
Effects of Enrofloxacin and Ciprofloxacin on Growth and Toxin Production of Microcystis aeruginosa



Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Material
2.1.1. Algal Species Tested
2.1.2. Test Drugs
2.2. Methods
2.2.1. Preparation of BG11 Medium
2.2.2. Culture of M. aeruginosa
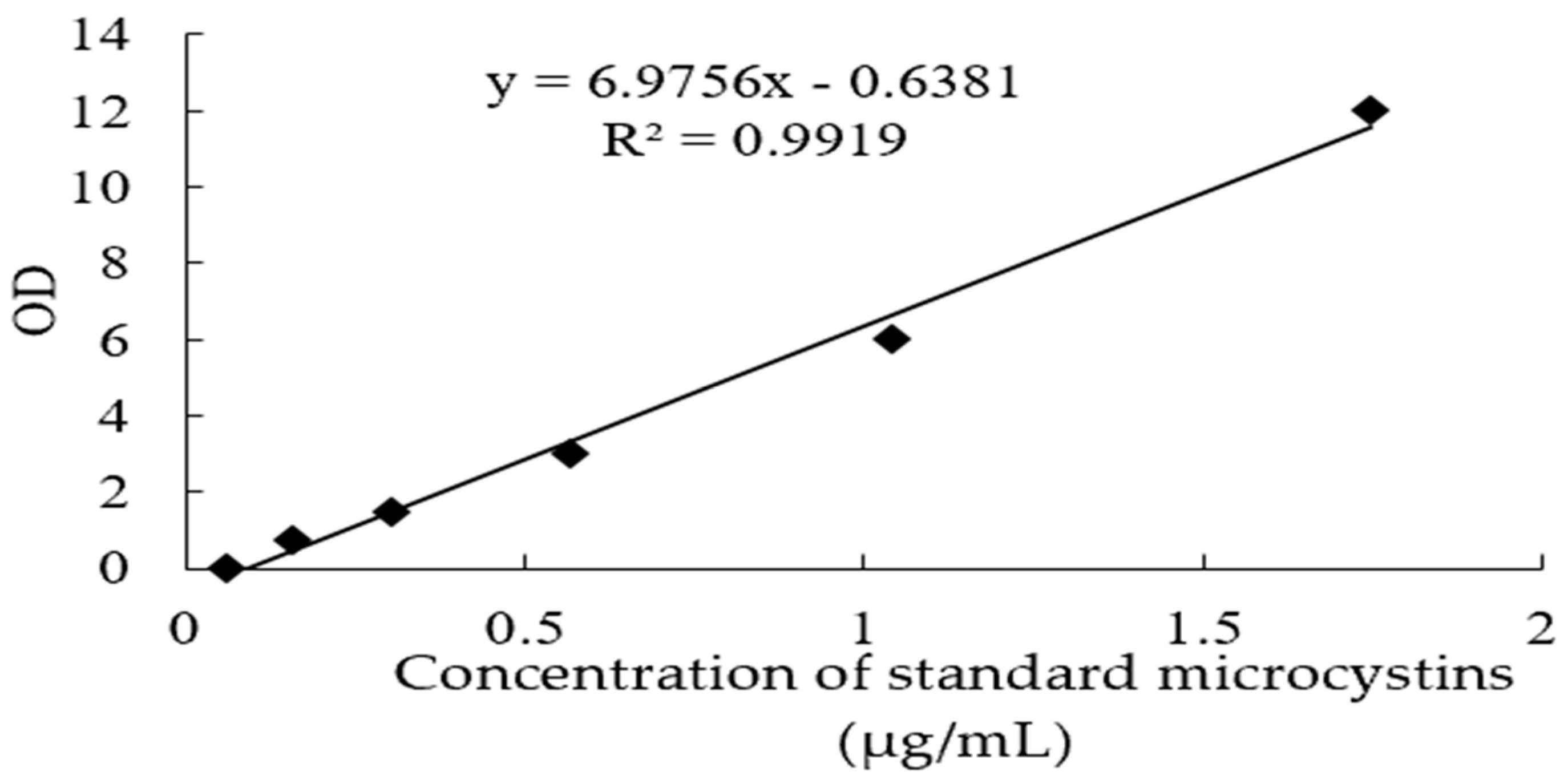
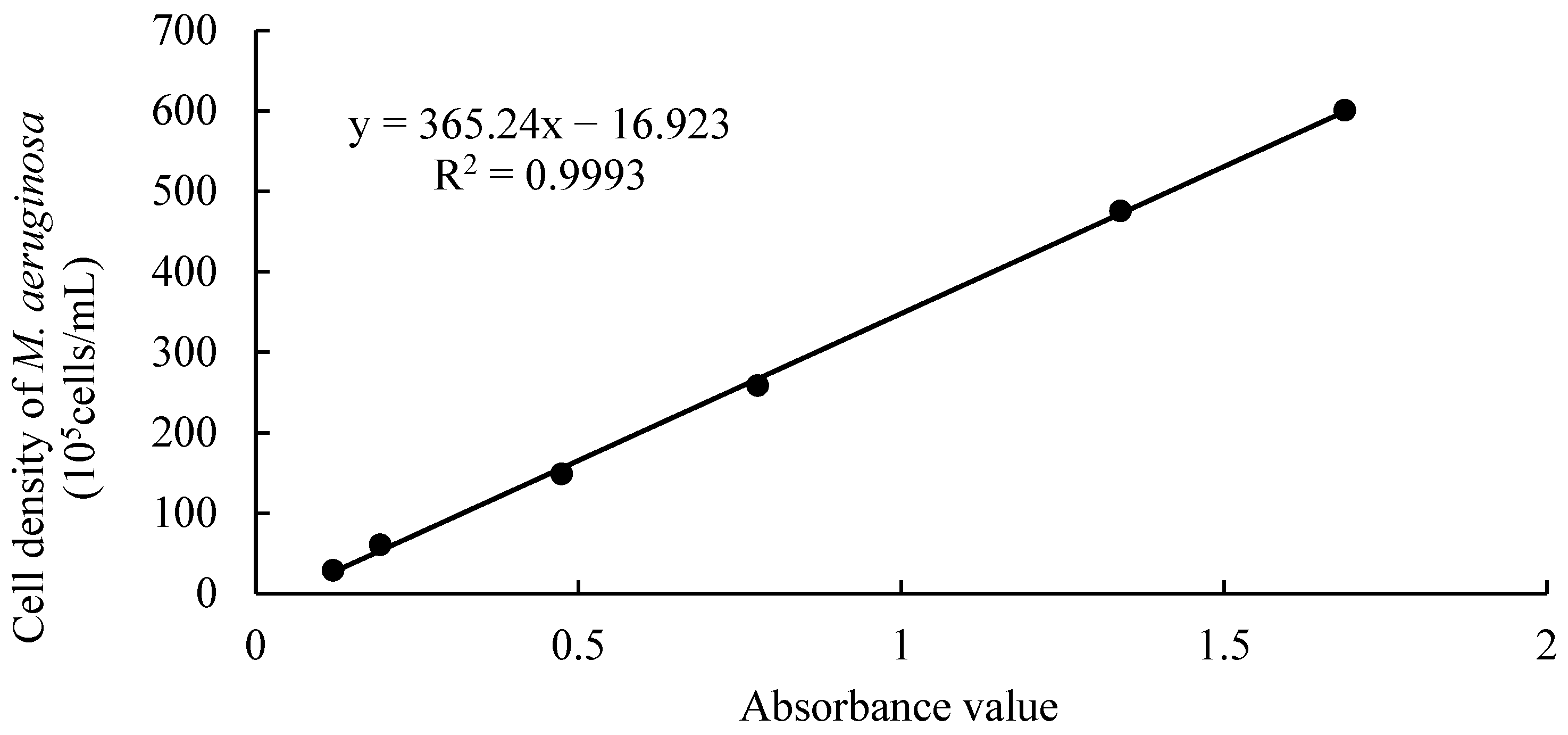
2.2.3. Establishment of Standard Curve
2.2.4. Toxicity Test of EF and CF on M. aeruginosa
2.2.5. Effects of EF and CF on the Toxicity Production of M. aeruginosa
2.3. Data Processing and Statistics
3. Results
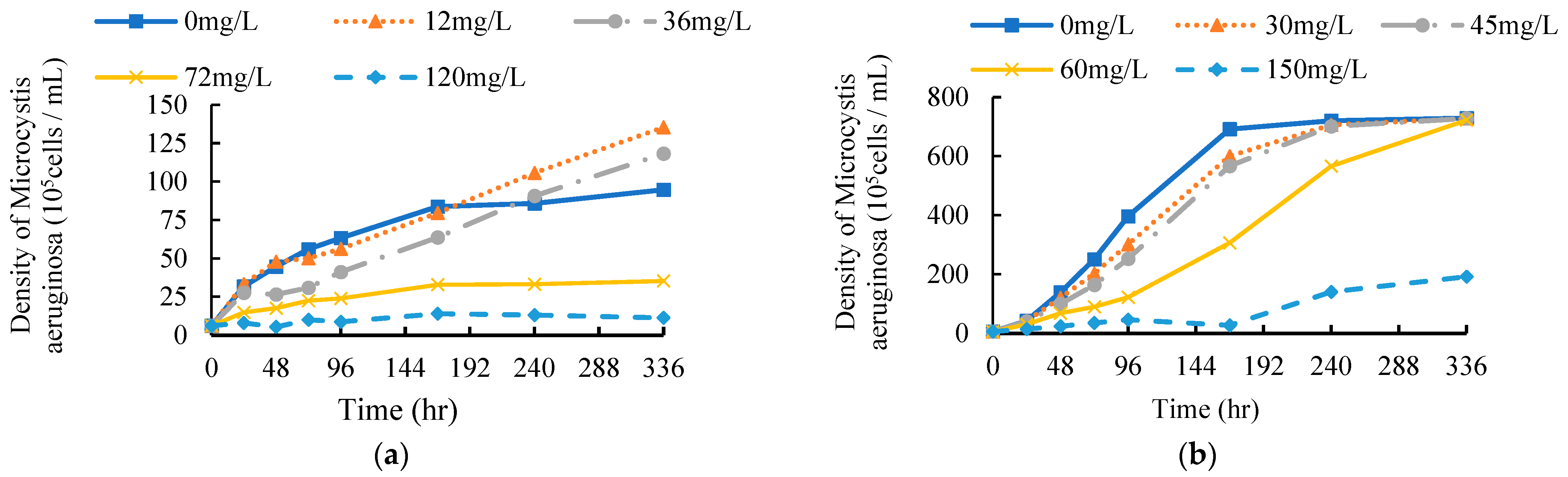
3.1. Effects of EF and CF on the Population Density of M. aeruginosa
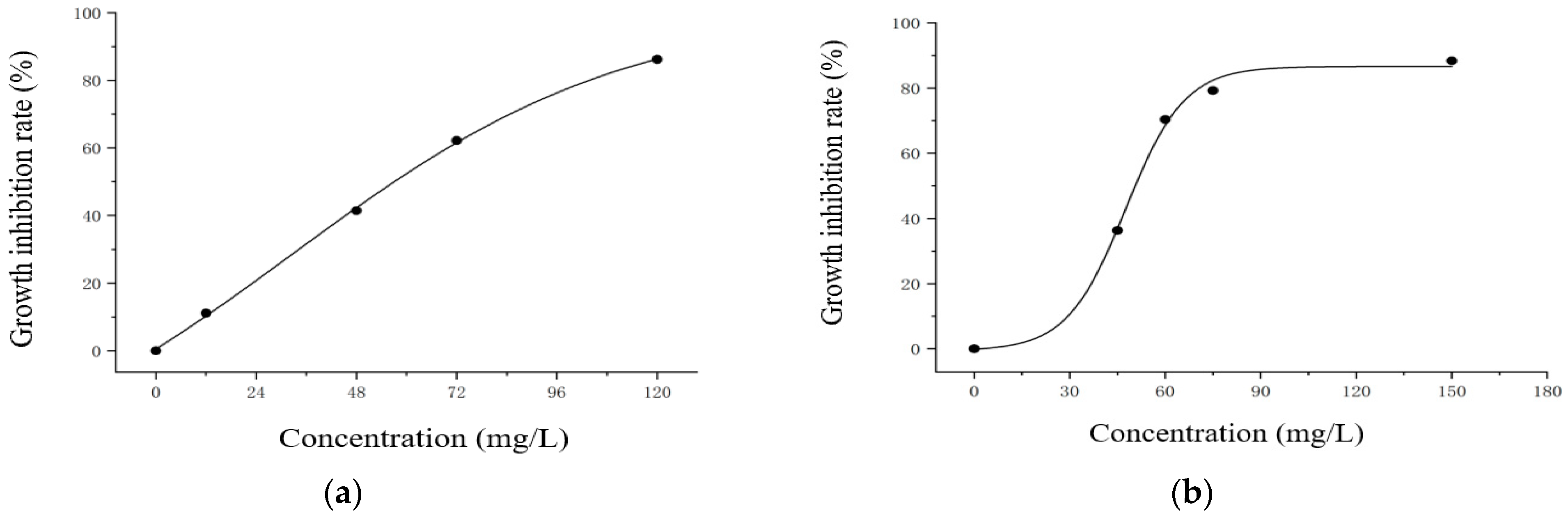
3.2. EC50 Values of EF and CF on M. aeruginosa
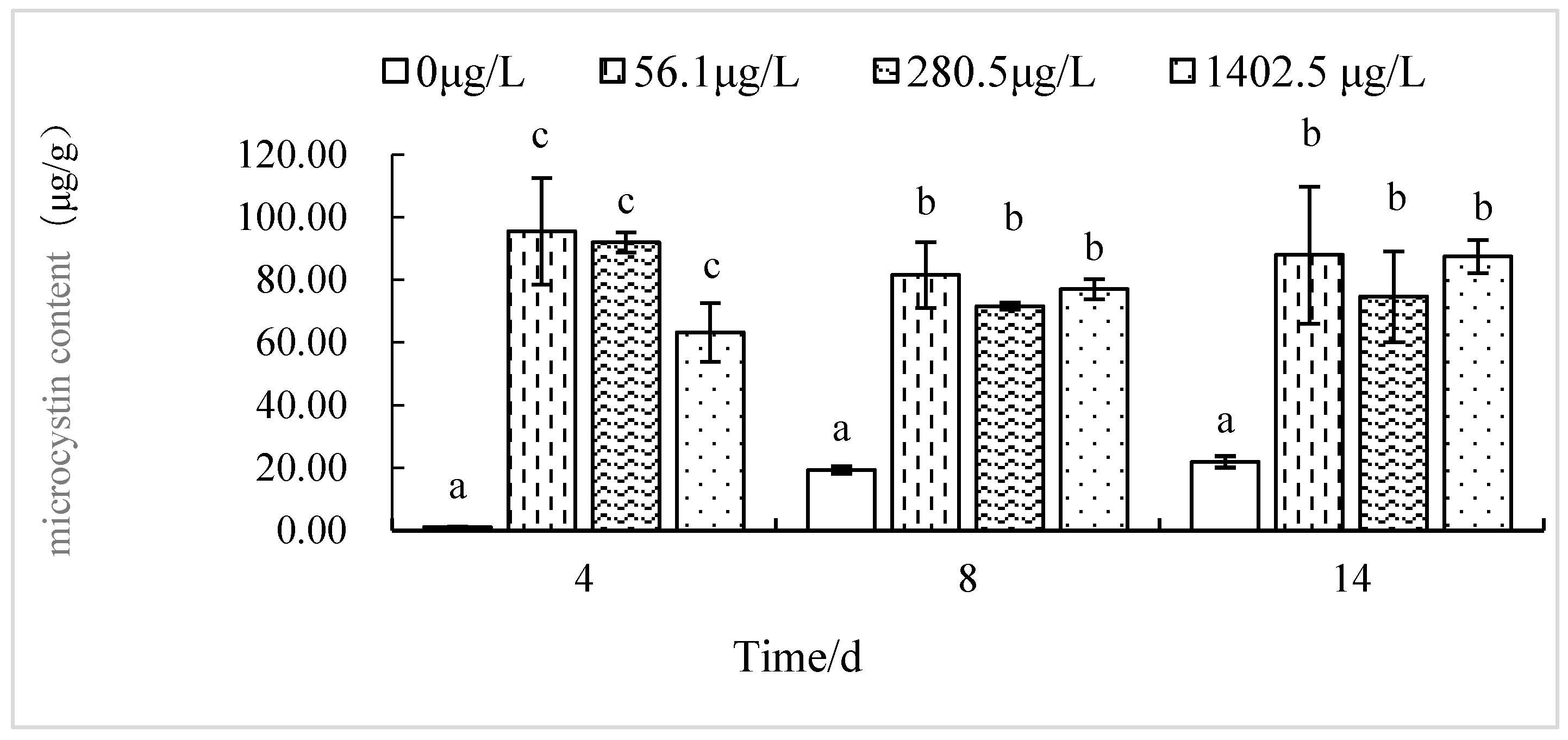
3.3. Effects of EF and CF on the Toxin Production of M. aeruginosa
4. Discussion
4.1. Toxic Effects of Antibiotics on M. aeruginosa
4.2. Effect of Antibiotics on Toxin Production by M. aeruginosa
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, Z.G. Distribution and Purification of Pharmaceutical and Personal Care Products (PPCPs) in Drinking Water. Ph.D. Thesis, Tsinghua University, Beijing, China, 2011. [Google Scholar]
