
Inhibition of Pathological Increased Matrix Metalloproteinase (MMP) Activity for Improvement of Bone Regeneration in Diabetes

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
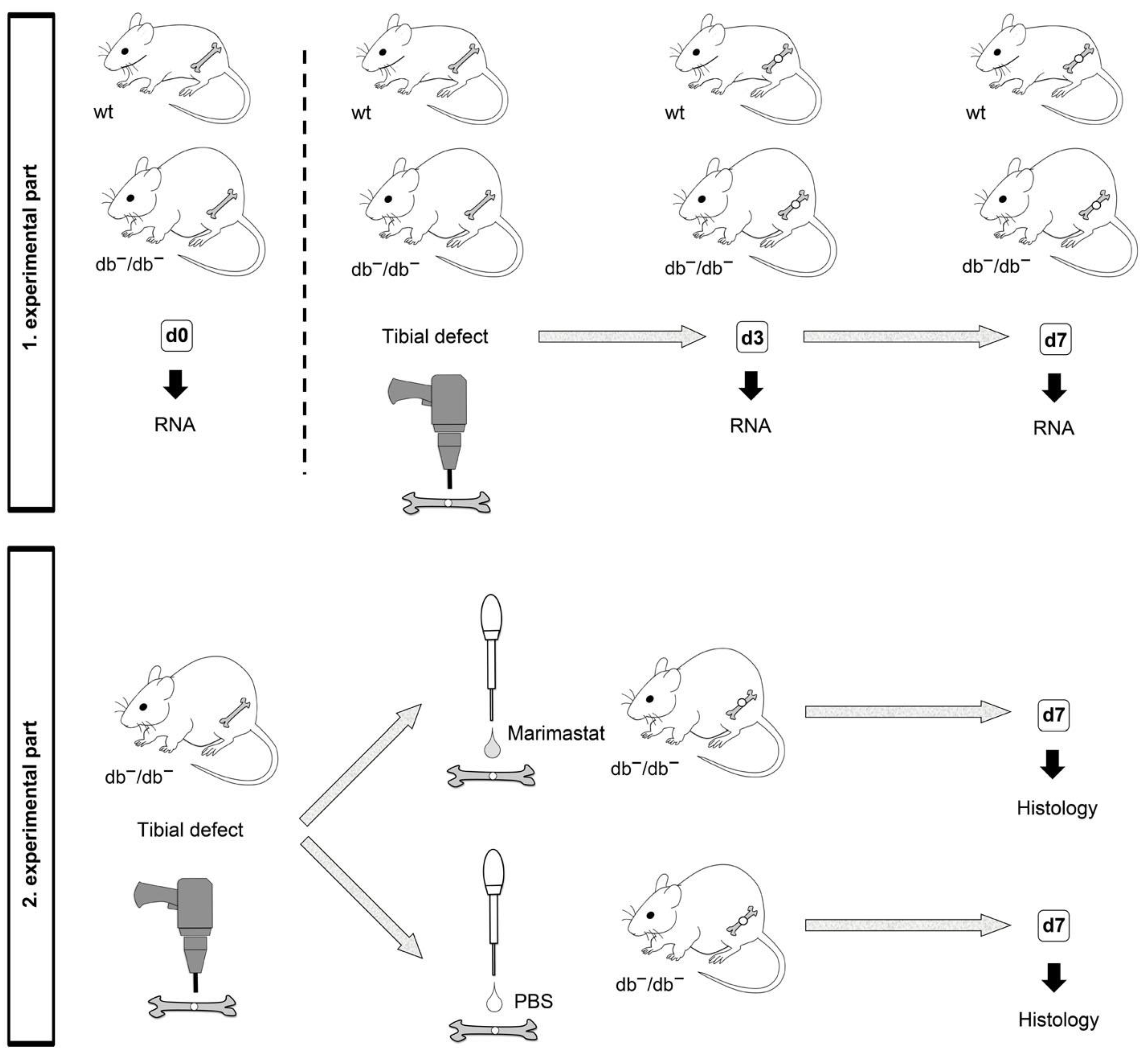
2.1. Surgery of Animals
2.2. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
2.3. Preparation of Tissues and Characterization of Bone Formation after Marimastat Application
2.4. Histological Analyses
2.5. Tartrate-Resistant Acid Phosphatase (TRAP) Staining
2.6. Statistical Analysis
3. Results
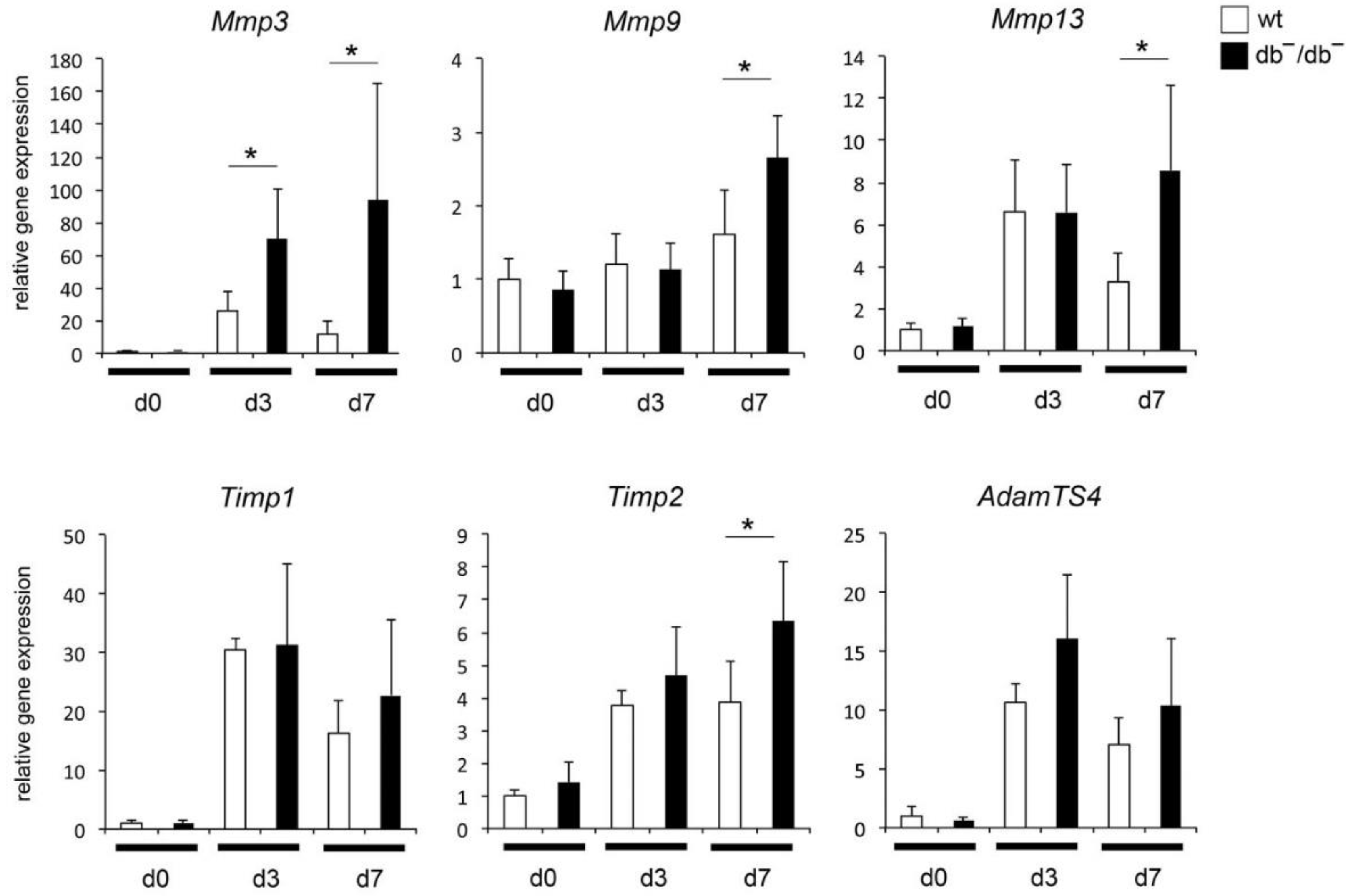
3.1. Increased Expression of MMP-Related Genes in Regenerating Bones of db−/db− Mice
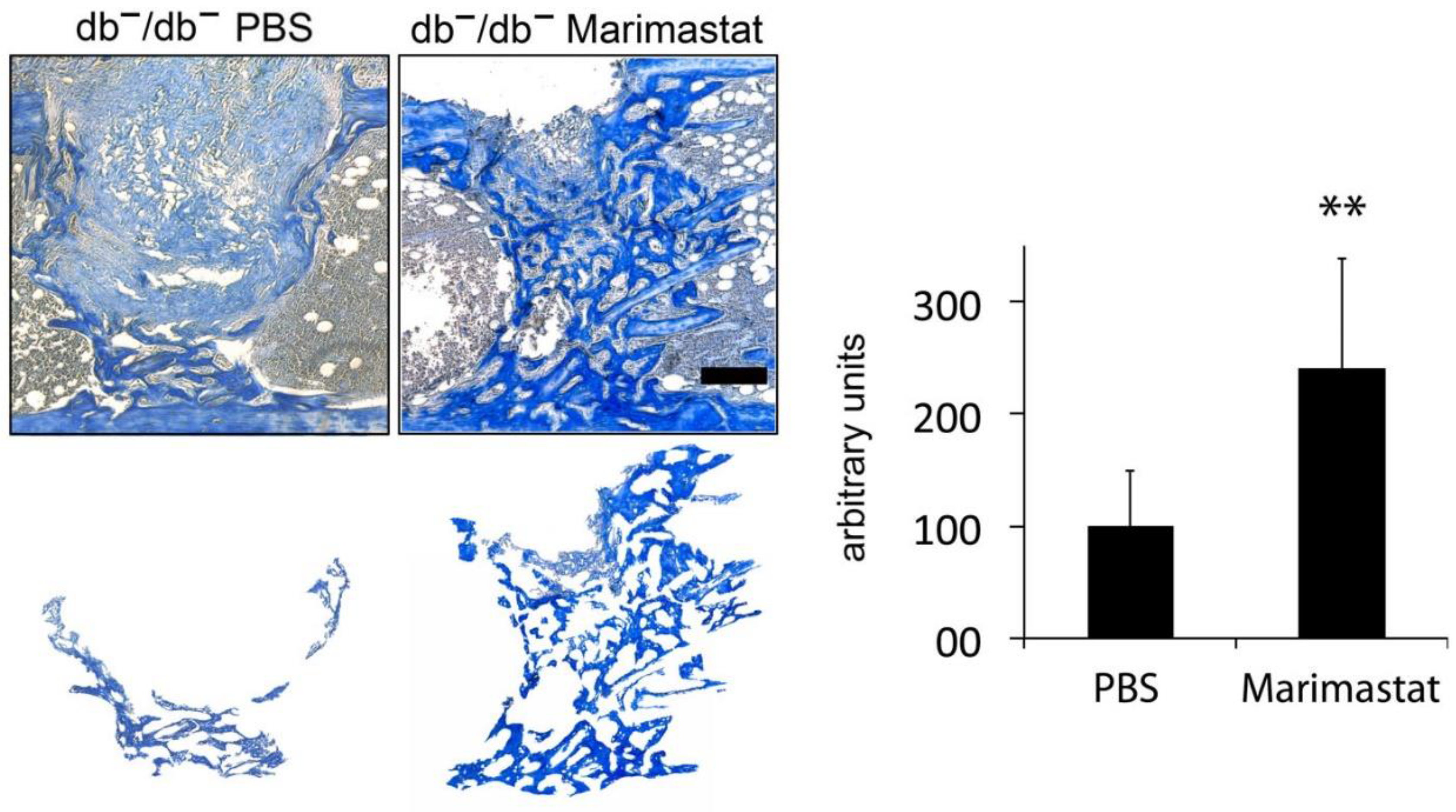
3.2. Enhanced New Bone Formation by Reducing MMP Activity in Bones of db−/db− Mice
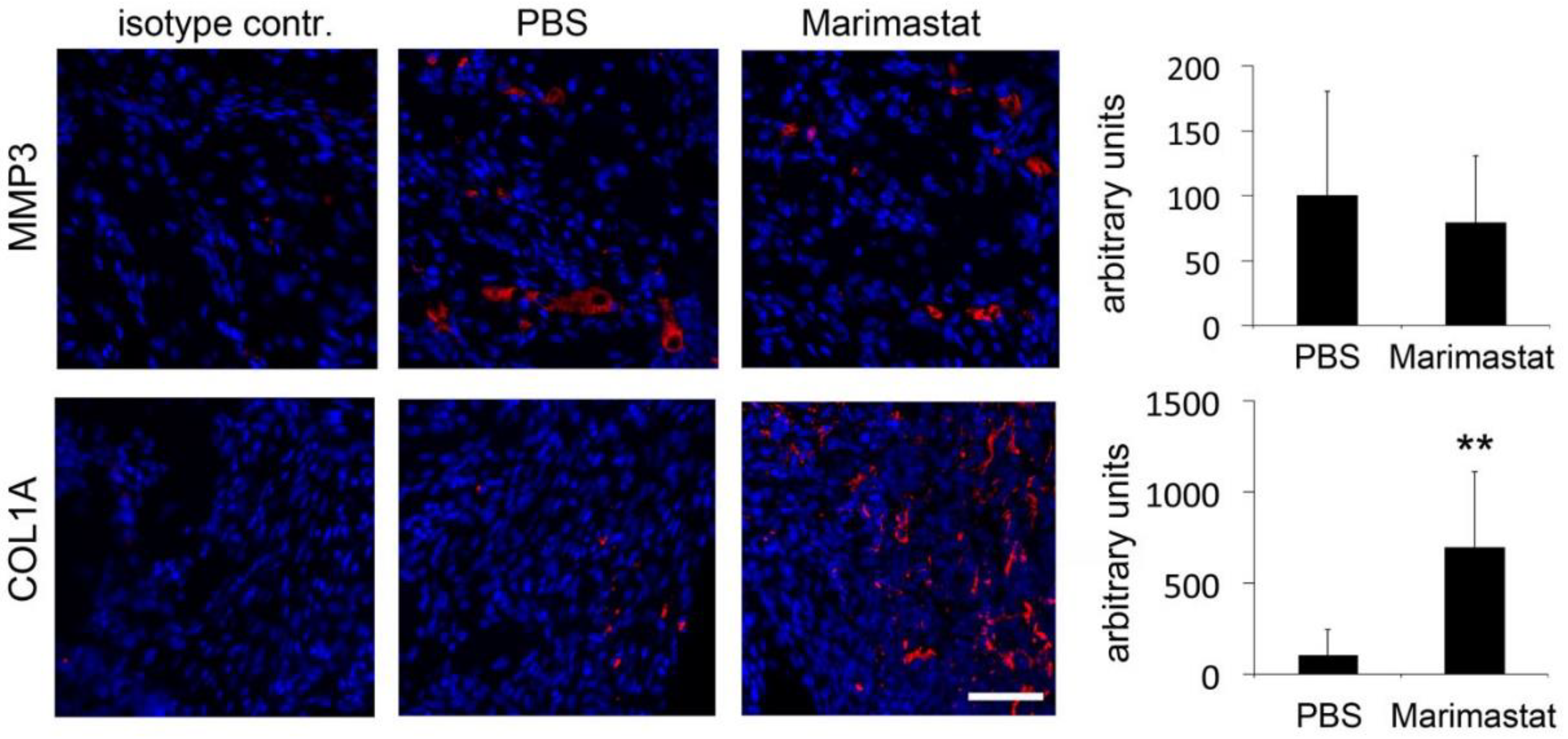
3.3. Enhanced Collagen Content by Inhibition of MMP Activity in Bones of db−/db− Mice
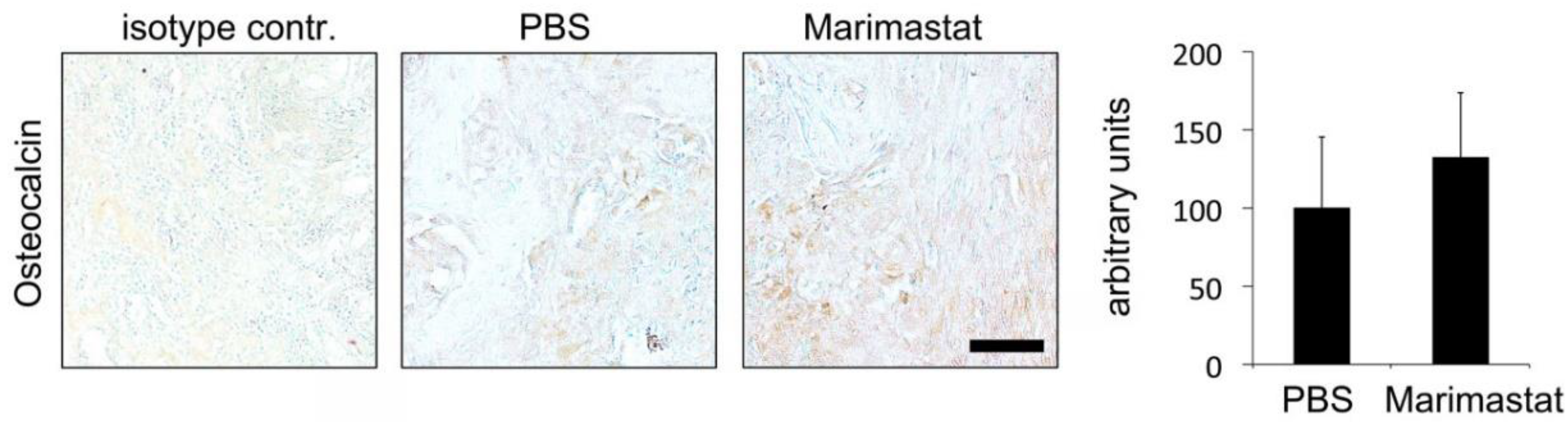
3.4. No Observable Effect of Marimastat on Osteocalcin Level Seven Days Postoperatively
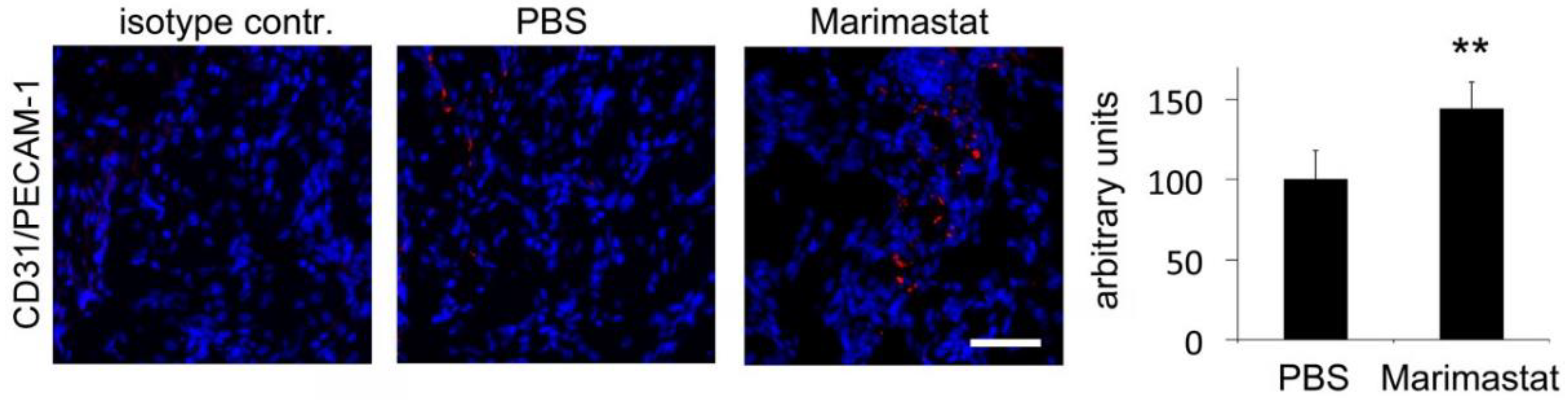
3.5. Enhanced Angiogenesis by Inhibition of MMP Activity in Bones of db−/db− Mice
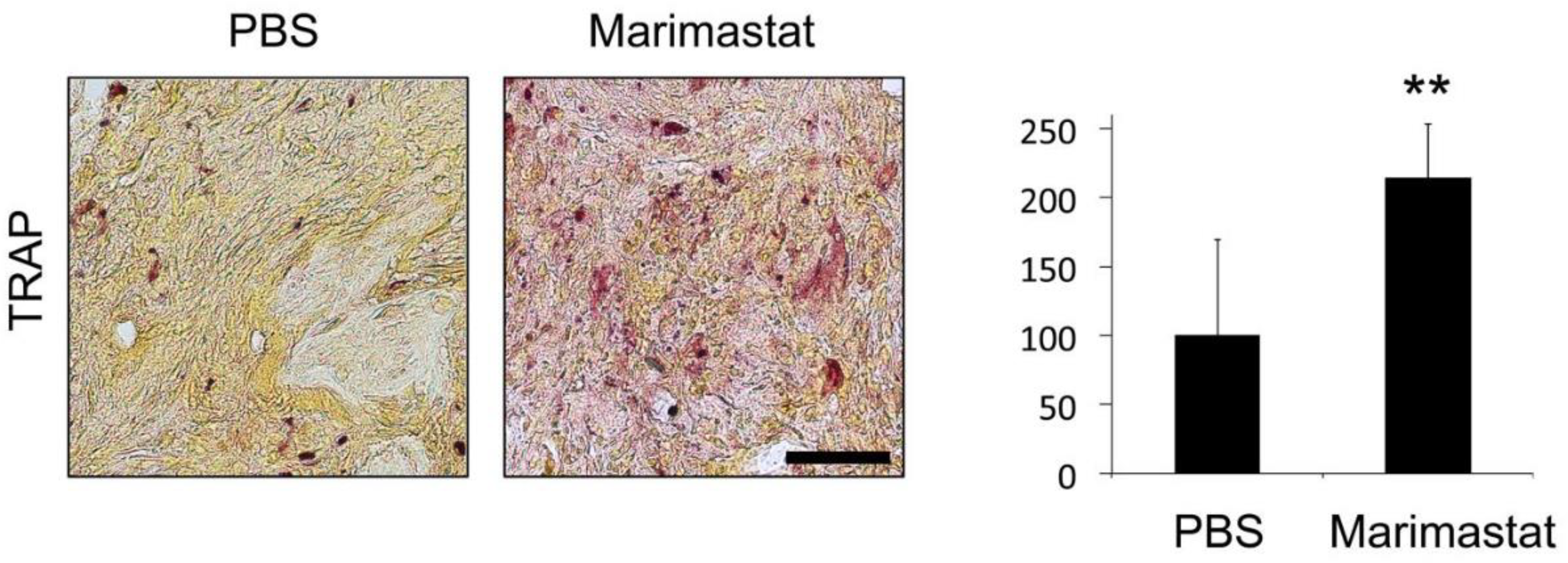
3.6. Increased Osteoclast Invasion by Inhibition of MMP Activity in Bones of db−/db− Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaston, M.S.; Simpson, A.H.R.W. Inhibition of fracture healing. J. Bone Jt. Surg. Br. Vol. 2007, 89, 1553–1560. [Google Scholar] [CrossRef]
- Hernandez, R.K.; Do, T.P.; Critchlow, C.W.; Dent, R.E.; Jick, S.S. Patient-related risk factors for fracture-healing complications in the United Kingdom general practice research database. Acta Orthop. 2012, 83, 653–660. [Google Scholar] [CrossRef]
