

Changes in Ground Cover Layers, Biomass and Diversity of Vascular Plants/Mosses in the Clear-Cuts Followed by Reforested Scots Pine until Maturity Age


Abstract
1. Introduction
2. Materials and Methods
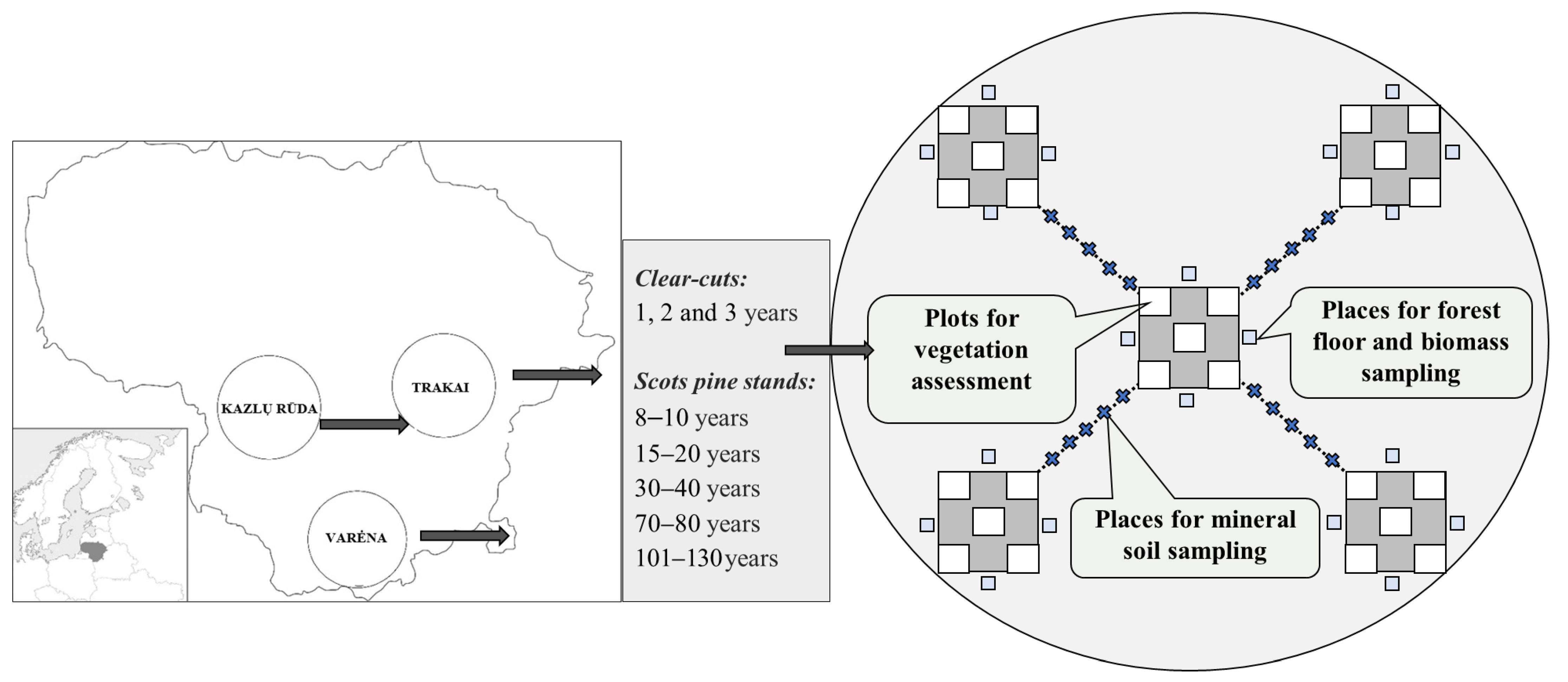
2.1. Study Design and Characteristics
2.2. Assessment of Ground Vegetation Cover
2.3. Assessment of Forest Floor, Ground Vegetation Mass, and Dead Wood Accumulation
2.4. Canopy Density Assessment
2.5. Calculations and Statistical Analysis
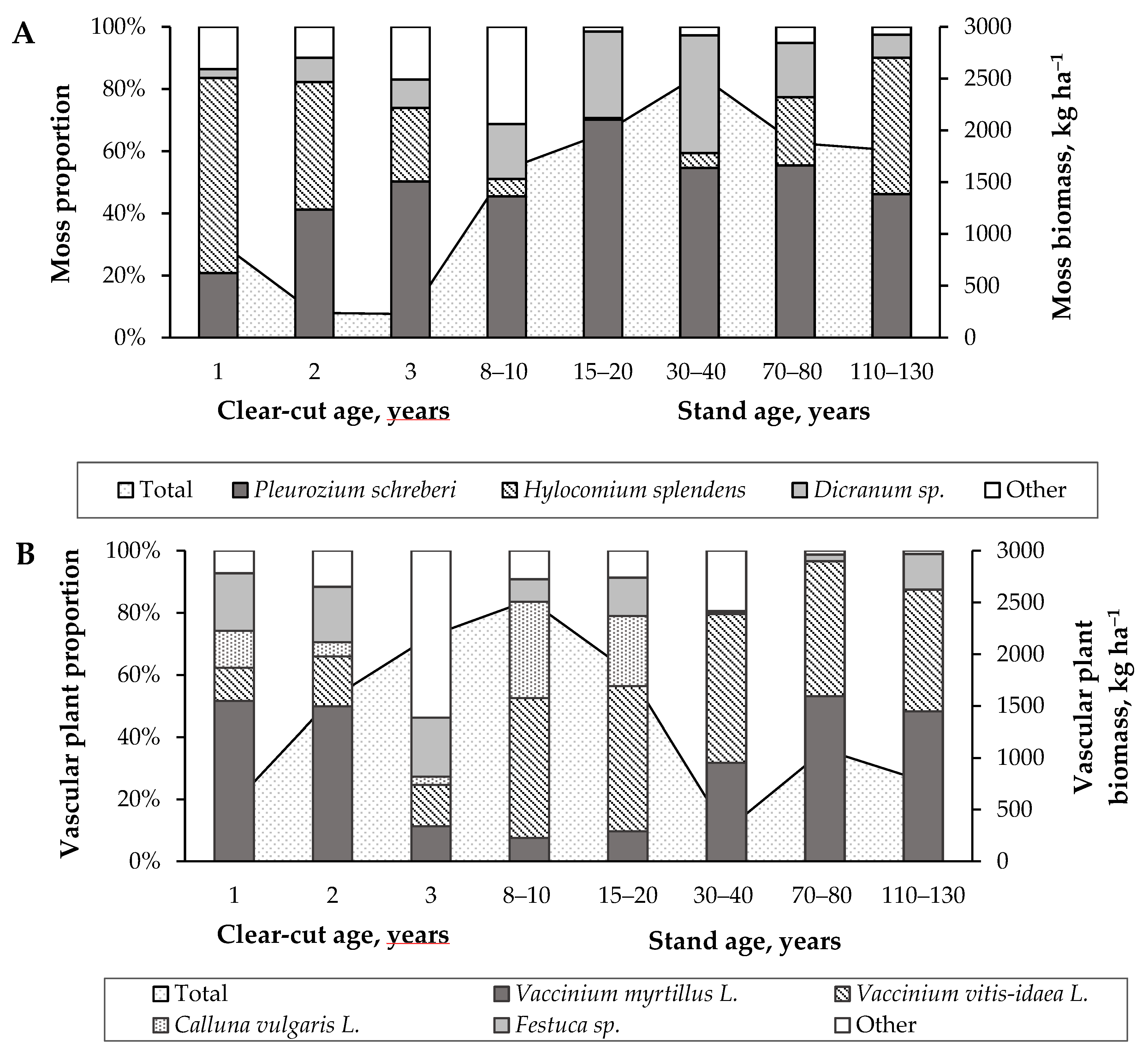
3. Results
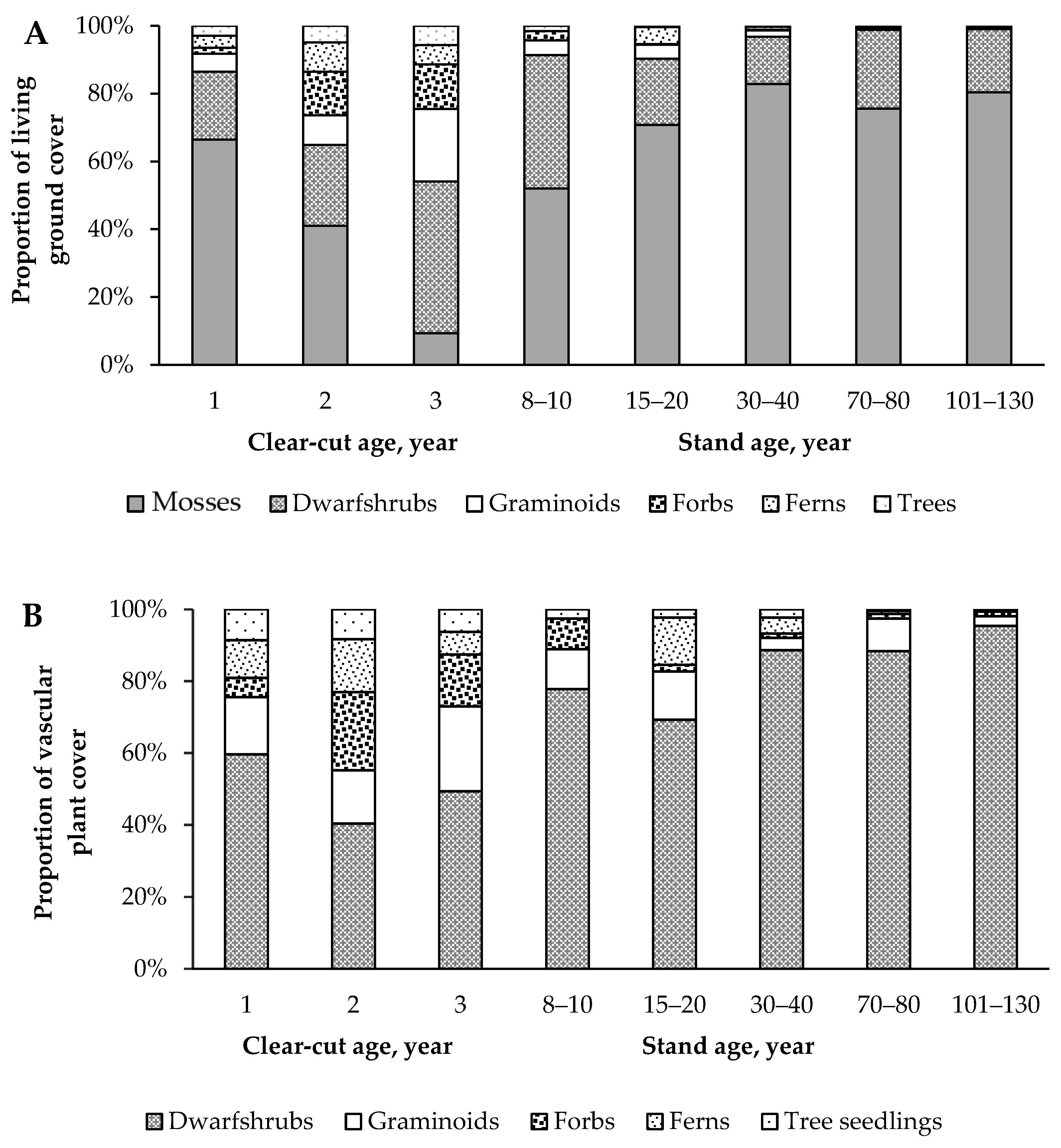
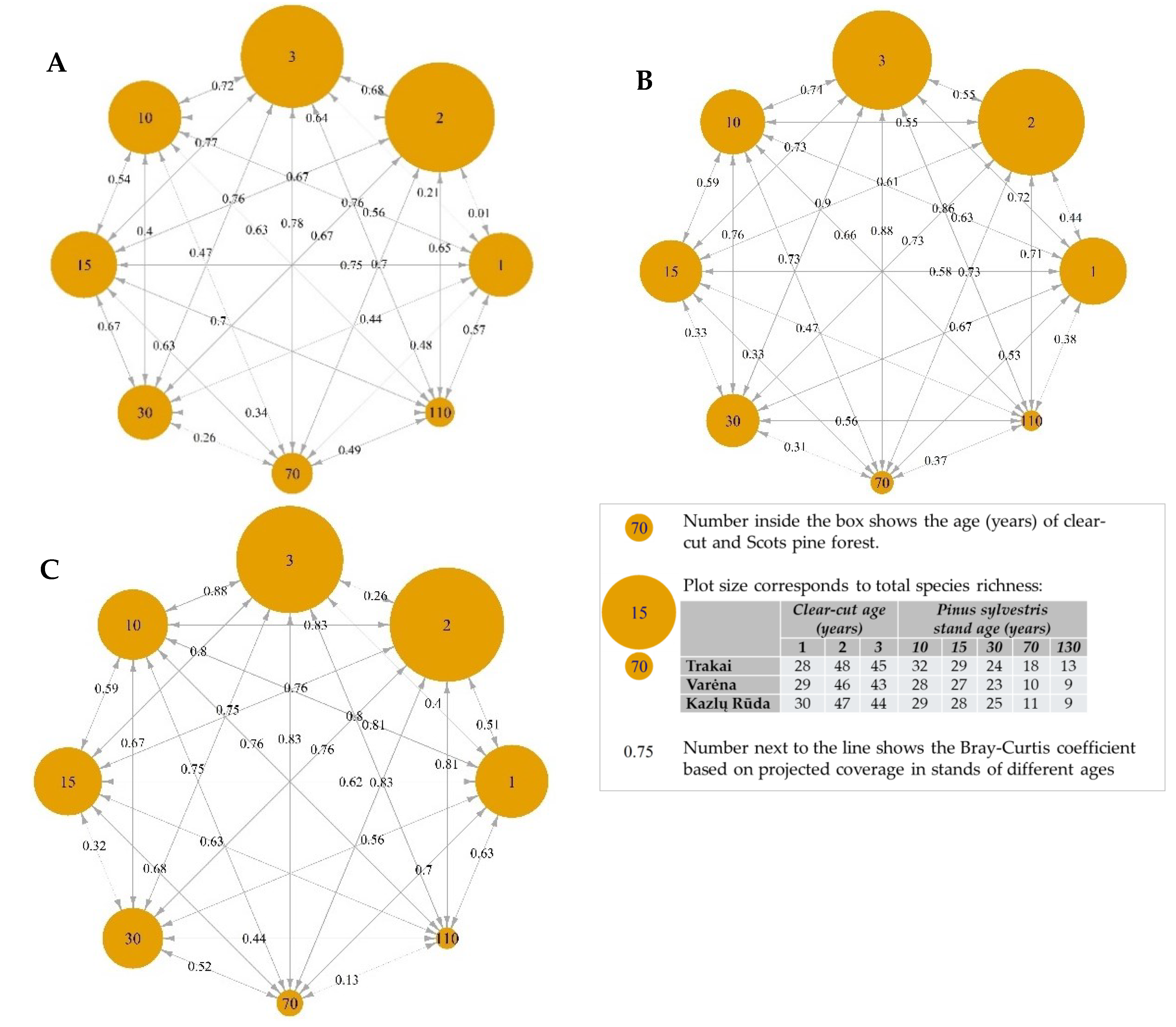
3.1. Change of Living Ground Cover at Different Forest Succession Stages
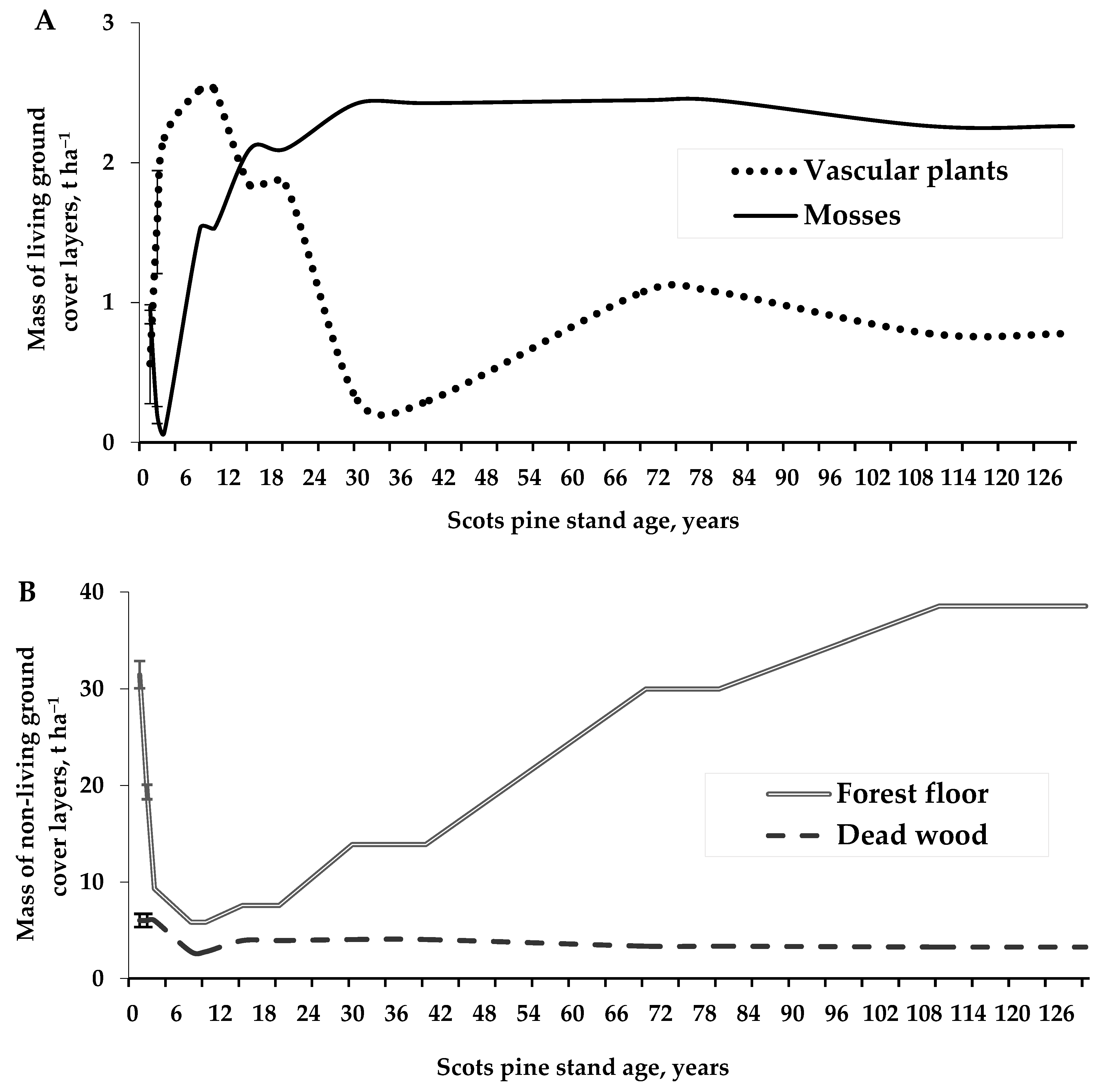
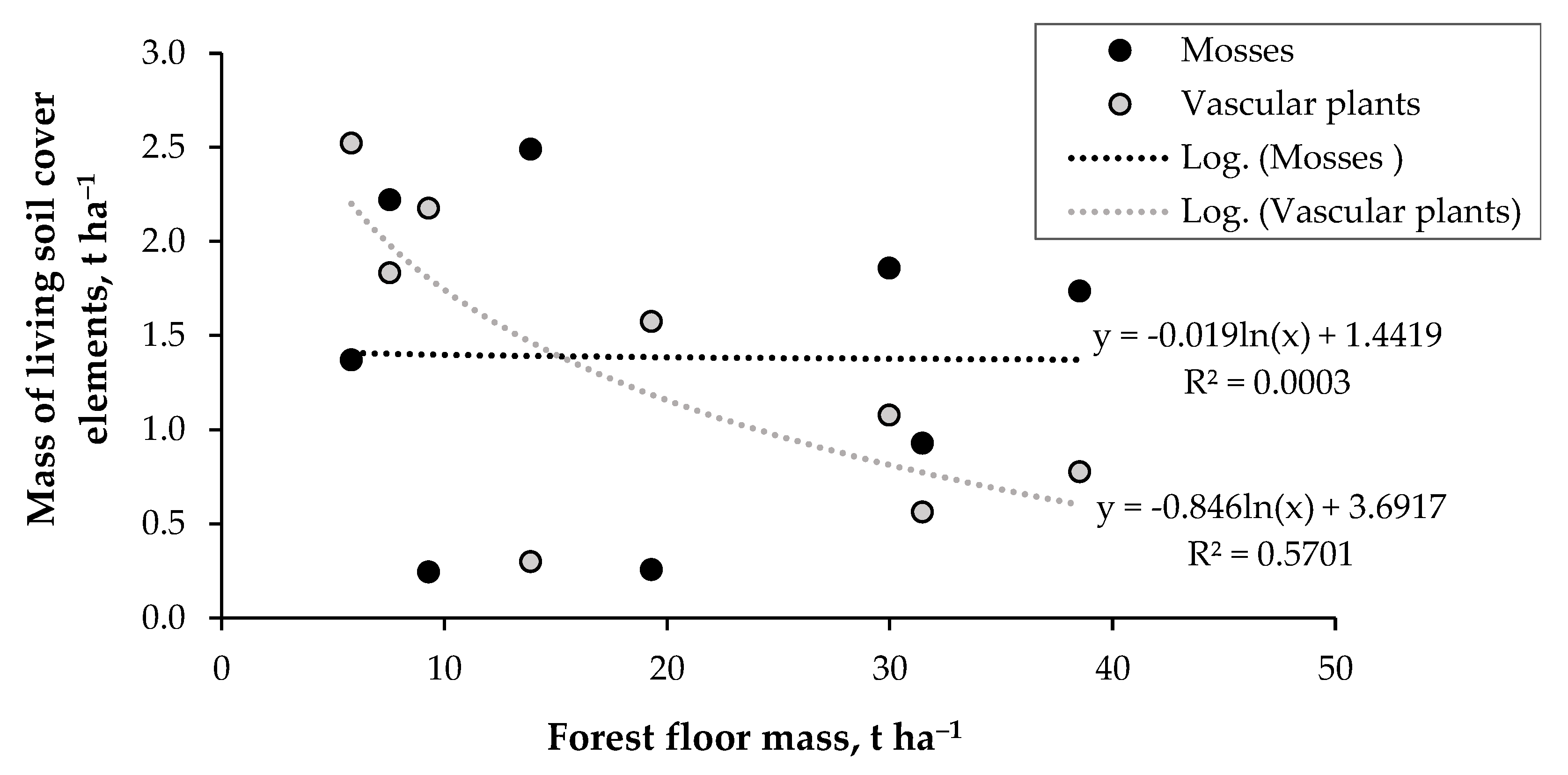
3.2. Ground Cover Layer Mass Dynamics and Relationships
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clarke, N.; Gundersen, P.; Jönsson-Belyazid, U.; Kjønaas, O.J.; Persson, T.; Sigurdsson, B.D.; Stupak, I.; Vesterdal, L. Influence of different tree-harvesting intensities on forest soil carbon stocks in boreal and northern temperate forest ecosystems. For. Ecol. Manag. 2015, 351, 9–19. [Google Scholar] [CrossRef]
- Česonienė, L.; Daubaras, R.; Bimbiraitė-Survilienė, K.; Kaškonienė, V.; Kaškonas, P.; Tiso, N.; Maruška, A.; Zych, M. Effects of clear-cuts in Scots pine-dominated forests on Vaccinium myrtillus and Vaccinium vitis-idaea vegetative characteristics, and accumulation of phenolic compounds. Balt. For. 2019, 24, 278–286. [Google Scholar]
- Mayer, M.; Prescott, C.E.; Abaker, W.E.A.; Augusto, L.; Cécillon, L.; Ferreira, G.W.D.; James, J.; Jandl, R.; Katzensteiner, K.; Laclau, J.-P.; et al. Tamm Review: Influence of forest management activities on soil organic carbon stocks: A knowledge synthesis. For. Ecol. Manag. 2020, 466, 118127. [Google Scholar] [CrossRef]
- Finér, L.; Mannerkoski, H.; Piirainen, S.; Starr, M. Carbon and nitrogen pools in an old-growth, Norway spruce mixed forest in eastern Finland and changes associated with clear-cutting. For. Ecol. Manag. 2003, 174, 51–63. [Google Scholar] [CrossRef]
- Palviainen, M. Logging residues and ground vegetation in nutrient dynamics of a clear-cut boreal forest. Diss. For. 2005, 12, 38. [Google Scholar] [CrossRef]
