. 2012, 57, 449–468. [Google Scholar] [CrossRef]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.S.; Meusemann, K.; Petersen, M.; Mayer, C.; Wilbrandt, J.; Ziesmann, T.; Donath, A.; Kjer, K.M.; Aspöck, U.; Aspöck, H.; et al. The evolutionary history of holometabolous insects inferred from transcriptome-based phylogeny and comprehensive morphological data. BMC Evol. Biol. 2014, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Casper, A.; Van Doren, M. The control of sexual identity in the Drosophila germline. Development 2006, 133, 2783–2791. [Google Scholar] [CrossRef]
- Hempel, L.U.; Kalamegham, R.; Smith, J.E., 3rd; Oliver, B. Drosophila germline sex determination: Integration of germline autonomous cues and somatic signals. Curr. Top. Dev. Biol. 2008, 83, 109–150. [Google Scholar] [PubMed]
- Murray, M.; Yang, Y.; Van, M. Germ cell sex determination: A collaboration between soma and germline. Curr. Opin. Cell. Biol. 2010, 22, 722–729. [Google Scholar] [CrossRef]
- Ryner, L.C.; Goodwin, S.F.; Castrillon, D.H.; Anand, A.; Villella, A.; Baker, B.S.; Hall, J.C.; Taylor, B.J.; Wasserman, S.A. Control of male sexual behavior and sexual orientation in Drosophila by the fruitless gene. Cell 1996, 87, 1079–1089. [Google Scholar] [CrossRef]
- Heinrichs, V.; Ryner, L.C.; Baker, B.S. Regulation of sex-specific selection of fruitless 5′ splice sites by transformer and transformer-2. Mol. Cell. Biol. 1998, 18, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Usui-Aoki, K.; Ito, H.; Ui-Tei, K.; Takahashi, K.; Lukacsovich, T.; Awano, W.; Nakata, H.; Piao, Z.F.; Nilsson, E.E.; Tomida, J.; et al. Formation of the male-specific muscle in female Drosophila by ectopic fruitless expression. Nat. Cell Biol. 2000, 2, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Fujitani, K.; Usui, K.; Shimizu-Nishikawa, K.; Tanaka, S.; Yamamoto, D. Sexual orientation in Drosophila is altered by the satori mutation in the sex-determination gene fruitless that encodes a zinc finger protein with a BTB domain. Proc. Natl. Acad. Sci. USA 1996, 93, 9687–9692. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Villella, A.; Ryner, L.C.; Carlo, T.; Goodwin, S.F.; Song, H.J.; Gailey, D.A.; Morales, A.; Hall, J.C.; Baker, B.S.; et al. Molecular genetic dissection of the sex-specific and vital functions of the Drosophila melanogaster sex determination gene fruitless. Genetics 2001, 158, 1569–1595. [Google Scholar] [CrossRef]
- Lee, G.; Hall, J.C. Abnormalities of male-specific FRU protein and serotonin expression in the CNS of fruitless mutants in Drosophila. J. Neurosci. 2001, 21, 513–526. [Google Scholar] [CrossRef]
- Lee, G.; Villella, A.; Taylor, B.J.; Hall, J.C. New reproductive anomalies in fruitless-mutant Drosophila males: Extreme lengthening of mating durations and infertility correlated with defective serotonergic innervation of reproductive organs. J. Neurobiol. 2001, 47, 121–149. [Google Scholar] [CrossRef]
- Demir, E.; Dickson, B.J. Fruitless splicing specifies male courtship behavior in Drosophila. Cell 2005, 121, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Gailey, D.A.; Billeter, J.C.; Liu, J.H.; Bauzon, F.; Allendorfer, J.B.; Goodwin, S.F. Functional conservation of the fruitless male sex-determination gene across 250 Myr of insect evolution. Mol. Biol. Evol. 2006, 23, 633–643. [Google Scholar] [CrossRef]
- Bertossa, R.C.; van de Zande, L.; Beukeboom, L.W. The Fruitless gene in Nasonia displays complex sex-specific splicing and contains new zinc finger domains. Mol. Biol. Evol. 2009, 26, 1557–1569. [Google Scholar] [CrossRef] [PubMed]
- Boerjan, B.; Tobback, J.; De Loof, A.; Schoofs, L.; Huybrechts, R. Fruitless RNAi knockdown in males interferes with copulation success in Schistocerca gregaria. Insect Biochem. Mol. Biol. 2011, 41, 340–347. [Google Scholar] [CrossRef]
- Clynen, E.; Ciudad, L.; Bellés, X.; Piulachs, M.D. Conservation of fruitless’ role as master regulator of male courtship behavior from cockroaches to flies. Dev. Genes. Evol. 2011, 221, 43–48. [Google Scholar] [CrossRef]