- Chen, D.J. Industrialization of antibiotics in China and global research and development of new drugs. In Proceedings of the 7th China Summit Forum on Industrial Biotechnology Development, Tianjin, China, 9–12 May 2013. [Google Scholar]
- Zhang, Q.Q.; Ying, G.G.; Pan, C.G.; Liu, Y.S.; Zhao, J.L. Comprehensive evaluation of antibiotics emission and fate in the river basins of China: Source analysis, multimedia modeling, and linkage to bacterial resistance. Environ. Sci. Technol. 2015, 49, 6772–6782. [Google Scholar] [CrossRef]
- Tang, Q.; Song, P.; Li, J.J.; Kong, F.L.; Sun, L.; Xu, L.Z. Control of antibiotic resistance in China must not be delayed: The current state of resistance and policy suggestions for the government, medical facilities, and patients. Biosci. Trends 2016, 10, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Zhang, M.; Wang, K.; Zhu, K.; Xu, C.; Xie, J.; Xu, J. Aquaculture impacts on China’s marine wild fisheries over the past 30 years. Front. Mar. Sci. 2021, 8, 710124. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, J.H.; Zhao, Y.; Wang, G.X. Antimicrobial susceptibility test of 20 traditional Chinese medicines against Vibrio anguillarum. Prog. Vet. Med. 2005, 26, 77–79. [Google Scholar]
