
Satellite DNA: An Evolving Topic

Departamento de Genética, Facultad de Ciencias, Universidad de Granada, 18071 Granada, Spain
Genes 2017, 8(9), 230; https://doi.org/10.3390/genes8090230
Submission received: 3 August 2017
/
Revised: 12 September 2017
/
Accepted: 13 September 2017
/
Published: 18 September 2017
(This article belongs to the Special Issue Chromosomal Evolution)
{kind=link}
{kind=link}
Abstract
Satellite DNA represents one of the most fascinating parts of the repetitive fraction of the eukaryotic genome. Since the discovery of highly repetitive tandem DNA in the 1960s, a lot of literature has extensively covered various topics related to the structure, organization, function, and evolution of such sequences. Today, with the advent of genomic tools, the study of satellite DNA has regained a great interest. Thus, Next-Generation Sequencing (NGS), together with high-throughput in silico analysis of the information contained in NGS reads, has revolutionized the analysis of the repetitive fraction of the eukaryotic genomes. The whole of the historical and current approaches to the topic gives us a broad view of the function and evolution of satellite DNA and its role in chromosomal evolution. Currently, we have extensive information on the molecular, chromosomal, biological, and population factors that affect the evolutionary fate of satellite DNA, knowledge that gives rise to a series of hypotheses that get on well with each other about the origin, spreading, and evolution of satellite DNA. In this paper, I review these hypotheses from a methodological, conceptual, and historical perspective and frame them in the context of chromosomal organization and evolution.
1. Introduction
Eukaryotic genomes are composed of a large amount of different classes of repetitive DNA sequences, either dispersed (mainly transposons and retrotransposons as well as retrotransposed sequences and some protein-coding gene families) or arranged in tandem (ribosomal RNA, protein-coding gene families, satellites, telomeric DNA, centromeric DNA) [1,2]. Repetitive DNA would explain the C-value enigma [3]. Thus, there is not a correlation between C-value (the DNA content) and the organism’s complexity. Differences in genome size between species, even closely related species, are at times thousands of orders of magnitude [4,5,6,7,8,9]. Among repetitive sequences, transposable elements (TEs) are mostly responsible for the pronounced differences between genomes. Thus, for example, TEs represent the 45% of the human genome [10], the 52% of the opossum genome [11], or the 85% of the maize genome [12]. However, in addition to TEs, satellite DNA (satDNA) also contributes greatly to genomes. Satellite DNA families are organized in large tandem arrays of highly repetitive non-coding short sequences.
There are several considerations to do concerning the distribution of satDNA families among species and their contribution to the C-value:
(i) Different satDNA families may be present in one species. For example, there are up to 15 families in Pisum sativum [13], 62 families in Locusta migratoria [14], or 9 satDNA families within the human genome [15,16]. However, there are usually one or a few predominant satDNA families in each species [13,14,15,16,17,18,19]. For example, the most abundant satDNA family within the human genome, the centromeric α satDNA, represents more than half of the total content of satDNA in the human genome [10,15,16]. On the other hand, the number of satDNA families must be taken relatively at times, as some of these satDNA families might be related to each other or on the contrary comprise different subfamilies of the same satDNA family.
(ii) Related species may share a set or a library of satDNA families. As mentioned in the previous point, among the families that make up the library is typical for each species to have one or a few predominant satDNA families. For example, different satDNA families are shared by the genomes of different species of the beetles of the family Tenebrionidae, each species having one or two predominant major satDNA families [17,18,19], as occur for the major satDNA of Tribolium castaneum (TCAST1) [20,21].
(iii) Concerning a particular satDNA family, this may be species-specific, which is in accord with the accepted view that satDNA is the evolutionarily fastest part of the genome (see below). However, in addition to species-specific satDNAs [22,23], there are also satDNA families shared by several related species within a genus [24,25,26,27,28], by the whole group of species composing a genus [23], by several genera from a family [29], by a whole family [30,31,32,33], by several families [34], or by a whole order [35,36,37,38]. Furthermore, a satDNA family might be preserved in various taxa of a phylum [39,40].
(iv) SatDNAs can represent a large proportion of the genome of one eukaryotic species, but not always. In plants, for example, satDNA can represent between 0.1% and 36% of the genome [41,42,43,44,45,46,47]. There is also a wide range of variation in satDNA content within the animal kingdom with values differing between less than 0.5% and more than 50%, both in invertebrates and vertebrates [15,16,17,18,19,20,21,48,49,50,51]. In both cases, plants and animals, the differences in satDNA content might significantly contribute to considerable genome-size differences between related species or even between cytotypes of the same species [9,44,45,47,51]. At times, these differences between species are due to the differential amplification of one particular satDNA while other times the differences are mediated by the sum of all satDNA families from each species. These are two examples among many of them: (a) the genomic content of the FriSAT1 satDNA varies greatly (0.1–36%) between different species of the plant genus Fritillaria [45]; (b) it is estimated that Drosophila simulans has 5% of the genome composed of satDNA but only the 0.5% of the Drosophila erecta genome is satDNA (reviewed in [9]). However, there are species such as Drosophila melanogaster with over 20% of the genome or Drosophila virilis with nearly 50% of the genome composed of satDNA [9].
Undoubtedly, the data that we have about the proportion of satDNAs in the eukaryotic genomes are fragmentary. Firstly, the data on satDNA genomic content was many times based only on rough estimations and, most times, the total abundance of satDNA in most eukaryotic genomes was probably higher than the estimated one. For decades, rough estimations of satDNA content within genomes have been used by indirect quantification methods (see below). Secondly, until a few years ago, satDNAs were underrepresented in genomic projects (see below). Further, the main procedure to identify and isolate satDNA families from the eukaryotic genomes during the last four decades (genomic restriction digestion and electrophoresis yielding a ladder pattern) is a biased method for the identification of usually one or, sometimes, a few satDNA families of a given genome but not to the detection of the complete set of satDNA families from one species. Thus, for example, in addition to satDNA families identified by conventional methods, several other satDNAs were identified after a genomic approach in several species [9,13,52].
Since its discovery more than 50 years ago, satDNA is still today one of the most intriguing, but also fascinating, parts of the genome. However, things are changing because the methodology in the analysis and the knowledge of genome are also changing. The classic review by Maxine F. Singer [53] put the accent on a new way of studying satDNA, which was a starter point for many researchers who opened up the ways for the decipherment of satDNA. Singer emphasized the conception of how methods change through time and how these changes lead to changing concepts. Today, after 35 years, new methods of genomic analysis are also provoking conceptual changes on the topic of satDNA. This review tries to summarize all the advances in the knowledge of satDNA, its organization, its function, and its evolution. Today, we have a better perspective on the knowledge of these aspects. The review focuses on the approach to the organization, to the function and to the evolution of satDNA also from this perspective of the evolution of all we have been learning during the years.
2. Changing Methods
The existence of repetitive DNAs in the genomes of eukaryotes was first unveiled in 1961 by Kit et al. [54] and by Seuoka et al. [55]. These pioneering works revealed that genomic DNA from mouse exhibits two buoyant DNA bands in density-gradient ultracentrifugation of DNA using cesium chloride. One of these two bands, the minor component, representing about 10% of the genome, was called satellite DNA. The DNA of this band underwent higher rates of renaturation than the rest of the nuclear mouse DNA or than the simplest genomes composed mostly of unique sequences, a proof that the satellite peak of the mouse genome was composed of repeated nucleotide sequences [56,57,58,59]. Britten and colleagues [56,57,58,59] developed the Cot analysis, based on the principles of DNA renaturation kinetics according to which the DNA sequences re-associates at a rate that is directly proportional to the number of times it occurs in the genome. They demonstrated that a variable proportion of every eukaryotic genomic DNA is composed of repetitive elements and that eukaryote genome sequences can be divided into highly repetitive, moderately repetitive, or single-copy classes of DNA sequences according to their reiteration frequency. Cot analysis thus represented a powerful tool by which highly repetitive, moderately repetitive and single/low-copy DNA can be selectively and efficiently fractionated, cloned, and characterized determining their complexity, composition and abundance [60].
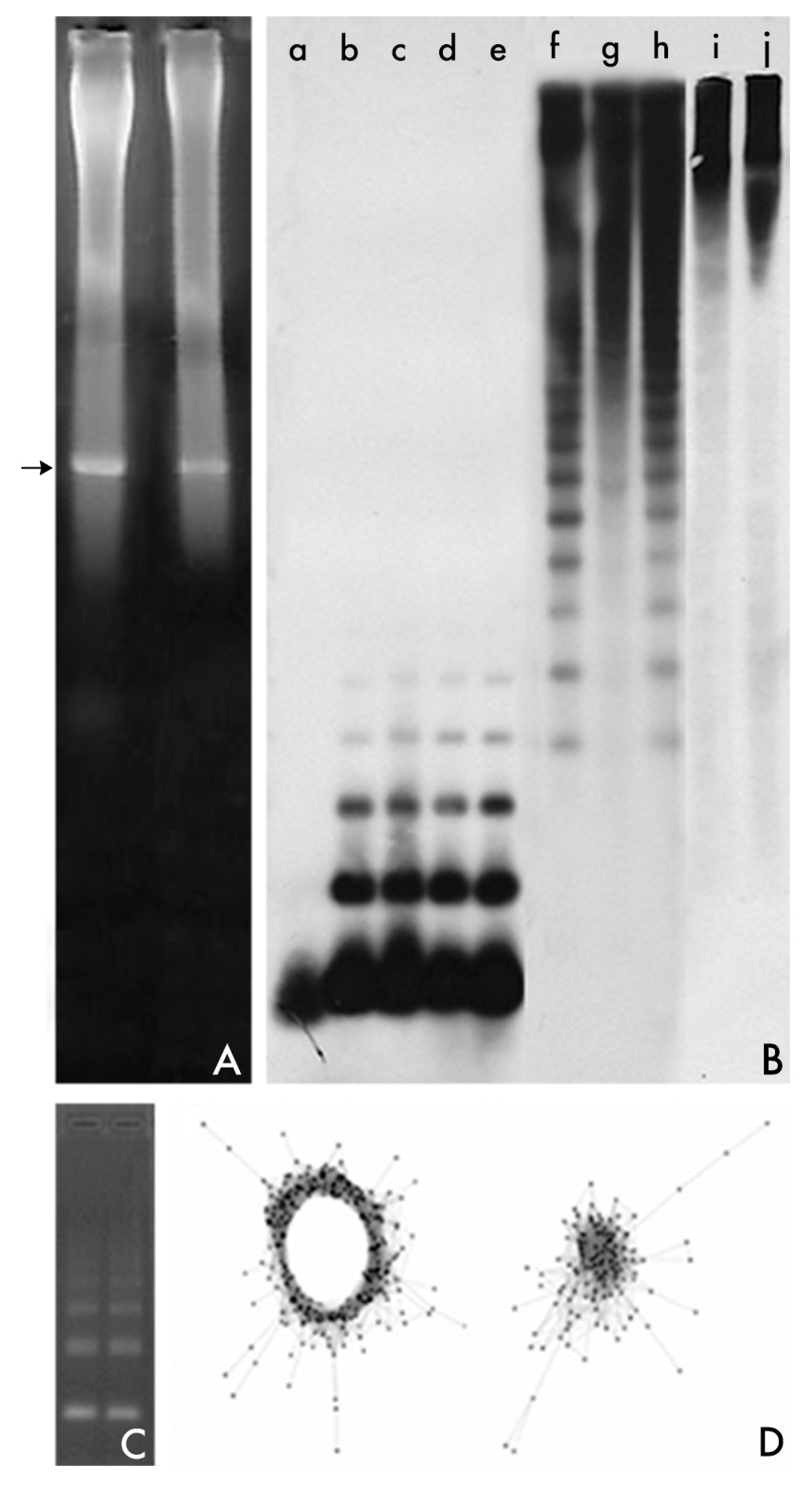
These two methods dominated analysis of repeated DNA sequences during the 1960s and 1970s [53]. These methods were substituted by the isolation from restriction endonuclease treatment of genomic DNA. The method simplified and popularized the study of satDNA, assisted to separate satDNA families from other repetitive sequences, and aided in uncovering cryptic satDNAs [53]. Following digestion of genomic DNA with a restriction enzyme and electrophoresis of the DNA fragments generated on agarose gels, satDNA sequences are revealed as a prominent band against the background smear [53]. In this way, the individual members of a repeated set of sequences are available for cloning after the prominent band is excised from the agarose gel, the agarose slice is melted and the DNA purified (Figure 1). These DNA fragments isolated from agarose gels have been found to be the source of numerous studies of satDNA families from a large number of eukaryotic species.
Roughly, quantification and global genomic organization may be addressed by using dot-blot and Southern blot hybridization techniques (Figure 1). In dot-blot analyses, defined amounts of total genomic DNA as well as the defined amounts of the unlabeled probe (repeat units of the satellite DNA family) are denatured and immobilized on nylon and hybridized with the labeled probe. The relative amounts of the satDNA family in the genome are estimated then by comparisons between densitometric scans of hybridization signals in genomic DNAs and those obtained for the reconstruction standards. The dot-blot hybridization was also commonly used for the detection of satDNA families of one species within the genomes of related species, a method that even permitted the use of this technique in an applied way for the establishment of phylogenetic relationships by the cladistic association of species in monophytletic groups. The organization of a repetitive DNA family in a genome may be analyzed by Southern blot hybridization (Figure 1). Three typical patterns can be observed after hybridization [53,61]. Southern blot hybridization patterns give a rough indication of sequence variation within and between species and contain certain useful phylogenetic signals [62].
In addition to Southern blot hybridization, the developmet of in situ techniques of hybridization was a breakthrough in the characterization of a satDNA (Figure 2). First attempts using radiolabeled probes were promising [63] and revealed that satellite sequences were located in the heterochromatin, in this case, the centromeric heterochromatin of mouse chromosomes. Notwithstanding, the real revolution, popularization, and potential of this technique came from the use of nonradioactive methods [64] and above all with the advent of fluorescent in situ hybridization (FISH), a powerful tool that may combine the use of different labeled probes with different fluorochromes [64,65].
The sequence of a satDNA family can be obtained by the entire set of monomeric units purified from melted excised agarose gels. Unambiguous consensus sequences are frequently obtained with uncloned sets [53,66,67]. Although this method allowed determining a rough estimate of satDNA variation [66,67], the sequencing of cloned members of the set has been the common procedure during the last four decades. The advent and the advantages of the polymerase chain reaction (PCR) technique extended its use for the isolation of repeats of a satDNA family from one species or from several related species, both by the selection of one primers pair or the combined use of several primers pair in order to uncover the whole variant type sequences found in each genome [68]. Sequencing of repeats of satDNA families obtained by both cloning and PCR, combined with Southern blot and in situ hybridization, has provided a wealth of information about the organization and location of satDNA repeats, the length and copy number of satDNA repeats, the satDNA repeat variability, the functional role of satDNA sequences, and the satDNA evolution as well as its (moderate) use in phylogenetic analysis and in taxonomic studies [1,2,41,69,70,71,72].
Meanwhile, over decades, satDNA sequences were left out of the great genome projects since difficulties arose in the assembly of contigs containing repeat sequences. It is obvious that a genomic perspective in the isolation and analysis of satDNA repeats would overcome the bias of the sequence data obtained by cloning or PCR methods. Thus, for example, there are many monomers that escape to cloning when they are isolated from prominent bands, which were excised from agarose gels, containing the monomer subset obtained after complete digestion. In addition, this procedure omits the cloning of the multimers resulted from undigested repeats lacking the site for the restriction enzyme utilized. This bias in the isolation procedure would be maintained in subsequent PCR experiments which use primers designed from information gathered by the aforementioned procedure. Furthermore, there are added difficulties when a satDNA family is poorly represented in a genome. Obstacles also arise when one is dealing with species having high genome size or with small quantities of satDNA. Further, one may raise the question of how many satDNA families are present within a genome. Some of these families may be overlooked in a routine examination conducted using restriction enzymes for their identification. In this context, the integration of Cot analysis, DNA cloning, and high-throughput sequencing was proposed as an attractive methodology that facilitates genome characterization [60].
Thus, satDNA analysis has found an ally in high-throughput sequencing of genomes using Next-Generation Sequencing (NGS). NGS and high-throughput in silico analysis of the information contained in NGS reads have transformed the study of repetitive DNA [41,73]. An efficient pipeline called RepeatExplorer [74,75] has been developed which allows for the de novo identification of repetitive DNA families in species lacking a reference genome [13,73,74,75,76]. RepeatExplorer follows a similarity-based read clustering approach that allows detection of repetitive sequences, which are identified as groups of frequently overlapping sequence reads in all-to-all read comparisons [13,73,74,75,76]. The clustering procedure employs graph-based methods that transform read similarities to a virtual graph, where reads are represented as nodes and their similarities by edges connecting the nodes. The identification of communities of densely connected nodes allows for the identification of various families of repetitive DNA sequences (Figure 1). The reads within the sequence clusters can be assembled to generate contigs that represent the repeats they contain [13,73,74,75,76]. The combination of NGS and computer analysis favours an in-depth global genomic analysis on the repetitive content of genomes and gives us the opportunity to uncover satDNA families whose isolation was elusive by other methods [8,13,14,43,52,77,78,79,80]. In a further step in the use of RepeatExplorer, Ruiz-Ruano et al. [14] have implemented a bioinformatic toolkit (satMiner) which allows for the identification of satDNA families that are extremely rare in the genome. This pipeline consists of several rounds of RepeatExplorer clustering separated by filtering out the reads containing already known satellites, thus increasing the likelihood of finding new rare satellite families. The method is highly reliable for species with high genome size or with small quantities of satDNA. In a further improvement of RepeatExplorer, Novak et al. [81] have developed Tandem Repeat Analyzer (TAREAN), a computational pipeline for unsupervised identification of satellite repeats from unassembled sequence reads. The pipeline uses low-pass whole genome sequence reads and performs their graph-based clustering. Resulting clusters, representing all types of repeats, are then examined for the presence of circular structures characteristic for tandem repeats. Reads from these clusters are then decomposed to k-mers and fractions of the most frequent k-mers are used for reconstructing representative monomer sequences for each satellite repeat [81].
RepeatExplorer, satMiner, and TAREAN are being extensively used in the analysis of the repetitive DNA content of many plant and some animal species [14,73,82]. The development of all these computer tools is opening new opportunities to uncover the core details of satDNA evolution and to gain insights on the different repeat families making up a given genome, their relative abundance and variability, as well as their roles in different genetic and genomic processes [74,75]. Further, this new perspective greatly contributes to the development of comparative genomics and of phylogenomics [74,75]. However, the graph-based genomics approaches are not the only ones. Wei et al. [9] have developed a method called k-Seek that analyzes unassembled Illumina sequence reads for identify and quantify short tandemly repeating sequences (kmers) of 2–10 bp, repeat lengths that are usual among satDNAs in Drosophila. While the existence of a reference genome is not needed for the use of RepeatExplorer, satMiner, or TAREAN pipelines, or for the use of k-Seek, the existence of a reference genome has facilitated the development of other computer programs for the analysis of the satDNA content of model species such as Caenorhabditis elegans or Tribolium castaneum. In fact, there are up to 25 conventional programs designed to retrieve tandem repeats (usually, short tandem repeats) from complete or or nearly complete genomes, which were not intended for processing the billions of short reads generated by Illumina or 454 sequencing in an operative time [83,84]. Even so, Subirana and Messeguer [48] have recently developed SATFIND and used it for the identification and analysis of the satellite families in Caenorhabditis [49]. Also, Pavlek et al. [85] have recovered the use of the Tandem Repeat Finder (TRF) algorithm [86] through the Tandem Repeats Database (TRDB) [87] for the identification of satDNA families of Tribolium castaneum.
3. Changing Concepts
The use of the term “satellite DNA” to define highly repetitive DNA sequences organized in tandem arrays was first proposed by Pech et al. [88] and coined and popularized as such by Singer [53], regardless of whether or not these sequences form classical satellite peaks upon isopycnic centrifugation [53]. The term covers a wide diversity of sequences that represent a highly variable part of the eukaryotic genome. SatDNA families differ from each other by their location, their repeat unit length, their abundance, and their nucleotide sequence. Together with satDNA evolution and function, those issues will be reviewed in the following subsections attempting to display how concepts about the satDNA topic have been changing during last decades.
3.1. Nucleotide Sequence Composition and Repeat Organization
In theory, satDNA repeats could be generated from any random sequence. Several satDNA families found in one genome may be related in origin. In these cases, the repeats forming each current different satDNA family were originated from the same shorter sequence [14,25]. However, most satDNA families found within the genome of a species are unrelated in sequence. As a matter of fact, it has been found that current repeats of a satDNA family might be the result of a complex evolutionary process that has led to the current repeat unit from shorter repetitions. Thus, repeat sequences are often composed of direct sub-repeats or motifs that remain as remnants of past events of sequence duplications. Thus, for example, the centromeric satDNAs of tilapias, sparids, or mice are the result of several cycles of duplication and sequence divergence of a basic shorter monomer composed of nine nucleotides [30]. A complex scenario of duplications and expansions was observed in the formation of the HindIII satDNA of sturgeons [89] or in the case of the centromeric satDNA of the wedge sole [90]. Current repeats originated from shorter repeat units were also found in several plant species [25,47,91,92].
Further, monomer sequences of some satDNAs might form higher-order repeat (HOR) units. HORs are the result of the simultaneous amplification and homogenization of two or more adjacent monomers [70]. Alpha satellite DNA is the most abundant satDNA of primate centromeres, at least of the simian primates [38]. The alpha satellite of humans adopts two different organizations. One is in the form of monomeric tandem repeats of about 170 bp. The other is organized into HOR structures that consist of multiple (from 2 to 34) head-to-tail basic repeat units [71,93,94,95]. Nucleotide sequence identities vary between 70% and 90% when comparing monomeric repeats of the same multiple unit. However, the similarity is higher than 95% when a monomeric unit of one multiple unit is compared to a counterpart monomer (located at the same position) in another multiple unit [38,93,94]. HOR organization is found at the centromere of human chromosomes while the pericentromeric heterochromatin is composed of single alpha satDNA monomers (50–100% sequence identity), which can coexist with HORs [70,71,96,97,98]. Different chromosome-specific alpha-satellite subfamilies have been described in humans, characterized by HOR organization plus their related monomer sequences [93,94,99,100]. HOR organization was believed to be restricted to hominids but it has been demonstrated that this type of organization is extended to hominoids [101,102,103] and further to the Old World monkeys (Catarrhini) [98,102,103,104,105] and also to the New World monkeys (Platyrrhini) [38,106]. In all characterized species, the monomeric units have a length of 170 bp but during the course of primate evolution this fundamental seeding unit has experienced a number of sequence and structural variations [70]. Old and New World monkeys and gibbons lack chromosome-specific subfamilies [70,101,105].