- Gundersen, P.; Schmidt, I.K.; Raulund-Rasmussen, K. Leaching of nitrate from temperate forests-effects of air pollution and forest management. Environ. Rev. 2006, 14, 1–57. [Google Scholar] [CrossRef]
- Hedwall, P.-O.; Grip, H.; Linder, S.; Lövdahl, L.; Nilsson, U.; Bergh, J. Effects of clear-cutting and slash removal on soil water chemistry and forest-floor vegetation in a nutrient optimised Norway spruce stand. Silva Fenn. 2013, 47, 933. [Google Scholar] [CrossRef]
- Häkkinen, M.; Heikkinen, J.; Mäkipää, R. Soil carbon stock increases in the organic layer of boreal middle-aged stands. Biogeosciences 2011, 8, 1279–1289. [Google Scholar] [CrossRef]
- Schelker, J.; Eklof, K.; Bishop, K.; Laudon, H. Effects of forestry operations on dissolved organic carbon concentrations and export in boreal first-order streams. J. Geophys. Res. Biogeosci. 2012, 117, G1. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, W.; Chen, H.; Mo, J. Impacts of nitrogen deposition on soil nitrogen cycle in forest ecosystems: A review. Acta Ecol. Sin. 2015, 35, 35–43. [Google Scholar] [CrossRef]
- Balandier, P.; Collet, C.; Miller, J.H.; Reynolds, P.E.; Zedaker, S.M. Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry 2005, 79, 3–27. [Google Scholar] [CrossRef]
- Palviainen, M.; Finér, L.; Mannerkoski, H.; Piirainen, S.; Starr, M. Responses of ground vegetation species to clear-cutting in a boreal forest: Aboveground biomass and nutrient contents during the first 7 years. Ecol. Res. 2005, 20, 652–660. [Google Scholar] [CrossRef]
- Parker, W.C.; Pitt, D.G.; Morneault, A.E. Influence of woody and herbaceous competition on microclimate and growth of eastern white pine (Pinus strobus L.) seedlings planted in a central Ontario clearcut. For. Ecol. Manag. 2009, 258, 2013–2025. [Google Scholar] [CrossRef]
- Vanha-Majamaa, I.; Shorohova, E.; Kushnevskaya, H.; Jalonen, J. Resilience of understory vegetation after variable retention felling in boreal Norway spruce forests—A ten-year perspective. For. Ecol. Manag. 2017, 393, 12–28. [Google Scholar] [CrossRef]
- Česonienė, L.; Daubaras, R.; Kaškonas, P.; Kaškonienė, V.; Maruška, A.S.; Tiso, N.; Zych, M. Initial impact of clear-cut logging on dynamics of understory vascular plants and pollinators in Scots pine-dominated forests in Lithuania. Turkish JAF Sci. Tech. 2018, 42, 433–443. [Google Scholar] [CrossRef]
