
The Microalga Chlorella vulgaris as a Natural Bioenergetic System for Effective CO2 Mitigation—New Perspectives against Global Warming


Abstract
1. Introduction
2. Materials and Methods
2.1. Organism and Cultivation Conditions
2.2. Microalgae Growth Determination
2.3. GC-TCD Measurements
2.4. Photosynthetic and Respiratory Activity Measurements
2.5. Fluorescence Induction Measurements: OJIP-Test
2.6. Lipid Extraction and Quantification
2.7. Carbohydrate Extraction and Quantification
2.8. Data Analysis
3. Results
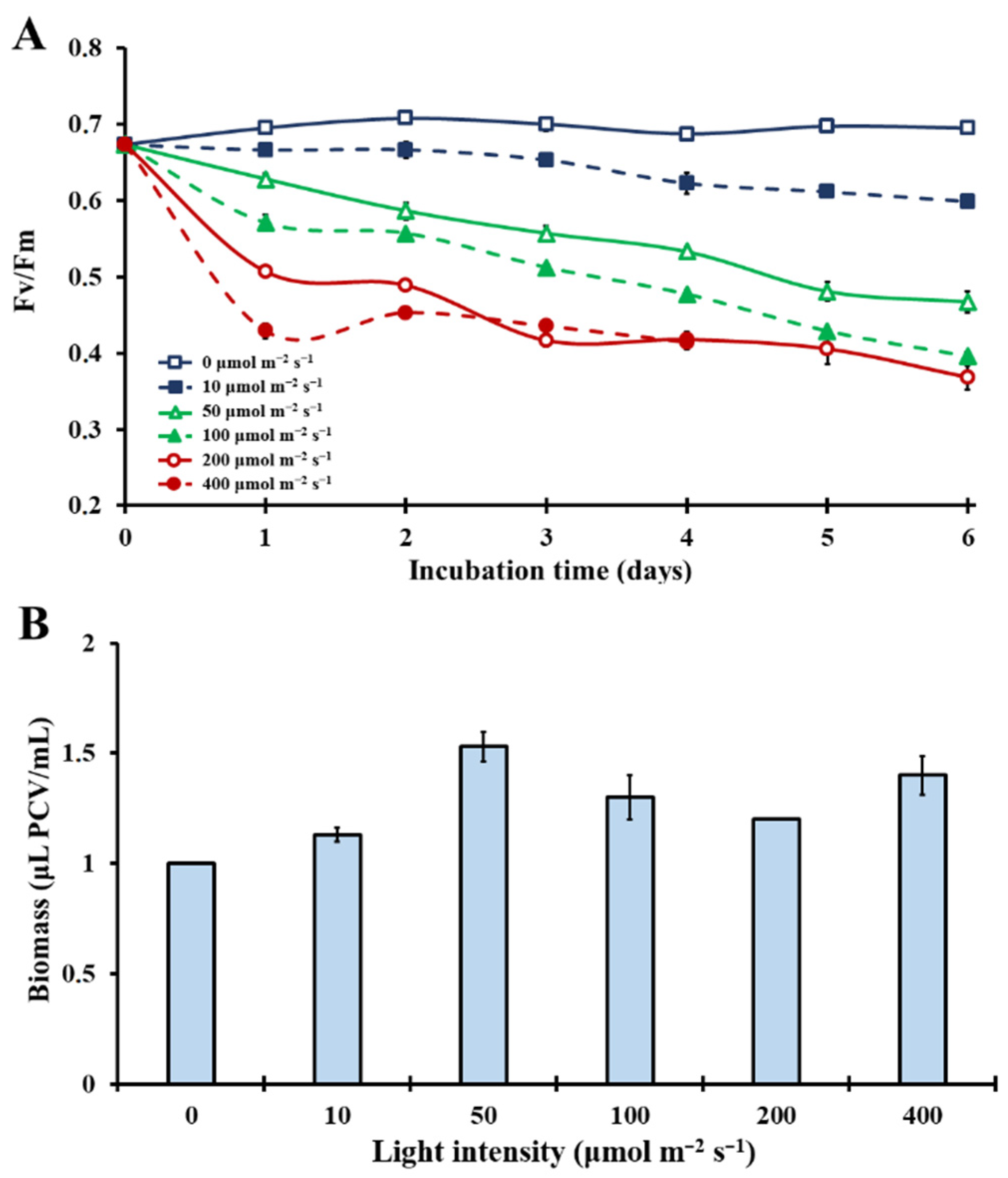
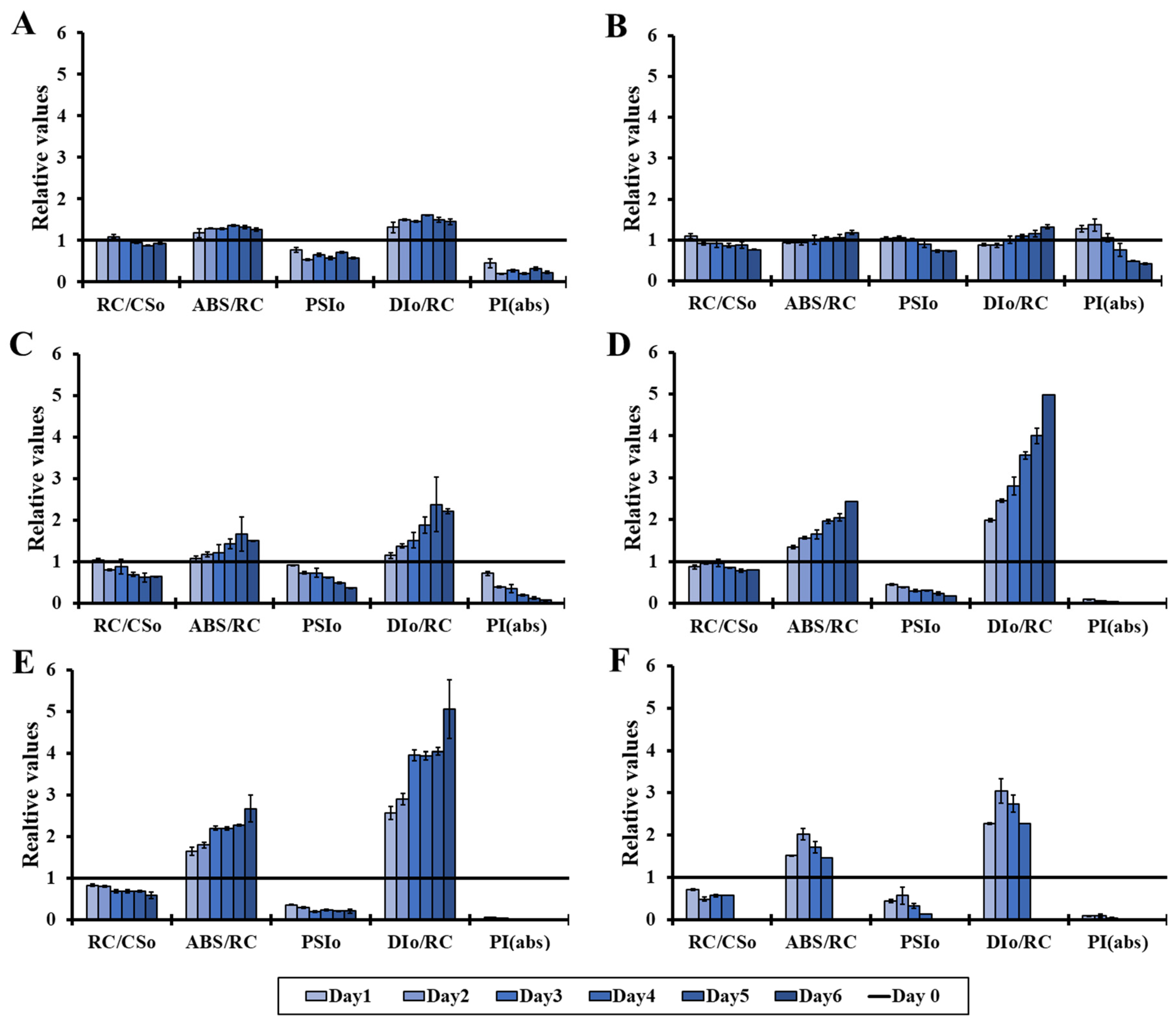
3.1. Effect of Different Light Intensities on the Microalgal Photosynthetic Mechanism
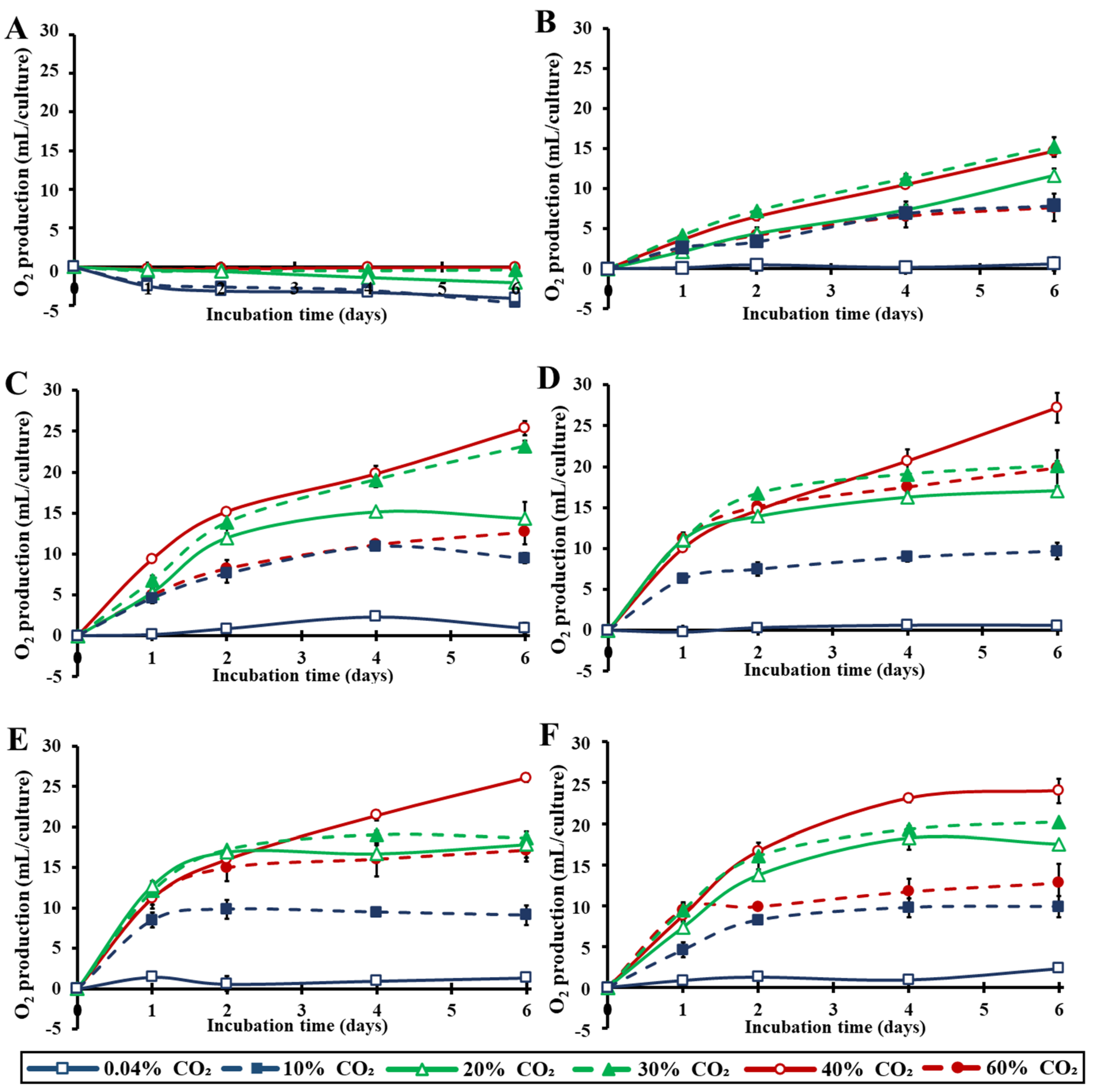
3.2. Effect of Various Extreme CO2 Concentrations at the Photosynthetic Mechanism under Several Light Intensities