- Retzepi, M.; Donos, N. The effect of diabetes mellitus on osseous healing. Clin. Oral Implant. Res. 2010, 21, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Khazai, N.B.; Beck, G.R., Jr.; Umpierrez, G.E. Diabetes and fractures: An overshadowed association. Curr. Opin. Endocrinol. Diabetes Obes. 2009, 16, 435–445. [Google Scholar] [CrossRef]
- Isidro, M.L.; Ruano, B. Bone disease in diabetes. Curr. Diabetes Rev. 2010, 6, 144–155. [Google Scholar] [CrossRef]
- Janghorbani, M.; van Dam, R.; Willett, W.C.; Hu, F.B. Systematic review of type 1 and type 2 diabetes mellitus and risk of fracture. Am. J. Epidemiol. 2007, 166, 495–505. [Google Scholar] [CrossRef]
- Heidemann, C.; Du, Y.; Schubert, I.; Rathmann, W.; Scheidt-Nave, C. Prävalenz und zeitliche Entwicklung des bekannten Diabetes mellitus: Ergebnisse der Studie zur Gesundheit Erwachsener in Deutschland (DEGS1). Bundesgesundheitsblatt Gesundheitsforsch. Gesundh. 2013, 56, 668–677. [Google Scholar] [CrossRef]
- Kahn, S.E.; Cooper, M.E.; Del Prato, S. Pathophysiology and treatment of type 2 diabetes: Perspectives on the past, present, and future. Lancet 2014, 383, 1068–1083. [Google Scholar] [CrossRef]
- Ritchie, N.D.; Baucom, K.J.; Sauder, K.A. Current perspectives on the impact of the national diabetes prevention program: Building on successes and overcoming challenges. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 2949–2957. [Google Scholar] [CrossRef]
- Wallner, C.; Schira-Heinen, J.; Wagner, J.M.; Schulte, M.; Fischer, S.; Hirsch, T.; Richter, W.; Abraham, S.; Kneser, U.; Lehnhardt, M.; et al. Application of VEGFA and FGF-9 enhances angiogenesis, osteogenesis and bone remodeling in type 2 diabetic long bone regeneration. PLoS ONE 2015, 10, e0118823. [Google Scholar] [CrossRef] [PubMed]