- Moreaux, C.; Martin, P.; Hereş, A.-M.; Yuste, J.C. Effects of forestry practices on the conservation of soil and forest ecosystem health and functioning: An umbrella review protocol. OSFPreprints 2022, 1–44. [Google Scholar] [CrossRef]
- Stefańska-Krzaczek, E.; Szymura, T.H. Species diversity of forest floor vegetation in age gradient of managed Scots pine stands. Balt. For. 2015, 21, 233–243. [Google Scholar]
- Chen, J.; Saunders, S.C.; Crow, T.R.; Naiman, R.J.; Brosofske, K.D.; Mroz, G.D.; Brookshire, B.L.; Franklin, J.F. Microclimate in forest ecosystem and landscape ecology. BioScience 1999, 49, 288–297. [Google Scholar] [CrossRef]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Harvest impacts on soil carbon storage in temperate forests. For. Ecol. Manag. 2010, 259, 857–866. [Google Scholar] [CrossRef]
- Tonteri, T.; Salemaa, M.; Rautio, P.; Hallikainen, V.; Korpela, L.; Merilä, P. Forest management regulates temporal change in the cover of boreal plant species. For. Ecol. Manag. 2016, 381, 115–124. [Google Scholar] [CrossRef]
- Gustienė, D.; Varnagirytė-Kabašinskienė, I.; Stakėnas, V. Ground Vegetation in Pinus sylvestris Forests at different successional stages following clear cuttings: A case study. Plants 2022, 11, 2651. [Google Scholar] [CrossRef] [PubMed]
- Keenan, R.J.; Kimmins, J.P. The ecological effects of clear-cutting. Environ. Rev. 1993, 1, 121–144. [Google Scholar] [CrossRef]
- Karazija, S. Age-related dynamics of pine forest communities in Lithuania. Balt. For. 2002, 9, 50–62. [Google Scholar]
- Hart, S.A.; Chen, H.Y.H. Fire, logging, and overstory affect understory abundance, diversity, and composition in boreal forest. Ecol. Monogr. 2008, 78, 123–140. [Google Scholar] [CrossRef]
- Kumar, P.; Chen, H.Y.H.; Thomas, S.C.; Shahi, C. Linking resource availability and heterogeneity to understorey species diversity through succession in the boreal forest of Canada. J. Ecol. 2017, 106, 1266–1276. [Google Scholar] [CrossRef]
- Boch, S.; Prati, D.; Müller, J.; Socher, S.; Baumbach, H.; Buscot, F.; Gockel, S.; Hemp, A.; Hessenmöller, D.; Kalko, E.K.V.; et al. High plant species richness indicates management-related disturbances rather than the conservation status of forests. Basic Appl. Ecol. 2013, 14, 496–505. [Google Scholar] [CrossRef]