3.3. Photosynthetic and Respiratory Activities under Different Light Intensities and CO2 Concentrations
3.4. Microalgal Biomass Production under Different Light Intensities and CO2 Concentrations
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peter, S.C. Reduction of CO2 to Chemicals and Fuels: A Solution to Global Warming and Energy Crisis. ACS Energy Lett. 2018, 3, 1557–1561. [Google Scholar] [CrossRef]
- McNeil, B.I.; Matear, R.J. Southern Ocean acidification: A tipping point at 450-ppm atmospheric CO2. Proc. Natl. Acad. Sci. USA 2008, 105, 18860–18864. [Google Scholar] [CrossRef]
- Etheridge, D.M.; Steele, L.P.; Langenfelds, R.L.; Francey, R.J.; Barnola, J.-M.; Morgan, V.I. Natural and anthropogenic changes in atmospheric CO2 over the last 1000 years from air in Antarctic ice and firn. J. Geophys. Res. Atmos. 1996, 101, 4115–4128. [Google Scholar] [CrossRef]
- Earth System Research Laboratory National Oceanic and Atmospheric Administration. Available online: https://www.esrl.noaa.gov/gmd/ccgg/trends/ (accessed on 4 May 2021).
- Lüthi, D.; Le Floch, M.; Bereiter, B.; Blunier, T.; Barnola, J.-M.; Siegenthaler, U.; Raynaud, D.; Jouzel, J.; Fischer, H.; Kawamura, K.; et al. High-resolution carbon dioxide concentration record 650,000–800,000 years before present. Nature 2008, 453, 379–382. [Google Scholar] [CrossRef]
- Ritchie, H.; Roser, M. CO2 and Greenhouse Gas Emissions. 2020. Available online: https://ourworldindata.org/co2-and-other-greenhouse-gas-emissions (accessed on 1 August 2020).