- Behr, B.; Leucht, P.; Longaker, M.T.; Quarto, N. Fgf-9 is required for angiogenesis and osteogenesis in long bone repair. Proc. Natl. Acad. Sci. USA 2010, 107, 11853–11858. [Google Scholar] [CrossRef]
- Becerikli, M.; Reinkemeier, F.; Dadras, M.; Wallner, C.; Wagner, J.M.; Drysch, M.; Sogorski, A.; von Glinski, M.; Lehnhardt, M.; Hahn, S.A.; et al. Tgf-beta pathway inhibition as the therapeutic acceleration of diabetic bone regeneration. J. Orthop. Res. 2021. [Google Scholar] [CrossRef]
- Crockett, J.C.; Rogers, M.J.; Coxon, F.P.; Hocking, L.J.; Helfrich, M.H. Bone remodelling at a glance. J. Cell Sci. 2011, 124, 991–998. [Google Scholar] [CrossRef]
- Hardy, E.; Fernandez-Patron, C. Destroy to rebuild: The connection between bone tissue remodeling and matrix metalloproteinases. Front. Physiol. 2020, 11. [Google Scholar] [CrossRef]
- Baragi, V.M.; Becher, G.; Bendele, A.M.; Biesinger, R.; Bluhm, H.; Boer, J.; Deng, H.; Dodd, R.; Essers, M.; Feuerstein, T.; et al. A new class of potent matrix metalloproteinase 13 inhibitors for potential treatment of osteoarthritis: Evidence of histologic and clinical efficacy without musculoskeletal toxicity in rat models. Arthritis Rheum. 2009, 60, 2008–2018. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, G.; Aoki, T.; Nakamura, H.; Tanzawa, K.; Okada, Y. Inhibition of ADAMTS4 (aggrecanase-1) by tissue inhibitors of metalloproteinases (TIMP-1, 2, 3 and 4). FEBS Lett. 2001, 494, 192–195. [Google Scholar] [CrossRef]