- Polianciuc, S.I.; Gurzău, A.E.; Kiss, B.; Ştefan, M.G.; Loghin, F. Antibiotics in the environment: Causes and consequences. Med. Pharm. Rep. 2020, 93, 231–240. [Google Scholar] [CrossRef]
- Lalumera, G.M.; Calamari, D.; Galli, P.; Castiglioni, S.; Crosa, G.; Fanelliet, R. Preliminary investigation on the environmental occurrence and effects of antibiotics used in aquaculture in Italy. Chemosphere 2004, 54, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Verlicchi, P.; Galletti, A.; Petrovic, M.; Barceló, D. Hospital effluents as a source of emerging pollutants: An overview of micropollutants and sustainable treatment options. J. Hydrol. 2010, 389, 416–428. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Song, C.; Chen, C.C. Research progress on the application of quinolone antibiotics in aquaculture. Jiangsu Agric. Sci. 2019, 47, 32–36. [Google Scholar]
- He, X.; Wang, Z.; Nie, X.; Yang, Y.; Pan, D.; Leung, A.O.; Cheng, Z.; Yang, Y.; Li, K.; Chen, K. Residues of fluoroquinolones in marine aquaculture environment of the Pearl River Delta, South China. Environ. Geochem. Health 2012, 34, 323–335. [Google Scholar] [CrossRef]