- Szulejko, J.E.; Kumar, P.; Deep, A.; Kim, K.-H. Global warming projections to 2100 using simple CO2 greenhouse gas modeling and comments on CO2 climate sensitivity factor. Atmos. Pollut. Res. 2017, 8, 136–140. [Google Scholar] [CrossRef]
- Nazarenko, L.; Schmidt, G.A.; Miller, R.L.; Tausnev, N.; Kelley, M.; Ruedy, R.; Russell, G.L.; Aleinov, I.; Bauer, S.; Bleck, R.; et al. Future climate change under RCP emission scenarios with GISS ModelE2. J. Adv. Model. Earth Syst. 2015, 7, 244–267. [Google Scholar] [CrossRef]
- Riahi, K.; van Vuuren, D.P.; Kriegler, E.; Edmonds, J.; O’Neill, B.C.; Fujimori, S.; Bauer, N.; Calvin, K.; Dellink, R.; Fricko, O.; et al. The Shared Socioeconomic Pathways and their energy, land use, and greenhouse gas emissions implications: An overview. Glob. Environ. Chang. 2017, 42, 153–168. [Google Scholar] [CrossRef]
- Walsh, B.; Ciais, P.; Janssens, I.A.; Peñuelas, J.; Riahi, K.; Rydzak, F.; van Vuuren, D.P.; Obersteiner, M. Pathways for balancing CO2 emissions and sinks. Nat. Commun. 2017, 8, 14856. [Google Scholar] [CrossRef] [PubMed]
- Rosa, G.M.; Moraes, L.; Cardias, B.B.; Souza, M.; Costa, J. Chemical absorption and CO2 biofixation via the cultivation of Spirulina in semicontinuous mode with nutrient recycle. Bioresour. Technol. 2015, 192, 321–327. [Google Scholar] [CrossRef] [PubMed]
- El Mekawy, A.; Hegab, H.M.; Mohanakrishna, G.; Elbaz, A.F.; Bulut, M.; Pant, D. Technological advances in CO2 conversion electro-biorefinery: A step toward commercialization. Bioresour. Technol. 2016, 215, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Duarte, J.H.; Fanka, L.S.; Costa, J.A.V. Utilization of simulated flue gas containing CO2, SO2, NO and ash for Chlorella fusca cultivation. Bioresour. Technol. 2016, 214, 159–165. [Google Scholar] [CrossRef]
- Wang, B.; Li, Y.; Wu, N.; Lan, C.Q. CO2 bio-mitigation using microalgae. Appl. Microbiol. Biotechnol. 2008, 79, 707–718. [Google Scholar] [CrossRef]
- Singh, J.; Dhar, D.W. Overview of Carbon Capture Technology: Microalgal Biorefinery Concept and State-of-the-Art. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef]
- Johnson, M.P. Photosynthesis. Essays Biochem. 2016, 60, 255–273. [Google Scholar] [CrossRef]
- Buick, R. When did oxygenic photosynthesis evolve? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 2731–2743. [Google Scholar] [CrossRef]
- Lyons, T.W.; Reinhard, C.T.; Planavsky, N.J. The rise of oxygen in Earth’s early ocean and atmosphere. Nature 2014, 506, 307–315. [Google Scholar] [CrossRef]
- Keller, D.P.; Lenton, A.; Littleton, E.W.; Oschlies, A.; Scott, V.; Vaughan, N.E. The Effects of Carbon Dioxide Removal on the Carbon Cycle. Curr. Clim. Chang. Rep. 2018, 4, 250–265. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Yeh, K.-L.; Aisyah, R.; Lee, D.-J.; Chang, J.-S. Cultivation, photobioreactor design and harvesting of microalgae for biodiesel production: A critical review. Bioresour. Technol. 2011, 102, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, L.; Oliveira, A.C. Microalgae as a raw material for biofuels production. J. Ind. Microbiol. Biotechnol. 2008, 36, 269–274. [Google Scholar] [CrossRef]
- Papazi, A.; Korelidou, A.; Andronis, E.; Parasyri, A.; Stamatis, N.; Kotzabasis, K. Bioenergetic reprogramming plasticity under nitrogen depletion by the unicellular green alga Scenedesmus Obliq. Planta 2018, 247, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Papazi, A.; Makridis, P.; Divanach, P.; Kotzabasis, K. Bioenergetic changes in the microalgal photosynthetic apparatus by extremely high CO2 concentrations induce an intense biomass production. Physiol. Plant. 2008, 132, 338–349. [Google Scholar] [CrossRef]