Regular HORs similar to those found in humans, but usually dimeric, have been found in several species of beetles (reviewed in [107,108]). However, the formation of complex HORs, shaped from interspersed and/or inversely oriented monomers and frequently with extraneous sequence elements, is usual (reviewed in [107,108]). Complex HORs have been found in non-human mammals, such as mouse, swine, bovids, horse, dog, and elephant and in insects (reviewed in [107,108]).
Interestingly, several classes of repetitive DNA sequences can be the seed for the formation of a satDNA family. For example, some satDNA families originated from part of the intergenic spacer of ribosomal RNA genes (rDNA) [41,70] or from parts of different classes of TEs [109]. Conversely, tandem repeats have also been found as integral components of TEs [109] and some satDNA were probably derived from tandem repeats present in a hypothetical miniature inverted–repeat transposable element (MITE)-like element [109,110,111,112]. Thus, it has been suggested that mobile elements may be an important source of satDNAs in diverse genomes [109,113,114,115].
3.2. Defining satellite DNA
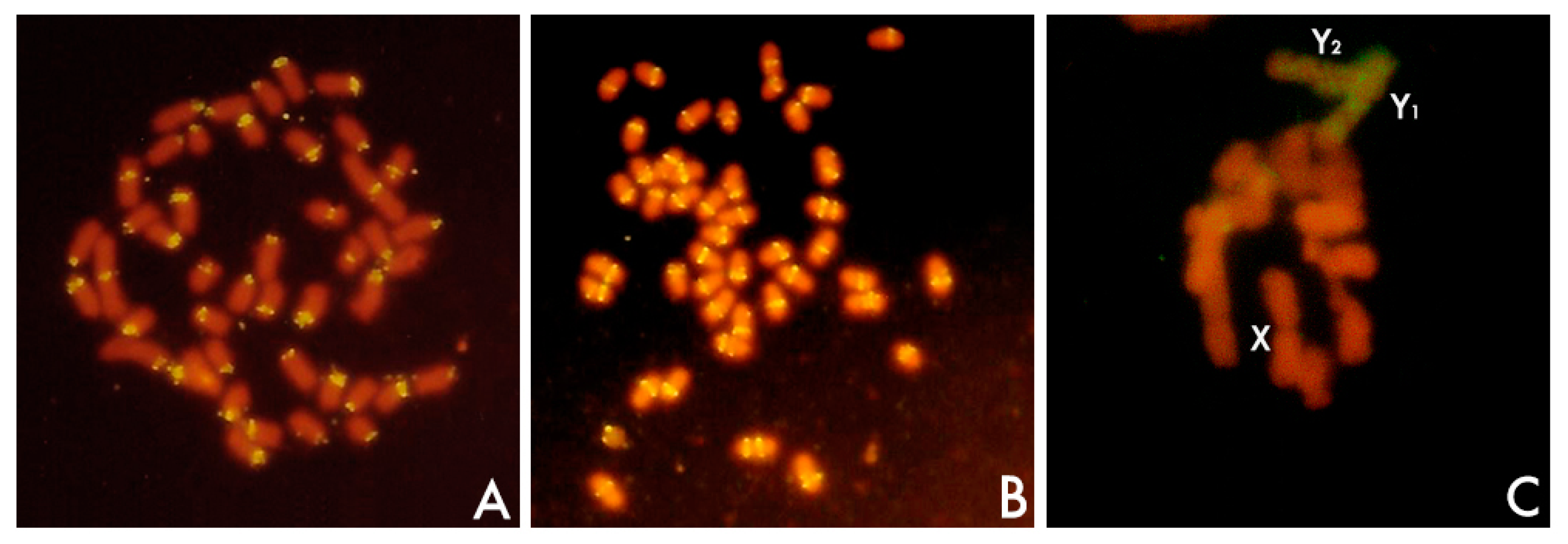
Although location, repeat length, and copy number are features than can be conveniently analyzed separately, I want to connect all of them in the following paragraphs because the variability shown by satDNAs concerning these features is connected also with the definition of satDNA itself. SatDNAs are the main component of the heterochromatin, which is found specifically at pericentromeric and subtelomeric locations of the chromosomes (Figure 2) [1,41,69,70,71]. Additionally, heterochromatin might be found occupying interstitial loci of chromosomes in specific positions of the chromosome arms between the subtelomeric and the pericentromeric regions, especially in invertebrates and plants. Further, heterochromatin might also be found in specific chromosomes such as sex chromosomes (Figure 2), supernumerary chromosomes, or in particular regions of one particular chromosome (see below). SatDNA is also a widespread element nucleating centromeres. However, in addition to satDNA, heterochromatin and centromeres might be occupied also by TEs [41,42,71,109,116,117,118,119,120,121]. Further, simple sequence repeats (SSRs) (also known as single tandem repeats -STRs- or microsatellites) as well as minisatellites are commonly found forming the pericentromeric and the subtelomeric heterochromatin [122,123,124].
Microsatellites are defined classically as tandem repeats of less than 10 bp in length in arrays less than 1 Kb, distributed in loci scattered throughout the genome. Minisatellites are similar but characterized by longer repeats (>10 bp). Most animal and plant satDNA sequences commonly have monomer unit lengths of about 150–180 bp or 300–360 bp, although exceptions to this assumption are far from being exceptional [35,39,41,70]. Even in the event that the repeat length intervals between satDNAs and microsatellites (≤10 bp) and minisatellites (10–100 bp) sometimes overlap, they have been conventionally differentiated by their location. Thus, while satDNAs would be composed of families of tandem repeats located as long arrays at the heterochromatin, micro-, and minisatellites would be proper of euchromatic regions. It has been claimed that simple repeats, when present and organized in arrays of many thousand copies in heterochromatin, they are satDNAs [41]. In fact, today, the term satDNA is applied to any tandem of hundreds to thousands repeat units located in the constitutive heterochromatin. Thus, in a broad sense, megabase-long arrays of repeating units of only few bp should be considered also satDNA [41,125]. Indeed, repeat lengths of ‘classical’ satDNAs vary between a few and several thousand base pairs. Thus, while repeat lengths of some satDNAs are longer than one or two Kb in cetaceans and bovids [35,51] or 5.9 Kb in potato [92], several satDNA of the pericentromeric heterochromatin of human chromosomes are composed of repeats as short or even shorter than the repeat length of common microsatellites [126]. In Drosophila, with the exception of three satDNAs, most known satellites are tandem repeats of simple sequences (≤10 bp) [9].
Furthermore, recently it has been demonstrated the existence in the euchromatin of loci composed of tandems with repeats of lengths in no way within the range of the micro- or minisatellite classification. Thus, short arrays of satDNAs dispersed along the euchromatin have been described. There are also satDNAs present not exclusively within heterochromatin but also dispersed as single repeats or short arrays within euchromatin [14,21,80,85,127,128,129,130,131]. Similarly, shorter repeats showed the same capability to cluster both in euchromatin and in heterochromatin. In fact, Ruiz-Ruano et al. [14] demonstrated that microsatellites, minisatellites, and satellites show similarities at genomic and cytological levels, with intragenomic dissemination preceding clustering, and claimed that the fact that all kinds of satDNA (micro-, mini-, and satellites) can show non-clustered and clustered states suggests that all these elements are mostly similar, except for repeat length. Also, variable number of tandem repeats (VNTRs) in bacteria showed similar features to non-clustered satDNAs [14].
These new views on the concept of satDNA became possible thanks to the evolving methodology used in the approach of satDNA analysis. These new perspectives lead to the coining of new terms such as those that arise from the high-throughput analysis of genomes. Thus, Ruiz-Ruano et al. have proposed the term ‘satellitome’ for the whole collection of different satDNA families in a genome [14]. The computer analysis of NGS reads allows us to obtain new insights on the origin and evolution of the satellitome, a part of the ‘repeatome’, the term proposed by Kim et al. [82] for the collective set of the repetitive elements of a genome (TEs, tandem repeats, etc.). Inter- and intraspecific satellitome comparative analysis provides information on satDNA organization and evolution as well as chromosome organization and evolution [14].
3.3. Principle of the Equilocal Distribution of Heterochromatin
The principle of equilocality of heterochromatin distribution would explain the tendency of heterochromatin to occupy similar location on non-homologous chromosomes [132]. The principle affect to all different sites of heterochromatin—pericentric, interstitial, and subtelomeric—which tend to have different cytogenetic properties [132]. That is, according to this principle, the heterochromatin accumulates at equivalent positions in each chromosome within a genome: pericentromeric and subtelomeric regions, principally. When heterochromatin is built up in an interstiticial region between the centromere and the telomere of a chromosome arm, all of the members of the chromosome set, or a particular subset of chromosomes, may carry heterochromatin at comparable locations [132]. The concept applies to satDNA and assumes that different satDNAs families occupy different equilocal sites. In fact, even in one species with many satDNA families such as Locusta migratoria, different satDNAs families displayed similar equilocal distribution across non-homologous chromosomes supporting the principle of satDNA equilocality [14]. Thus, usually, the pericentromeric heterochromatin of one species is composed of a particular satDNA family which is different from the satDNA family found in the subtelomeric region. When several satDNA families are located in one given chromosomal region, pericentromerically for example, they are arranged in differentially located arrays and this arrangement is maintained in all the chromosomes, as occur in mouse minor and major satDNA families ([133] and references herein).
The concept of equilocality assumes that subtelomeric regions are presumptive sites where heterochromatin amplification tends to be initiated, and from which satDNA sequences are spread to interstitial sites in a process known as interstitialization, which may be facilitated by telomere reunion at the first meiotic prophase bouquet [132,134]. Interstitialization has ocurred in different species through the transference of satDNA sequences from the telomeres towards interstitial sites [135]. The subtelomeric pSc119.2 satellite DNA tended to spread towards new interstitial sites during the diversification of the most primitive form of the genus Secale towards the most advanced taxa [135]. This highly repetitive sequence has supported events of interstitialization also in Hordeum [122]. DraI is a subtelomeric satDNA family shared by several fish species of the Sparidae family (Figure 2). In addition to subtelomeric location, there are several interstitial loci in some chromosomes of these species [29]. However, the process of interstitialization cannot explain alone the formation of new satDNA families in interstitial loci (see below).
There are clear exceptions to the principle of equilocality. Non-equilocal arrangement of a satDNA family could be due to chromosome reorganization such as inversions and/or transpositions or Robertsonian translocations [27].
Absence of exchange between non-homologous chromosomes may lead to the independent evolution of satDNAs in two subgenomes, even at equilocal heterocromatic sites, and to the emergence of different satDNA families, as occur in Sus scrofa domestica. This species has a bimodal karyotype 12 meta-/submetacentric (Mc) and 6 acrocentric (Ac) chromosomes (Mc and Ac ‘subgenomes’). The centromeres of each ‘subgenome’ differ by having a different satDNA family. This difference was explained by the spatial arrangement of chromosomes in meiotic pachytene nuclei [136].
On the other hand, an exceptional case of non-equilocal distribution of satDNA is found in the larger chromosome of the plant species Muscari comosum which is the result of a massive amplification of the Muscari comosum satDNA family (MCSAT), constitutes the 5% of the genome, and leads to the characteristic asymmetry of the karyotype of this species [46]. Additionally, the MCSAT repeat family varies between 0.8% and 5% of the genome between species of the subgenus Leopoldia of Muscari, which largely contributes to the different degrees of asymmetry of their karyotypes [46].
Alteration of the equilocal distribution of satDNAs also included satDNA amplifications in specific regions of particular chromosomes such as B and sex chromosomes as well as microchromosomes. Sex chromosomes have arisen independently several times in several different groups of organisms from a pair of autosomes. Genetic differentiation of undifferentiated non-heteromorphic sex chromosomes leads to the establishment of sex chromosome heteromorphism [137]. Thus, the gradual suppression of recombination between the sex chromosomes is thought to lead to their progressive divergence and to the erosion of the Y chromosome [138] which results in the loss of function of many genes within the Y chromosome and accumulation of distinct classes of repetitive DNAs such as TEs and satDNAs [137,138,139,140,141,142,143]. The human male-specific region of the Y chromosome (MSY) is a mosaic of heterochromatic sequences and three classes of euchromatic sequences [140]. Among the repetitive sequences, these authors found that the heterochromatin of MSY encompasses at least six distinct satDNA sequences. In Orthoptera, the X0/XX sex-determining system is considered modal but eventually, diverse sex chromosome systems involving neo-Y formation evolved several times [144]. In one of these species, Eneoptera surinamensis, a high-throughput analysis of the satellitome has revealed that the neo-Y chromosome harbors the highest diversity of satDNAs documented to date, representing 39 distinct families, with seven being exclusive to this chromosome [144]. Several satDNA families were also deeply analyzed in dioecious plant species. Among flowering plants, only a few dioecious plant species have chromosome-mediated sex determination systems [141,142,145]. The most common case is the existence of XX/XY chromosomal complements although there are other alternatives involving more complex chromosomal systems. Sex chromosomes in plants are in general evolutionarily young existing a great variety of situations from non-heteromorphic sex chromosomes to highly differentiated heteromorphic sex chromosomes through several cases of different intermediate stages or degrees of sex-chromosome differentiation [41,141,142,145]. The Y chromosomes of XX/XY1Y2 species of the genus Rumex, but not those of XX/XY species, are heterochromatic and have accumulated a set of diverse repetitive sequences [25,52,146,147,148,149,150,151]. TEs were found in Y chromosomes of Rumex acetosa but satDNA prevented their significant expansion [52]. Kejnovský et al. [152] suggested that microsatellites are probably targets for insertions of transposable elements [152]. Microsatellite accumulation on the Y chromosome in Silene latifolia also suggested that the spread of microsatellites predates other structural changes that occurred during the Y-chromosome evolution [153]. SatDNA accumulation is occurring even in an early stage of sex-chromosome evolution such as that of Silene latifolia [154]. A high-throughput sequence analysis of repetitive DNA revealed generally low divergence in repeat composition between the sex chromosomes in S. latifolia, as expected according their relatively recent origin [155]. A comprehensive review on the impact of repetitive DNA sequences on plant sex chromosomes [156] and various papers on the evolution of sex chromosomes in different systems are treated in this volume and I refer to these articles for a broad view of the subject.
Chromosome-specific satDNAs are also found in particular chromosomes such as B chromosomes [157]. Supernumerary chromosomes or B chromosomes are dispensable genetic material found in about 15% of eukaryote organisms, both plants and animals [158]. They are derived from the A chromosome complement from the same (intraspecific origin) or a different (interspecific origin) species [158]. Interspecific origin may be a byproduct of interspecific hybridization [159] or a product of interspecies introgression [14,158,160,161]. Satellitome analysis has been revealed as a powerful tool to shed light on B chromosome origin and evolution in several cases. Thus, for example, the B chromosome in the grasshopper Eumigus monticola most likely arose from the proximal third of one autosome through a breakpoint that it could be delimited by the authors, as deduced from satellitome analysis [161]. In addition, two specific satDNAs in the B chromosome could be originated intra-specifically since seeds of smaller satDNA repeats probably involved in their formation were already present in the B-lacking genome, suggesting their massive amplification in the B chromosome after the origin of the supernumerary chromosome [161]. Similarly, A and B chromosomes of Secale cereale contained a similar proportion of repeats but differed significantly in composition by an additional massive accumulation of B-specific satDNAs [157]. Some B-enriched sequences are unique to the supernumerary chromosome, but not all of them, which suggest that B originated from A chromosomes [157]. Rye satDNA families including those clustered on the B chromosome are transcriptionally active. These B transcripts could have a function as scaffold RNA in the organization and regulation of Bs [162]. Indeed, B chromosomes, though initially inert elements, possess transcribed sequences and even transcribed pseudogenes and protein coding genes [162,163,164,165,166]. For a review of this topic, see the paper of Ruban et al. [167] in this volume. The same as B chromosomes, supernumerary chromosome segments have also been described, some of them being heterochromatic and plenty of satDNA sequences [168,169,170], but neither all supernumerary chromosomes nor all supernumerary chromosome segments are heterochromatic [41].
Bimodal karytoypes composed of macro- and microchromosomes are frequent in birds and reptiles. In some cases, there are microchromosome-specific satDNAs suggesting an independent evolution of these chromosomes with respect to the macrochromosomes [171]. For example, centromeric repetitive sequences have been isolated in struthioniformes, galliformes, and one species of turtle ([171] and references therein). In contrast, Matsubara et al. [171] analyzed three satDNA families in two species of snakes—the habu snake (Protobothrops flavoviridis, Viperidae) and the Burmese python (Python bivittatus, Pythonidae)—and either satDNA sequences showed no genomic compartmentalization between the macrochromosomes and microchromosomes as was found previously in lizards and skinks ([171] and references therein). On the basis of chromosome number, the sturgeons can be divided into two groups: the first includes those with a diploid number of 120 and the second includes those with a diploid number of about 240–260. Additionally, their karyotypes are characterized by the presence of numerous microchromosomes. The HindIII satDNA family is located in the centromeres of a reduced set of chromosomes which correlates with species ploidy [172]. Thus, diploid species have 8 HindIII centromeres while tetraploids have from 50 to 80 HindIII centromeres. In both cases, there was no distinction between micro- and macrochormosomes [172], but the results suggest the existence of other satDNA families or variants of the HindIII family populating the rest of centromeres. The DBC-150 satDNA family is restricted to a single pair of microchromosomes in species from the Drosophila buzzatii cluster [173].
3.4. Satellite DNA Evolution
According to the hypothesis developed by Ruiz-Ruano et al. [14], the birth of a satDNA family implies the de novo duplication of a genomic sequence of two or more base pairs that gives rise to a short tandem repeat at a unique genomic location. The formation of this locus can occur by various mechanisms such as, for example, the slippage of one DNA strand during DNA replication or the reinsertion of replicate copies from extrachromosomal circular DNA intermediaries. In a second step, this repetitive family can be disseminated throughout the genome by mechanisms such as transposition or reinsertion of replicate sequences from extrachromosomal DNA intermediaries. At that point, some loci may be amplified locally. Genomic restrictions and natural selection would henceforth place rigid limits on the accumulation of satellite DNA, especially in prokaryotes. In eukaryotes, however, some loci can be amplified locally and exceed the detection threshold of cytogenetic techniques (such as fluorescence in situ hybridization -FISH-) thus becoming a satDNA locus visible by FISH. Local amplification involves a rapid increase in the size of the tandem that could take place, for example, by unequal crossing-over. On the basis of this model, tandem repeats of 15 bp or less may appear by chance in many places in the genome in such a way that both microsatellites and shorter minisatellites can begin their life cycle in the second stage. These shorter repeat units can in turn give rise to longer repeat units through several consecutive cycles of duplication and divergence [25,30,89,90]. Similarly, HORs are evolutionary units of amplification and homogenization of smaller monomers [70].
3.4.1. The “Library” Hypothesis
The hypothesis about the origin of satDNA families of Ruiz-Ruano et al. [14], integrates the “library” hypothesis on the expansion of different satDNA families in different lineages [14,17,70,109,174,175]. Related species may share an ancestral set of different conserved satDNA families, each of which may be differentially amplified in each species. When a satDNA family is amplified differentially in one species, low-copy counterparts of it are found in other related species. Monomer variants comparisons of this satDNA family show high interspecific sequence conservation and absence of species-diagnostic mutations, as found in the beetle Palorus [17]. This hypothesis has been proven in insects [17,18,175,176] and plants [26,177,178].
Historically, both transposition and replication of extrachromosomal circles of tandem repeats by the rolling-circle mechanism and reinsertion of replicated arrays [66,67] have been postulated as the main mechanisms for the spreading of satDNA families. However, there was no evidence of such mechanisms as driving forces of satDNA dissemination. Cohen et al. [179,180] found evidence for the rolling-circle mechanism. However, it was not until very recently when the structural and functional liaisons between transposable elements and satDNAs have been found [109]. The data would probe that TEs may act as a substrate for satDNA emergence and mobility [109,114,115,181,182]. Mobile elements may significantly contribute to satDNA evolution by generating a library of tandem repeats that can be dispersed through the genome and in some cases amplified into long arrays of new satDNAs [109]. Šatović and Plohl [114] proposed that onset and spread of tandem repeats can be intimately linked to processes of transposition in more cases than expected previoulsy. Pavlek et al. [85] found that, in addition to other mechanisms, transposition might play an important role in the efficient spread of satDNA in Tribulium castaneum. According to Šatović and Plohl [114], interspersed TEs and satDNAs, shape eukaryotic genomes and drive their evolution.
These two mechanisms as well as unequal crossing-over might be the responsible of differential amplification of different satDNA families of the ancestral library found in the common ancestor of different descendant species. This model of evolution of satellite DNA may also explain the differential amplification of the subfamilies of a satellite DNA family [68,183,184]. Thus, HinfI is a subtelomeric satDNA family [185], first isolated in the genera Carthamus and Centaurea [183,185], conserved in all species of the Centaureinae subtribe [68] and in the genome of the species of the Cardueae tribe [184], the largest in the family Compositae. At least nine HinfI subfamilies were present in the common ancestor of Cardueae, each of which has spread differently in different genera of each of the subtribes of this tribe.
3.4.2. Concerted Evolution
Once a satDNA family has been spread out throughout the genome, each repeat at every tandem constituting the different satDNA loci of a species could follow an independent evolution freely diverging from one another or, on the contrary, they could follow a cohesive evolution [41,70]. In fact, contrary to what would be expected in the absence of selective restrictions, members of a satDNA family would show a high degree of intra-specific similarity and inter-specific divergence, following a pattern of concerted evolution [41,70]. This cohesive and gradual evolution would be allowed through a two-step evolutionary process called molecular drive [186,187]. According to this model, new variants that appear by mutation in individual units are expanded to the rest of repets by means of mechanisms like unequal crossing-over, transposition, or reinsertion of replicated extrachromosomal forms, which together with gene conversion, are involved in the homogenization of satDNAs. In a second step, the new expanded variants are fixed in a population through sexual reproduction.
A main topic of interest concerning concerted evolution is the extent of population differentiation. Up to the present, interpopulation concerted evolution of satDNAs was detected only in satDNAs of pupfish [188]. No other studies on satDNA divergence at the population level did succeed in identifying population-specific mutations or other population-specific sequence features of satDNAs (reviewed in [189]). Feliciello et al. [189] have found evidence for the existence of population-specific satDNA profiles. In Tribolium castaneum, there are two highly similar subfamilies of the Tribolium castaneum 2 satDNA (TCAST2). They differ by nine point mutations, each homogenized and fixed within a particular subfamily. Sequences of satellite subfamilies do not differ among T. castaneum strains but they were differentially amplified among strains. According to the proposal of Feliciello et al. [189], these results would support the hypothesis that satDNA could act as driver of genome divergence at the population level. Variation and population differentiation in satDNA abundance among lines of Drosophila melanogaster have also been exposed by Wei et al. [9].