- Liu, X.; Bao, W.K. Understory plant assemblages present distinct short-term responses to the clear-cutting of an old-growth spruce forest near an alpine timberline. Can. J. For. Res. 2014, 44, 562–571. [Google Scholar] [CrossRef]
- Bačkaitis, J. Paprastosios Pušies (Pinus sylvestris L.) Žėlimui Įtaką Darantys Aplinkos Veiksniai: Daktaro disertacija [Environmental Factors Influencing Scots Pine (Pinus sylvestris L.) Reneneration. Ph.D. Thesis, Lithuanian University of Agriculture, Akademija, Lithuania, 2005. (In Lithuanian). [Google Scholar]
- Heinrichs, S.; Schmidt, W. Short-term effects of selection and clear-cutting on the shrub and herb layer vegetation during the conversion of even-aged Norway spruce stands into mixed stands. For. Ecol. Manag. 2009, 258, 667–678. [Google Scholar] [CrossRef]
- Kumar, P.; Chen, H.Y.H.; Searle, E.B.; Shahi, C. Dynamics of understorey biomass, production and turnover associated with long-term overstorey succession in Boreal forest of Canada. For. Ecol. Manag. 2018, 427, 152–161. [Google Scholar] [CrossRef]
- Petersson, L.; Holmström, E.; Lindbladh, M.; Felton, A. Tree species impact on understory vegetation: Vascular plant communities of Scots pine and Norway spruce managed stands in northern Europe. For. Ecol. Manag. 2019, 448, 330–345. [Google Scholar] [CrossRef]
- Juodvalkis, A.; Kairiūkštis, L. Medynų Formavimas ir Kirtimai [Stand Forming and Felling]; Lututė: Kaunas, Lithuania, 2009. (In Lithuanian) [Google Scholar]
- Vaičys, M.; Karazija, S.; Kuliešis, A.; Rutkauskas, A. Miškų Augavietės. Miško Augaviečių Tipai [Forest Sites. Forest Site Types]; Lututė: Kaunas, Lithuania, 2006; p. 95. (In Lithuanian) [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014; International Soil Classification System for Naming 615 Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015; p. 192. [Google Scholar]
- Karazija, S. Lietuvos Miškų Tipai [Lithuanian Forest Types]; Mokslas: Vilnius, Lithuania, 1988. (In Lithuanian) [Google Scholar]
- Gustienė, D. Dirvožemio Dangos Dinamika Paprastosios Pušies Medynuose po Plynųjų Kirtimų [Dynamics of Ground Cover in Scots Pine Stands after Clear Cuttings]. Ph.D. Thesis, Vytautas Magnus University, Kaunas, Lithuania, 2024. (In Lithuanian with English Summary). [Google Scholar]