- Zhang, Q.; Xin, Q.; Zhu, J.M. The antibiotic contaminations in the main water bodies in China and the associated environmental and human health impacts. Environ. Chem. 2014, 33, 1075–1083. [Google Scholar]
- Yuan, S.G.; Cui, Y.F.; Zhang, W.J. Residual levels of antibiotics in aquatic products in Beijing market. Asian J. Ecotoxicol. 2015, 10, 311–317. [Google Scholar]
- He, X.T.; Deng, M.C.; Wang, Q.; Yang, Y.T.; Yang, Y.F.; Nie, X.P. Residues and health risk assessment of quinolones and sulfonamides in cultured fish from Pearl River Delta, China. Aquaculture 2016, 458, 38–46. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Ali, S.S.; Ramadan, H.; El-Aswar, E.I.; Eltawab, R.; Ho, S.H.; Elsamahy, T.; Li, S.; El-Sheekh, M.M.; Schagerl, M.; et al. Microalgae-based wastewater treatment: Mechanisms, challenges, recent advances, and future prospects. Environ. Sci. Ecotechnol. 2022, 8, 100205. [Google Scholar] [CrossRef] [PubMed]
- Cid, A.; Prado, R.; Rioboo, C.; Suarez-Bregua, P.; Herrero, C. Use of Microalgae as Biological Indicators of Pollution: Looking for New Relevant Cytotoxicity Endpoints. In Microalgae: Biotechnology, Microbiology and Energy; Johnsen, M.N., Ed.; Nova Science Publishers: New York, NY, USA, 2012; pp. 311–323. [Google Scholar]