3.4.3. Factors Influencing Satellite DNA Evolution
In the absence of selective and biological restrictions, the concerted evolution rate of a satDNA family depends basically on evolutionary time [190]. In these cases, it may be expected that transitional stages during the propagation of a variant from its onset by mutation to fixation can be clearly classified [191,192,193]. However, concerted evolution depends on several intrinsic and extrinsic factors and in many cases this pattern of evolution is affected and altered in such a way that the differences between species are not always as noticeable as might be expected. Contrary to that, the differences between sequences of the same species might be similar to the differences found between different species. As indicated above, time is important and an elevated number of shared polymorphisms between repeats of two different species might simply indicate a short divergence time between the compared species [190,194]. Not always however, since the rate of sequence evolution of a satDNA family could be slowed or accelerated by the effect of location, organization, and repeat-copy number [26], population and evolutionary factors [33,183], biological factors [195,196,197], or functional constraints [19]. Several examples illustrating these effects are found in the following paragraphs.
3.4.3.1. Chromosomal Location, Organization, and Repeat-Copy Number
Analysis of satDNA in the genus Rumex demonstrated that the chromosomal location affects to the evolutionary rate of this type of repetitive sequences because the location might affect to its recombination rate [25,26]. Eurasian and American dioecious species of the plant genus Rumex (Polygonaceae) form a monophyletic group which is divided in two phylogenetic clades: one composed of species with a simple sex chromosome system XX/XY and another composed by species with a complex sex chromosome system XX/XY1Y2, this later derived from an ancestor with a XX/XY system. Analysis of the synaptonemic complexes and the cytogenetic characterization of sex chromosomes of species XX/XY and XX/XY1Y2, revealed that XX/XY species represent an early stage of genetic differentiation between sex chromosomes. In contrast, species with a sex determination system XX/XY1Y2 showed an advanced state of genetic differentiation between the X and the Y chromosomes [26,149]. In such a way that the Y chromosomes do not mate with each other, pairing only at one end with each of the X ends in a sexual trivalent. Thus, there is no recombination, or is very limited among the Ys, being very restricted between X and Y [149,198]. XX/XY1Y2 species have heterochromatic Y chromosomes but not the XX/XY species [149]. Up to seven satDNA families have been characterized in XX/XY1Y2 species [25,52,146,147,151,168]. One family, the RAE730 family, is unique to the heterochromatic segment of one autosome. The RAYSI family is unique to the Y chromosomes and is therefore not present in females. The RAE180 family is in the Y chromosomes and, in addition, in a small locus of the pair 2 of the karyotype. Therefore, this satellite is present in the genome of males and females, although in females RAE180 is poorly represented [26]. Mechanisms eluding recombination negatively influence concerted evolution. Thus, RAE180 and RAYSI satellites of the Y chromosomes of dioecious species have an evolutionary rate that is half the rate of change of the RAE730 satellite of autosomes [25]. Unlike the other two satellites, RAE180 is also present in dioecious XX/XY species but in a smaller amount and in an autosomal locus, visible by FISH only in some species. Interestingly, comparing XX/XY and XX/XY1Y2 species, the same satellite DNA (RAE180) in different locations has different rates of evolution [26]. Thus, the RAE180 has rates of evolutionary change three times superior when in autosomes (in XX/XY) species than when in Y chromosomes (in XX/XY1Y2 species). In addition, RAE180 sequences located in autosomes show a pattern of concerted evolution that is not observed when comparing sequences of the Y chromosomes [26].
Pavlek et al. [85] have found that concerted evolution acts more efficiently on longer than shorter arrays. Thus, it is probable that ancestral variants in low copy number remnants of the library could escape from the homogenization mechanisms [85,197]. In this sense, it has been proposed that evolutionary periods of stasis would keep variability by the reduced action of molecular mechanisms of non-reciprocal exchange which could be a fact for low repeat-copy number [26]. In this context, it is very interesting also that the repeats placed in the center of one array tend to be homogenized more efficiently than those occurring toward the proximal and distal ends of the array [121,199]. While homogenization mechanisms maintain sequence similarity of repeats within the array, they would also provide divergent repeat variants at the array ends which can be amplified and may be used as a source of new satDNAs [200].
Different arrays on the same or in different chromosomes may experience independent homogenization for arrays- or chromosome-specific repeat variants [122,127,131,201]. Thus, the analysis of euchromatic and heterochromatic repeats from 52 arrays showed that the homogenization of satDNA repeats of the 1688 satDNA family of Drosophila occurred differentially for distinct genomic regions, from euchromatin to heterochromatin and from local arrays to chromosomes [127]. Further, the lack of chromosome transfer between non-homologous chromosomes may lead to chromosome-specific subfamilies within a genome [202]. Thus reduction or elimination of exchanges between non-homologous chromosomes would give rise to the formation of satDNA subfamilies as occur for RAYSI satDNA in Rumex [148], for PIM357 satDNA of the beetle genus Pimelia [203], and for satDNA in spiders of the genus Tetragnatha [204]. Absence of inter-chromosome exchange may lead even to the independent evolution of satDNAs in two subgenomes even at equilocal sites as occur in Sus scrofa domestica (see above). The observed low rates of homogenization of the DBC-150 family might be related to a presumed reduction or suppression of meiotic recombination in the microchromosomes of Drosophila buzzatii [173].
3.4.3.2. Population and Evolutionary Factors
Gene flow between taxa reduces their genetic differences, but also leads to increased intra-specific variation. Therefore, contrary to expectations about the model of concerted evolution, in an evolutionary scenario of reticulated evolution, such as that of the evolutionary history of sturgeons [205], we would find similar or even higher levels of intra-specific variation than inter-specific divergence. This is what occurs in the case of the HindIII and PstI satellite DNAs of sturgeons [33,89]. The repeat sequences of these satellites are not grouped by species affinity in phylogenetic analyses, but the sequences isolated from species belonging to the same phylobiogeographic clade appear intermixed. In addition, the following was observed: (a) high intraspecific variability; (b) low levels of concerted evolution between species belonging to the same phylobiogeographic clade; and (c) a high number of shared polymorphisms between distant species belonging to different clades. Facts, all of them, which could be explained under the pattern of reticulated evolution of these species and interspecific hybridization, common in sturgeons [33,89]. In Cardueae, up to nine subfamilies of the HinfI satDNA have been described. Although phylogenetic trees group the sequences by subfamily affinity rather than by specific provenance, when comparing the repeats of the same subfamily, in most cases, the degree of divergence between any pair of sequences is related to the evolutionary distance between the species compared. However, there are exceptions to this rule, which appear when comparing sequences of species of some genera, such as Centaurea, in which reticulated evolution has been a key factor in its evolution [68,183,184].
3.4.3.3. Biological Factors
Luchetti et al. [195] found that sexuality acts as a driving force in the fixation of sequence variants within a satDNA family thus generating intrapopulation cohesiveness and interpopulation discontinuities, and that parthenogenesis has a slowing effect on molecular turnover processes. However, spreading of new variants in unisexual specimens by gene conversion events was also observed. Therefore, given enough time, sequence homogenization can take place in a unisexual species. A similar situation is found for satDNAs of Y chromosomes of the plant Rumex acetosa. While the Y chromosomes of R. acetosa do not recombine, sister-chromatid interchanges would explain gene conversion homogenizing events which should lead to concerted evolution of Y-linked satDNA subfamilies [148], in resemblance to the human Y chromosome [140]. Accumulated data suggest that evolution of satDNA in ants follows the concerted evolution pattern but that this process is slow in relation with other organisms, probably due to the eusociality and haplodiploidy of these insects [197,206]. As explained in Lorite et al. [197], the haploid males of ants lack meiotic recombination and so the mutation rate in males could counteract the effectiveness of the genome turnover mechanisms. According to these authors, this could partially explain the slowdown in the homogenization and fixation processes [197]. On the other hand, they also explain that eusociality hinders random mating and reduces the number of reproductive individuals to a few units, leading to a high level of variability uniformly distributed among related taxa [196,197]. The effect of autogamy apparently leads to a lack of species-specific variants of ATR-2 satDNA in Arachis species (Leguminosae) [178].
3.4.3.4. Functional Constraints
SatDNA is one of the most dynamic components of genomes, undergoing rapid changes in array size and sequence composition within a short evolutionary period [41,70]. Expansion and shrinkage of satDNAs contribute significantly to the array-length polymorphism as well as to the replacement of the most abundant variant with a different variant [41,70]. Further, rapid amplification of one satDNA family from the library, rapidly changes any profile of genomic satDNA [17,70]. Thus, satDNA sequences are rapidly evolving sequences that might cause reproductive barriers between organisms and promote speciation [17,71,189,207,208,209,210,211,212]. However, some satDNAs exhibit sequence conservation of part or of the whole monomer sequence for long evolutionary periods [18,19,24,33,34,35,36,37,38,39,40]. Functional constraints might be influencing in the preservation of satDNAs found in beetles, clams, or several plant groups where some monomer sequences might be evolutionarily preferred in comparison with others [18,19,24,39,40,72]. Some monomers may be preferred because of their functional potential and/or simply because particular combinations of nucleotides and structural features of the DNA molecule are favored by homogenization mechanisms [69,70]. However, biological factors might be responsible for sequence conservation [33,72]. Some satDNAs repeats conserve motifs that are remnants of shorter ancestral repeat monomers which led to the current repeats by duplication and divergence [91]. In contrast, sequence motif conservation might be the consequence of selection drive action. For example, in Schizosaccharaomyces pombe, small interfering RNAs (siRNAs) involved in heterochromatin formation derive preferentially from the most conserved regions of heterochromatic repeats [213], which suggest that conservation in this case is due to functional constraints [72,213]. The satDNs’ most known motif is the centromere protein B (CENP-B) box, a 17-bp long motif that is preserved in repeats of primate alpha satDNA harboring the centromeres and in the unrelated centromeric satellite of mouse [214,215,216,217]. The DNA binding domain of the CENP-B protein recognizes and binds the CENP-B box in the centromeric alpha satDNA and mouse minor satDNA. The CENP-B protein and the CENP-B box are largely conserved in mammals [214,218,219,220,221,222]. CENP-B protein facilitates centromere formation and plays an important role in the assembly of specific centromere structures in interphase nuclei and on mitotic chromosomes [221]. In humans, CENP-B is involved in centromere functions, such as de novo centromere protein A (CENP-A) chromatin assembly, CENP-A nucleosome stabilization, and fidelity enhancement of chromosome segregation [222]. Contrary to CENP-B, other centromeric proteins, such as CENP-A, essential for centromere identity and function, are not DNA sequence-specific binding proteins (see below). In the next section, I address this and other questions related to the functional roles of satDNA.
3.5. Satellite DNA Function
Over time, it has been accepted that satDNA would be composed of non-coding and non-transcribed repeats associated with heterochromatin with no immediate use, “junk” DNA, even worse “selfish” DNA [41,223,224,225]. Further, the role of satDNA sequences in any biological process was discarded. It was intriguing thus the transcriptional activity of some specific tandem repetitive repeats in newts during embryogenesis [226,227,228,229,230]. Therefore, transcripts of satDNA in oocyte lampbrush chromosomes of newts were viewed as a failure of normal transcription termination probably as a result of read-through from upstream structural gene promoters [226,227,228,229,230]. Similar conclusions were obtained from the analysis of the satDNA transcription in lampbrush chromosomes of pigeon and chickens. In these cases, it was proposed that transcription of repeat sequences was related to its genomic organization [231,232]. Nowadays, however, there is evidence for specific roles of satDNA transcripts on gene and genome regulation. In addition, the early view of satDNA as junk or even garbage DNA has evolved. SatDNA repeats form the centromere locus and the heterochromatin of the pericentromeric area [69,70,71]. In addition, other roles for satDNAs have also been suggested such as chromosome organization, pairing, and segregation [69,70,71]. Furthermore, satDNA transcripts may be involved in the assembly of the kinetochore, in the control of telomere elongation, capping, and replication, in the epigenetic regulation of heterochromatin establishment and maintenance, in the transcriptional response during stress and in the modulation of gene expression [72,233,234].
3.5.1. Centromeres and Pericentromeric Heterochromain
The centromere is a critical locus for the perpetuation of the genetic material that regulates chromosome segregation during mitosis and meiosis by assembling and directing the organization of the kinetochore, a proteinaceous complex that attaches chromosomes to the spindle [69,70,71,235,236,237]. On the other hand, pericentromeric satellite repeats are essential elements that stabilize interactions with DNA binding proteins, maintain heterochromatin architecture, sustain kinetochore formation, maintain sister-chromatid cohesion, and drive chromosomal segregation during mitosis and meiosis [2,69,70,71,72,233,238,239].
As mentioned before, the centromeres of most eukaryotes are mainly composed of satDNA. However, there are exceptions. The centromeres of the budding yeast Saccharomyces cerevisiae span only about 125 bp of a single copy sequence divided in three conserved DNA elements and are assembled into a single Cse4 (a centromeric H3 histone variant, CENH3) nucleosome that captures a single microtubule [71,235,236,237]. These so-called “point centromeres”, constitute the most known exception to the alternative, most common, “regional centromere” consisting of multiple CENH3 nucleosomes that capture several microtubules [71,235,236,237]. The regional centromere of fission yeast Schizosaccharomyces pombe is formed of unique or low-copy sequences flanked by heterochromatic regions [71,235,236,237]. In most animal and plant species, the regional centromere is a “satellite centromere” [237]. The centromere contains large arrays of satDNA sequences which, especially in the case of plants, might be interrupted by mobile elements and it is also flanked by heterochromatin [70,71,96,236,237,240]. However, these centromeric satDNA repeats varies substantially among species, conserved at times only between closely related species, and often being species-specific [41,70,71,77]. In fact, cases have been found in which two or more unrelated satDNA can be found at different sets of centromeres within the same species [241,242,243]. Furthermore, there are functional centromeres lacking satDNA. Six of the twelve potato chromosomes have tandem repeat-based centromeres, but five centromeres do not contain a tandem repeat [242]. It has been found that while most chromosomes of the chicken, horse, and orangutan complement are composed of satDNA, some of their chromosomes contain non-repetitive DNA [236,244,245,246,247]. The centromere in these cases resembles that of neocentromeres formed when centromeres are disrupted in humans [236,248]. Thus, functional neocentromeres may lack centromere-specific sequences and heterochromatin both in plants and animals [236,237,249]. Also, only one of the two centromeres containing the DNA elements of functional centromeres is active in dicentric chromosomes [250,251,252]. Beyond, centromeric retrotransposons are important elements of centromeres of a wide range of angiosperm species [117]. In rice and maize, all centromeres have different retrotransposon families [118,120], and retrotransposons are the main component of banana and some wheat centromeres [42,119].
All these data prove that DNA sequences alone are insufficient to determine centromere identity and have revealed the importance of epigenetic factors regulating the centromere identity and function. Regional centromeres of both plants and animals are characterized by the presence of a centromere-specific histone H3 variant CENH3 and are organized as euchromatic domains of CENH3, flanked by heterochromatic domains [70,71,236,237,240,253,254]. Within the centromeric chromatin, CENH3 is interspersed with histone H3 dimethylated at lysine 4 (H3K4me2) and associated repeats are hypomethylated with respect to those found in heterochromatin. Conversely, pericentromeric chromatin is enriched for nucleosomes containing histone H3 methylated at lysine 9 (H3K9me), a mark associated with heterochromatin. For example, a typical human centromere is composed of a fraction of HOR α satDNA built from subdomains of nucleosomes containing centromeric CENP-A (the human histone H3 variant CENH3) interspersed with H3K4me2 and the remainder HORs and monomers of α satDNA are assembled into heterochromatin enriched for nucleosomes containing H3K9me [70,71,253]. Similar to human centromeres, the CENH3 nucleosomes of other analyzed species typically occupy only a portion of the satellite repeats and the rest of repetitive elements are embedded in flanking pericentromeric heterochromatin [240,249,254]. The total size of CENH3 domains is similar between different chromosomes independently of their length or the length of satellite arrays [249].
Most centromeric retrotransposons tested to date are actively transcribed and these transcript might have a role in centromere formation, maintenance, and function [117]. Interestingly, many centromeric satDNA are also transcribed. Transcription of pericentromeric and centromeric repetitive sequences seems to have roles not strictly related to heterochromatin establishment but also in maintaining centromere identity, being an important functional component of the centromere/kinetochore complex [72,233,255,256,257]. Therefore, changes in centromeric satDNA transcription may affect chromosome stability and segregation [72,258,259]. There is a growing body of work showing that transcription of centromeric satDNA in long, single-stranded transcripts encompassing a few satellite monomers [260,261,262], contributes to assembly of the kinetochore, being important for the localization of CENP-A as well as of the kinetochore proteins centromere protein C (CENP-C), inner centromere protein (INCENP) and surviving (an INCENP-interacting protein) at the human centromere [234,260,262]. Recently, it has been showed that centromeric RNAs are processed during mitosis, and that recruitment of the splicing machinery to these transcripts is important for kinetochore assembly [263]. However, it appears that recruitment of the RNA processing machinery to the centromere, but not the persistence or even complete synthesis of mature centromeric RNAs, is important for kinetochore assembly [234]. Transcripts of α satellite play an important role in kinetochore formation but also in the establishment of pericentromeric heterochromatin and are indispensable for the proper cell division [257]. SAT III RNA of Drosophila melanogaster is an integral part of centromere indentity by binding to the kinetochore component CENP-C, and because is involved in the correct localization of the centromere-defining proteins, CENP-A and CENP-C, as well as outer kinetochore proteins [264]. Long RNAs not processed into siRNA are also characteristic of centromeres of rice (40 nt), wallaby (34–42 nt), mouse (120 nt), and beetles (multiple transcripts of heterogenous size longer than 500 nt, up to more than 5 kb) [72,258,265,266,267,268]. In rice, long transcripts from centromeric satDNA are processed in RNAs of 40 nt and they act together with 21–24 nt long siRNAs that might derive from the pericentromeric portion of the same satellite [265]. Similarly, the centromeric RNAs of wallaby and mouse are processed from longer double stranded RNAs [266,267]. Thus, RNAi together to an alternative mechanism involving longer RNA might be operating in kinetochore establishment [72].
There is still another type of centromere in addition to point and regional centromeres, the holocentromere, which is spread along the entire length of the chromosome. Holocentric chromosomes lack a primary constriction, in contrast to monocentrics, forming kinetochores distributed along the entire chromosome and microtubules can attach along most of the poleward facing surface [269,270]. The holocentric chromosomes have also been called diffuse-kinetochore chromosomes, holokinetic chromosomes, and polykinetic chromosomes [269,270]. Holocentric chromosomes evolved at least four independent times in plants and at least nine independent times in animals [269,270]. The holocentric chromosomes have been found in the monocots families Juncaceae and Cyperaceae and in the genus Chionographis (family Melanthiaceae). Not all genera in Cyperaceae and Juncaceae families are holocentric, but there are some of their genera, as Luzea, with holocentric chromosomes. Holocentric eudicots are limited to two genera, Drosera (family Droseraceae) and Cuscuta (family Convulvulaceae), but not all the species of Cuscuta have holocentric chromosomes [269,270]. Recently, Neumann et al. [241,271,272] have identified in species of the legume genera Lathyrus and Pisum the so-called meta-polycentric chromosomes, monocentric chromosomes with multiple centromere domains, representing a putative intermediate between monocentric and holocentric chromosomes. These domains in Pisum sativum are almost entirely composed of repetitive DNA sequences belonging to 13 satDNA distinct families and 1 family of centromeric retrotransposons, all of which are unevenly distributed among pea chromosomes [241]. In the animal kingdom, holocentric chromosomes have been found in two phyla, Nematoda in which holocentric chromosomes arose once and Arthropoda in which this type of chromosomes arose at least eight times [269].
Plant holocentromeres have been extensively studied in the Cyperaceae species Rhynchospora [273,274]. Holocentromeres of Rhynchospora pubera are highly enriched by the “Tyba” satDNA family that occurs as genome-wide interspersed arrays [273]. This satDNA family is centromere-specific and associates with CENH3 [273]. Centromeric arrays vary in length from 3 to 16 kb and are intermingled with gene-coding sequences and transposable elements [273]. In contrast to other satDNAs, Tyba is dispersed in interphase and with the onset of mitotic condensation, linear satDNA structures along all chromosomes rather than clustered blocks are formed [273]. Thus, holocentromeres of metaphase chromosomes in this species are composed of multiple centromeric units rather than possessing a diffuse organization. A cell-cycle-dependent shuffling of multiple centromeric units results in the formation of functional polycentromeres during mitosis. Holocentricity influences the chromosomal organization of different satDNA families in different species of Rhynchospora [274]. Thus, while more conserved centromeric repeats revealed linear signals, noncentromeric species-specific satDNA formed distinct clusters along the mitotic chromosomes. In interphase, the centromeric satDNAs appeared dispersed while non-centromeric species-specific satDNAs appeared clustered [274]. No specific repeats have been found to be associated with CENH3 loci in Luzula elegans despite previous efforts to find them [270,275]. In fact, CENH3 has been independently lost in four insect lineages that transitioned from monocentricity to holocentricity [237,276].
Holocentric nematodes have a large number of satDNAs, apparently very few evolutionarily conserved, scattered throughout their genome. In contrast, no scattered satDNAs are found in the monocentric nematode Trichinella spiralis [48]. Centromere-like satDNAs described in Caenorhabditis elegans may accumulate CENH3, are located in regions with high CENH3 affinity, promote kinetochore formation during mitosis, and have sequence features similar to satDNAs of monocentric species [49]. Subirana et al. [49] found that satellites are randomly distributed in domains of either low or high affinity for CENH3, but that all the longest satellites with longest repeat had higher affinity for CENH3. However, these results contrast with those of Gassmann et al. [277] and those of Steiner and Henikoff [278], who found that CENH3 occupies nonrepeated regions. Steiner and Henikoff [278] did find that holocentromeres show a polycentric distribution, with each site containing a single-wrap centromeric nucleosome and that centromeric sites correspond to transcription factor hotspots, which are bound by multiple transcription factors, but which lack their characteristic sequence-specific DNA-binding motifs [237,278], and do not coincide with satellite positions [49]. Gassman et al. [277] did find that CENH3 occupies nonrepeated regions of 10–12 kb dispersed across about half of the genome and is excluded from loci that are transcribed in the germline and early embryo.
Independently of sequence conservation, satDNAs may stabilize CENH3 nucleosomes. Thus, satDNA would maintain sequence homogeneity crucial for centromere stability and at the same time it can be a source of extremely rapid changes at centromere [70,71]. Stabilization of CENH3 nucleosomes could explain how new satellites arise and populate centromeres and how neocentromeres based on unique sequences can evolve to repetitive centromeres presumably to stabilize them [236,249]. Tandem duplication of a new sequence with a selective advantage for CENH3 stabilization or transposition of existing satDNAs could populate an epigenetically defined neocentromere [242,249].