- Liang, H.P.H.; Xu, J.; Xue, M.; Jackson, C.J. Matrix metalloproteinases in bone development and pathology: Current knowledge and potential clinical utility. Met. Med. 2016, 3, 93–102. [Google Scholar] [CrossRef]
- Sobue, T.; Hakeda, Y.; Kobayashi, Y.; Hayakawa, H.; Yamashita, K.; Aoki, T.; Kumegawa, M.; Noguchi, T.; Hayakawa, T. Tissue Inhibitor of Metalloproteinases 1 and 2 directly stimulate the bone-resorbing activity of isolated mature osteoclasts. J. Bone Miner. Res. 2001, 16, 2205–2214. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Eckhardt, S.G. Development of matrix metalloproteinase inhibitors in cancer therapy. J. Natl. Cancer Inst. 2001, 93, 178–193. [Google Scholar] [CrossRef]
- Millar, A.W.; Brown, P.D.; Moore, J.; Galloway, W.A.; Cornish, A.G.; Lenehan, T.J.; Lynch, K.P. Results of single and repeat dose studies of the oral matrix metalloproteinase inhibitor marimastat in healthy male volunteers. Br. J. Clin. Pharmacol. 1998, 45, 21–26. [Google Scholar] [CrossRef]
- Thomas, A.L.; Steward, W.P. Marimastat: The clinical development of a matrix metalloproteinase inhibitor. Expert Opin. Investig. Drugs 2000, 9, 2913–2922. [Google Scholar] [CrossRef]
- Wallner, C.; Abraham, S.; Wagner, J.M.; Harati, K.; Ismer, B.; Kessler, L.; Zöllner, H.; Lehnhardt, M.; Behr, B. Local application of isogenic adipose-derived stem cells restores bone healing capacity in a type 2 diabetes model. Stem Cells Transl. Med. 2016, 5, 836–844. [Google Scholar] [CrossRef]
- Vannucci, L.; Fossi, C.; Quattrini, S.; Guasti, L.; Pampaloni, B.; Gronchi, G.; Giusti, F.; Romagnoli, C.; Cianferotti, L.; Marcucci, G.; et al. Calcium intake in bone health: A focus on calcium-rich mineral waters. Nutrients 2018, 10, 1930. [Google Scholar] [CrossRef]