- Zerveas, S.; Kydonakis, E.; Moutidis, P.; Maragkoudakis, A.; Kotzabasis, K. Microalgae strategy in anoxic atmospheres with various CO2 concentrations—Environmental and (astro)biotechnological perspectives. Environ. Exp. Bot. 2021, 187, 104474. [Google Scholar] [CrossRef]
- Anjos, M.; Fernandes, B.D.; Vicente, A.A.; Teixeira, J.A.; Dragone, G. Optimization of CO2 bio-mitigation by Chlorella vulgaris. Bioresour. Technol. 2013, 139, 149–154. [Google Scholar] [CrossRef]
- Lam, M.K.; Lee, K.T. Effect of carbon source towards the growth of Chlorella vulgaris for CO2 bio-mitigation and biodiesel production. Int. J. Greenh. Gas Control 2013, 14, 169–176. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, F.; Hao, L.; Shedayi, A.A.; Guo, L.; Ma, C.; Huang, B.; Xu, M. The optimal CO2 concentrations for the growth of three perennial grass species. BMC Plant Biol. 2018, 18, 27. [Google Scholar] [CrossRef]
- Safi, C.; Zebib, B.; Merah, O.; Pontalier, P.-Y.; Vaca-Garcia, C. Morphology, composition, production, processing and applications of Chlorella vulgaris: A review. Renew. Sustain. Energy Rev. 2014, 35, 265–278. [Google Scholar] [CrossRef]
- Ru, I.T.K.; Sung, Y.Y.; Jusoh, M.; Wahid, M.E.A.; Nagappan, T. Chlorella vulgaris: A perspective on its potential for combining high biomass with high value bioproducts. J. Appl. Phycol. 2020, 1, 2–11. [Google Scholar] [CrossRef]
- Görs, M.; Schumann, R.; Hepperle, D.; Karsten, U. Quality analysis of commercial Chlorella products used as dietary supplement in human nutrition. J. Appl. Phycol. 2009, 22, 265–276. [Google Scholar] [CrossRef]
- Kholif, A.E.; Morsy, T.A.; Matloup, O.H.; Anele, U.Y.; Mohamed, A.G.; El-Sayed, A.B. Dietary Chlorella vulgaris microalgae improves feed utilization, milk production and concentrations of conjugated linoleic acids in the milk of Damascus goats. J. Agric. Sci. 2016, 155, 508–518. [Google Scholar] [CrossRef]
- Ahmad, M.T.; Shariff, M.; Yusoff, F.; Goh, Y.M.; Banerjee, S. Applications of microalga Chlorella vulgaris in aquaculture. Rev. Aquac. 2018, 12, 328–346. [Google Scholar] [CrossRef]
- Schreiber, C.; Schiedung, H.; Harrison, L.; Briese, C.; Ackermann, B.; Kant, J.; Schrey, S.D.; Hofmann, D.; Singh, D.; Ebenhoh, O.; et al. Evaluating potential of green alga Chlorella vulgaris to accumulate phosphorus and to fertilize nutrient-poor soil substrates for crop plants. J. Appl. Phycol. 2018. [Google Scholar] [CrossRef]
- Faheed, F.A.; Abd-El Fattah, Z. Effect of Chlorella vulgaris as bio-fertilizer on growth parameters and metabolic aspects of lettuce plant. J. Agri. Soc. Sci. 2008, 4, 165–169. [Google Scholar]
- Singh, A.; Nigam, P.S.; Murphy, J.D. Renewable fuels from algae: An answer to debatable land based fuels. Bioresour. Technol. 2011, 102, 10–16. [Google Scholar] [CrossRef]
- Kim, K.H.; Choi, I.S.; Kim, H.M.; Wi, S.G.; Bae, H.-J. Bioethanol production from the nutrient stress-induced microalga Chlorella vulgaris by enzymatic hydrolysis and immobilized yeast fermentation. Bioresour. Technol. 2014, 153, 47–54. [Google Scholar] [CrossRef]
- Asadi, P.; Rad, H.A.; Qaderi, F. Lipid and biodiesel production by cultivation isolated strain Chlorella sorokiniana pa.91 and Chlorella vulgaris in dairy wastewater treatment plant effluents. J. Environ. Health Sci. Eng. 2020, 18, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Beyerinck, M.W. Culturversuche mit Zoochlorellen, Lichenengonidien und anderen niederen Algen. Bot. Ztg. 1890, 47, 725–739, 741–754, 757–768, 781–785. [Google Scholar]
- Wong, Y.K.; Ho, Y.H.; Ho, K.C.; Leung, H.M.; Yung, K.K.L. Growth Medium Screening for Chlorella vulgaris Growth and Lipid Production. J. Aquac. Mar. Biol. 2017, 6, 00143. [Google Scholar] [CrossRef]