The CENH3 variant is the defining chromatin component of centromeres in most eukaryotes [276]. Notwithstanding the foregoing, species-specific CENH3 proteins have been identified in all eukaryotes investigated so far, including humans (CENP-A), budding yeast (Cse4), fission yeast (Cnp1), Caenorhabditis elegans (HCP-3), Drosophila melanogaster (CID), and different plant species [71]. The “centromere drive” model proposes the unequal transmission of competing variant centromeres in the asymmetric female meiosis [208,211,279]. Thus, it has been proposed that positive selection for mutations in genes coding for kinetochore proteins that favor the affinity of these proteins (e.g., CENH3) for centromere sequence variants [71,208,211]. Co-evolution of centromeric satDNAs and kinetochore proteins would explain in this way the variety of eukaryotic centromeres and the rapid evolution of both, sequences and proteins, at centromeres [71,208,211,279]. Rapid changes among individuals in the centromere would lead to reduced compatibility of homologous chromosomes in hybrids and ultimately to postzygotic isolation, thus triggering speciation [71,211,280,281].
According to the centromere drive model, adaptively evolving CENH3 has not been detected in lineages having symmetric meiosis [208,220,279,282,283,284,285,286]. This is because evolutionary changes of CENH3 depend also on the chromosomal structure in terms of kinetochore formation [279]. Thus, most studies addressing CENH3 evolution were carried out on species with monocentric centromeres [279,284]. Also, studies were conducted on Caenorhabditis that, although they have holokinetic chromosomes in mitosis, form a cup-like kinetochore in meiosis with ambiguous utilization of CenH3 [279,284]. Thus, the centromere drive model has been questioned recently by Neumann et al. [271]. Two paralogous variants of CENH3 were identified in Fabaceae, CENH3-1 and CENH3-2, which originated from a duplication event in the common ancestor of Fabeae species. Subsequently, while the CENH3-1 gene was lost or silenced in the lineage leading to Vicia and Lens, having monocentric centromeres, CENH3-1 and CENH3-2 have been retained in the polycentromeric species of Pisum and Lathyrus. Both genes appear to have evolved under purifying selection and produce functional CENH3 proteins which are fully colocalized [271]. These results are contrary to the predictions of the centromere drive model. Indeed, Zedek and Bureš [279], analyzing CENH3 isoforms in 13 Luzula species did not detect positive selection, but on the contrary they found relaxed selection on CENH3. These results have been interpreted from an evolutionary point of view in such a way that holocentromeric chromosomes might have evolved as a defense, suppresing centromere drive [279,287]. Thus, it is possible that holokinetism has evolved only in lineages with asymmetric meiosis where it might be a necessary defense against centromere drive, while in lineages with symmetric meiosis such a defence has no purpose [287].
3.5.2. Subtelomeric Heterochromatin
The telomeres protect chromosomes from degradation and repair activities and prevent chromosome shortening that results from replication of the end of the linear chromosomes [288,289]. Different telomere-specific proteins build telomeres to accomplish these functions [288]. Concerning DNA sequences, telomeres are organized in an ancestral conserved structure based in the repetition of short tandem repeats of about 6 bp: 5′-TTGGGG-3′ (Tetrahymena), 5′-TTAGG-3′ (insects), 5′-TTAGGG-3′ (vertebrates), 5′-TTTAGGG-3′ (plants). However, there are some exceptions to this general rule. The vertebrate telomere was also found in marine invertebrates [290]. Many species of Asparagales have telomeres composed by the human-type TTAGGG repeats instead of the plant-type TTTAGGG repeats [291,292]. In Cestrum (Solanaceae), the telomeres have the TTTTTTAGGG motif [293]. Despite some variation, the human-type repeat appears to be the ancestral and the most common telomere repeat in eukaryotes [294]. However, alternative motifs have been replacing it along the evolutionary time of diverse eukaryotic lineages [294].
Even in the event that telomeric repeats are not included within the category of satDNA, they share many similarities with satDNA sequences and are intimately related with subtelomeric satDNA repeats. However, they are originated and amplified by a different mechanism depending on telomerase, a reverse transcriptase that adds telomeric DNA to telomeres using associated RNA as a template.
Notwithstanding, there are exceptions to this general rule. Thus, Drosophila telomere maintenance is different and it is based on targeted transposition of three non-LTR (long terminal repeat) retrotransposons [295], which reinforce the suggestion that a retrotransposon gene was “domesticated” for a cellular role early in the evolution of eukaryotes [1]. On the other hand, Allium species lack any known telomeric sequence [296]. Instead, the chromosomal ends of Allium species consist of satDNA [297], which suggest a functional role for satDNA in telomeric function, at least in such exceptional cases (see below).
Subtelomeric heterochromatin is composed of tandem repeats with a variety of lengths and repeat numbers [41,298]. However, it has been described some cases of high level of sequence conservation between species [42]. Subtelomeric repeats have been reported in numerous plant and animal species. For example, the human subtelomere structure and variation has been extensively analyzed [299,300]. In rye, several satDNA families have been described with different repeat lengths and great variability in organization in the subtelomeric heterochromatin [301,302,303,304]. CL14 and CL34 repeats show a complex satellite repeat composition of subtelomeres through Solanum species [305] as well as khipu and jumper satDNAs at subtelomeres of Phaseolus [306]. Subtelomeres are fast evolving regions [304,307]. Many subtelomeric satDNA sequences are species-specific, often chromosome-specific. The existence of several subtelomeric satDNA families or even subfamilies in one species is also common [301,308]. Rearrangements by recombination between non-homologous chromosome ends may lead to the formation of new satDNA families [91,305].
Frequently, short and often degenerate tracts of telomeric repeats are interspersed within the subtelomeric region and even occasionally within the single copy sequences adjacent to the subtelomeric repeats [27,29,47,300,309,310]. Interstitial telomeric repeats can also be found at centromeres as evolutionary relics derived from chromosomal rearrangements [27,311], although telomeric repeats invaded and were amplified within the functional centromeres of Solanum species [312].
Several functional roles has been proposed for subtelomeric DNA [298,300,313,314]: (a) genome stability; (b) faithful chromosome replication; (c) faithful chromosome pairing and segregation; (d) cell cycle regulation; (e) cellular aging and immortalization; (f) movements and localization of chromosomes within the nucleus; (g) transcriptional regulation of subtelomeric genes; (h) buffer terminal genes against the dynamic processes of loss and adition of telomeres. In addition, subtelomeric repeats may have an incidental role in cases of losing of conventional telomeric repeats. In such cases, chromosomal ends have been stabilized by subtelomeric satDNA [297,315]. In any case, as mentioned, these roles are sequence-independent. Further, it has been demonstrated that telomere-like sequences interspersed within subtelomeric DNA may play a role in subtelomeric recombination and transcription, in telomere maintenance via the alternative lengthening of telomeres (ALT) pathway, and in telomere healing [300,316,317].
Telomeric repeat-containing RNAs (TERRA) are long non-coding RNAs with sizes ranging between 100 nt and 9 kb, transcribed from telomeric C-rich strand of telomeres, initiating in subtelomeric regions, thus containing subtelomeric and telomeric sequences [233,318,319]. These RNAs were detected first in human and rodent as nuclear restricted RNAs that showed high levels in all adult tissues with low or no telomerase activity while showing low levels in cancer cells [318,319,320]. TERRA RNAs remain associated to telomeric chromatin as components of mammalian telomeres where negatively regulate telomere length and are involved in shortening of telomeres. TERRA transcripts have been detected associated with heterochromatin marks typical of telemores (H3K9me3) as well as with different telomere proteins including heterochromatin protein 1 (HP1) and the telomere repeat factors 1 and 2 (TRF1 and TRF2), among others, thus playing an important role in the maintenance of telomeric heterochromatin [233,321]. Over-elongation of telomeres leads to synthesize longer TERRAs which are involved in the recruitment of histone methyltransferases with the subesquent histone H3 methylation and the recovery of HP1, which in turn induce the transcriptional repression of TERRAs, preventing the heterochromatin hyperformation [318,319,320,321,322]. TERRA is thus involved in heterochromatin establishment but also has a role in the regulation of telomere length, in telomere capping and replication [233]. TERRA RNAs are evolutionarily conserved [319]. They have been described in different vertebrates [320] and yeast [323], as well as in Arabidopsis [324]. A portion of TERRA transcripts of Arabidopsis are processed into siRNAs, which promote methylation of cytosines in telomeric repeats and contribute to the maintenance of telomeric chromatin [324].
3.5.3. Heterochromatin Assembly
Heterochromatin plays an essential role in preservation of epigenetic information, transcriptional repression of repetitive DNA, and proper chromosome segregation [72]. Even in the event of the difficulties for its analysis, today there is a great accumulation of data revealing the importance of heterochromatin in those roles. Specifically, the role of pericentromeric and subtelomeric satDNA transcripts is being brought to light.
Transcription from pericentromeric repetitive sequences has been reported in a number of plants and animals [71,72,239,325]. Small 20–25 bp long RNAs derived from centromeric DNA repeats of the fission yeast S. pombe are involved in heterochromatin assembly by the means of RNA interference (RNAi) processes [326]. For heterochromatin establishment, which in turn leads to a transcriptionally silent state, only low level of expression of repetitive DNA is necessary. This silencing mechanism of the chromatin is accomplished by RNA-induced transcriptional silencing complex (RITS) built by the association of Argonaute protein and siRNAs. Interestingly, these siRNAs are derived from the processing by Dicer of double-stranded RNA that arises from bidirectional transcription of repeated centromeric DNA [326]. RITS complex contributes to the formation of pericentromeric heterochromatin, whose establishment is necessary for proper chromosomal segregation in mitosis and meiosis [326]. Formation of heterochromatin also requires histone H3 methylation at lysine 9 (H3K9me2), essential for the recruitment of heterochromatin protein 1 (HP1). This promotes silencing gene expression, recombination, kinetochore assembly, and prevention of erroneous microtubule attachment to the kinetochores [72,326]. The RITS complex uses base pairing interactions between the loaded siRNA and nascent transcripts (along with H3K9me2 interactions) to localize to pericentric regions. Localized RITS complexes recruit histone methyltransferases, leading to H3K9 methylation, HP1 recruitment (Swi6 in S. pombe), heterochromatic silencing, and generation of more siRNAs through recruitment of Dicer and an RNA-dependent RNA polymerase (RdRP) [327,328]. In this way, heterochromatin is maintained through a self-reinforcing positive feedback loop; siRNA promotes H3K9 methylation which promotes siRNA production, and both siRNA and H3K9me2 interact with the RITS complex to facilitate its localization to pericentric heterochromatin [327,328].
Like in fission yeast, small RNAs contribute to heterochromatin regulation in fungi, ciliates, plants, and worms acting through RNA-dependent RNA polymerase (RdRP) , self-reinforcing feedback loops and histone methylation [327,328]. In plants, in addition to histone methylation, siRNAs also direct the methylation of the DNA from which they derived [329]. The regulation of heterochromatin assembly by siRNAs of 21–24 nt derived from satDNAs has been proven in several plant species such as Arabidopsis, rice, and sugar beet [265,329,330]. However, the involvement of RNAi in heterochromatin formation in other eukaryotic organisms is still debated [72,233]. Dicer and Argonaute protein families, components of the RNAi machinery, have been identified in the genome of the beetle Tribolium castaneum, but not the RdRP [72] which appearently has been lost in most insects and vertebrates [328]. Evidence for the regulation of heterochromatin assembly through an RNAi mechanism has been found in chicken [331], in C. elegans [332], in Drosophila melanogaster [333], and in mouse [334]. However, it is not still clear how heterochromatin regulation is mediated in these organisms. It appears that siRNAs are not involved in heterochromatin remodeling in mammals. However, it appears that a RNA is required but this RNA is still unindetified [335]. An RNAi-dependent mechanism that involves telomerase reverse transcriptase in heterochromatin establishment and maintenance has been recently proposed [233,336].
A relation between misregulation of centromeric and pericentromeric satDNA transcription and tumors has been established. De-condensation of pericentromeric heterochromatin and over-expression of pericentromeric satellite repeats is characteristic of some tumors and genetic disorders [72,233,325,337,338,339]. Three different studies revealed the overexpression of pericentromeric satDNAs in human tumors [325,337,338]. For example, the pericentromeric human satellite II (HSATII) is aberrantly overexpressed in a wide variety of epithelial cancers [338]. Induction of HSATII transcription is triggered by growth of cells under nonadherent conditions. The HSATII transcripts are reversely transcribed and this RNA-derived DNA is reintegrated with the subsequent expansion of HSATII loci [239]. HSATII copy number gain is a common feature in primary human colon tumors and is associated with a lower overall survival. Thus, cancer-associated derepression of specific repetitive sequences can promote their RNA driven genomic expansion, with potential implications on pericentromeric architecture [239]. Further, this study illustrates the point that satDNA profile might evolve rapidly in a species by retrotransposition of some satellites over others, at least under certain circumstances such as in the setting of cancer.
On the other hand, chromosomal instability induced by overexpression of α satellite transcripts appear involved in the development of breast and colorectal cancers [340,341,342]. Thus, although some alpha satellite transcription may be necessary for proper centromere and/or heterochromatin function, overexpression may have damaging effects [328].
3.5.4. Gene Regulation
Promoter elements and transcription start sites, as well as binding motifs for transcription factors, have been mapped within some satellites which could influence nearby genes, probably by the mechanism of transcriptional interference [72,343].
Human satellite III DNA is transcribed in response to stress [72,233,344]. In human cells, the heat shock response is characterized by the transcriptional activation of both protein coding heat shock genes and noncoding repeated satellite III DNA sequences located at pericentromeric heterochromatin [345]. The transcriptional activation of sat III by the HSF1 protein (Heat Shock Factor 1, a transcription factor present in the nucleus and cytoplasm of cell before heat shock) could represent a powerful way to trigger a rapid remodeling of gene expression at a genome-wide scale in heat-shocked cells [345]. In addition to heat shock, various cellular stresses may also influence the expression of satDNA [233]. In humans, an example of gene expression mediated by heterochromatin transcripts is found in the testis-specific RNA transcripts from the MSY distal heterochromatic block of the Y chromosome which are involved in trans-splicing with CDC2L2 kinase mRNA generating a testis-specific isoform [71,346].
On the other hand, PRAT and PSUB satDNAs of beetles of the genus Palorus are differentially expressed depending on developmental stages and tissue types [258,268]. In such a way, transcription of many satDNAs have been found to depend on development during which satDNA transcripts may act as epigenetic signals required for centromere stability [347,348] and organization of pericentromeric heterochromatin, and might be necessary for differentiation and developmental progression [72,233].
Thus, several examples suggest that transcripts from satDNA might regulate gene expression in particular cases [72,233,327]. However, there are few experimental studies proving it. Nonetheless, there are two recent studies that demonstrate the specific role of siRNAs derived from processed noncoding RNAs transcribed from satDNA. The TCAST1 satDNA of the beetle Tribolium castaneum comprises the pericentromeric heterochromatin but is also dispersed within euchromatin [21,128,130]. Heat shock induces TCAST1 expression. Transcripts of this satDNA are long double-strand transcripts that are processed into 21–30 nt siRNAs which trigger an increase in repressive epigenetic modifications of histones at satDNA regions in heterochromatin [130,349]. Feliciello et al. [130] have demonstrated that short stretches of euchromatic TCAST1 satDNA loci modulate gene expression by enhanced suppression of activity of TCAST1-associated genes and slower recovery of their activity after long-term heat stress. The level of gene suppression is stronger as the number of copies of TCAST1 repeats is higher. The gene suppression consists in heterochromatin assembly mediated by siRNAs at euchromatic TCAST1 loci and surrounding regions. These authors found differences in the pattern of distribution of TCAST1 elements between populations which according to their proposal might contribute to gene expression diversity among populations of this species. Furthermore, they proposed that this diversity might have an adaptative impact on populations to different environmental conditions. Vlahović et al. [108] have proposed that HORs could act as gene regulatory elements and that variation in HOR composition among individuals or populations can generate gene expression diversity and contribute to the evolution of gene regulatory network. The second example focus in the control of dosage compensation in Drosophila melanogaster males, in which satDNA directs male-specific gene expression [350]. The resulting imbalance in gene dosage between males and females in D. melanogaster is compensated by increased expression from the single X chromosome of males. Processed siRNAs from transcripts of a family of X-linked satDNA repeats promote X recognition [351]. Joshi and Meller [352] have demonstrated that these small noncoding RNAs generated from X-linked satDNA repeats are cis-acting elements that guide dosage compensation machinery preferentially find X sequences. The siRNA pathway therefore promotes recognition guided by chromosome-specific repetitive sequences [352].
In addition, Hall et al. [353], have demonstrated also the crucial role of human HSATII satDNA in gene regulation. Small blocks of HSATII are found on the pericentromeres of 11 human chromosomes, but chromosome 1, and to a lesser extent chromosome 16, carries very large (~5–6 Mb) blocks of this ~26-bp tandem repeat. This study provides evidence for the biological significance of HSATII. They postulate that it may have a normal function during development, such as global epigenomic programming in early development, or gametogenesis. This proposal is based on the capacity of both HSATII DNA and RNA to amass and sequester accumulations of two epigenetic factors that regulate heterochromatin, the polycomb group (PcG) complex PRC1 (protein regulator of cytokinesis 1), and the MeCP2 (methyl-DNA binding protein 2). In cancer, PRC1 bodies form on the demethylated HSATII of the mega-satellite in chromosome 1, while MeCP2 bodies form on HSATII RNA, potentially leading to further changes in the epigenome. Thus, HSATII DNA and RNA can bind and impact distribution of chromatin regulatory proteins, which goes awry in cancer. In altered cells, two types of cancer-specific nuclear bodies are formed, which is caused by locus-specific deregulation of HSATII. The study indicates that HSATII demethylation not only may be caused by, but may contribute to epigenetic instability in cancer, possibly providing a survival advantage (e.g., a bigger ‘sponge’ for PcG proteins) [353].
4. Concluding Remarks and Perspectives
“An evolving topic” has tried to show the evolution of the concept of satellite DNA through time since its discovery in the early 1960s until today. The cause of this evolution has been the continuous advance of the technology used to address the issue. Although the definition of satDNA as highly repetitive DNA organized in tandem is still valid, nowaday we have a wider view of the term and we know that the concept includes a large variety of highly variable sequence types in terms of nucleotide composition, length and structure of repetitive units, organization, and localization. The effort of many researchers over these years has allowed us to better understand various aspects of the very nature of the diversity of satDNA sequences but also of the evolution of satDNA and of the multiple factors influencing it. The approach to the study of satDNA from a genomic perspective has also helped us to better understand the complete set of satDNA families composing the genome of eukaryotic species and their evolution, as well as to know the relationships between the different satDNA families and between chromosomes. On the other hand, satDNA, initially considered ‘junk’ DNA, the largest ‘garbage’ among the ‘rubbish’ that supposedly constituted the plentiful panoply of types of repeated DNA sequences in the eukaryotic genome [41], is now revealed as a part of the genome that has important implications in chromatin regulation as well as in gene regulation, and which plays important structural roles in vital functions, such as segregation or preservation of the genetic material.
After a few years in the early 21st century in which the technique relegated ostracism to the advance of knowledge about satellite DNA, today we can be optimistic and believe that both the genomic approach and the most advanced technology in the field of genes and genomics regulation will soon allow us to find out additional, even unknown, details of this fascinating part of the eukaryotic genome. The advance in the knowledge of functional roles of satDNA as well as its role in cancer onset and progression will be of special interest.
Acknowledgments
The research in our laboratory is currently financed by Grupos de Investigación de la Junta de Andalucía (Group number BIO109). The author thanks J.P.M. Camacho (University of Granada) for the critical reading of the article.
Conflicts of Interest
The author declares no conflict of interest.