- Ingrid, C.; Jamie, B. Toxic Cyanobacteria in Water; E&FN Spon Publisher: London, UK; New York, NY, USA, 1999; 416p. [Google Scholar]
- Su, Y.L.; Deng, Y.R. Microcystins in eutrophic lakes and their controlling and removing methods. Environ. Sci. Technol. 2013, 36, 62–66+84. [Google Scholar]
- Singh, D.P.; Tyagi, M.B.; Kumar, A.; Thakur, J.K.; Kumar, A. Antialgal activity of a hepatotoxin-producing cyanobacterium, Microcystis aeruginosa. World J. Microbiol. 2001, 17, 15–22. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, M.; Wang, R.N. The temporal and spation variation of the cyanobacteria which caused the water bloom in the Dianchi Lake, Kunming‚ China. J. Yunnan Univ. 2005, 3, 272–276. (In Chinese) [Google Scholar]
- Sukenik, A.; Kaplan, A. Cyanobacterial harmful algal blooms in aquatic ecosystems: A comprehensive outlook on current and emerging mitigation and control approaches. Microorganisms 2021, 9, 1472. [Google Scholar] [CrossRef]
- Yang, W.W.; Hien, V.T.T.; Wu, Y.X.; Zhang, W.H. Toxicity of enrofloxacin and erythromycin thiocyanate on Microcystis aeruginosa. China Environ. Sci. 2013, 33, 1829–1834. [Google Scholar]