- Ehlers, T.; Berch, S.; Mackinnon, A. Inventory of non-timber forest product plant and fungal species in the Robson Valley. BC J. Ecosyst. Manag. 2003, 4, 1–15. [Google Scholar] [CrossRef]
- Stakėnas, V.; Varnagirytė-Kabašinskienė, I.; Sirgedaitė-Šėžienė, V.; Armolaitis, K.; Araminienė, V.; Muraškienė, M.; Žemaitis, P. Dead wood density and carbon estimates for the main tree species in the Lithuanian hemiboreal forest. Eur. J. For. Res. 2020, 139, 1045–1055. [Google Scholar] [CrossRef]
- Gustienė, D.; Varnagirytė-Kabašinskienė, I.; Stakėnas, V. Ground vegetation, forest floor and mineral topsoil in a clear-cutting and reforested Scots pine stands of different ages: A case study. J. For. Res. 2022, 33, 1247–1257. [Google Scholar] [CrossRef]
- Hart, S.A.; Chen, H.Y.H. Understory vegetation dynamics of North American boreal forests. Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Arduini, I.; Ercoli, L. Recovery of understory vegetation in clear-cut stone pine (Pinus pinea L.) plantations. Plant Biosyst. 2012, 146, 244–258. [Google Scholar] [CrossRef]
- Reich, P.B.; Frelich, L.E.; Voldseth, R.A.; Bakken, P.; Adair, E.C. Understorey diversity in southern boreal forests is regulated by productivity and its indirect impacts on resource availability and heterogeneity. J. Ecol. 2011, 100, 539–545. [Google Scholar] [CrossRef]
- Halpern, C.B.; Lutz, J.A. Canopy closure exerts weak controls on understory dynamics: A 30-year study of overstory–understory interactions. Ecol. Monogr. 2013, 83, 221–237. [Google Scholar] [CrossRef]
- Pykälä, J. Immediate increase in plant species richness after clear-cutting of boreal herb-rich forests. Appl. Veg. Sci. 2004, 7, 29–34. [Google Scholar] [CrossRef]
- Bainbridge, E.L.; Strong, W.L. Pinus contorta understory vegetation dynamics following clearcutting in west-central Alberta, Canada. For. Ecol. Manag. 2005, 213, 133–150. [Google Scholar] [CrossRef]
- Kelly, D.L.; Connolly, A. A review of the plant communities associated with Scots pine (Pinus sylvestris L.) in Europe, and an evaluation of putative indicator/specialist species. For. Syst. 2000, 9, 15–39. [Google Scholar] [CrossRef]
- Navasaitis, M.; Ozolinčius, R.; Smaliukas, D.; Balevičienė, J. Lietuvos Dendroflora [Dendroflora of Lithuania]; Lututė: Kaunas, Lithuania, 2003. (In Lithuanian) [Google Scholar]