References
- López-Flores, I.; Garrido-Ramos, M.A. The repetitive DNA content of eukaryotic genomes. Genome Dyn. 2012, 7, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Biscotti, M.A.; Olmo, E.; Heslop-Harrison, J.S. Repetitive DNA in eukaryotic genomes. Chromosome Res. 2015, 23, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Gregory, T.R. Genome size evolution in animals. In The Evolution of the Genome; Gregory, T.R., Ed.; Elsevier: Burlington, NJ, USA, 2005; pp. 3–87. [Google Scholar]
- Jaillon, O. Genome duplication in the teleost fish Tetraodon nigroviridis reveals the early vertebrate proto-karyotype. Nature 2004, 431, 946–957. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.D.; Leitch, I.J. Genome size evolution in plants. In The Evolution of the Genome; Gregory, T.R., Ed.; Elsevier: Burlington, NJ, USA, 2005; pp. 89–162. [Google Scholar]
- Piegu, B.; Guyot, R.; Picault, N.; Roulin, A.; Saniyal, A.; Kim, H.; Collura, K.; Brar, D.S.; Jackson, S.; Win, R.A.; et al. Doubling genome size without polyploidization: Dynamics of retrotransposition-driven genomic expansions in Oryza australiensis, a wild relative of rice. Genome Res. 2006, 16, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.T.; Pattyn, P.; Bakker, E.G.; Cao, J.; Cheng, J.-F.; Clark, R.M.; Fahlgren, N.; Fawcett, J.A.; Grimwood, J.; Gundlach, H.; et al. The Arabidopsis lyrata genome sequence and the basis of rapid genome size change. Nat. Genet. 2011, 43, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Piednoël, M.; Aberer, A.J.; Schneeweiss, G.M.; Macas, J.; Novak, P.; Gundlach, H.; Temsch, E.M.; Renner, S.S. Next-generation sequencing reveals the impact of repetitive DNA across phylogenetically closely related genomes of Orobanchaceae. Mol. Biol. Evol. 2012, 29, 3601–3611. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.H.C.; Grenier, J.K.; Barbash, D.A.; Clark, A.G. Correlated variation and population differentiation in satellite DNA abundance among lines of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2014, 111, 18793–18798. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, T.S.; Wakefield, M.J.; Aken, B.; Amemiya, C.T.; Chang, J.L.; Duke, S.; Garber, M.; Gentles, A.J.; Goodstadt, L.; Heger, A.; et al. Genome of the marsupial Monodelphis domestica reveals innovation in non-coding sequences. Nature 2007, 447, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Macas, J.; Neumann, P.; Navratilova, A. Repetitive DNA in the pea (Pisum sativum L.) genome: Comprehensive characterization using 454 sequencing and comparison to soybean and Medicago truncatula. BMC Genom. 2007, 8, 427. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ruano, F.J.; López-León, M.D.; Cabrero, J.; Camacho, J.P.M. High-throughput analysis of the satellitome illuminates satellite DNA evolution. Sci. Rep. 2016, 6, 28333. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.; Sutton, G.; Ng, P.C.; Feuk, L.; Halpern, A.L.; Walenz, B.P.; Axelrod, N.; Huang, J.; Kirkness, E.F.; Denisov, G.; et al. The diploid genome sequence of an individual human. PLoS Biol. 2007, 5, e254. [Google Scholar] [CrossRef] [PubMed]
- Miga, K.H. Completing the human genome: The progress and challenge of satellite DNA assembly. Chromosome Res. 2015, 23, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Meštrović, N.; Plohl, M.; Mravinac, B.; Ugarković, D. Evolution of satellite DNAs from the genus Palorus-experimental evidence for the “library” hypothesis. Mol. Biol. Evol. 1998, 15, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Mravinac, B.; Plohl, M.; Meštrović, N.; Ugarković, D. Sequence of PRAT satellite DNA “frozen” in some Coleopteran species. J. Mol. Evol. 2002, 54, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Mravinac, B.; Plohl, M.; Ugarković, D. Preservation and high sequence conservation of satellite DNAs suggest functional constraints. J. Mol. Evol. 2005, 61, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Ugarković, D.; Podnar, M.; Plohl, M. Satellite DNA of the red flour beetle Tribolium castaneum-comparative study of satellites from the genus Tribolium. Mol. Biol. Evol. 1996, 13, 1059–1066. [Google Scholar] [CrossRef] [PubMed]
- Feliciello, I.; Chinali, G.; Ugarković, Đ. Structure and evolutionary dynamics of the major satellite in the red flour beetle Tribolium castaneum. Genetica 2011, 139, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, L.; Venanzetti, F.; Sbordoni, V. Characterization of a species- specific satellite DNA family of Dolichopoda schiavazzii (Orthoptera, Rhaphidophoridae) cave crickets. J. Mol. Evol. 1994, 39, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Martinsen, L.; Venanzetti, F.; Johnsen, A.; Sbordoni, V.; Bachmann, L. Molecular evolution of the pDo500 satellite DNA family in Dolichopoda cave crickets (Rhaphidophoridae). BMC Evol. Biol. 2009, 9, 301. [Google Scholar] [CrossRef] [PubMed]
- Cafasso, D.; Chinali, G. An ancient satellite DNA has maintained repetitive units of the original structure in most species of the living fossil plant genus Zamia. Genome 2014, 57, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Navajas-Pérez, R.; de la Herrán, R.; Jamilena, M.; Lozano, R.; Ruiz Rejón, C.R.; Ruiz Rejón, M.; Garrido-Ramos, M.A. Reduced rates of sequence evolution of Y-linked satellite DNA in Rumex (Polygonaceae). J. Mol. Evol. 2005, 60, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Navajas-Pérez, R.; Quesada del Bosque, M.E.; Garrido-Ramos, M.A. Effect of location, organization and repeat-copy number in satellite-DNA evolution. Mol. Genet. Gen. 2009, 282, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Navajas-Pérez, R.; Schwarzacher, T.; Ruiz Rejón, M.; Garrido-Ramos, M.A. Characterization of RUSI, a telomere-associated satellite DNA, in the genus Rumex (Polygonaceae). Cytogenet. Genome Res. 2009, 124, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Navajas-Pérez, R.; Schwarzacher, T.; Ruiz Rejón, M.; Garrido-Ramos, M.A. Molecular cytogenetic characterization of Rumex papillaris, a dioecious plant with an XX/XY1Y2 sex chromosome system. Genetica 2009, 135, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ramos, M.A.; de la Herran, R.; Ruiz Rejón, M.; Ruiz Rejón, C. A satellite DNA of the Sparidae family (Pisces, Perciformes) associated with telomeric sequences. Cytogenet. Cell Genet. 1998, 83, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ramos, M.A.; Jamilena, M.; Lozano, R.; Ruiz Rejón, C.; Ruiz Rejón, M. The EcoRI centromeric satellite DNA of the Sparidae family (Pisces, Perciformes) contains a sequence motive common to other vertebrate centromeric satellite DNAs. Cytogenet. Cell. Genet. 1995, 71, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ramos, M.A.; de la Herran, R.; Jamilena, M.; Lozano, R.; Ruiz Rejón, C.; Ruiz Rejón, M. Evolution of centromeric satellite-DNA and its use in phylogenetic studies of the Sparidae family (Pisces, Perciformes). Mol. Phyl. Evol. 1999, 12, 200–204. [Google Scholar] [CrossRef] [PubMed]
- De la Herrán, R.; Ruiz Rejón, C.; Ruiz Rejón, M.; Garrido-Ramos, M.A. The molecular phylogeny of the Sparidae (Pisces, Perciformes) based on two satellite DNA families. Heredity 2001, 87, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Robles, F.; de la Herrán, R.; Ludwig, A.; Ruiz Rejón, C.; Ruiz Rejón, M.; Garrido-Ramos, M.A. Evolution of ancient satellite DNAs in sturgeon genomes. Gene 2004, 338, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Árnason, Ú.; Widegren, B. Pinniped phylogeny enlightened by molecular hybridizations using highly repetitive DNA. Mol. Biol. Evol. 1986, 3, 356–365. [Google Scholar]
- Arnason, U. Phylogeny of marine mammals-evidence from chromosomes and DNA. Chromosomes Today 1990, 10, 267–278. [Google Scholar]
- Arnason, U.; Grettarsdottir, S.; Widegren, B. Mysticete (baleen whale) relationships based upon the sequence of the common cetacean DNA satellite. Mol. Biol. Evol. 1992, 9, 1018–1028. [Google Scholar] [PubMed]
- Gretarsdottir, G.; Arnason, U. Evolution of the common cetacean highly repetitive DNA component and the systematic position of Orcaella brevirostris. J. Mol. Evol. 1992, 34, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Sujiwattanarat, P.; Thapana, W.; Srikulnath, K.; Hirai, Y.; Hirai, H.; Koga, A. Higher-order repeat structure in alpha satellite DNA occurs in New World monkeys and is not confined to hominoids. Sci. Rep. 2015, 5, 10315. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, S.; Goyal, V. Repetitive sequences in plant nuclear DNA: Types, Distribution, Evolution and Function. Genom. Proteom. Bioinform. 2014, 12, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Petraccioli, A.; Odierna, G.; Capriglione, T.; Barucca, M.; Forconi, M.; Olmo, E.; Biscotti, M.A. A novel satellite DNA isolated in Pecten jacobaeus shows high sequence similarity among mollusks. Mol. Genet. Genom. 2015, 290, 1717–1725. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ramos, M.A. Satellite DNA in Plants: More than Just Rubbish. Cytogenet. Genome Res. 2015, 146, 153–170. [Google Scholar] [CrossRef] [PubMed]
- Čížková, J.; Hřibová, A.; Humplíková, L.; Christelová, P.; Suchánková, P.; Doležel, J. Molecular analysis and genomic organization of major DNA satellites in banana (Musa spp.). PLoS ONE 2013, 8, e54808. [Google Scholar] [CrossRef] [PubMed]
- Hribová, E.; Neumann, P.; Matsumoto, T.; Roux, N.; Macas, J.; Dolezel, J. Repetitive part of the banana (Musa acuminata) genome investigated by low-depth 454 sequencing. BMC Plant Biol. 2010, 10, 204. [Google Scholar] [CrossRef] [PubMed]
- Macas, J.; Požárková, D.; Navrátilová, A.; Nouzová, M.; Neumann, P. Two new families of tandem repeats isolated from genus Vicia using genomic self-priming PCR. Mol. Gen. Genet. 2000, 263, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Ambrožová, K.; Mandáková, T.; Bureš, P.; Neumann, P.; Leitch, I.J.; Koblížková, A.; Macas, J.; Lysak, M.A. Diverse retrotransposon families and an AT-rich satellite DNA revealed in giant genomes of Fritillaria lilies. Ann. Bot. 2011, 107, 255–268. [Google Scholar] [CrossRef] [PubMed]
- De la Herrán, R.; Robles, F.; Cuñado, N.; Santos, J.L.; Ruiz Rejón, M.; Garrido-Ramos, M.A.; Ruiz Rejón, C. A heterochromatic satellite DNA is highly amplified in a single chromosome of Muscari (Hyacinthaceae). Chromosoma 2001, 110, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Emadzade, K.; Jang, T.S.; Macas, J.; Kovařík, A.; Novák, P.; Parker, J.; Weiss-Schneeweiss, H. Differential amplification of satellite PaB6 in chromosomally hypervariable Prospero autumnale complex (Hyacinthaceae). Ann. Bot. 2014, 114, 1597–1608. [Google Scholar] [CrossRef] [PubMed]
- Subirana, J.A.; Messeguer, X. A Satellite explosion in the genome of holocentric nematodes. PLoS ONE 2013, 8, e62221. [Google Scholar] [CrossRef] [PubMed]
- Subirana, J.A.; Albà, M.; Messeguer, X. High evolutionary turnover of satellite families in Caenorhabditis. BMC Evol. Biol. 2015, 15, 218. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ramos, M.A.; Jamilena, M.; Lozano, R.; Ruiz Rejón, C.; Ruiz Rejón, M. Cloning and characterization of a fish centromeric satellite DNA. Cytogenet. Cell Genet. 1994, 65, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Kopecna, O.; Kubickova, S.; Cernohorska, H.; Cabelova, K.; Vahala, J.; Martinkova, N.; Rubes, J. Tribe-specific satellite DNA in non-domestic Bovidae. Chromosome Res. 2014, 22, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Steflova, P.; Tokan, V.; Vogel, I.; Lexa, M.; Macas, J.; Novák, P.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Contrasting patterns of transposable element and satellite distribution on sex chromosomes (XY1Y2) in the dioecious plant Rumex acetosa. Genome Biol. Evol. 2013, 5, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.F. Highly rRepeated sequences in mammalian genomes. Int. Rev. Cytol. 1982, 76, 67–112. [Google Scholar] [PubMed]
- Kit, S. Equilibrium sedimentation in density gradients of DNA preparations from animal tissues. J. Mol. Biol. 1961, 3, 711–716. [Google Scholar] [CrossRef]
- Sueoka, N. Variation and heterogeneity of base composition of deoxyribonucleic acids: A compilation of old and new data. J. Mol. Biol. 1961, 3, 31–40. [Google Scholar] [CrossRef]
- Waring, M.; Britten, R.J. Nucleotide sequence repetition: A rapidly reassociating fraction of mouse DNA. Science 1966, 154, 791–794. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.J.; Kohne, D.E. Repeated sequences in DNA. Science 1968, 161, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.J.; Graham, D.E.; Neufeld, B.R. Analysis of repeating DNA sequences by reassociation. Methods Enzymol. 1974, 29, 363–405. [Google Scholar] [PubMed]
- Britten, R.J.; Davidson, E.H. Hybridisation strategy. In Nucleic Acid Hybridization; Hames, B.D., Higgins, S.J., Eds.; IRL Press: Washington, DC, USA, 1986; pp. 3–15. [Google Scholar]
- Peterson, D.G.; Schulze, S.R.; Sciara, E.B.; Lee, S.A.; Bowers, J.E.; Nagel, A.; Jiang, N.; Tibbitts, D.C.; Wessler, S.R.; Paterson, A.H. Integration of Cot Analysis, DNA Cloning, and High-Throughput sequencing facilitates genome characterization and gene discovery. Genome Res. 2002, 12, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Horz, W.; Zachau, H.G. Characterization of distinct segments in mouse satellite DNA by restriction nucleases. Eur. J. Biochem. 1977, 73, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ramos, M.A.; Jamilena, M.; Lozano, R.; Cárdenas, S.; Ruiz Rejón, C.; Ruiz Rejón, M. Phylogenetic relationships of the Sparidae family (Pisces, Perciformes) inferred from satellite DNA. Hereditas 1995, 122, 1–6. [Google Scholar] [CrossRef]
- Pardue, M.L.; Gall, J.G. Chromosomal localization of mouse satellite DNA. Science 1970, 168, 1356–1358. [Google Scholar] [CrossRef] [PubMed]
- Gall, J.G. The origin of in situ hybridization—A personal history. Methods 2016, 98, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Schwarzacher, T.; Heslop-Harrison, P. Practical In Situ Hybridization; BIOS Scientific Publishers Ltd.: Oxford, UK, 2000. [Google Scholar]
- Feliciello, I.; Picariello, O.; Chinali, G. The first characterisation of the overall variability of repetitive units in a species reveals unexpected features of satellite DNA. Gene 2005, 349, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Feliciello, I.; Picariello, O.; Chinali, G. Intra-specific variability and unusual organization of the repetitive units in a satellite DNA from Rana dalmatina: Molecular evidence of a new mechanism of DNA repair acting on satellite DNA. Gene 2006, 383, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Quesada del Bosque, M.E.; López-Flores, I.; Suárez-Santiago, V.N.; Garrido-Ramos, M.A. Differential spreading of HinfI satellite DNA variants during radiation in Centaureinae. Ann. Bot. 2013, 112, 1793–1802. [Google Scholar] [CrossRef] [PubMed]
- Plohl, M.; Luchetti, A.; Meštrović, N.; Mantovani, B. Satellite DNAs between selfishness and functionality: Structure, genomics and evolution of tandem repeats in centromeric (hetero)chromatin. Gene 2008, 409, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Plohl, M.; Meštrović, N.; Mravinac, B. Satellite DNA evolution. Genome Dyn. 2012, 7, 126–152. [Google Scholar] [PubMed]
- Plohl, M.; Meštrović, N.; Mravinac, B. Centromere identity from the DNA point of view. Chromosoma 2014, 123, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Pezer, Z.; Brajković, J.; Feliciello, I.; Ugarković, Đ. Satellite DNA-Mediated Effects on Genome Regulation. Genome Dyn. 2012, 7, 153–169. [Google Scholar] [PubMed]
- Weiss-Schneeweiss, H.; Leitch, A.R.; McCann, J.; Jang, T.S.; Macas, J. Employing next generation sequencing to explore the repeat landscape of the plant genome. In Next Generation Sequencing in Plant Systematics Regnum Vegetabile; Hörandl, E., Appelhans, M., Eds.; Koeltz Scientific Books: Königstein, Germany, 2015; pp. 155–179. [Google Scholar]
- Novák, P.; Neumann, P.; Macas, J. Graph-based clustering and characterization of repetitive sequences in next-generation sequencing data. BMC Bioinform. 2010, 11, 378. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Neumann, P.; Pech, J.; Steinhaisl, J.; Macas, J. RepeatExplorer: A Galaxy-based web server for genome-wide characterization of eukaryotic repetitive elements from next generation sequence reads. Bioinformatics 2013, 29, 792–793. [Google Scholar] [CrossRef] [PubMed]
- Pagan, H.J.T.; Macas, J.; Novak, P.; McCulloch, E.S.; Stevens, R.D.; Ray, D.A. Survey sequencing reveals elevated DNA transposon activity, novel elements, and variation in repetitive landscapes among bats. Genome Biol. Evol. 2012, 4, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Melters, D.P.; Bradnam, K.R.; Young, H.A.; Telis, N.; May, M.R.; Ruby, J.G.; Sebra, R.; Peluso, P.; Eid, J.; Rank, D.; et al. Comparative analysis of tandem repeats from hundreds of species reveals unique insights into centromere evolution. Genome Biol. 2013, 14, R10. [Google Scholar] [CrossRef] [PubMed]
- Camacho, J.P.M.; Ruiz-Ruano, F.J.; Martín-Blázquez, R.; López-León, M.D.; Cabrero, J.; Lorite, P.; Cabral-de-Mello, D.C.; Bakkali, M. A step to the gigantic genome of the desert locust: Chromosome sizes and repeated DNAs. Chromosoma 2015, 124, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Hribova, E.; Neumann, P.; Koblizkova, A.; Dolezel, J.; Macas, J. Genome-wide analysis of repeat diversity across the family Musaceae. PLoS ONE 2014, 9, e98918. [Google Scholar] [CrossRef] [PubMed]
- Pita, S.; Panzera, F.; Mora, P.; Vela, J.; Cuadrado, Á.; Sánchez, A.; Palomeque, T.; Lorite, P. Comparative repeatome analysis on Triatoma infestans Andean and Non-Andean lineages, main vector of Chagas disease. PLoS ONE 2017, 12, e0181635. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Ávila Robledillo, P.; Koblížková, A.; Vrbová, I.; Neumann, P.; Macas, J. TAREAN: A computational tool for identification and characterization of satellite DNA from unassembled short reads. Nucleic Acids Res. 2017, 45, e111. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.B.; Oh, J.H.; McIver, L.J.; Rashkovetsky, E.; Michalak, K.; Garner, H.R.; Kang, L.; Nevo, E.; Korol, A.B.; Michalak, P. Divergence of Drosophila melanogaster repeatomes in response to a sharp microclimate contrast in Evolution Canyon, Israel. Proc. Natl. Acad. Sci. USA 2014, 111, 10630–10635. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.G.; Kwoh, C.K.; Hsu, Y.L.; Wirawan, A. Review of tandemrepeat search tools: A systematic approach to evaluating algorithmic performance. Brief. Bioinform. 2012, 14, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Monjo, T.; Hoang, P.H.; Yoshimura, J.; Yurino, H.; Mitsui, J.; Ishiura, H.; Takahashi, Y.; Ichikawa, Y.; Goto, J.; et al. Rapid detection of expanded short tandem repeats in personal genomics using hybrid sequencing. Bioinformatics 2014, 30, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Pavlek, M.; Gelfand, Y.; Plohl, M.; Meštrović, N. Genome wide analysis of tandem repeats in Tribolium castaneum genome reveals abundant and highly dynamic tandem repeat families with satellite DNA features in euchromatic chromosomal arms. DNA Res. 2015, 22, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Gelfand, Y.; Rodriguez, A.; Benson, G. TRDB—The Tandem Repeats Database. Nucleic Acids Res. 2006, 35, D80–D87. [Google Scholar] [CrossRef] [PubMed]
- Pech, M.; Igo-Kemenes, T.; Zachau, H.G. Nucleotide sequence of a highly repetitive component of rat DNA. Nucleic Acids Res. 1979, 7, 417–432. [Google Scholar] [CrossRef] [PubMed]
- De la Herrán, R.; Fontana, F.; Lanfredi, M.; Congiu, L.; Leis, M.; Rossi, R.; Ruiz Rejón, C.; Ruiz Rejón, M.; Garrido-Ramos, M.A. Slow rates of evolution and sequence homogenization in an ancient satellite DNA family of sturgeons. Mol. Biol. Evol. 2001, 18, 432–436. [Google Scholar] [CrossRef] [PubMed]
- De la Herrán, R.; Robles, F.; Navas, J.I.; López-Flores, I.; Herrera, M.; Hachero, I.; Garrido-Ramos, M.A.; Ruiz Rejón, C.; Ruiz Rejón, M. The centromeric satellite of the wedge sole (Dicologoglossa cuneata, Pleuronectiformes) is composed mainly of a sequence motif conserved in other vertebrate centromeric DNAs. Cytogenet. Genome Res. 2008, 121, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Macas, J.; Navrátilová, A.; Koblížková, A. Sequence homogenization and chromosomal localization of VicTR-B satellites differ between closely related Vicia species. Chromosoma 2006, 115, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Stupar, R.M.; Song, J.; Tek, A.L.; Cheng, Z.; Dong, F.; Jiang, J. Highly condensed potato pericentromeric heterochromatin contains rDNA-related tandem repeats. Genetics 2002, 162, 1435–1444. [Google Scholar] [PubMed]
- Willard, H.F.; Waye, J.S. Hierarchical order in chromosome-specific human alpha satellite DNA. Trends Genet. 1987, 3, 192–198. [Google Scholar] [CrossRef]
- Warburton, P.E.; Willard, H.F. Genomic analysis of sequence variation in tandemly repeated DNA. Evidence for localized homogeneous sequence domains within arrays of alpha-satellite DNA. J. Mol. Biol. 1990, 216, 3–16. [Google Scholar] [CrossRef]
- Miga, K.H.; Newton, Y.; Jain, M.; Altemose, N.; Willard, H.F.; Kent, W.J. Centromere reference models for human chromosomes X and Y satellite arrays. Genome Res. 2014, 24, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Schueler, M.G.; Higgins, A.W.; Rudd, M.K.; Gustashaw, K.; Willard, H.F. Genomic and genetic definition of a functional human centromere. Science 2001, 294, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Rudd, M.K.; Wray, G.A.; Willard, H.F. The evolutionary dynamics of alpha-satellite. Genome Res. 2006, 16, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Cacheux, L.; Ponger, L.; Gerbault-Seureau, M.; Richard, F.A.; Escudé, C. Diversity and distribution of alpha satellite DNA in the genome of an Old World monkey: Cercopithecus solatus. BMC Genom. 2016, 17, 916. [Google Scholar] [CrossRef] [PubMed]