- Meier, N.; Käppeli, S.C.; Hediger, N.M.; Billeter, J.C.; Goodwin, S.F.; Bopp, D. Genetic control of courtship behavior in the housefly: Evidence for a conserved bifurcation of the sex-determining pathway. PLoS ONE 2013, 8, e62476. [Google Scholar] [CrossRef]
- Salvemini, M.; D’Amato, R.; Petrella, V.; Aceto, S.; Nimmo, D.; Neira, M.; Alphey, L.; Polito, L.C.; Saccone, G. The orthologue of the fruitfly sex behaviour gene fruitless in the mosquito Aedes aegypti: Evolution of genomic organisation and alternative splicing. PLoS ONE 2013, 8, e48554. [Google Scholar] [CrossRef] [PubMed]
- Villella, A.; Hall, J.C. Courtship anomalies caused by doublesex mutations in Drosophila melanogaster. Genetics 1996, 143, 331–344. [Google Scholar] [CrossRef]
- Rideout, E.J.; Billeter, J.C.; Goodwin, S.F. The sex-determination genes fruitless and doublesex specify a neural substrate required for courtship song. Curr. Biol. 2007, 17, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Baker, B.S. Genetic identification and separation of innate and experience-dependent courtship behaviors in Drosophila. Cell 2014, 156, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Billeter, J.C.; Villella, A.; Allendorfer, J.B.; Dornan, A.J.; Richardson, M.; Gailey, D.A.; Goodwin, S.F. Isoform-specific control of male neuronal differentiation and behavior in Drosophila by the fruitless gene. Curr. Biol. 2006, 16, 1063–1076. [Google Scholar] [CrossRef] [PubMed]
- Beukeboom, L.W.; Kamping, A.; van de Zande, L. Sex determination in the haplodiploid wasp Nasonia vitripennis (Hymenoptera: Chalcidoidea): A critical consideration of models and evidence. Semin. Cell. Dev. Biol. 2007, 18, 371–378. [Google Scholar] [CrossRef]
- Hasselmann, M.; Gempe, T.; Schiøtt, M.; Nunes-Silva, C.G.; Otte, M.; Beye, M. Evidence for the evolutionary nascence of a novel sex determination pathway in honeybees. Nature 2008, 454, 519–522. [Google Scholar] [CrossRef]