- Nichols, H.W.; Bold, H.C. Trichosarcina polymorpha Gen. et Sp. Nov. J. Phycol. 1965, 1, 34–38. [Google Scholar] [CrossRef]
- Serra-Maia, R.; Bernard, O.; Gonçalves, A.; Bensalem, S.; Lopes, F. Influence of temperature on Chlorella vulgaris growth and mortality rates in a photobioreactor. Algal Res. 2016, 18, 352–359. [Google Scholar] [CrossRef]
- Seyfabadi, J.; Ramezanpour, Z.; Amini Khoeyi, Z. Protein, fatty acid, and pigment content of Chlorella vulgaris under different light regimes. J. Appl. Phycol. 2010, 23, 721–726. [Google Scholar] [CrossRef]
- Zerveas, S.; Mente, M.; Tsakiri, D.; Kotzabasis, K. Microalgal photosynthesis induces alkalization of aquatic environment as a result of H+ uptake independently from CO2 concentration—New perspectives for environmental applications. J. Environ. Manage. 2021, 289, 112546. [Google Scholar] [CrossRef]
- Navakoudis, E.; Vrentzou, K.; Kotzabasis, K. A polyamine- and LHCII protease activity-based mechanism regulates the plasticity and adaptation status of the photosynthetic apparatus. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 261–271. [Google Scholar] [CrossRef]
- Strasser, B.J.; Strasser, R.J. Measuring Fast Fluorescence Transients to Address Environmental Questions: The JIP-Test. In Photosynthesis: From Light to Biosphere; Mathis, P., Ed.; Kluwer Academic Press: Dordrecht, The Netherlands, 1995; pp. 977–980. [Google Scholar] [CrossRef]
- Srivastava, A.; Strasser, R.J.; Govindjee. Polyphasic rise of chlorophyll a fluorescence in herbicide-resistant D1 mutants of Chlamydomonas reinardtii. Photosynth. Res. 1995, 43, 131–141. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence. Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 19. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Sati, H.; Mitra, M.; Mishra, S.; Baredar, P. Microalgal lipid extraction strategies for biodiesel production: A review. Algal Res. 2019, 38, 101413. [Google Scholar] [CrossRef]
- Park, J.; Jeong, H. Easy and rapid quantification of lipid contents of marine dinoflagellates using the sulpho-phospho-vanillin method. Algae 2016, 31, 391–401. [Google Scholar] [CrossRef]
- Schulze, C.; Wetzel, M.; Reinhardt, J.; Schmidt, M.; Felten, L.; Mundt, S. Screening of microalgae for primary metabolites including β-glucans and the influence of nitrate starvation and irradiance on β-glucan production. J. Appl. Phycol. 2016, 28, 2719–2725. [Google Scholar] [CrossRef]
- Navakoudis, E.; Lütz, C.; Langebartels, C.; Lütz-Meindl, U.; Kotzabasis, K. Ozone impact on the photosynthetic apparatus and the protective role of polyamines. Biochim. Biophys. Acta. 2003, 1621, 160–169. [Google Scholar] [CrossRef]
- Lütz, C.; Navakoudis, E.; Seidlitz, H.K.; Kotzabasis, K. Simulated solar irradiation with enhanced UV-B adjust plastid- and thylakoid-associated polyamine changes for UV-B protection. Biochim. Biophys. Acta 2005, 1710, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Sfichi, L.; Ioanidis, E.N.; Kotzabasis, K. Thylakoid-associated polyamines adjust the UVB-sensitivity of the photosynthetic apparatus by means of LHCII changes. Photochem. Photobiol. 2004, 80, 499–506. [Google Scholar] [CrossRef]
- Sfakianaki, M.; Sfichi, L.; Kotzabasis, K. The involvement of LHCII-associated polyamines in the response of the photosynthetic apparatus to low temperature. J. Photochem. Photobiol. B 2006, 84, 181–188. [Google Scholar] [CrossRef]
- Jin, H.F.; Lim, B.R.; Lee, K. Influence of nitrate feeding on carbon dioxide fixation by microalgae. J. Environ. Sci. Health A 2006, 41, 2813–2824. [Google Scholar] [CrossRef] [PubMed]