- Česonienė, L.; Daubaras, R.; Tamutis, V.; Kaškonienė, V.; Kaškonas, P.; Stakėnas, V.; Zych, M. Effect of clear-cutting on the understory vegetation, soil and diversity of litter beetles in Scots pine-dominated forest. J. Sustain. For. 2019, 38, 791–808. [Google Scholar] [CrossRef]
- Fierer, N.; Strickland, M.S.; Liptzin, D.; Bradford, M.A.; Cleveland, C.C. Global patterns in belowground communities. Ecol. Lett. 2009, 12, 1238–1249. [Google Scholar] [CrossRef] [PubMed]
- Waring, B.; Neumann, M.; Prentice, I.C.; Adams, M.; Smith, P.; Siegert, M. Forests and decarbonization–Roles of natural and planted forests. Front. For. Glob. Chang. 2020, 3, 534891. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y.H. Is understory plant species diversity driven by resource quantity or resource heterogeneity? Ecology 2010, 91, 1931–1938. [Google Scholar] [CrossRef]
- Arróniz-Crespo, M.; Bougoure, J.; Murphy, D.V.; Cutler, N.A.; Souza-Egipsy, V.; Chaput, D.L.; Jones, D.L.; Ostle, N.; Wade, S.C.; Clode, P.L.; et al. Revealing the transfer pathways of cyanobacterial-fixed N into the boreal forest through the feather-moss microbiome. Front. Plant Sci. 2022, 13, 1036258. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Object | Stand Composition * | Stand Age (Years) | DBH (cm) | Height (m) | Stocking Level *** | Volume (m3 ha−1) |
|---|---|---|---|---|---|---|
| Trakai 54°44′ N, 24°80′ E 54°44′ N, 24°80′ E | 90P10E | 2 ** | - | - | 6000 trees ha−1 | - |
| 90P10E | 2 ** | - | - | 6000 trees ha−1 | - | |
| 100P | 10 | 5 | 4.5 | 0.9 | 20 | |
| 100P | 15 | 9 | 7 | 0.8 | 60 | |
| 90P10E | 30 | 14 | 15 | 0.8 | 160 | |
| 100P | 70 | 25 | 26 | 0.7 | 280 | |
| 100P | 135 | 46 | 30 | 0.7 | 415 | |
| Varėna 54°26′ N, 24°53′ E 54°25′ N, 24°53′ E | 90P10B | 2 ** | - | - | 6000 trees ha−1 | - |
| 100P | 2 ** | - | - | 6000 trees ha−1 | - | |
| 90P10B | 8 | 3 | 2.5 | 0.9 | 14 | |
| 90P10B | 15 | 5 | 6 | 1.0 | 50 | |
| 100P | 39 | 13 | 16 | 0.8 | 120 | |
| 100P | 70 | 21 | 24 | 0.9 | 300 | |
| 100P | 110 | 34 | 27 | 0.8 | 360 | |
| Kazlų Rūda 54°76′ N, 23°40′ E 54°73′ N, 23°47′ E | 100P | 2 ** | - | - | 7000 trees ha−1 | - |
| 100P | 2 ** | - | - | 5000 trees ha−1 | - | |
| 90P10B | 8 | 4 | 3.1 | 0.9 | 19 | |
| 90P10B | 15 | 8 | 4.8 | 0.9 | 70 | |
| 90P10B | 30 | 14 | 15 | 0.9 | 150 | |
| 100P | 77 | 29 | 28 | 0.9 | 400 | |
| 90P10E | 117 | 38 | 31 | 0.7 | 320 |