- Wagner, J.M.; Reinkemeier, F.; Dadras, M.; Wallner, C.; Huber, J.; Sogorski, A.; Sacher, M.; Schmidt, S.; Drysch, M.; Dittfeld, S.; et al. Local Wnt3a treatment restores bone regeneration in large osseous defects after surgical debridement of osteomyelitis. J. Mol. Med. 2020, 98, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.M.; Reinkemeier, F.; Wallner, C.; Dadras, M.; Huber, J.; Schmidt, S.V.; Drysch, M.; Dittfeld, S.; Jaurich, H.; Becerikli, M.; et al. Adipose-derived stromal cells are capable of restoring bone regeneration after post-traumatic osteomyelitis and modulate B-cell response. Stem Cells Transl. Med. 2019, 8, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Becerikli, M.; Jaurich, H.; Schira-Heinen, J.; Schulte, M.; Döbele, C.; Wallner, C.; Abraham, S.; Wagner, J.M.; Dadras, M.; Kneser, U.; et al. Age-dependent alterations in osteoblast and osteoclast activity in human cancellous bone. J. Cell. Mol. Med. 2017, 21, 2773–2781. [Google Scholar] [CrossRef] [PubMed]
- Cignachi, N.P.; Pesquero, J.B.; Oliveira, R.B.; Etges, A.; Campos, M.M. Kinin B1 receptor deletion affects bone healing in type 1 diabetic mice. J. Cell. Physiol. 2015, 230, 3019–3028. [Google Scholar] [CrossRef]
- Ogasawara, A.; Nakajima, A.; Nakajima, F.; Goto, K.-I.; Yamazaki, M. Molecular basis for affected cartilage formation and bone union in fracture healing of the streptozotocin-induced diabetic rat. Bone 2008, 43, 832–839. [Google Scholar] [CrossRef]
- Henriksen, K.; Karsdal, M.A. Type I collagen. In Biochemistry of Collagens, Laminins and Elastin; Elsevier: Amsterdam, The Netherlands, 2019; pp. 1–12. [Google Scholar]