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.-J.; Chang, J.-S. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Friedlingstein, P.; O’Sullivan, M.; Jones, M.W.; Andrew, R.M.; Hauck, J.; Olsen, A.; Peters, G.P.; Peters, W.; Pongratz, J.; Sitch, S.; et al. Global Carbon Budget. Earth Syst. Sci. Data 2020, 12, 3269–3340. [Google Scholar] [CrossRef]
- Carrasquillo, R. ISS ECLSS Technology Evolution for Exploration. In Proceedings of the 43rd AIAA Aerospace Sciences Meeting and Exhibit, Reno, NV, USA, 10–13 January 2005. [Google Scholar] [CrossRef]
- Yang, L.; Li, H.; Liu, T.; Zhong, Y.; Ji, C.; Lu, Q.; Fan, L.; Li, J.; Leng, L.; Li, K.; et al. Microalgae biotechnology as an attempt for bioregenerative life support systems: Problems and prospects. J. Chem. Technol. Biotechnol. 2019, 94, 3039–3048. [Google Scholar] [CrossRef]
- Battistuzzi, M.; Cocola, L.; Salasnich, B.; Erculiani, M.S.; Alei, E.; Morosinotto, T.; Claudi, R.; Poletto, L.; La Rocca, N. A new remote sensing-based system for the monitoring and analysis of growth and gas exchange rates of photosynthetic microorganisms under simulated non-terrestrial conditions. Front. Plant Sci. 2020, 11, 182. [Google Scholar] [CrossRef] [PubMed]
- Jones, H. Design Rules for Space Life Support Systems. SAE Tech. Pap. 2003, 2356. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| [CO2] | Light Intensity | Carbohydrates (% DW) ± SE% | Carbohydrates (mg/L) ± SE% | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Day 0 | Day 1 | Day 3 | Day 6 | Day 0 | Day 1 | Day 3 | Day 6 | ||
| 0.04% | 0 μmol m−2 s−1 | 24.0 ± 1.0 | 17.9 ± 0.5 | 15.2 ± 1.2 | 13.5 ± 0.7 | 42.2 ± 1.8 | 31.5 ± 0.9 | 26.7 ± 2.0 | 23.7 ± 1.1 |
| 10 μmol m−2 s−1 | 24.0 ± 1.0 | 17.9 ±1.0 | 8.6 ± 0.4 | 10.2 ± 0.2 | 42.2 ± 1.8 | 31.5 ± 1.8 | 23.1 ± 0.7 | 30.5 ± 0.2 | |
| 50 μmol m−2 s−1 | 24.0 ± 1.0 | 14.6 ± 0.9 | 11.9 ± 1.0 | 11.3 ± 1.1 | 42.2 ± 1.8 | 26.1 ± 1.5 | 28.8 ± 2.5 | 30.5 ± 3.0 | |
| 100 μmol m−2 s−1 | 24.0 ± 1.0 | 17.1 ± 1.2 | 12.5 ± 2.2 | 12.1 ± 1.0 | 42.2 ± 1.8 | 30.0 ± 2.2 | 26.4 ± 4.6 | 27.7 ± 2.2 | |
| 200 μmol m−2 s−1 | 24.0 ± 1.0 | 20.1 ± 2.2 | 18.5 ± 0.4 | 18.4 ± 1.8 | 42.2 ± 1.8 | 35.3 ± 3.9 | 35.9 ± 1.0 | 38.8 ± 3.7 | |
| 400 μmol m−2 s−1 | 24.0 ± 1.0 | 11.9 ± 0.4 | 8.6 ± 0.1 | 10.0 ± 0.1 | 42.2 ± 1.8 | 20.9 ± 0.8 | 16.8 ± 0.3 | 21.0 ± 0.4 | |
| 30% | 0 μmol m−2 s−1 | 24.0 ± 1.0 | 21.5 ± 2.9 | 21.4 ± 1.2 | 15.0 ± 0.5 | 42.2 ± 1.8 | 37.8 ± 5.1 | 37.6 ± 3.4 | 26.4 ± 1.0 |
| 10 μmol m−2 s−1 | 24.0 ± 1.0 | 17.1 ± 2.7 | 35.5 ± 0.7 | 23.4 ± 0.7 | 42.2 ± 1.8 | 70.8 ± 4.3 | 79.6 ± 1.7 | 73.1 ± 2.0 | |
| 50 μmol m−2 s−1 | 24.0 ± 1.0 | 37.2 ± 1.3 | 14.0 ± 1.6 | 18.7 ± 1.0 | 42.2 ± 1.8 | 51.8± 3.3 | 90.2 ± 6.9 | 116.2 ± 5.8 | |
| 100 μmol m−2 s−1 | 24.0 ± 1.0 | 31.0 ± 1.7 | 20.1 ± 2.0 | 15.5 ± 3.0 | 42.2 ± 1.8 | 79.0 ± 4.2 | 114.3 ± 8.5 | 104.1±20.3 | |
| 200 μmol m−2 s−1 | 24.0 ± 1.0 | 34.8 ± 1.1 | 19.4 ± 0.6 | 21.4 ± 0.5 | 42.2 ± 1.8 | 67.1 ± 2.7 | 83.4 ± 1.9 | 138.6 ± 3.2 | |
| 400 μmol m−2 s−1 | 24.0 ± 1.0 | 34.4 ± 2.3 | 28.7 ± 2.8 | 16.5 ± 0.7 | 42.2 ± 1.8 | 87.8 ± 5.7 | 151.7±14.6 | 112.9± 4.7 | |
| 60% | 0 μmol m−2 s−1 | 24.0 ± 1.0 | 19.5 ± 0.7 | 19.0 ± 0.5 | 15.6 ± 0.4 | 42.2 ± 1.8 | 34.3 ± 1.2 | 32.9 ± 1.0 | 27.4 ± 0.8 |
| 10 μmol m−2 s−1 | 24.0 ± 1.0 | 14.3 ± 0.4 | 14.8 ± 0.7 | 12.2 ± 0.8 | 42.2 ± 1.8 | 27.6 ± 0.9 | 56.4 ± 1.5 | 33.7 ± 2.3 | |
| 50 μmol m−2 s−1 | 24.0 ± 1.0 | 27.0 ± 0.4 | 21.5 ± 2.0 | 22.8 ± 2.6 | 42.2 ± 1.8 | 52.3 ± 0.7 | 60.6 ± 5.8 | 85.4 ± 9.7 | |
| 100 μmol m−2 s−1 | 24.0 ± 1.0 | 27.1 ± 2.0 | 20.7 ± 1.3 | 16.1 ± 1.6 | 42.2 ± 1.8 | 52.4 ± 4.0 | 68.1 ± 4.2 | 74.4 ± 7.4 | |
| 200 μmol m−2 s−1 | 24.0 ± 1.0 | 26.3 ± 1.0 | 22.8 ± 1.5 | 29.1 ± 1.2 | 42.2 ± 1.8 | 67.5 ± 1.8 | 72.2 ± 4.7 | 111.3 ± 4.8 | |
