
Novel Haplotype in the HHEX Gene Promoter Associated with Body Length in Pigs


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sampling
2.2. DNA and RNA Extraction
2.3. Semiquantitative Reverse Transcription PCR (SqRT-PCR) and Quantitative Real-Time PCR (qRT-PCR)
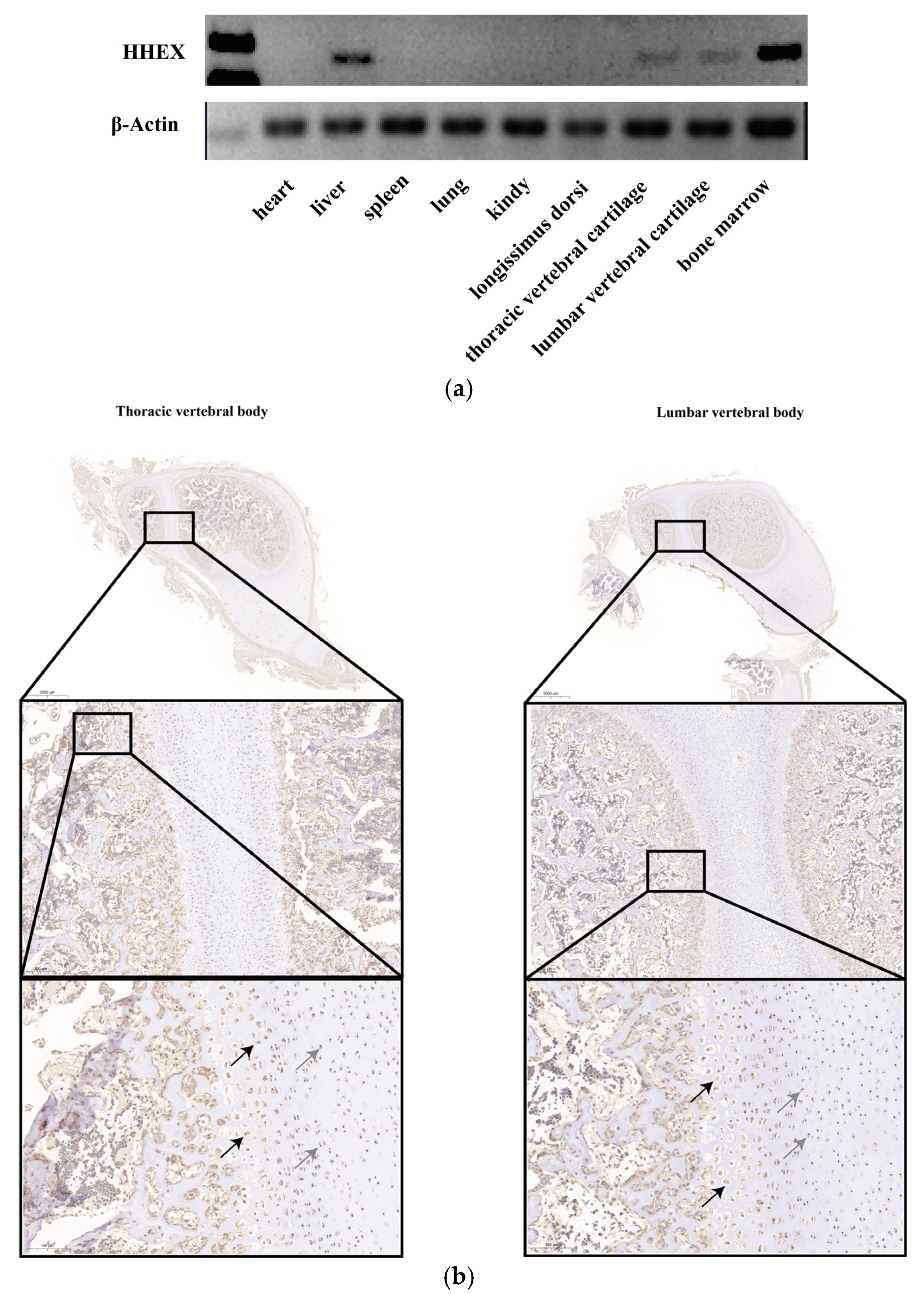
2.4. Immunohistochemistry (IHC)
2.5. Rapid Amplification of cDNA Ends (RACE)
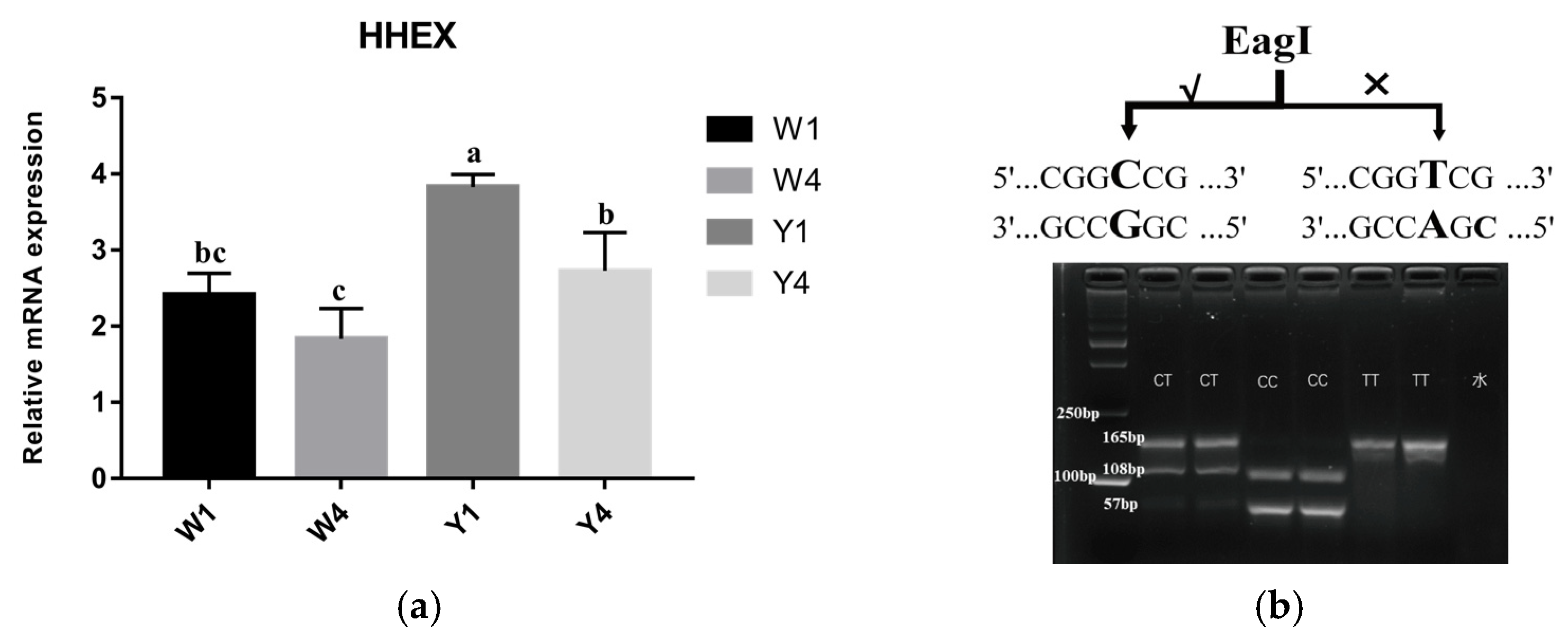
2.6. PCR Amplification, Sequencing and Genotyping
2.7. Reporter Plasmid Construction
2.8. Cell Culture and Luciferase Assay
2.9. Prediction of the Changes in Transcription Factor-Binding Sites (TFBSs)
2.10. Statistical Analysis
3. Results
3.1. Porcine HHEX Gene Expression Pattern
3.2. Polymorphism of the Porcine HHEX Gene
3.3. Comparison of Haplotype Frequencies of the HHEX Gene among Pig Breeds
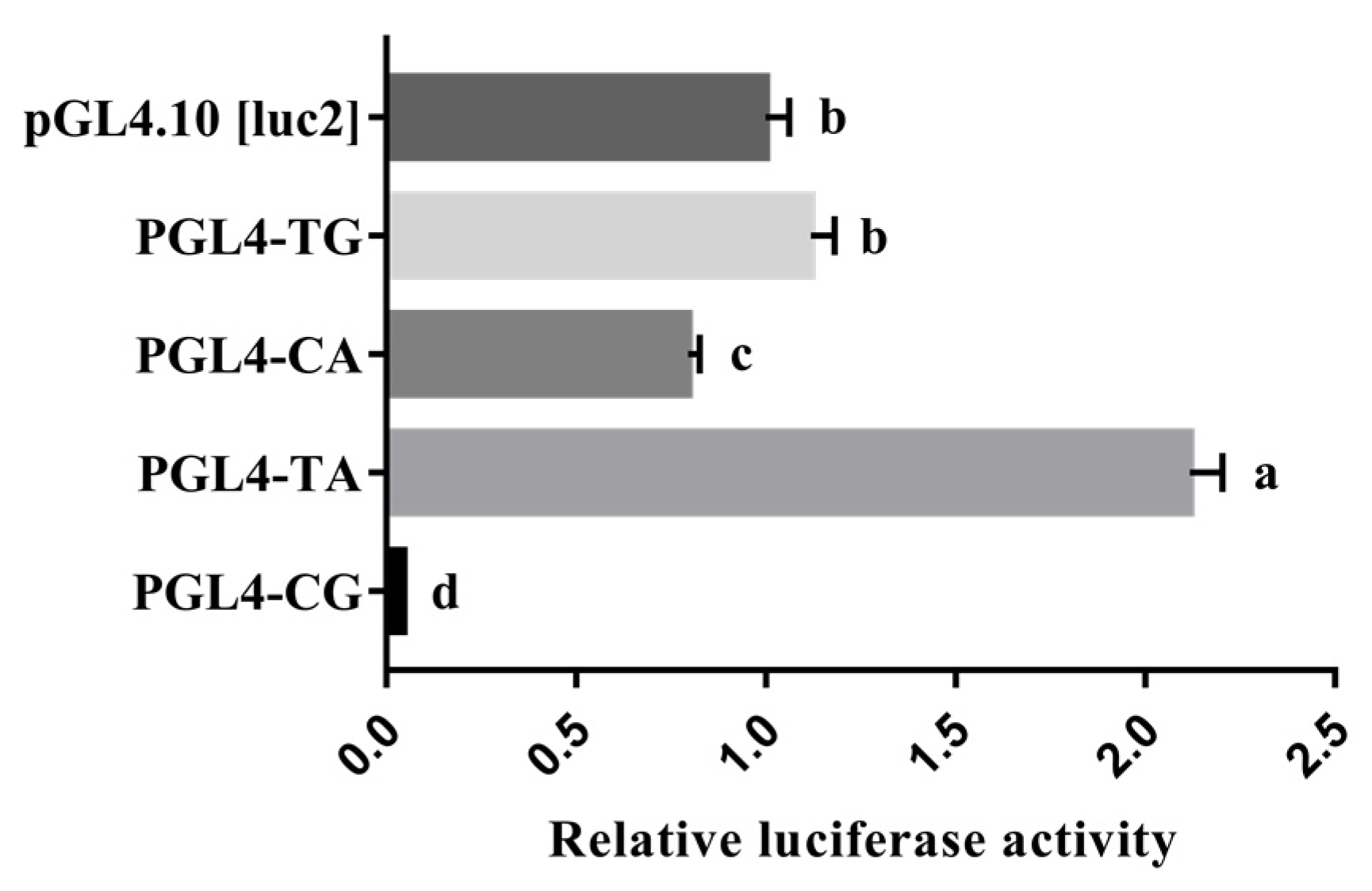
3.4. Promoter Activity Analyses of the Porcine HHEX Gene
3.5. Effect of Haplotype on the Promoter Activity of HHEX
3.6. The TA > CG Haplotype Adds a Transcription Suppressor Factor-Binding Site