- Paiva, K.B.; Granjeiro, J.M. Matrix Metalloproteinases in Bone Resorption, Remodeling, and Repair. Prog. Mol. Biol. Transl. Sci. 2017, 148, 203–303. [Google Scholar] [CrossRef] [PubMed]
- Tokito, A.; Jougasaki, M. Matrix metalloproteinases in non-neoplastic disorders. Int. J. Mol. Sci. 2016, 17, 1178. [Google Scholar] [CrossRef]
- Pap, T.; Korb-Pap, A. Cartilage damage in osteoarthritis and rheumatoid arthritis—Two unequal siblings. Nat. Rev. Rheumatol. 2015, 11, 606–615. [Google Scholar] [CrossRef]
- Klimiuk, P.; Sierakowski, S.; Latosiewicz, R.; Cylwik, B.; Skowronski, J.; Chwiecko, J. Serum matrix metalloproteinases and tissue inhibitors of metalloproteinases in different histological variants of rheumatoid synovitis. Rheumatology 2002, 41, 78–87. [Google Scholar] [CrossRef][Green Version]
- Vu, T.H.; Werb, Z. Matrix metalloproteinases: Effectors of development and normal physiology. Genes Dev. 2000, 14, 2123–2133. [Google Scholar] [CrossRef] [PubMed]
- Haeusler, G.; Walter, I.; Helmreich, M.; Egerbacher, M. Localization of matrix metalloproteinases, (MMPs) their tissue inhibitors, and vascular endothelial growth factor (VEGF) in growth plates of children and adolescents indicates a role for MMPs in human postnatal growth and skeletal maturation. Calcif. Tissue Res. 2005, 76, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Kim, H.; Liu, X.; Sugiura, H.; Kohyama, T.; Fang, Q.; Wen, F.-Q.; Abe, S.; Wang, X.; Atkinson, J.J.; et al. Matrix metalloproteinase-9 activates TGF-β and stimulates fibroblast contraction of collagen gels. Am. J. Physiol. Cell. Mol. Physiol. 2014, 306, L1006–L1015. [Google Scholar] [CrossRef] [PubMed]