- Shepelev, V.A.; Alexandrov, A.A.; Yurov, Y.B.; Alexandrov, I.A. The evolutionary origin of man can be traced in the layers of defunct ancestral alpha satellites flanking the active centromeres of human chromosome. PLoS Genet. 2009, 5, e1000641. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, I.A.; Mitkevich, S.P.; Yurov, Y.B. The phylogeny of human chromosome specific alpha satellites. Chromosoma 1988, 96, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Cellamare, A.; Catacchio, C.R.; Alkan, C.; Giannuzzi, G.; Antonacci, F.; Cardone, M.F.; Della Valle, G.; Malig, M.; Rocchi, M.; Eichler, E.E.; et al. New insights into centromere organization and evolution from the white-cheeked gibbon and marmoset. Mol. Biol. Evol. 2009, 26, 1889–1900. [Google Scholar] [CrossRef] [PubMed]
- Terada, S.; Hirai, Y.; Hirai, H.; Koga, A. Higher-order repeat structure in alpha satellite DNA is an attribute of hominoids rather than hominids. J. Hum. Genet. 2013, 58, 752–754. [Google Scholar] [CrossRef] [PubMed]
- Koga, A.; Hirai, Y.; Terada, S.; Jahan, I.; Baicharoen, S.; Arsaithamkul, V.; Hirai, H. Evolutionary Origin of Higher-Order Repeat Structure in Alpha-Satellite DNA of Primate Centromeres. DNA Res. 2014, 21, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Pike, L.M.; Carlisle, A.; Newell, C.; Hong, S.B.; Musich, P.R. Sequence and evolution of rhesus monkey alphoid DNA. J. Mol. Evol. 1986, 23, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Alkan, C.; Ventura, M.; Archidiacono, N.; Rocchi, M.; Sahinalp, S.C.; Eichler, E.E. Organization and evolution of primate centromeric DNA from whole-genome shotgun sequence data. PLoS Comput. Biol. 2007, 3, 1807–1818. [Google Scholar] [CrossRef] [PubMed]
- Alves, G.; Seuánez, H.N.; Fanning, T. Alpha satellite DNA in neotropical primates (Platyrrhini). Chromosoma 1994, 103, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Palomeque, T.; Lorite, P. Satellite DNA in insects—A review. Heredity 2008, 100, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Vlahović, I.; Glunčić, M.; Rosandić, M.; Ugarković, Đ.; Paar, V. Regular higher order repeat structures in beetle Tribolium castaneum genome. Genome Biol. Evol. 2016, evw174. [Google Scholar] [CrossRef] [PubMed]
- Meštrović, N.; Mravinac, B.; Pavlek, M.; Vojvoda-Zeljko, T.; Šatović, E.; Plohl, M. Structural and functional liaisons between transposable elements and satellite DNAs. Chromosome Res. 2015, 23, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Plohl, M.; Cornudella, L. Characterization of a complex satellite DNA in the mollusc Donax trunculus: Analysis of sequence variations and divergence. Gene 1996, 169, 157–164. [Google Scholar] [CrossRef]
- López-Flores, I.; de la Herrán, R.; Garrido-Ramos, M.A.; Boudry, P.; Ruiz-Rejón, C.; Ruiz-Rejón, M. The molecular phylogeny of oysters based on a satellite DNA related to transposons. Gene 2004, 339, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Plohl, M.; Petrovic, V.; Luchetti, A.; Ricci, A.; Satovic, E.; Passamonti, M.; Mantovani, B. Long-term conservation vs high sequence divergence: The case of an extraordinarily old satellite DNA in bivalve mollusks. Heredity 2010, 104, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Macas, J.; Koblizkova, A.; Navratilova, A.; Neumann, P. Hypervariable 3′ UTR region of plant LTR-retrotransposons as a source of novel satellite repeats. Gene 2009, 448, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Šatović, E.; Plohl, M. Tandem repeat-containing MITEs in the clam Donax trunculus. Genome Biol. Evol. 2013, 5, 2549–2559. [Google Scholar] [CrossRef] [PubMed]
- Satović, E.; Zeljko, T.V.; Luchetti, A.; Mantovani, B.; Plohl, M. Adjacent sequences disclose potential for intra-genomic dispersal of satellite DNA repeats and suggest a complex network with transposable elements. BMC Genom. 2016, 17, 997. [Google Scholar] [CrossRef] [PubMed]
- Schueler, M.G.; Dunn, J.M.; Bird, C.P.; Ross, M.T.; Viggiano, L.; NISC Comparative Sequencing Program; Rocchi, M.; Willard, H.F.; Green, E.D. Progressive proximal expansion of the primate X chromosome centromere. Proc. Natl. Acad. Sci. USA 2005, 102, 10563–10568. [Google Scholar] [CrossRef] [PubMed]
- Neumann, P.; Navrátilová, A.; Koblížková, A.; Kejnovský, E.; Hřibová, E.; Hobza, R.; Widmer, A.; Doležel, J.; Macas, J. Plant centromeric retrotransposons: A structural and cytogenetic perspective. Mob. DNA 2011, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Zhang, W.; Yang, Q.; Zhang, Y.; Han, B.; Gu, M.; Xue, Y.; Cheng, Z. Diversity of centromeric repeats in two closely related wild rice species, Oryza officinalis and Oryza rhizomatis. Mol. Genet. Genom. 2006, 275, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Choulet, F.; Heng, Y.; Hao, W.; Paux, P.; Liu, Z.; Yue, W.; Jin, W.; Feuillet, C.; Zhang, X. Wheat centromeric retrotransposons: The new ones take a major role in centromeric structure. Plant J. 2013, 73, 952–965. [Google Scholar] [CrossRef] [PubMed]
- Nagaki, K.; Neumann, P.; Zhang, D.F.; Ouyang, S.; Buell, C.R.; Cheng, Z.K.; Jiang, J.M. Structure, divergence, and distribution of the CRR centromeric retrotransposon family in rice. Mol. Biol. Evol. 2005, 22, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Khost, D.E.; Eickbush, D.G.; Larracuente, A.M. Single-molecule sequencing resolves the detailed structure of complex satellite DNA loci in Drosophila melanogaster. Genome Res. 2017, 27, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Carmona, A.; Friero, E.; de Bustos, A.; Jouve, N.; Cuadrado, A. Cytogenetic diversity of SSR motifs within and between Hordeum species carrying the H genome: H. vulgare L. and H. bulbosum L. Theor. Appl. Genet. 2013, 126, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, A.; Carmona, A.; Jouve, N. Chromosomal characterization of the three subgenomes in the polyploids of Hordeum murinum L.: New insight into the evolution of this complex. PLoS ONE 2013, 8, e81385. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, A.; Jouve, N. The nonrandom distribution of long clusters of all possible classes of trinucleotide repeats in barley chromosomes. Chromosome Res. 2007, 15, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Ananiev, E.V.; Chamberlin, M.A.; Klaiber, J.; Svitashev, S. Microsatellite megatracts in the maize (Zea mays L.) genome. Genome 2005, 48, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Prosser, J.; Frommert, M.; Paul, C.; Vincent, P.C. Sequence relationships of three human satellite DNAs. J. Mol. Riol. 1986, 187, 145–155. [Google Scholar] [CrossRef]
- Kuhn, G.S.C.; Küttler, H.; Moreira-Filho, O.; Heslop-Harrison, J.S. The 1.688 Repetitive DNA of Drosophila: Concerted Evolution at Different Genomic Scales and Association with Genes. Mol. Biol. Evol. 2012, 29, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Brajković, J.; Feliciello, I.; Bruvo-Mađarić, B.; Ugarković, Ð. Satellite DNA-like elements associated with genes within euchromatin of the beetle Tribolium castaneum. G3 Genes Genomes Genet. 2012, 2, 931. [Google Scholar] [CrossRef] [PubMed]
- Larracuente, A.M. The organization and evolution of the Responder satellite in species of the Drosophila melanogaster group: Dynamic evolution of a target of meiotic drive. BMC Evol. Biol. 2014, 14, 233. [Google Scholar] [CrossRef] [PubMed]
- Feliciello, I.; Akrap, I.; Ugarković, Đ. Satellite DNA Modulates Gene Expression in the Beetle Tribolium castaneum after Heat Stress. PLoS Genet 2015, 11, e1005466. [Google Scholar]
- De Lima, L.G.; Svartman, M.; Kuhn, G.C.S. Dissecting the satellite DNA landscape in three cactophilic Drosophila sequenced genomes. G3 Genes Genomes Genet. 2017. [Google Scholar] [CrossRef] [PubMed]
- John, B.; King, M.; Schweizer, D.; Mendelak, M. Equilocality of heterochromatin distribution and heterochromatin heterogeneity in acridid grasshoppers. Chromosoma 1985, 91, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Guenatri, M.; Bailly, D.; Maison, C.; Almouzni, G. Mouse centric and pericentric satellite repeats form distinct functional heterochromatin. J. Cell. Biol. 2004, 166, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, D.; Loidl, J. A model for heterochromatin dispersion and the evolution of C-band patterns. In Chromosomes Today; Hayman, D.L., Rofe, R.H., Sharp, P.J., Eds.; Allen and Unwin: London, UK, 1987; Volume 9, pp. 61–74. [Google Scholar]
- Cuadrado, A.; Jouve, N. Evolutionary trends of different repetitive DNA sequences during speciation in the genus Secale. J. Hered. 2002, 93, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Jantsch, M.; Hamilton, B.; Mayr, B.; Schweizer, D. Meiotic chromosome behaviour reflects levels of sequence divergence in Sus scrofa domestica satellite DNA. Chromosoma 1990, 99, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D. Plant sex determination and sex chromosomes. Heredity 2002, 88, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Filatov, D.A.; Moneger, F.; Negrutiu, I.; Charlesworth, D. Low variability in a Y-linked plant gene and its implications for Y-chromosome evolution. Nature 2000, 404, 388–390. [Google Scholar] [CrossRef] [PubMed]
- Steinemann, M.; Steinemann, S. The enigma of the Y chromosome degeneration: TRAM, a novel retrotransposon is preferentially located on the neo-Y chromosome of Drosophila miranda. Genetics 1997, 145, 261–266. [Google Scholar] [PubMed]
- Skaletsky, H.; Kuroda-Kawaguchi, T.; Minx, P.J.; Cordum, H.S.; Hillier, L.; Brown, L.G.; Repping, S.; Pyntikova, T.; Ali, J.; Bieri, J.; et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 2003, 423, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Hobza, R.; Kubat, Z.; Cegan, R.; Jesionek, W.; Vyskot, B.; Kejnovsky, E. Impact of repetitive DNA on sex chromosome evolution in plants. Chromosome Res. 2015, 23, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Vyskot, B.; Hobza, R. The genomics of plant sex chromosomes. Plant Sci. 2015, 236, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Blackmon, H.; Ross, L.; Bachtrog, D. Sex determination, sex chromosomes, and karyotype evolution in insects. J. Hered. 2017, 108, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Gimenez, O.M.; Dias, G.B.; de Lima, L.G.; Kuhn, G.C.E.S.; Ramos, É.; Martins, C.; Cabral-de-Mello, D.C. High-throughput analysis of the satellitome revealed enormous diversity of satellite DNAs in the neo-Y chromosome of the cricket Eneoptera surinamensis. Sci. Rep. 2017, 7, 6422. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D. Plant Sex Chromosomes. Annu. Rev. Plant Biol. 2016, 67, 397–420. [Google Scholar] [CrossRef] [PubMed]
- Shibata, F.; Hizume, M.; Kurori, Y. Chromosome painting of Y chromosomes and isolation of a Y chromosome-specific repetitive sequence in the dioecious plant Rumex acetosa. Chromosoma 1999, 108, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Shibata, F.; Hizume, M.; Kurori, Y. Differentiation and the polymorphic nature of the Y chromosomes revealed by repetitive sequences in the dioecious plant, Rumex acetosa. Chromosome Res. 2000, 8, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Navajas-Pérez, R.; Schwarzacher, T.; de la Herrán, R.; Ruiz Rejón, C.; Ruiz Rejón, M.; Garrido-Ramos, M.A. The origin and evolution of the variability in a Y-specific satellite-DNA of Rumex acetosa and its relatives. Gene 2006, 368, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Cuñado, N.; Navajas-Pérez, R.; de la Herrán, R.; Ruiz Rejón, C.; Ruiz Rejón, M.; Santos, J.L.; Garrido-Ramos, M.A. The evolution of sex chromosomes in the genus Rumex (Polygonaceae): Identification of a new species with heteromorphic sex chromosomes. Chromosome Res. 2007, 15, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, B.; Navajas-Pérez, R.; Lozano, R.; Parker, J.S.; de la Herrán, R.; Ruiz Rejón, C.; Ruiz Rejón, M.; Garrido-Ramos, M.A.; Jamilena, M. Cloning and characterization of dispersed repetitive DNA derived from microdissected sex chromosomes of Rumex acetosa. Genome 2006, 49, 114–121. [Google Scholar] [PubMed]
- Mariotti, B.; Manzano, S.; Kejnovský, E.; Vyskot, B.; Jamilena, M. Accumulation of Y-specific satellite DNAs during the evolution of Rumex acetosa sex chromosomes. Mol. Genet. Genom. 2009, 281, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Kejnovský, E.; Michalovova, M.; Steflova, P.; Kejnovska, I.; Manzano, S.; Hobza, R.; Kubat, Z.; Kovarik, J.; Jamilena, M.; Vyskot, B. Expansion of microsatellites on evolutionary young Y chromosome. PLoS ONE 2013, 8, e45519. [Google Scholar] [CrossRef] [PubMed]
- Kubat, Z.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Microsatellite accumulation on the Y chromosome in Silene latifolia. Genome 2008, 51, 350–356. [Google Scholar] [PubMed]
- Hobza, R.; Lengerova, M.; Svoboda, J.; Kubekova, H.; Kejnovsky, E.; Vyskot, B. An accumulation of tandem DNA repeats on the Y chromosome in Silene latifolia during early stages of sex chromosome evolution. Chromosoma 2006, 115, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Macas, J.; Kejnovský, E.; Neumann, P.; Novák, P.; Koblížková, A.; Vyskot, B. Next generation sequencing-based analysis of repetitive DNA in the model dioceous plant Silene latifolia. PLoS ONE 2011, 6, e27335. [Google Scholar] [CrossRef]
- Hobza, R.; Kubat, Z.; Cegan, R.; Jesione, W.; Kejnovsky, E.; Vyskot, B. Sex chromosome evolution in plants—Insight into the Y chromosome formation. Submitted.
- Klemme, S.; Banaei-Moghaddam, A.M.; Macas, J.; Wicker, T.; Novák, P.; Houben, A. High-copy sequences reveal distinct evolution of the rye B chromosome. New Phytol. 2013, 199, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Camacho, J.P.M. B chromosomes. In The Evolution of the Genome; Gregory, T.R., Ed.; Elsevier: Burlington, NJ, USA, 2005; pp. 223–285. [Google Scholar]
- Perfectti, F.; Werren, J.H. The interspecific origin of B chromosomes: Experimental evidence. Evolution 2001, 55, 1069–1073. [Google Scholar] [CrossRef]
- Tosta, V.C.; Marthe, J.B.; Tavares, M.G.; Fernandes-Salomão, T.M.; Pompolo, S.G.; Recco-Pimentel, S.M.; Perfectti, F.; Campos, L.A.; Camacho, J.P. Possible introgression of B chromosomes between bee species (genus Partamona). Cytogenet. Genome Res. 2014, 144, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ruano, F.J.; Cabrero, J.; López-León, M.D.; Camacho, J.P.M. Satellite DNA content illuminates the ancestry of a supernumerary (B) chromosome. Chromosoma 2017, 126, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Banaei-Moghaddama, A.M.; Martis, M.M.; Macas, J.; Gundlach, H.; Himmelbach, A.; Altschmied, L.; Mayer, K.F.X.; Houben, A. Genes on B chromosomes: Old questions revisited with new tools. Biochim. Biophys. Acta 2015, 1849, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Carchilan, M.; Kumke, K.; Mikolajewski, S.; Houben, A. Rye B chromosomes are weakly transcribed and might alter the transcriptional activity of a chromosome sequences. Chromosoma 2009, 118, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Estevez, M.; Lopez-Leon, M.D.; Cabrero, J.; Camacho, J.P.M. B-chromosome ribosomal DNA is functional in the grasshopper Eyprepocnemis plorans. PLoS ONE 2012, 7, e36600. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, V.A.; Dementyeva, P.V.; Larkin, D.M.; O’Brien, P.C.; Perelman, P.L.; Yang, F.; Ferguson-Smith, M.A.; Graphodatsky, A.S. Transcription of a protein-coding gene on B chromosomes of the Siberian roe deer (Capreolus pygargus). BMC Biol. 2013, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Valente, G.T.; Conte, M.A.; Fantinatti, B.E.; Cabral-de-Mello, D.C.; Carvalho, R.F.; Vicari, M.R.; Kocher, T.D.; Martins, C. Origin and evolution of B chromosomes in the cichlid fish Astatotilapia latifasciata based on integrated genomic analyses. Mol. Biol. Evol. 2014, 31, 2061–2072. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.; Schmutzer, T.; Scholz, U.; Houben, A. Origin and evolution of B chromosomes—What did we learn from next generation sequencing data analysis? Submitted.
- Shibata, F.; Hizume, M.; Kurori, Y. Molecular cytogenetic analysis of supernumerary heterochromatic segments in Rumex acetosa. Genome 2000, 43, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Camacho, J.P.M.; Cabrero, J. New hypotheses about the origin of supernumerary chromosome segments in grasshoppers. Heredity 1987, 58, 341–343. [Google Scholar] [CrossRef]
- Lopez-Leon, M.D.; Cabrero, J.; Camacho, J.P.M. Male and female segregation distortion for heterochromatic supernumerary segments on the B8 chromosome of the grasshopper Chortippus Jacobsi. Chromosoma 1992, 101, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Uno, Y.; Srikulnath, K.; Seki, R.; Nishida, N.; Matsuda, Y. Molecular cloning and characterization of satellite DNA sequences from constitutive heterochromatin of the habu snake (Protobothrops flavoviridis, Viperidae) and the Burmese python (Python bivittatus, Pythonidae). Chromosoma 2015, 124, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Lanfredi, M.; Congiu, L.; Garrido-Ramos, M.A.; dela Herrán, M.; Leis, M.; Chicca, M.; Rossi, R.; Tagliavini, J.; Ruiz Rejón, C.; Ruiz Rejón, M.; et al. Chromosomal location and evolution of a satellite DNA family in seven sturgeons species. Chromosome Res. 2001, 9, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, G.C.S.; Franco, F.F.; Manfrin, M.H.; Moreira-Filho, O.; Sene, F.M. Low rates of homogenization of the DBC-150 satellite DNA family restricted to a single pair of microchromosomes in species from the Drosophila buzzatii cluster. Chromosome Res. 2007, 15, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Fry, K.; Salser, W. Nucleotide sequences of HS- alpha satellite DNA from kangaroo rat Dipodomys ordii and characterization of similar sequences in other rodents. Cell 1977, 12, 1069–1084. [Google Scholar] [CrossRef]
- Pons, J.; Bruvo, B.; Petitpierre, E.; Plohl, M.; Ugarkovic, D.; Juan, C. Complex structural features of satellite DNA sequences in the genus Pimelia (Coleoptera: Tenebrionidae): Random differential amplification from a common ‘satellite DNA library’. Heredity 2004, 92, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Cesari, M.; Luchetti, A.; Passamonti, M.; Scali, V.; Mantovani, B. Polymerase chain reaction amplification of the Bag320 satellite family reveals the ancestral library and past gene conversion events in Bacillus rossius (Insecta Phasmatodea). Gene 2003, 312, 289–295. [Google Scholar] [CrossRef]
- Quesada del Bosque, M.E.; Navajas-Pérez, R.; Panero, J.L.; Fernández-González, A.; Garrido-Ramos, M.A. A satellite DNA evolutionary analysis in the North American endemic dioecious plant Rumex hastatulus (Polygonaceae). Genome 2011, 54, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Samoluk, S.S.; Robledo, G.; Bertioli, D.; Seijo, J.G. Evolutionary dynamics of an AT-rich satellite DNA and its contribution to karyotype differentiation in wild diploid Arachis species. Mol. Genet. Genom. 2017, 292, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Agmon, N.; Yacobi, K.; Mislovati, M.; Segal, D. Evidence for rolling circle replication of tandem genes in Drosophila. Nucleic Acids Res. 2005, 33, 4519–4526. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Agmon, N.; Sobol, O.; Segal, D. Extrachromosomal circles of satellite repeats and 5S ribosomal DNA in human cells. Mob. DNA 2010, 1, 11. [Google Scholar] [CrossRef] [PubMed]
- Dias, G.B.; Svartman, M.; Delprat, A.; Ruiz, A.; Kuhn, G.C.S. Tetris is a foldback transposon that provided the building blocks for an emerging satellite DNA of Drosophila virilis. Genome Biol. Evol. 2014, 6, 1302–1313. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, A. terMITEs: Miniature inverted-repeat transposable elements (MITEs) in the termite genome (Blattodea: Termitoidae). Mol. Genet. Genom. 2015, 290, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Santiago, V.N.; Blanca, G.; Ruiz-Rejón, M.; Garrido-Ramos, M.A. Satellite-DNA evolutionary patterns under a complex evolutionay scenario: The case of Acrolophus subgroup (Centaurea L., Compositae) from the western Mediterranean. Gene 2007, 404, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Quesada del Bosque, M.E.; López-Flores, I.; Suárez-Santiago, V.N.; Garrido-Ramos, M.A. Satellite-DNA diversification and the evolution of major lineages in Cardueae (Carduoideae, Asteraceae). J. Plant Res. 2014, 127, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Raina, S.N.; Sharma, S.; Sasakuma, T.; Kishii, M.; Vaishnavi, S. Novel repeated DNA sequences in safflower (Carthamus tinctorius L.) (Asteraceae): Cloning, sequencing, and physical mapping by fluorescence in situ hybridization. J. Hered. 2005, 96, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Dover, G. Molecular drive: A cohesive mode of species evolution. Nature 1982, 299, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Dover, G. Molecular drive. Trends Genet. 2002, 18, 587–589. [Google Scholar] [CrossRef]
- Elder, J.F., Jr.; Turner, B.J. Concerted evolution at the population level: Pupfish HindIII satellite DNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 994–998. [Google Scholar] [CrossRef] [PubMed]
- Feliciello, I.; Akrap, I.; Brajković, J.; Zlatar, I.; Ugarković, Đ. Satellite DNA as a driver of population divergence in the red flour beetle Tribolium castaneum. Genome Biol. Evol. 2014, 7, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Gutiérrez, M.A.; Suárez-Santiago, V.N.; López-Flores, I.; Romero, A.T.; Garrido-Ramos, M.A. Concerted evolution of satellite DNA in Sarcocapnos: A matter of time. Plant Mol. Biol. 2012, 78, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Strachan, T.; Webb, D.; Dover, G. Transition stages of molecular drive in multiple-copy DNA families in Drosophila. EMBO J. 1985, 4, 1701–1708. [Google Scholar] [PubMed]