- Available online: https://microbiology.ucdavis.edu/meeks/BG11medium.html (accessed on 9 October 2023).
- Xu, X.Y.; Cheng, T.Y.; Chen, L.; Zhang, W.; Liu, T.Z. Effects of phosphorus on Haematococcus pluvialis cell propagation and differentiation in two mediums. Chin. J. Process Eng. 2016, 16, 840–848. (In Chinese) [Google Scholar] [CrossRef]
- Liu, D.; Zeng, H.H.; Deng, Y. Combined stress toxicity of binary heavy metal mixture to Scenedesmus obliquus. Sci. Technol. Eng. 2019, 19, 374–383. (In Chinese) [Google Scholar]
- Shen, P.P.; Wang, Z.H.; Qi, Y.Z.; Xie, L.C.; Wang, Y. An optical density method for determination of microalgal biomass. J. Jinan Univ. Nat. Sci. 2001, 22, 115–119. (In Chinese) [Google Scholar]
- Lv, X.Y.; Zhang, E.; Yang, Y. Methodological research on measuring chlorella quantity by spectrophotometry. J. Anhui Agri. Sci. 2009, 37, 11104–11105. (In Chinese) [Google Scholar]
- Gu, D.X.; Song, G.L.; Ge, L.Y.; Deng, H.H.; Zhang, M.H. Study on toxicology of two antibiotics on Isochrysis galbana. Zhejiang Agri. Sci. 2013, 02, 179–182. [Google Scholar]
- MCs ElISA Kit (Shanghai Keshun Biotechnology Co., Ltd., Shanghai, China). Available online: https://ml.mbd.baidu.com/r/17wDuAsAPgQ?f=cp&u=cb76288e42ef58ca/ (accessed on 9 October 2023).