- Ren, P.; Hughes, M.; Krishnamoorthy, S.; Zou, S.; Zhang, L.; Wu, D.; Zhang, C.; Curci, J.A.; Coselli, J.S.; Milewicz, D.M.; et al. Critical role of ADAMTS-4 in the development of sporadic aortic aneurysm and dissection in mice. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Djouad, F.; Delorme, B.; Maurice, M.; Bony, C.; Apparailly, F.; Louis-Plence, P.; Canovas, F.; Charbord, P.; Noël, D.; Jorgensen, C. Microenvironmental changes during differentiation of mesenchymal stem cells towards chondrocytes. Arthritis Res. Ther. 2007, 9, R33. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Gao, W.; Zhu, L.; Ren, J.; Yao, H.; Wang, K.; Shi, D. TIMP-1 inhibits proliferation and osteogenic differentiation of hBMSCs through Wnt/β-catenin signaling. Biosci. Rep. 2019, 39, BSR20181290. [Google Scholar] [CrossRef] [PubMed]
- Piva, S.R.; Susko, A.M.; Khoja, S.S.; Josbeno, D.A.; Fitzgerald, G.K.; Toledo, F.G.S. Links between osteoarthritis and diabetes. Clin. Geriatr. Med. 2015, 31, 67–87. [Google Scholar] [CrossRef]
- Chen, T.-H.; Chen, L.; Hsieh, M.-S.; Chang, C.-P.; Chou, D.-T.; Tsai, S.-H. Evidence for a protective role for adiponectin in osteoarthritis. Biochim. Biophys. Acta Mol. Basis Dis. 2006, 1762, 711–718. [Google Scholar] [CrossRef]
- Zhu, W.-H.; Guo, X.; Villaschi, S.; Nicosia, R.F. Regulation of vascular growth and regression by matrix metalloproteinases in the rat aorta model of angiogenesis. Lab. Investig. 2000, 80, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Tamura, Y.; Kawao, N.; Okada, K.; Yano, M.; Okumoto, K.; Kaji, H. Influence of diabetic state and vitamin D deficiency on bone repair in female mice. Bone 2014, 61, 102–108. [Google Scholar] [CrossRef]