| Object | Ground Cover Layer | Clear-Cut (Years) | Scots Pine Stands (Years) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 8–10 | 15–20 | 30–40 | 70–80 | 110–130 | ||
| Cover (%) | |||||||||
| Trakai | Mosses | 56.2 ± 4.2 c | 18.1 ± 3.3 b | 6.5 ± 1.2 a | 53.0 ± 4.4 c | 59.8 ± 6.3 c | 92.5 ± 1.3 d | 96.0 ± 2.4 d | 88.9 ± 3.5 d |
| Vascular plants | 14.5 ± 2.1 a | 23.2 ± 2.3 b | 52.2 ± 2.2 d | 59.0 ± 4.4 d | 34.6 ± 1.4 c | 24.1 ± 7.1 b | 39.6 ± 3.1 c | 16.6 ± 3.1 a | |
| Forest floor | 95.8 ± 1.7 cd | 39.6 ± 4.7 a | 56.2 ± 5.3 b | 80.1 ± 4.8 c | 99.8 ± 0.1 d | 100.0 ± 0.0 d | 100.0 ± 0.0 d | 100.0 ± 0.0 d | |
| Varėna | Mosses | 44.0 ± 5.7 c | 12.0 ± 3.2 b | 2.6 ± 0.5 a | 43.0 ± 4.5 c | 64.9 ± 4.2 d | 96.0 ± 2.6 e | 92.6 ± 1.5 e | 90.3 ± 3.5 e |
| Vascular plants | 15.2 ± 1.7 a | 16. 7 ± 1.7 a | 40.7 ± 3.2 c | 28.6 ± 3.8 b | 23.8 ± 2.7 b | 12.4 ± 3.0 a | 21.3 ± 3.6 b | 23.3 ± 4.6 b | |
| Forest floor | 65.8 ± 5.2 b | 35.7 ± 5.2 a | 35.9 ± 4.7 a | 34.6 ± 7.7 a | 86.3 ± 3.3 c | 100.0 ± 0.0 d | 100.0 ± 0.0 d | 99.9 ± 0.2 d | |
| Kazlų Rūda | Mosses | 18.8 ± 2.0 c | 10.9 ± 0.8 b | 6.5 ± 0.5 a | 27.8 ± 1.7 d | 77.8 ± 4.3 e | 87.4 ± 6.8 e | 97.9 ± 11.9 f | 93.3 ± 9.8 f |
| Vascular plants | 33.6 ± 0.7 c | 29.6 ± 0.8 b | 66.9 ± 1.1 d | 53.2 ± 2.4 d | 28.2 ± 0.8 b | 21.5 ± 0.6 a | 34.9 ± 1.6 c | 26.7 ± 1.5 b | |
| Forest floor | 68.2 ± 4.1 a | 56.2 ± 3.9 a | 79.8 ± 4.2 b | 92.2 ± 5.5 c | 99.2 ± 0.6 c | 99.5 ± 0.5 c | 94.8 ± 5.0 c | 99.9 ± 0.2 c | |
| Species of Vascular Plants/Mosses | Clear-Cuts (Years) | Scots Pine Stands (Years) | ||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 8–10 | 15–20 | 30–40 | 70–80 | 110–130 | |
| Prominence Value | ||||||||
| Vaccinium myrtillus L. | 76.81 ± 31.57 | 46.38 ± 15.71 | 54.30 ± 19.07 | 25.81 ± 21.03 | 19.85 ± 9.42 | 88.02 ± 32.30 | 205.20 ± 64.96 | 148.82 ± 58.96 |
| Vaccinium vitis-idaea L. | 17.08 ± 7.53 | 16.43 ± 8.27 | 32.02 ± 15.25 | 174.07 ± 53.06 | 122.32 ± 64.94 | 44.28 ± 11.64 | 85.43 ± 37.94 | 44.37 ± 24.73 |
| Calluna vulgaris L. (Hull) | 12.24 ± 12.07 | 2.17 ± 1.96 | 3.55 ± 1.79 | 159.68 ± 73.25 | 20.63 ± 12.22 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Rubus idaeus L. | 6.35 ± 6.07 | 27.71 ± 25.30 | 111.35 ± 81.78 | 15.34 ± 15.10 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Festuca sp. | 20.92 ± 20.56 | 19.29 ± 19.28 | 84.48 ± 82.18 | 16.88 ± 16.81 | 34.40 ± 34.40 | 11.46 ± 11.46 | 3.69 ± 3.69 | 6.33 ± 6.33 |