| 400 μmol m−2 s−1 | 24.0 ± 1.0 | 29.6 ± 1.8 | 27.3 ± 0.7 | 16.1 ± 0.4 | 42.2 ± 1.8 | 62.7 ± 3.7 | 73.5 ± 2.1 | 78.0 ± 1.2 | |
| [CO2] | Light Intensity | Lipids (% DW) ± SE% | Lipids (mg/L) ± SE% | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Day 0 | Day 1 | Day 3 | Day 6 | Day 0 | Day 1 | Day 3 | Day 6 | ||
| 0.04% | 0 μmol m−2 s−1 | 13.2 ± 0.6 | 8.90 ± 0.4 | 13.1 ± 0.2 | 11.1 ± 0.1 | 23.2 ± 0.8 | 15.6 ± 0.7 | 23.0 ± 0.4 | 19.5 ± 0.1 |
| 10 μmol m−2 s−1 | 13.2 ± 0.6 | 14.2 ± 0.2 | 9.60 ± 1.0 | 9.70 ± 0.2 | 23.2 ± 0.8 | 25.1 ± 0.3 | 16.9 ± 1.8 | 19.2 ± 0.4 | |
| 50 μmol m−2 s−1 | 13.2 ± 0.6 | 12.4 ± 0.5 | 10.3 ± 0.2 | 8.50 ± 0.3 | 23.2 ± 0.8 | 20.3 ± 1.2 | 21.8 ± 0.7 | 25.1 ± 1.3 | |
| 100 μmol m−2 s−1 | 13.2 ± 0.6 | 13.5 ± 0.6 | 13.5 ± 0.7 | 10.6 ± 0.7 | 23.2 ± 0.8 | 23.7 ± 1.0 | 28.5 ± 1.5 | 24.2 ± 1.6 | |
| 200 μmol m−2 s−1 | 13.2 ± 0.6 | 13.3 ± 0.6 | 11.0 ± 0.1 | 9.30 ± 0.1 | 23.2 ± 0.8 | 23.4 ± 1.0 | 21.3 ± 0.3 | 19.7 ± 0.1 | |
| 400 μmol m−2 s−1 | 13.2 ± 0.6 | 13.7 ± 1.0 | 10.5 ± 0.4 | 8.20 ± 0.7 | 23.2 ± 0.8 | 24.1 ± 1.7 | 20.4 ± 0.8 | 17.2 ± 1.5 | |
| 30% | 0 μmol m−2 s−1 | 13.2 ± 0.6 | 12.0 ± 0.4 | 10.7 ± 0.4 | 12.4 ± 0.3 | 23.2 ± 0.8 | 21.0 ± 0.6 | 18.8 ± 0.7 | 21.8 ± 0.6 |
| 10 μmol m−2 s−1 | 13.2 ± 0.6 | 14.2 ± 0.4 | 9.30 ± 0.2 | 10.3 ± 0.2 | 23.2 ± 0.8 | 24.5 ± 0.8 | 20.9 ± 0.3 | 32.0 ± 0.6 | |
| 50 μmol m−2 s−1 | 13.2 ± 0.6 | 11.7 ± 0.4 | 8.30 ± 0.1 | 7.60 ± 0.4 | 23.2 ± 0.8 | 28.8 ± 1.5 | 35.1 ± 0.2 | 49.1 ± 0.9 | |
| 100 μmol m−2 s−1 | 13.2 ± 0.6 | 10.4 ± 0.6 | 13.9 ± 0.8 | 8.60 ± 0.1 | 23.2 ± 0.8 | 26.6 ± 1.6 | 57.9 ± 3.2 | 58.2 ± 0.5 | |
| 200 μmol m−2 s−1 | 13.2 ± 0.6 | 10.6 ± 0.7 | 7.20 ± 0.3 | 8.00 ± 0.2 | 23.2 ± 0.8 | 27.0 ± 1.7 | 36.2 ± 1.8 | 51.8 ± 1.3 | |
| 400 μmol m−2 s−1 | 13.2 ± 0.6 | 13.6 ± 0.6 | 7.50 ± 0.3 | 6.80 ± 0.4 | 23.2 ± 0.8 | 34.8 ± 1.6 | 39.4 ± 1.8 | 46.2 ± 3.0 | |
| 60% | 0 μmol m−2 s−1 | 13.2 ± 0.6 | 11.4 ± 0.5 | 11.2 ± 1.4 | 13.9 ± 0.8 | 23.2 ± 0.8 | 20.1 ± 0.8 | 19.8 ± 2.4 | 24.5 ± 1.4 |
| 10 μmol m−2 s−1 | 13.2 ± 0.6 | 15.8 ± 0.1 | 12.3 ± 0.4 | 14.0 ± 0.1 | 23.2 ± 0.8 | 30.7 ± 0.2 | 31.0 ± 1.2 | 38.6 ± 0.2 | |
| 50 μmol m−2 s−1 | 13.2 ± 0.6 | 13.8 ± 0.2 | 11.2 ± 0.1 | 11.7 ± 0.4 | 23.2 ± 0.8 | 26.7 ± 0.4 | 31.5 ± 0.5 | 44.0 ± 2.2 | |
| 100 μmol m−2 s−1 | 13.2 ± 0.6 | 16.4 ±0.5 | 18.3 ± 0.4 | 8.60 ± 0.2 | 23.2 ± 0.8 | 31.8 ± 0.9 | 62.0 ± 1.3 | 39.8 ± 1.0 | |
| 200 μmol m−2 s−1 | 13.2 ± 0.6 | 19.0 ± 0.5 | 10.5 ± 0.3 | 8.80 ± 0.3 | 23.2 ± 0.8 | 24.6 ± 1.0 | 33.2 ± 0.9 | 33.7 ± 1.1 | |
| 400 μmol m−2 s−1 | 13.2 ± 0.6 | 13.4 ± 1.1 | 13.6 ± 0.4 | 8.10 ± 0.3 | 23.2 ± 0.8 | 28.4 ± 2.3 | 36.7 ± 1.0 | 24.2 ± 0.8 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mountourakis, F.; Papazi, A.; Kotzabasis, K. The Microalga Chlorella vulgaris as a Natural Bioenergetic System for Effective CO2 Mitigation—New Perspectives against Global Warming. Symmetry 2021, 13, 997. https://doi.org/10.3390/sym13060997
Mountourakis F, Papazi A, Kotzabasis K. The Microalga Chlorella vulgaris as a Natural Bioenergetic System for Effective CO2 Mitigation—New Perspectives against Global Warming. Symmetry. 2021; 13(6):997. https://doi.org/10.3390/sym13060997
Chicago/Turabian StyleMountourakis, Fanourios, Aikaterini Papazi, and Kiriakos Kotzabasis. 2021. "The Microalga Chlorella vulgaris as a Natural Bioenergetic System for Effective CO2 Mitigation—New Perspectives against Global Warming" Symmetry 13, no. 6: 997. https://doi.org/10.3390/sym13060997
APA StyleMountourakis, F., Papazi, A., & Kotzabasis, K. (2021). The Microalga Chlorella vulgaris as a Natural Bioenergetic System for Effective CO2 Mitigation—New Perspectives against Global Warming. Symmetry, 13(6), 997. https://doi.org/10.3390/sym13060997