- van Meerloo, J.; Kaspers, G.J.; Cloos, J. Cell sensitivity assays: The MTT assay. Methods Mol. Biol. 2011, 731, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Plumb, J.A. Cell sensitivity assays: The MTT assay. Methods Mol. Med. 2004, 88, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Carraschi, S.P.; Cruz, C.D.; Basile, A.G.; Pitelli, R.A. Effects of fungicides for non target fungi Alternaria cassiae. Int. J. Environ. Agric. Biotech. 2017, 2, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Y.; Huang, Q.F.; Wang, Q.; Li, X.J.; Duan, H.B. Hazardous waste classification and management. Environ. Pollut. Prev. 2006, 1, 34–36. (In Chinese) [Google Scholar]
- Kasai, K.; Kanno, T.; Endo, Y.; Endo, Y.; Wakasa, K.; Tozawa, Y. Guanosine tetra- and pentaphosphate synthase activity in chloroplasts of a higher plant: Association with 70S ribosomes and inhibition by tetracycline. Nucleic Acids Res. 2004, 32, 5732–5741. [Google Scholar] [CrossRef]
- Lopes, T.O.M.; Passos, L.S.; Vieira, L.V.; Pinto, E.; Dorr, F.; Scherer, R.; de Andrade Salustriano, N.; Carneiro, M.T.W.D.; Postay, L.F.; Gomes, L.C. Metals, arsenic, pesticides, and microcystins in tilapia (Oreochromis niloticus) from aquaculture parks in Brazil. Environ. Sci. Pollut. Res. Int. 2020, 27, 20187–20200. [Google Scholar] [CrossRef]
- Pan, Y.; Dong, J.; Wan, L.; Sun, S.; MacIsaac, H.J.; Drouillard, K.G.; Chang, X. Norfloxacin pollution alters species composition and stability of plankton communities. J. Hazard. Mater. 2020, 385, 121625. [Google Scholar] [CrossRef]
- Qin, H.W.; Chen, L.F.; Liu, N.; Qin, W.C.; Yuan, X. Toxic effect of ofloxacin on Scenedesmus obliquus. Environ. Chem. 2011, 30, 885–886. (In Chinese) [Google Scholar]
- Su, Z.X.; Xiao, H.; Li, C. Toxic effects of enrofloxacin hydrochloride on Chlorella pyrenoidosa. Chin. J. Prev. Vet. Med. 2017, 6, 96–99. (In Chinese) [Google Scholar]
- Wan, J.J. Study the Response of Freshwater Microalgae to Several Antibiotics Stress. Ph.D. Thesis, Huaqiao University, Xiamen, China, 2014. [Google Scholar]
- Shi, L. The Hormesis Effect of Three Quinolone Antibiotics in Chlorella pyrenoidosa. Ph.D. Thesis, Harbin University of Commerce, Harbin, China, 2018. [Google Scholar]
- Lian, P.; Ge, L.Y.; Deng, H.H.; Zhao, C.R.; Xu, X.X. Toxic effects of two quinolone antibiotics on Platymonas subcordiformis. Environ. Sci. Manag. 2014, 39, 46–48. [Google Scholar]
- Hu, Z.Q.; Liu, Y.D.; Xiao, B. Effects of microcystin on the growth and photosynthetic activity of algae in freshwater. Ecol. Environ. 2008, 3, 885–890. [Google Scholar]
- Zhang, H. Effects of the Ratio of Nitrogen and Phosphorus Concentration on Growth and Microcystin of Microcystis aeruginosa. Ph.D. Thesis, Hefei University of Technology, Hefei, China, 2020. [Google Scholar]
- Oh, H.M.; Lee, S.J.; Jang, M.H.; Yoon, B.D. Microcystin production by Microcystis aeruginosa in a phosphorus-limited chemostat. Appl. Environ. Microbiol. 2000, 66, 176–179. [Google Scholar] [CrossRef]
- Wiedner, C.; Visser, P.M.; Fastner, J.; Metcalf, J.S.; Codd, G.A.; Mur, L.R. Effects of light on the microcystin content of Microcystis strain PCC 7806. Appl. Environ. Microbiol. 2003, 69, 1475–1481. [Google Scholar] [CrossRef]