- Bachmann, L.; Sperlich, D. Gradual evolution of a specific satellite DNA family in Drosophila ambigua, D. tristis, and D. obscura. Mol. Biol. Evol. 1993, 10, 647–659. [Google Scholar] [PubMed]
- Navajas-Pérez, R.; Ruiz Rejón, M.; Garrido-Ramos, M.A.; Aznarte, J.L.; Rubio-Escudero, C. SatDNA Analyzer: A computing tool for satellite-DNA evolutionary analysis. Bioinformatics 2007, 23, 767–768. [Google Scholar] [CrossRef] [PubMed]
- Strachan, T.; Coen, E.; Webb, D.; Dover, G. Modes and rates of change of complex DNA families of Drosophila. J. Mol. Biol. 1982, 158, 37–54. [Google Scholar] [CrossRef]
- Luchetti, A.; Cesari, M.; Carrara, G.; Cavicchi, S.; Passamonti, M.; Scali, V.; Mantovani, B. Unisexualityand molecular drive: Bag320 sequence diversity in Bacillus taxa (Insecta Phasmatodea). J. Mol. Evol. 2003, 56, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, A.; Marini, M.; Mantovani, B. Non-concerted evolution of RET76 satellite DNA family in Reticulitermes taxa (Insecta, Isoptera). Genetica 2006, 128, 123–132. [Google Scholar]
- Lorite, P.; Muñoz-López, M.; Carrillo, J.A.; Sanllorente, O.; Vela, J.; Mora, P.; Tinaut, A.; Torres, M.I.; Palomeque, T. Concerted evolution, a slow process for ant satellite DNA: Study of the satellite DNA in the Aphaenogaster genus (Hymenoptera, Formicidae). Org. Divers Evol. 2017. [Google Scholar] [CrossRef]
- Ruiz Rejón, C.; Jamilena, M.; Garrido-Ramos, M.A.; Parker, J.S.; Ruiz Rejón, M. Cytogenetic and molecular analysis of the multiple sex chromosome system of Rumex acetosa. Heredity 1994, 72, 209–215. [Google Scholar] [CrossRef]
- McAllister, B.F.; Werren, J.H. Evolution of tandemly repeated sequences: What happens at the end of an array? J. Mol. Evol. 1999, 48, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Mravinac, B.; Plohl, M. Satellite DNA junctions identify the potential origin of new repetitive elements in the beetle Tribolium madens. Gene 2007, 394, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Plohl, M.; Ugarković, D. Analysis of divergence of Alphitobius diaperinus satellite DNA—Roles of recombination, replication slippage and gene conversion. Mol. Gen. Genet. 1994, 242, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Harrison, G.E.; Heslop-Harrison, J.S. Centromeric repetitive DNA sequences in the genus Brassica. Theor. Appl. Genet. 1995, 90, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Pons, J.; Petitpierre, E.; Juan, C. Evolutionary dynamics of satellite DNA family PIM357 in species of the genus Pimelia (Tenebrionidae, Coleoptera). Mol. Biol. Evol. 2002, 19, 1329–1340. [Google Scholar] [CrossRef] [PubMed]
- Pons, J.; Gillespie, R.G. Common origin of the satellite DNAs of the Hawaiian spiders of the genus Tetragnatha: Evolutionary constraints on the length and nucleotide composition of the repeats. Gene 2003, 313, 169–177. [Google Scholar] [CrossRef]
- Vasil’ev, V.P. Polyploidization by reticular speciation in Acipenseriform evolution: A working hypothesis. J. Appl. Ichthyol. 1999, 15, 29–31. [Google Scholar] [CrossRef]
- Lorite, P.; Carrillo, J.A.; Tinaut, A.; Palomeque, T. Evolutionary dynamics of satellite DNA in species of the genus Formica (Hymenoptera, Formicidae). Gene 2004, 332, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Malik, H.S.; Henikoff, S. Adaptive evolution of Cid, a centromere specific histone in Drosophila. Genetics 2001, 157, 1293–1298. [Google Scholar] [PubMed]
- Henikoff, S.; Ahmad, K.; Malik, H.S. The centromere paradox: Stable inheritance with rapidly evolving DNA. Science 2001, 293, 1098–1102. [Google Scholar] [CrossRef] [PubMed]
- Ferree, P.M.; Barbash, D.A. Species-specific heterochromatin prevents mitotic chromosome segregation to cause hybrid lethality in Drosophila. PLoS Biol. 2009, 7, e1000234. [Google Scholar] [CrossRef] [PubMed]
- Meštrović, N.; Castagnone-Sereno, P.; Plohl, M. Interplay of selective pressure and stochastic events directs evolution of the MEL172 satellite DNA library in root-knot nematodes. Mol. Biol. Evol. 2006, 23, 2316–2325. [Google Scholar] [CrossRef] [PubMed]
- Malik, H.S.; Henikoff, S. Major Evolutionary Transitions in Centromere Complexity. Cell 2009, 138, 1067–1082. [Google Scholar] [CrossRef] [PubMed]
- Mravinac, B.; Plohl, M. Parallelism in evolution of highly repetitive DNAs in sibling species. Mol. Biol. Evol. 2010, 27, 1857–1867. [Google Scholar] [CrossRef] [PubMed]
- Djupedal, I.; Kos-Braun, I.C.; Mosher, R.A.; Söderholm, N.; Simmer, F.; Hardcastle, T.J.; Fender, A.; Heidrich, N.; Kagansky, A.; Bayne, E.; et al. Analysis of small RNA in fission yeast; centromeric siRNAs are potentially generated through a structured RNA. EMBO J. 2009, 28, 3832–3844. [Google Scholar] [CrossRef] [PubMed]
- Haaf, T.; Mater, A.G.; Wienberg, J.; Ward, D.C. Presence and abundance of CENP-B box sequences in great ape subsets of primate-specific alpha-satellite DNA. J. Mol. Evol. 1995, 41, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Masumoto, H.; Masukata, H.; Muro, Y.; Nozaki, N.; Okazaki, T. A human centromere antigen (CENP-B) interacts with a short specific sequence in alphoid DNA, a human centromeric satellite. J. Cell Biol. 1989, 109, 1963–1973. [Google Scholar] [CrossRef] [PubMed]
- Muro, Y.; Masumoto, H.; Yoda, K.; Nozaki, N.; Ohashi, M.; Okazaki, T. Centromere protein B assembles human centromeric alpha-satellite DNA at the 17-bp sequence, CENP-B box. J. Cell Biol. 1992, 116, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Masumoto, H.; Nakano, M.; Ohzeki, J. The role of CENP-B and alpha-satellite DNA: De novo assembly and epigenetic maintenance of human centromeres. Chromosome Res. 2004, 12, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Bulazel, K.; Metcalfe, C.; Ferreri, G.C.; Yu, J.; Eldridge, M.D.; O’Neill, R.J. Cytogenetic and molecular evaluation of centromereassociated DNA sequences from a marsupial (Macropodidae: Macropus rufogriseus) X chromosome. Genetics 2006, 172, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Alkan, C.; Cardone, M.F.; Catacchio, C.R.; Antonacci, F.; O’Brien, S.J.; Ryder, O.A.; Purgato, S.; Zoli, M.; Della Valle, G.; Eichler, E.E.; et al. Genome-wide characterization of centromeric satellites from multiple mammalian genomes. Genome Res. 2011, 21, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Schueler, M.G.; Swanson, W.; Thomas, P.J. NISC Comparative Sequencing Program & Green, E.D. Adaptive evolution of foundation kinetochore proteins in primates. Mol. Biol. Evol. 2010, 27, 1585–1597. [Google Scholar] [PubMed]
- Fachinetti, D.; Han, J.S.; McMahon, M.A.; Ly, P.; Abdullah, A.; Wong, A.J.; Cleveland, D.W. DNA sequence-specific binding of CENP-B enhances the fidelity of human centromere function. Dev. Cell 2015, 33, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Kugou, K.; Hirai, H.; Masumoto, H.; Koga, A. Formation of functional CENP-B boxes at diverse locations in repeat units of centromeric DNA in New World monkeys. Sci. Rep. 2016, 6, 27833. [Google Scholar] [CrossRef] [PubMed]
- Graur, D.; Zheng, Y.; Price, N.; Azevedo, R.B.R.; Zufall, R.A.; Elhaik, E. On the immortality of television sets: “function” in the human genome according to the evolution-free gospel of ENCODE. Genome Biol. Evol. 2013, 5, 578–590. [Google Scholar] [CrossRef] [PubMed]
- Graur, D.; Zheng, Y.; Azevedo, R.B.R. An Evolutionary Classification of Genomic Function. Genome Biol. Evol. 2015, 7, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, W.F. Is junk DNA bunk? A critique of ENCODE. Proc. Natl. Acad. Sci. USA 2013, 110, 5294–5300. [Google Scholar] [CrossRef] [PubMed]
- Varley, J.M.; Macgregor, H.C.; Erba, H.P. Satellite DNA is transcribed on lampbrush chromosomes. Nature 1980, 283, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Varley, J.M.; Macgregor, H.C.; Nardi, I.; Andrews, C.; Erba, H.P. Cytological evidence of transcription of highly repeated DNA sequences during the lampbrush stage in Triturus cristatus carnifex. Chromosoma 1980, 80, 289–307. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.O.; Barsacchi-Pilone, G.; Mahon, K.A.; Gall, J.G. Transcripts from both DNA strands of a satellite DNA occur on lampbrush chromosome loops of the newt Notophthalmus. Cell 1981, 24, 649–659. [Google Scholar] [CrossRef]
- Gall, J.G.; Stephenson, E.C.; Erba, H.P.; Diaz, M.O.; Barsacchi-Pilone, G. Histone genes are located at the sphere loci of newt lampbrush chromosomes. Chromosoma 1981, 84, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Epstein, L.M.; Mahon, K.A.; Gall, J.G. Transcription of a satellite DNA in the newt. J. Cell Biol. 1986, 103, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Solovei, I.V.; Joffe, B.I.; Gaginskaya, E.R.; Macgregor, H.C. Transcription of lampbrush chromosomes of a centromerically localized highly repeated DNA in pigeon (Columba) relates to sequence arrangement. Chromosome Res. 1996, 4, 588–603. [Google Scholar] [CrossRef] [PubMed]
- Hori, T.; Suzuki, Y.; Solovei, I.; Saitoh, Y.; Hutchison, N.; Ikeda, J.E.; Macgregor, H.; Mizuno, S. Characterization of DNA sequences constituting the terminal heterochromatin of the chicken Z chromosome. Chromosome Res. 1996, 4, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Biscotti, M.A.; Canapa, A.; Forconi, M.; Olmo, E.; Barucca, M. Transcription of tandemly repetitive DNA: Functional roles. Chromosome Res. 2015, 23, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Grenfell, A.W.; Strzelecka, M.; Heald, R. Transcription brings the complex(ity) to the centromere. Cell Cycle 2017, 16, 235–236. [Google Scholar] [CrossRef] [PubMed]
- Verdaasdonk, J.S.; Bloom, K. Centromeres: Unique chromatin structures that drive chromosome segregation. Nat. Rev. Mol. Cell Biol. 2011, 12, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Fukagawa, T.; Earnshaw, W.C. The Centromere: Chromatin Foundation for the Kinetochore Machinery. Dev. Cell 2014, 30, 496–508. [Google Scholar] [CrossRef] [PubMed]
- Steiner, F.A.; Henikoff, S. Diversity in the organization of centromeric chromatin. Curr. Opin. Genet. Dev. 2015, 31, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Bierhoff, H.; Postepska-Igielska, A.; Grummt, I. Noisy silence: Non-coding RNA and heterochromatin formation at repetitive elements. Epigenetics 2013, 9, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Bersani, F.; Lee, E.; Kharchenko, P.V.; Xu, A.W.; Liu, M.; Xega, K.; MacKenzie, O.C.; Brannigan, B.W.; Wittner, B.S.; Jung, H.; et al. Pericentromeric satellite repeat expansions through RNA-derived DNA intermediates in cancer. Proc. Natl. Acad. Sci. USA 2015, 112, 15148–15153. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, X.; Jin, W. An overview of plant centromeres. J. Genet. Genom. 2009, 36, 529–537. [Google Scholar] [CrossRef]
- Neumann, P.; Navratilova, A.; Schroeder-Reiter, E.; Koblizkova, A.; Steinbauerova, V.; Chocholova, E.; Novak, P.; Wanner, G.; Macas, J. Stretching the rules: Monocentric chromosomes with multiple centromere domains. PLoS Genet. 2012, 8, e1002777. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Wu, Y.; Koblízková, A.; Torres, G.A.; Wang, K.; Iovene, M.; Neumann, P.; Zhang, W.; Novák, P.; Buell, C.R.; et al. Repeatless and repeat-based centromeres in potato: Implications for centromere evolution. Plant Cell 2012, 24, 3559–3574. [Google Scholar] [CrossRef] [PubMed]
- Iwata, A.; Tek, A.L.; Richard, M.M.S.; Abernathy, B.; Fonseca, A.; Schmutz, J.; Chen, N.W.G.; Thareau, V.; Magdelenat, G.; Li, Y.; et al. Identification and characterization of functional centromeres of the common bean. Plant J. 2013, 76, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Wade, C.M.; Giulotto, E.; Sigurdsson, S.; Zoli, M.; Gnerre, S.; Imsland, F.; Lear, T.L.; Adelson, D.L.; Bailey, E.; Bellone, R.R.; et al. Genome sequence, comparative analysis, and population genetics of the domestic horse. Science 2009, 326, 865–867. [Google Scholar] [CrossRef] [PubMed]
- Shang, W.H.; Hori, T.; Toyoda, A.; Kato, J.; Popendorf, K.; Sakakibara, Y.; Fujiyama, A.; Fukagawa, T. Chickens possess centromeres with both extended tandem repeats and short non-tandem-repetitive sequences. Genome Res. 2010, 20, 1219–1228. [Google Scholar] [CrossRef] [PubMed]
- Locke, D.P.; Hillier, L.W.; Warren, W.C.; Worley, K.C.; Nazareth, L.V.; Muzny, D.M.; Yang, S.P.; Wang, Z.; Chinwalla, A.T.; Minx, P.; et al. Comparative and demographic analysis of orangutan genomes. Nature 2011, 469, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, F.; Gamba, R.; Mazzagatti, A.; Piras, F.M.; Cappelletti, E.; Belloni, E.; Nergadze, S.G.; Raimondi, E.; Giulotto, E. The major horse satellite DNA family is associated with centromere competence. Mol. Cytogenet. 2016, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Hasson, D.; Cheung, F.; Warburton, P.E. A paucity of heterochromatin at functional human neocentromeres. Epigenet. Chromatin 2010, 3, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Talbertc, P.B.; Zhang, W.; Wua, Y.; Yang, Z.; Henikoff, J.G.; Henikoff, S.; Jianga, J. The CentO satellite confers translational and rotational phasing on cenH3 nucleosomes in rice centromeres. Proc. Natl. Acad. Sci. USA 2013, 110, E4875–E4883. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Fu, S.; Dong, Q.; Han, F.; Birchler, J.A. Inactivation of a centromere during the formation of a translocation in maize. Chromosome Res. 2011, 19, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Lv, Z.; Pang, J.; Liu, Y.; Guo, X.; Fu, S.; Li, J.; Dong, Q.; Wu, H.-J.; Gao, Z.; et al. Formation of a functional maize centromere after loss of centromeric sequences and gain of ectopic sequences. Plant Cell 2013, 25, 1979–1989. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Gao, Z.; Birchler, J.; Han, F. Dicentric chromosome formation and epigenetics of centromere formation in plants. J. Genet. Genom. 2012, 39, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Schueler, M.G.; Sullivan, B.A. Structural and functional dynamics of human centromeric chromatin. Annu. Rev. Genom. Hum. Genet. 2006, 7, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lee, H.R.; Koo, D.H.; Jiang, J. Epigenetic modification of centromeric chromatin: Hypomethylation of DNA sequences in the CENH3-associated chromatin in Arabidopsis thaliana and maize. Plant Cell 2008, 20, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Gent, J.I.; Dawe, R.K. RNA as a structural and regulatory component of the centromere. Annu. Rev. Genet. 2012, 46, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Chan, F.L.; Marshall, O.J.; Saffery, R.; Kim, B.W.; Earle, E.; Choo, K.H.; Wong, L.H. Active transcription and essential role of RNA polymerase II at the centromere during mitosis. Proc. Natl. Acad. Sci. USA 2012, 109, 1979–1984. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, J.H.; Jakubsche, J.N.; Martins, N.M.; Kagansky, A.; Nakano, M.; Kimura, H.; Kelly, D.A.; Turner, B.M.; Masumoto, H.; Larionov, V.; et al. Epigenetic engineering: Histone H3K9 acetylation is compatible with kinetochore structure and function. J. Cell Sci. 2012, 125, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Pezer, Z.; Ugarković, D. Role of non-coding RNA and heterochromatin in aneuploidy and cancer. Semin. Cancer Biol. 2008, 18, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Frescas, D.; Guardavaccaro, D.; Kuchay, S.M.; Kato, H.; Poleshko, A.; Basrur, V.; Elenitoba-Johnson, K.S.; Katz, R.A.; Pagano, M. KDM2A represses transcription of centromeric satellite repeats and maintains the heterochromatic state. Cell Cycle 2008, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.H.; Brettingham-Moore, K.H.; Chan, L.; Quach, J.M.; Anderson, M.A.; Northrop, E.L.; Hannan, R.; Saffery, R.; Shaw, M.L.; Williams, E.; et al. Centromere RNA is a key component for the assembly of nucleoproteins at the nucleolus and centromere. Genome Res. 2007, 17, 1146–1160. [Google Scholar] [CrossRef] [PubMed]
- Saffery, R.; Sumer, H.; Hassan, S.; Wong, L.H.; Craig, J.M.; Todokoro, K.; Anderson, M.; Stafford, A.; Choo, K.H. Transcription within a functional human centromere. Mol. Cell 2003, 12, 509–516. [Google Scholar] [CrossRef]
- Quénet, D.; Dalal, Y. A long non-coding RNA is required for targeting centromeric protein A to the human centromere. Elife 2014, 3, e03254. [Google Scholar] [CrossRef] [PubMed]
- Grenfell, A.W.; Heald, R.; Strzelecka, M. Mitotic noncoding RNA processing promotes kinetochore and spindle assembly in Xenopus. J. Cell Biol. 2016, 214, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Rošic, S.; Köhler, F.; Erhardt, S. Repetitive centromeric satellite RNA is essential for kinetochore formation and cell division. J. Cell Biol. 2014, 207, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-R.; Neumann, P.; Macas, J.; Jiang, J. Transcription and evolutionary dynamics of the centromeric satellite repeat CentO in rice. Mol. Biol. Evol. 2006, 23, 2505–2520. [Google Scholar] [CrossRef] [PubMed]
- Carone, D.M.; Longo, M.S.; Ferreri, G.C.; Hall, L.; Harris, M.; Shook, N.; Bulazel, K.V.; Carone, B.R.; Obergfell, C.; O’Neill, M.J.; et al. A new class of retroviral and satellite encoded small RNAs emanates from mammalian centromeres. Chromosoma 2009, 118, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Pezer, Z.; Ugarković, D. Transcription of pericentromeric heterochromatin in beetles—Satellite DNAs as active regulatory elements. Cytogenet. Genome Res. 2009, 124, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Bouzinba-Segard, H.; Guais, A.; Francastel, C. Accumulation of small murine minor satellite transcripts leads to impaired centromeric architecture and function. Proc. Natl. Acad. Sci. USA 2006, 103, 8709–8714. [Google Scholar] [CrossRef] [PubMed]
- Melters, D.P.; Paliulis, L.V.; Korf, I.F.; Chan, S.W. Holocentric chromosomes: Convergent evolution, meiotic adaptations, and genomic analysis. Chromosome Res. 2012, 20, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Cuacos, M.; Franklin, F.C.H.; Heckmann, S. Atypical centromeres in plants—What they can tell us. Front. Plant Sci. 2015, 6, 913. [Google Scholar] [CrossRef] [PubMed]
- Neumann, P.; Pavlíková, Z.; Koblížková, A.; Fuková, I.; Jedlicková, V.; Novák, P.; Macas, J. Centromeres off the hook: Massive changes in centromere size and structure following duplication of CenH3 gene in Fabeae species. Mol.Biol. Evol. 2015, 32, 1862–1879. [Google Scholar] [CrossRef] [PubMed]
- Neumann, P.; Schubert, V.; Fuková, I.; Manning, J.E.; Houben, A.; Macas, J. Epigenetic Histone Marks of Extended Meta-Polycentric Centromeres of Lathyrus and Pisum Chromosomes. Front. Plant Sci. 2016, 7, 234. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.; Ribeiro, T.; Neumann, P.; Macas, J.; Novák, P.; Schubert, V.; Pellino, M.; Fuchs, J.; Ma, W.; Kuhlmann, M.; et al. Holocentromeres in Rhynchospora are associated with genome-wide centromere-specific repeat arrays interspersed among euchromatin. Proc. Natl. Acad. Sci. USA 2015, 112, 13633–13638. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, T.; Marques, A.; Novák, P.; Schubert, V.; Vanzela, A.L.L.; Macas, J.; Houben, A.; Pedrosa-Harand, A. Centromeric and non-centromeric satellite DNA organization differs in holocentric Rhynchospora species. Chromosoma 2017, 126, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, S.; Macas, J.; Kumke, K.; Fuchs, J.; Schubert, V.; Ma, L.; Novak, P.; Neumann, P.; Taudien, S.; Platzer, M.; et al. The holocentric species Luzula elegans shows an interplay between centromere and large-scale genome organization. Plant J. 2013, 73, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Drinnenberg, I.A.; deYoung, D.; Henikoff, S.; Malik, H.S. Recurrent loss of CenH3 is associated with independent transitions to holocentricity in insects. Elife 2014, 3, e03676. [Google Scholar] [CrossRef] [PubMed]
- Gassmann, R.; Rechtsteiner, A.; Yuen, K.W.; Muroyama, A.; Egelhofer, T.; Gaydos, L.; Barron, F.; Maddox, P.; Essex, A.; Monen, J.; et al. An inverse relationship to germline transcription defines centromeric chromatin in C. elegans. Nature 2012, 484, 534–537. [Google Scholar] [CrossRef] [PubMed]
- Steiner, F.A.; Henikoff, S. Holocentromeres are dispersed point centromeres localized at transcription factor hotspots. Elife 2014, 3, e02025. [Google Scholar] [CrossRef] [PubMed]
- Zedek, F.; Bureš, P. CenH3 evolution reflects meiotic symmetry as predicted by the centromere drive model. Sci. Rep. 2016, 6, 33308. [Google Scholar] [CrossRef] [PubMed]
- Dawe, R.K.; Henikoff, S. Centromeres put epigenetics in the driver’s seat. Trends Biochem. Sci. 2006, 31, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Ugarković, Ð. Evolution of Alpha-Satellite DNA; eLS. John Wiley & Sons, Ltd.: Chichester, UK, 2013. [Google Scholar] [CrossRef]
- Talbert, P.B.; Bayes, J.J.; Henikoff, S. Evolution of centromeres and kinetochores: A two-part fugue. In The Kinetochore; De Wulf, P., Earnshaw, W.C., Eds.; Springer: Berlin, Germany, 2008; pp. 193–230. [Google Scholar]
- Zedek, F.; Bureš, P. Evidence for Centromere Drive in the Holocentric Chromosomes of Caenorhabditis. PLoS ONE 2012, 7, e30496. [Google Scholar] [CrossRef] [PubMed]
- Zedek, F.; Bureš, P. Absence of positive selection on CenH3 in Luzula suggests that holokinetic chromosomes may suppress centromere drive. Ann. Bot. 2016, 118, 1347–1352. [Google Scholar] [CrossRef] [PubMed]