- Kayal, R.A.; Tsatsas, D.; Bauer, M.A.; Allen, B.; Al-Sebaei, M.O.; Kakar, S.; Leone, C.W.; Morgan, E.F.; Gerstenfeld, L.C.; Einhorn, T.A.; et al. Diminished bone formation during diabetic fracture healing is related to the premature resorption of cartilage associated with increased osteoclast activity. J. Bone Miner. Res. 2007, 22, 560–568. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, J.M.; Reinkemeier, F.; Wallner, C.; Dadras, M.; Dittfeld, S.; Drysch, M.; Sogorski, A.; von Glinski, M.; Lehnhardt, M.; Behr, B.; et al. Inhibition of Pathological Increased Matrix Metalloproteinase (MMP) Activity for Improvement of Bone Regeneration in Diabetes. Life 2022, 12, 134. https://doi.org/10.3390/life12020134
Wagner JM, Reinkemeier F, Wallner C, Dadras M, Dittfeld S, Drysch M, Sogorski A, von Glinski M, Lehnhardt M, Behr B, et al. Inhibition of Pathological Increased Matrix Metalloproteinase (MMP) Activity for Improvement of Bone Regeneration in Diabetes. Life. 2022; 12(2):134. https://doi.org/10.3390/life12020134
Chicago/Turabian StyleWagner, Johannes Maximilian, Felix Reinkemeier, Christoph Wallner, Mehran Dadras, Stephanie Dittfeld, Marius Drysch, Alexander Sogorski, Maxi von Glinski, Marcus Lehnhardt, Björn Behr, and et al. 2022. "Inhibition of Pathological Increased Matrix Metalloproteinase (MMP) Activity for Improvement of Bone Regeneration in Diabetes" Life 12, no. 2: 134. https://doi.org/10.3390/life12020134
APA StyleWagner, J. M., Reinkemeier, F., Wallner, C., Dadras, M., Dittfeld, S., Drysch, M., Sogorski, A., von Glinski, M., Lehnhardt, M., Behr, B., & Becerikli, M. (2022). Inhibition of Pathological Increased Matrix Metalloproteinase (MMP) Activity for Improvement of Bone Regeneration in Diabetes. Life, 12(2), 134. https://doi.org/10.3390/life12020134