| Pteridium aquilinum (L.) | 10.14 ± 0.34 | 17.81 ± 2.16 | 23.23 ± 9.02 | 0.00 ± 0.00 | 47.48 ± 47.48 | 4.85 ± 4.85 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Pleurozium schreberi (Brid.) Mitt. | 200.71 ± 65.92 | 75.01 ± 29.23 | 12.58 ± 4.92 | 164.89 ± 76.84 | 323.01 ± 109.80 | 499.69 ± 15.99 | 478.67 ± 198.73 | 207.64 ± 42.94 |
| Hylocomium splendens (Hedw.) Scimp. | 85.96 ± 49.44 | 5.34 ± 4.37 | 1.29 ± 0.16 | 31.10 ± 26.79 | 33.84 ± 31.12 | 167.31 ± 84.32 | 323.81 ± 219.68 | 552.38 ± 77.06 |
| Dicranum sp. | 24.43 ± 11.83 | 11.97 ± 7.35 | 7.48 ± 6.26 | 20.68 ± 4.57 | 126.16 ± 13.03 | 157.36 ± 119.17 | 42.96 ± 36.84 | 20.68 ± 13.27 |
| Ptilium crista-castrensis (Hedw.) De Not | 4.98 ± 4.54 | 6.47 ± 6.46 | 5.09 ± 5.09 | 10.20 ± 10.20 | 9.88 ± 8.13 | 11.66 ± 11.03 | 50.17 ± 25.03 | 31.60 ± 28.85 |
| Cirriphyllum piliferum (Hed.) Grout | 00.00 ± 0.00 | 00.00 ± 0.00 | 00.00 ± 0.00 | 7.30 ± 7.30 | 83.64 ± 83.64 | 4.64 ± 4.64 | 00.00 ± 0.00 | 00.00 ± 0.00 |
| Polytrichum sp. | 00.00 ± 0.00 | 0.41 ± 0.36 | 0.47 ± 0.47 | 67.82 ± 44.00 | 12.83 ± 11.49 | 2.28 ± 2.16 | 6.91 ± 6.91 | 00.00 ± 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gustienė, D.; Varnagirytė-Kabašinskienė, I. Changes in Ground Cover Layers, Biomass and Diversity of Vascular Plants/Mosses in the Clear-Cuts Followed by Reforested Scots Pine until Maturity Age. Land 2024, 13, 1477. https://doi.org/10.3390/land13091477
Gustienė D, Varnagirytė-Kabašinskienė I. Changes in Ground Cover Layers, Biomass and Diversity of Vascular Plants/Mosses in the Clear-Cuts Followed by Reforested Scots Pine until Maturity Age. Land. 2024; 13(9):1477. https://doi.org/10.3390/land13091477
Chicago/Turabian StyleGustienė, Dovilė, and Iveta Varnagirytė-Kabašinskienė. 2024. "Changes in Ground Cover Layers, Biomass and Diversity of Vascular Plants/Mosses in the Clear-Cuts Followed by Reforested Scots Pine until Maturity Age" Land 13, no. 9: 1477. https://doi.org/10.3390/land13091477
APA StyleGustienė, D., & Varnagirytė-Kabašinskienė, I. (2024). Changes in Ground Cover Layers, Biomass and Diversity of Vascular Plants/Mosses in the Clear-Cuts Followed by Reforested Scots Pine until Maturity Age. Land, 13(9), 1477. https://doi.org/10.3390/land13091477