- Finseth, F.R.; Dong, Y.; Saunders, A.; Fishman, L. Duplication and Adaptive Evolution of a Key Centromeric Protein in Mimulus, a Genus with Female Meiotic Drive. Mol. Biol. Evol. 2015, 32, 2694–2706. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.E.; Rogers, K. Phylogenetic Analysis of Fungal Centromere H3 Proteins. Genetics 2006, 174, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- Bureš, P.; Zedek, F.; Marková, M. Holocentric chromosomes. In Plant Genome Diversity. Vol. 2. Physical Structure of Plant Genomes; Wendel, J., Greilhuber, J., Doležel, J., Leitch, I.J., Eds.; Springer: Heidelberg, Germany, 2013; pp. 187–208. [Google Scholar]
- Martínez, P.; Blasco, M.A. Telomeric and extra-telomeric roles for telomerase and the telomere-binding proteins. Nat. Rev. Cancer 2011, 11, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, D.C.; Londoño-Vallejo, A. Telomere dynamics in mammals. Genome Dyn. 2012, 7, 29–45. [Google Scholar] [PubMed]
- Mravinac, B.; Meštrović, N.; Čavrak, V.V.; Plohl, M. TCAGG, an alternative telomeric sequence in insects. Chromosoma 2011, 120, 367–376. [Google Scholar] [CrossRef] [PubMed]
- De la Herrán, R.; Cuñado, N.; Navajas-Pérez, R.; Santos, J.L.; Ruiz Rejón, C.; Garrido-Ramos, M.A.; Ruiz Rejón, M. The controversial telomeres of lily plants. Cytogenet. Genome Res. 2005, 109, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Sýkorová, E.; Lim, K.Y.; Kunická, Z.; Chase, M.W.; Bennett, M.D.; Fajkus, J.; Leitch, A.R. Telomere variability in the monocotyledonous plant order Asparagales. Proc. R. Soc. Lond. B 2003, 270, 1893–1904. [Google Scholar] [CrossRef] [PubMed]
- Peška, V.; Fajkus, P.; Fojtová, M.; Dvořáčková, M.; Hapala, J.; Dvořáček, V.; Polanská, P.; Leitch, A.R.; Sýkorová, E.; Fajkus, J. Characterisation of an unusual telomere motif (TTTTTTAGGG)n in the plant Cestrum elegans (Solanaceae), a species with a large genome. Plant J. 2015, 82, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Fulnečková, J.; Sěvčíková, T.; Fajkus, J.; Lukešová, A.; Lukeš, M.; Vlček, C.; Lang, B.F.; Kim, E.; Eliáš, M.; Sýkorová, E. A broad phylogenetic survey unveils the diversity and evolution of telomeres in eukaryotes. Genome Biol. Evol. 2013, 5, 468–483. [Google Scholar] [CrossRef] [PubMed]
- Silva-Sousa, R.; López-Panadès, E.; Casacuberta, E. Drosophila telomeres: An example of co- evolution with transposable Elements. Genome Dyn. 2012, 7, 46–67. [Google Scholar] [PubMed]
- Sýkorová, E.; Fajkus, J.; Meznikova, M.; Lim, K.Y.; Neplechova, K.; Blattner, F.R.; Chase, M.W.; Leitch, A.R. Minisatellite telomeres occur in the family Alliaceae but are lost in Allium. Am. J. Bot. 2006, 93, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Pich, U.; Schubert, I. Terminal heterochromatin and alternative telomeric sequences in Allium cepa. Chromosome Res. 1998, 6, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Louis, E.J.; Vershinin, A.V. Chromosome ends: Different sequences may provide conserved functions. Bioessays 2005, 27, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Riethman, H.; Ambrosini, A.; Castaneda, C.; Finklestein, J.; Hu, X.L.; Mudunuri, U.; Paul, S.; Wei, J. Mapping and initial analysis of human subtelomeric sequence assemblies. Genome Res. 2004, 14, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Riethman, H.; Ambrosini, A.; Paul, S. Human subtelomere structure and variation. Chromosome Res. 2005, 13, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, A.; Jouve, N. Mapping and organization of highly-repeated DNA sequences by means of simultaneous and sequential FISH and C-banding in 6x-triticale. Chromosome Res. 1994, 2, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, A.; Jouve, N. Fluorescent in situ hybridization and C-banding analyses of highly repetitive DNA sequences in the heterochromatin of rye (Secale montanum Guss.) and wheat incorporating S. montanum chromosome segments. Genome 1995, 38, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Vershinin, A.V.; Heslop-Harrison, J.S. Comparative analysis of the nucleosomal structure of rye, wheat and their relatives. Plant Mol. Biol. 1998, 36, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Contento, A.; Heslop-Harrison, J.S.; Schwarzacher, T. Diversity of a major repetitive DNA sequence in diploid and polyploid Triticeae. Cytogenet. Genome Res. 2005, 109, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Torres, G.A.; Gong, Z.; Iovene, M.; Hirsch, C.D.; Buell, C.R.; Bryan, G.J.; Novák, P.; Macas, J.; Jiang, J. Organization and evolution of subtelomeric satellite repeats in the potato genome. GE Genes Genomes Genet. 2011, 1, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, T.; Dos Santos, K.G.; Richard, M.M.; Sévignac, M.; Thareau, V.; Geffroy, V.; Pedrosa-Harand, A. Evolutionary dynamics of satellite DNA repeats from Phaseolus beans. Protoplasma 2017, 254, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Richard, M.M.S.; Chen, N.W.G.; Thareau, V.; Pflieger, E.; Blanchet, S.; Pedrosa-Harand, A.; Iwata, A.; Chavarro, C.; Jackson, S.A.; Geffroy, V. The subtelomeric khipu satellite repeat from Phaseolus vulgaris: Lessons learned from the genome analysis of the Andean genotype G19833. Front. Plant Sci. 2013, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kazama, Y.; Sugiyama, R.; Suto, Y.; Uchida, W.; Kawano, S. The clustering of four subfamilies of satellite DNA at individual chromosome ends in Silene latifolia. Genome 2006, 49, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ramos, M.A.; de la Herrán, R.; Ruiz Rejón, M.; Ruiz Rejón, C. A subtelomeric satellite DNA family isolated from the genome of the dioecious plant Silene latifolia. Genome 1999, 42, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Sýkorová, E.; Cartagena, J.; Horáková, M.; Fukui, K.; Fajkus, J. Characterization of telomere-subtelomere junctions in Silene latifolia. Mol. Gen. Genom. 2003, 269, 13–20. [Google Scholar]
- Lim, K.Y.; Kovarik, A.; Matyášek, R.; Chase, M.W.; Knapp, S.; McCarthy, E.; Clarkson, J.J.; Leitch, A.R. Comparative genomics and repetitive sequence divergence in the species of diploid Nicotiana section Alatae. Plant J. 2006, 48, 907–919. [Google Scholar] [PubMed]
- He, L.; Liu, J.; Torres, G.A.; Zhang, H.; Jiang, J.; Xie, C. Interstitial telomeric repeats are enriched in the centromeres of chromosomes in Solanum species. Chromosome Res. 2013, 21, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Feuerbach, F.; Galy, V.; Trelles-Sticken, E.; Fromont-Racine, M.; Jacquier, A.; Gilson, E.; Olivo-Marin, J.C.; Scherthan, H.; Nehrbass, U. Nuclear architecture and spatial positioning help establish transcriptional states of telomeres in yeast. Nat. Cell Biol. 2002, 4, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H. Telomeres and telomerase: Their mechanisms of action and the effects of altering their functions. FEBS Lett. 2005, 579, 859–862. [Google Scholar] [CrossRef] [PubMed]
- Jain, D.; Hebden, A.K.; Nakamura, T.M.; Miller, K.M.; Cooper, J.P. HAATI survivors replace canonical telomeres with blocks of generic heterochromatin. Nature 2010, 467, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Lundblad, V. Telomere maintenance without telomerase. Oncogene 2002, 21, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, A.; Garcia, F.; Azzalin, C.; Giulotto, E.; Egozcue, J.; Ponsa, M.; Garcia, M. Distribution of intrachromosomal telomeric sequences (ITS) on Macaca fascicularis (Primates) chromosomes and their implication for chromosome evolution. Hum. Genet. 2002, 110, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric repeat containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Cusanelli, E.; Chartrand, P. Telomeric repeat-containing RNA TERRA: A noncoding RNA connecting telomere biology to genome integrity. Front. Genet. 2015, 6, 143. [Google Scholar] [CrossRef] [PubMed]
- Schoeftner, S.; Blasco, M.A. Developmentally regulated transcription of mammalian telomeres by DNA-dependent RNA polymerase II. Nat. Cell Biol. 2008, 10, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Norseen, J.; Wiedmer, A.; Riethman, H.; Lieberman, P.M. TERRA RNA binding to TRF2 facilitates heterochromatin formation and ORC recruitment at telomeres. Mol. Cell 2009, 35, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Arnoult, N.; VanBeneden, A.; Decottignies, A. Telomere length regulates TERRA levels through increased trimethylation of telomeric H3K9 and HP1 alpha. Nat. Struct. Mol. Biol. 2012, 19, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Luke, B.; Panza, A.; Redon, S.; Iglesias, N.; Li, Z.; Lingner, J. The Rat1p 5′ to 3′ exonuclease degrades telomeric repeat-containing RNA and promotes telomere elongation in Saccharomyces cerevisiae. Mol. Cell 2008, 32, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Vrbsky, J.; Akimcheva, S.; Watson, J.M.; Turner, T.L.; Daxinger, L.; Vyskot, B.; Aufsatz, W.; Riha, K. siRNA-mediated methylation of Arabidopsis telomeres. PLoS Genet. 2010, 6, e1000986. [Google Scholar] [CrossRef] [PubMed]
- Eymery, A.; Horard, B.; El Atifi-Borel, M.; Fourel, G.; Berger, F.; Vitte, A.L.; Van den Broeck, A.; Brambilla, E.; Fournier, A.; Callanan, M.; et al. A transcriptomic analysis of human centromeric and pericentric sequences in normal and tumor cells. Nucleic Acids Res. 2009, 37, 6340–6354. [Google Scholar] [CrossRef] [PubMed]
- Volpe, T.A.; Kidner, C.; Hall, I.M.; Teng, G.; Grewal, S.I.S.; Martienssen, R. Regulation of heterochromatic silencing and histone H3 lysine- 9 methylation by RNAi. Science 2002, 297, 1833–1837. [Google Scholar] [CrossRef] [PubMed]
- Holoch, D.; Moazed, D. RNA-mediated epigenetic regulation of gene expression. Nat. Rev. Genet. 2015, 16, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.L.; Straight, A.F. RNA-mediated regulation of heterochromatin. Curr. Opin. Cell Biol. 2017, 46, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, F.; Weisshaar, B.; Fuchs, J.; Bannack, E.; Minoche, A.E.; Dohm, J.C.; Himmelbauer, H.; Schmidt, T. Epigenetic profiling of heterochromatic satellite DNA. Chromosoma 2011, 120, 409–422. [Google Scholar] [CrossRef] [PubMed]
- May, B.P.; Lippman, Z.B.; Fang, Y.; Spector, D.L.; Martienssen, R.A. Differential regulation of strandspecific transcripts from Arabidopsis centromeric satellite repeat. PLoS Genet. 2005, 1, e79. [Google Scholar] [CrossRef] [PubMed]
- Fukagawa, T.; Nogami, M.; Yoshikawa, M.; Ikeno, M.; Okazaki, T.; Takami, Y.; Nakayama, T.; Oshimura, M. Dicer is essential for formation of the heterochromatin structure in vertebrate cells. Nat. Cell Biol. 2004, 6, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Grishok, A.; Tabara, H.; Mello, C.C. Genetic requirements for inheritance of RNAi in C. elegans. Science 2000, 287, 2494–2497. [Google Scholar] [CrossRef] [PubMed]
- Pal-Bhadra, M.; Leibovitch, B.; Gandhi, S.; Rao, M.; Bhadra, U.; Birchler, J.; Elgin, S. Heterochromatic silencing and HP1 localization in Drosophila are dependent on the RNAi machinery. Science 2004, 303, 669–672. [Google Scholar] [CrossRef] [PubMed]
- Kanellopoulou, C.; Muljo, S.; Kung, A.; Ganesan, S.; Drapkin, R.; Jenuwein, T.; Livingston, D.; Rajewsky, K. Dicer-deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing. Genes Dev. 2005, 19, 489. [Google Scholar] [CrossRef] [PubMed]
- Saksouk, N.; Simboeck, E.; Déjardin, J. Constitutive heterochromatin formation and transcription in mammals. Epigenet. Chromatin 2015, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Maida, Y.; Yasukawa, M.; Okamoto, N.; Ohka, S.; Kinoshita, K.; Totoki, Y.; Ito, T.K.; Minamino, T.; Nakamura, H.; Yamaguchi, S.; et al. Involvement of telomerase reverse transcriptase in heterochromatin maintenance. Mol. Cell Biol. 2014, 34, 1576–1593. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Pao, G.M.; Huynh, A.M.; Suh, H.; Tonnu, N.; Nederlof, P.M.; Gage, F.H.; Verma, I.M. BRCA1 tumour suppression occurs via heterochromatin-mediated silencing. Nature 2011, 477, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Ting, D.T.; Lipson, D.; Paul, S.; Brannigan, B.W.; Akhavanfard, S.; Coffman, E.J.; Contino, G.; Deshpande, V.; Iafrate, A.J.; Letovsky, S.; et al. Aberrant overexpression of satellite repeats in pancreatic and other epithelial cancers. Science 2011, 331, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.P.; Meles, S.; Escudeiro, A.; Mendes-da-Silva, A.; Adega, F.; Chaves, R. Satellite Noncoding RNAs: The emerging players in cells, cellular pathways and cancer. Chromosome Res. 2015, 23, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Ichida, K.; Suzuki, K.; Kakizawa, N.; Fukui, T.; Takayama, Y.; Watanabe, F.; Kato, T.; Saito, M.; Futsuhara, K.; Rikiyama, T.; et al. Overexpression of satellite alpha transcript in breast cancer patients to impair specific chromosomes. J. Clin. Oncol. 2017, 35. [Google Scholar] [CrossRef]
- Ichida, K.; Suzuki, K.; Takayama, Y.; Muto, Y.; Fukui, T.; Kato, T.; Saito, M.; Watanabe, F.; Kakizawa, N.; Imoto, H.; et al. Detection of satellite alpha transcript in sera, as a surrogate marker for the risk of development of multiple cancers in colorectal cancer patients. J. Clin. Oncol. 2015, 33. [Google Scholar] [CrossRef]
- Kakizawa, N.; Suzuki, K.; Yoshizawa, A.; Ichida, K.; Saito, M.; Futsuhara, K. Toshiki Rikiyama Satellite alpha transcripts as a biomarker to predict the development of breast cancer with bilateral or multiple primary malignancies. J. Clin. Oncol. 2017, 35. [Google Scholar] [CrossRef]
- Ugarković, Đ. Functional elements residing within satellite DNAs. EMBO Rep. 2005, 6, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Valgardsdottir, R.; Chiodi, I.; Giordano, M.; Cobianchi, F.; Riva, S.; Biamonti, G. Structural and functional characterization of noncoding repetitive RNAs transcribed in stressed human cells. Mol. Biol. Cell 2005, 16, 2597–2604. [Google Scholar] [CrossRef] [PubMed]
- Col, E.; Hoghoughi, N.; Dufour, S.; Penin, J.; Koskas, S.; Faure, V.; Ouzounova, M.; Hernandez-Vargash, H.; Reynoird, N.; Daujat, S.; et al. Bromodomain factors of BET family are new essential actors of pericentric heterochromatin transcriptional activation in response to heat shock. Sci. Rep. 2017, 7, 5418. [Google Scholar] [CrossRef] [PubMed]
- Jehan, Z.; Vallinayagam, S.; Tiwari, S.; Pradhan, S.; Singh, L.; Suresh, A.; Reddy, H.M.; Ahuja, Y.R.; Jesudasan, R.A. Novel noncoding RNA from human Y distal heterochromatic block (Yq12) generates testis specific chimeric CDC2L2. Genome Res. 2007, 17, 433–440. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, R.; Baumann, C.; Viveiros, M.M. ATRX contributes to epigenetic asymmetry and silencing of major satellite transcripts in the maternal genome of the mouse embryo. Development 2015, 142, 1806–1817. [Google Scholar] [CrossRef] [PubMed]
- Noh, K.M.; Maze, I.; Zhao, D.; Xiang, B.; Wenderski, W.; Lewis, P.W.; Shen, L.; Li, H.; Allis, C.D. ATRX tolerates activitydependent histone H3 methyl/phos switching to maintain repetitive element silencing in neurons. Proc. Natl. Acad. Sci. USA 2015, 112, 6820–6827. [Google Scholar] [CrossRef] [PubMed]
- Pezer, Z.; Ugarkovic, D. Satellite DNA-associated siRNAs as mediators of heat shock response in insects. RNA Biol. 2012, 9, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Ferree, P.M. Sex Differences: Satellite DNA Directs Male-Specific Gene Expression. Curr. Biol. 2017, 27, 1866. [Google Scholar] [CrossRef] [PubMed]
- Menon, D.U.; Coarfa, C.; Xiao, W.; Gunaratne, P.H.; Meller, V.H. siRNAs from an X-linked satellite repeat promote X-chromosome recognition in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2014, 111, 16460–16465. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.S.; Meller, V.H. Satellite Repeats Identify X Chromatin for Dosage Compensation in Drosophila melanogaster males. Curr. Biol. 2017, 27, 1393–1402. [Google Scholar] [CrossRef] [PubMed]
- Hall, L.L.; Byron, M.; Carone, D.M.; Whitfield, T.W.; Pouliot, G.P.; Fischer, A.; Jones, P.; Lawrence, J.B. Demethylated HSATII DNA and HSATII RNA foci sequester PRC1 and MeCP2 into cancer-specific nuclear bodies. Cell Reports 2017, 18, 2943–2956. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(A) After electrophoresis of restriction digested genomic DNA, a distinct prominent band (arrow) against the background smear might be unveiled apparent which contains single fragments corresponding to the individual monomeric members of a satellite DNA (satDNA) family. (B) Southern blot hybridization patterns using satDNA probes against genomic DNA digested with different restriction enzymes. The occurrence of a restriction site for a particular enzyme leads to the complete digestion of an array which leads to a band of monomeric units (type A digestion, [53,61]). However, it is common that part of the array remains as a ‘ladder’ of oligomers of the repeat unit since some units have lost the restriction site by mutation (a, b, c, d, e). On the contrary, mutation may lead to the appearance of occasional restriction sites in some repeats within the array, which is observed as a type B digestion (f, g, h) [53,61]. Patterns of undigested satDNA are found in lines i, j. (C). Ladder-like patterns of satDNA amplification by polymerase chain reaction (PCR). (D). Examples of repeat clusters of satDNA visualized in the form of graphs where nodes represent sequence reads and edges connect reads with sequence similarities as obtained using RepeatExplorer [74,75].
Figure 1.
(A) After electrophoresis of restriction digested genomic DNA, a distinct prominent band (arrow) against the background smear might be unveiled apparent which contains single fragments corresponding to the individual monomeric members of a satellite DNA (satDNA) family. (B) Southern blot hybridization patterns using satDNA probes against genomic DNA digested with different restriction enzymes. The occurrence of a restriction site for a particular enzyme leads to the complete digestion of an array which leads to a band of monomeric units (type A digestion, [53,61]). However, it is common that part of the array remains as a ‘ladder’ of oligomers of the repeat unit since some units have lost the restriction site by mutation (a, b, c, d, e). On the contrary, mutation may lead to the appearance of occasional restriction sites in some repeats within the array, which is observed as a type B digestion (f, g, h) [53,61]. Patterns of undigested satDNA are found in lines i, j. (C). Ladder-like patterns of satDNA amplification by polymerase chain reaction (PCR). (D). Examples of repeat clusters of satDNA visualized in the form of graphs where nodes represent sequence reads and edges connect reads with sequence similarities as obtained using RepeatExplorer [74,75].

Figure 2.
Fluorescent in situ hybridization (FISH) using satDNA probes. Probes were labeled with digoxigenin-dUTP and detected with antidigoxigenin FITC-conjugate. FITC (Fluorescein IsoTioCyanate) is a fluorochrome derivative of fluorescein which fluoresces yellowish-green. In these pictures, yellowish-green signals correspond to detected probes while chromosomes are counterstained with ethidium propide (red). (A) EcoRI satDNA is the main component of the centromeres of the chromosomes of the fish species of the Sparidae family. This picture shows the detection of the hybridization signals of EcoRI repeats on the centromeres of all the chromosomes of Diplodus bellotti [30]. (B) DraI satDNA is located in the subtelomeric region of the chromosomes of some sparid species [29]. This picture shows the detection of the hybridization signals of DraI repeats at the subtelomeric region of many of the chromosomes of Pagrus auriga. The chromosomes of this species are all acrocentric. Also look for the presence of some interstitial loci in some of its chromosomes. (C) RAE180 satDNA is highly amplified in the Y chromosomes of males (XY1Y2) of the dioecius plant species Rumex acetosa [25,26,28]. This picture shows the detection of the hybridization signals of RAE180 repeats which are widespread in the major part of the two Y chromosomes. Sex chromosomes are indicated.
Figure 2.
Fluorescent in situ hybridization (FISH) using satDNA probes. Probes were labeled with digoxigenin-dUTP and detected with antidigoxigenin FITC-conjugate. FITC (Fluorescein IsoTioCyanate) is a fluorochrome derivative of fluorescein which fluoresces yellowish-green. In these pictures, yellowish-green signals correspond to detected probes while chromosomes are counterstained with ethidium propide (red). (A) EcoRI satDNA is the main component of the centromeres of the chromosomes of the fish species of the Sparidae family. This picture shows the detection of the hybridization signals of EcoRI repeats on the centromeres of all the chromosomes of Diplodus bellotti [30]. (B) DraI satDNA is located in the subtelomeric region of the chromosomes of some sparid species [29]. This picture shows the detection of the hybridization signals of DraI repeats at the subtelomeric region of many of the chromosomes of Pagrus auriga. The chromosomes of this species are all acrocentric. Also look for the presence of some interstitial loci in some of its chromosomes. (C) RAE180 satDNA is highly amplified in the Y chromosomes of males (XY1Y2) of the dioecius plant species Rumex acetosa [25,26,28]. This picture shows the detection of the hybridization signals of RAE180 repeats which are widespread in the major part of the two Y chromosomes. Sex chromosomes are indicated.

© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Garrido-Ramos, M.A. Satellite DNA: An Evolving Topic. Genes 2017, 8, 230. https://doi.org/10.3390/genes8090230
AMA Style
Garrido-Ramos MA. Satellite DNA: An Evolving Topic. Genes. 2017; 8(9):230. https://doi.org/10.3390/genes8090230
Chicago/Turabian StyleGarrido-Ramos, Manuel A. 2017. "Satellite DNA: An Evolving Topic" Genes 8, no. 9: 230. https://doi.org/10.3390/genes8090230
APA StyleGarrido-Ramos, M. A. (2017). Satellite DNA: An Evolving Topic. Genes, 8(9), 230. https://doi.org/10.3390/genes8090230
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.