- Verhulst, E.C.; Beukeboom, L.W.; van de Zande, L. Maternal control of haplodiploid sex determination in the wasp Nasonia. Science 2010, 328, 620–623. [Google Scholar] [CrossRef]
- The Honeybee Genome Sequencing Consortium. Insights into social insects from the genome of the honeybee Apis mellifera. Nature 2006, 443, 931–949. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
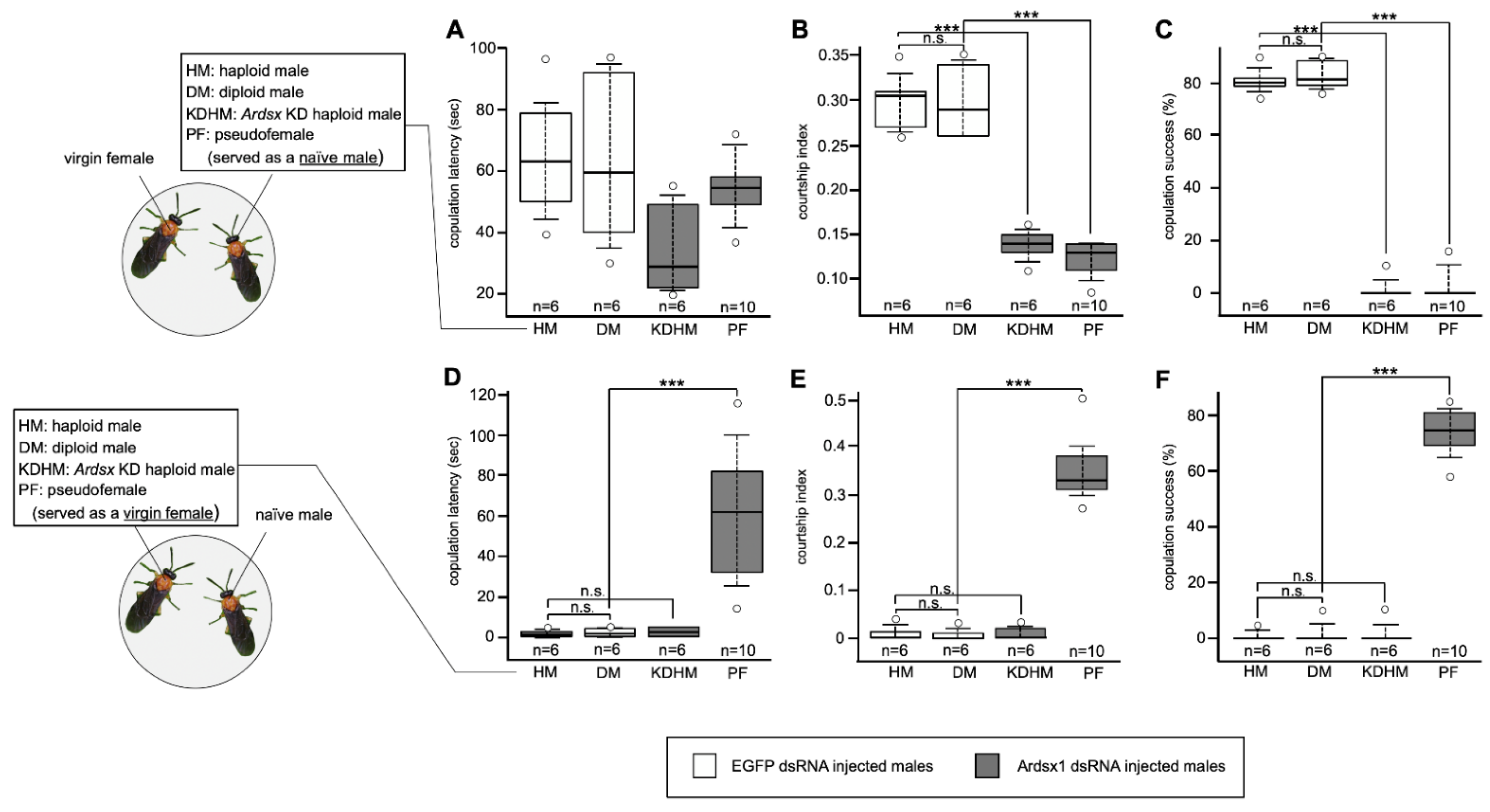{kind=link}
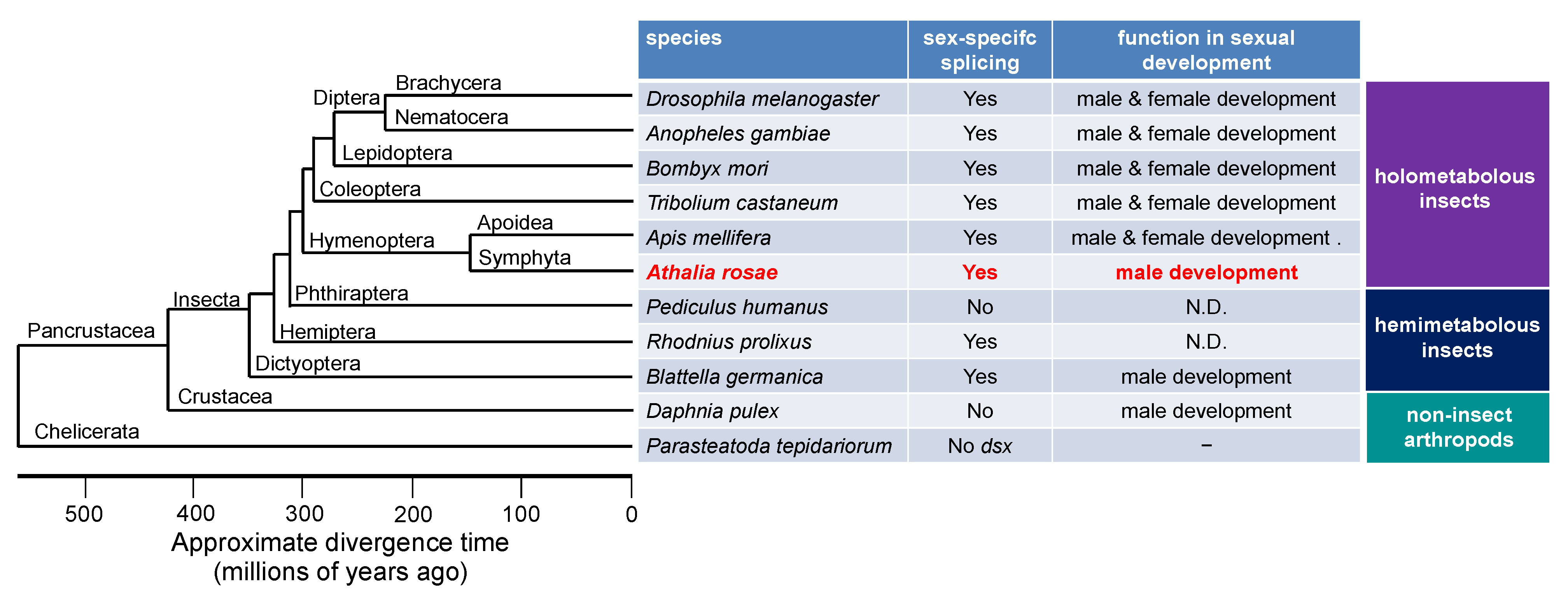{kind=link}
| dsRNA | Sexuality | No. of Injected Animals | No. of Investigated Adults | Sexual Phenotypes *2 | |||
|---|---|---|---|---|---|---|---|
| 2nd Instar | 3rd Instar | Last Instar | Normal | Sex-Reversal | |||
| EGFP | female | 50 | 26 | 19 | 12 | 12 | 0 |
| male (haploid) | 63 | 27 | 20 | 13 | 13 | 0 | |
| male (diploid) | N/A *1 | N/A | N/A | 7 | 7 | 0 | |
| Ardsx1 | female | 59 | 35 | 26 | 18 | 18 | 0 |
| male (haploid) | 54 | 29 | 20 | 13 | 1 | 12 | |
| male (diploid) | N/A | N/A | N/A | 12 | 1 | 11 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mine, S.; Sumitani, M.; Aoki, F.; Hatakeyama, M.; Suzuki, M.G. Effects of Functional Depletion of Doublesex on Male Development in the Sawfly, Athalia rosae. Insects 2021, 12, 849. https://doi.org/10.3390/insects12100849
Mine S, Sumitani M, Aoki F, Hatakeyama M, Suzuki MG. Effects of Functional Depletion of Doublesex on Male Development in the Sawfly, Athalia rosae. Insects. 2021; 12(10):849. https://doi.org/10.3390/insects12100849
Chicago/Turabian StyleMine, Shotaro, Megumi Sumitani, Fugaku Aoki, Masatsugu Hatakeyama, and Masataka G. Suzuki. 2021. "Effects of Functional Depletion of Doublesex on Male Development in the Sawfly, Athalia rosae" Insects 12, no. 10: 849. https://doi.org/10.3390/insects12100849
APA StyleMine, S., Sumitani, M., Aoki, F., Hatakeyama, M., & Suzuki, M. G. (2021). Effects of Functional Depletion of Doublesex on Male Development in the Sawfly, Athalia rosae. Insects, 12(10), 849. https://doi.org/10.3390/insects12100849