- Watanabe, M.F.; Oishi, S. Effects of environmental factors on toxicity of a cyanobacterium (Microcystis aeruginosa) under culture conditions. Appl. Environ. Microbiol. 1985, 49, 1342–1344. [Google Scholar] [CrossRef]
- Wu, R.; Cui, L.F.; Lu, S.; Shi, Y. Effects of temperature and light on the growth of Microcystis aeruginosa and the release of algal toxins. Environ. Sci. Technol. 2010, 6, 33–36+51. [Google Scholar]
- Wu, H.Y.; Su, J.; Shi, W. Study on the growth and toxicity Microcystis aeruginosa strain under different condition. J. Environ. Health 2006, 23, 304–307. [Google Scholar]
- Yi, W.L.; Wang, G.D.; Liu, X.W.; Ma, Z.Y. Effects of N/P ratios on the growth and some biochemical constituents of Microcystis aeruginosa. J. Northwest Sci-Tech Univ. Agric. For. Nat. Sci. Ed. 2005, 6, 151–154. [Google Scholar]
- Xu, D.M. Study on Distribution Characteristics and Influence Mechanism of Microcystins in Chaohu Lake. Ph.D. Thesis, Anhui Jianzhu University, Hefei, China, 2022. [Google Scholar]
- Long, B.M.; Jones, G.J.; Orr, P.T. Cellular microcystin content in N-Limited Microcystis aeruginosa can be predicted from growth rate. Appl. Environ. Microbiol. 2001, 67, 278–283. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types of Antibiotics | Concentration (mg/L) | ||||
|---|---|---|---|---|---|
| Enrofloxacin (EF) | 0 | 12 | 36 | 72 | 120 |
| Ciprofloxacin (CF) | 0 | 30 | 45 | 60 | 150 |
| Types of Antibiotics | CK | Concentration (μg/L) | ||
|---|---|---|---|---|
| Enrofloxacin | 0.0 | 56.1 (1 × SC) | 280.5 (5 × SC) | 1402.5 (25 × SC) |
| Ciprofloxacin | 0.0 | 49.8 (1 × SC) | 249.0 (5 × SC) | 1245.0 (25 × SC) |
| Types of Antibiotics | Regression Equation | R2 |
|---|---|---|
| Enrofloxacin | y = 110.35 + (−104.96 − 110.35)/(1 + exp((x − 2.88)/56.47)) | 0.9983 |
| Ciprofloxacin | y = 110.35 + (−104.96 − 110.35)/(1 + exp((x − 2.88)/56.47)) | 0.9984 |
| Grade Toxic | Aquatic Organisms 96 h LC50 (mg/L) | Rat Oral 96 h LC50 (mg/L) |
|---|---|---|
| Grade X (very high toxic) | <0.1 | <0.5 |
| Grade A (high toxic) | 0.1–1.0 | 0.5–5.0 |
| Grade B (medium toxic) | 1.0–10.0 | 5.0–50.0 |
| Grade C (medium-low toxic) | 10.0–100.0 | 50.0–500.0 |
| Grade D (low toxic) | >100.0 | >500.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, Y.; Xie, Q.-M.; Chu, T.-J. Effects of Enrofloxacin and Ciprofloxacin on Growth and Toxin Production of Microcystis aeruginosa. Water 2023, 15, 3580. https://doi.org/10.3390/w15203580
Xia Y, Xie Q-M, Chu T-J. Effects of Enrofloxacin and Ciprofloxacin on Growth and Toxin Production of Microcystis aeruginosa. Water. 2023; 15(20):3580. https://doi.org/10.3390/w15203580
Chicago/Turabian StyleXia, Yu, Qin-Ming Xie, and Ta-Jen Chu. 2023. "Effects of Enrofloxacin and Ciprofloxacin on Growth and Toxin Production of Microcystis aeruginosa" Water 15, no. 20: 3580. https://doi.org/10.3390/w15203580
APA StyleXia, Y., Xie, Q.-M., & Chu, T.-J. (2023). Effects of Enrofloxacin and Ciprofloxacin on Growth and Toxin Production of Microcystis aeruginosa. Water, 15(20), 3580. https://doi.org/10.3390/w15203580