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, T.; Lin, J. Variation of heart size and its correlation with growth performance and vascular space in domestic pigs. Anim. Sci. 1997, 64, 523–528. [Google Scholar] [CrossRef]
- Bogucka, J.; Kapelanski, W.; Elminowska-Wenda, G.; Walasik, K.; Lewandowska, K.L. Comparison of microstructural traits of Musculus longissimus lumborum in wild boars, domestic pigs and wild boar/domestic pig hybrids. Arch. Anim. Breed. 2008, 51, 359–365. [Google Scholar] [CrossRef]
- Ruusunen, M.; Puolanne, E. Histochemical properties of fibre types in muscles of wild and domestic pigs and the effect of growth rate on muscle fibre properties. Meat Sci. 2004, 67, 533–539. [Google Scholar] [CrossRef]
- Castillo-Contreras, R.; Mentaberre, G.; Aguilar, X.F.; Conejero, C.; Colom-Cadena, A.; Ráez-Bravo, A.; González-Crespo, C.; Espunyes, J.; Lavín, S.; López-Olvera, J.R. Wild boar in the city: Phenotypic responses to urbanisation. Sci. Total Environ. 2021, 773, 145593. [Google Scholar] [CrossRef] [PubMed]
- Larson, G.; Dobney, K.; Albarella, U.; Fang, M.; Matisoo-Smith, E.; Robins, J.; Lowden, S.; Finlayson, H.; Brand, T.; Willerslev, E.; et al. Worldwide phylogeography of wild boar reveals multiple centers of pig domestication. Science 2005, 307, 1618–1621. [Google Scholar] [CrossRef]
- Wang, J.; Liu, C.; Chen, J.; Bai, Y.; Wang, K.; Wang, Y.; Fang, M. Genome-Wide Analysis Reveals Human-Mediated Introgression from Western Pigs to Indigenous Chinese Breeds. Genes 2020, 11, 275. [Google Scholar] [CrossRef]
- Xinhong, L.; Zhen, L.; Fu, J.; Wang, L.; Yang, Q.; Li, P.; Li, Y. Quantitative proteomic profiling indicates the difference in reproductive efficiency between Meishan and Duroc boar spermatozoa. Theriogenology 2018, 116, 71–82. [Google Scholar] [CrossRef]
- Clapperton, M.; Bishop, S.C.; Glass, E.J. Innate immune traits differ between Meishan and Large White pigs. Vet. Immunol. Immunopathol. 2005, 104, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Dai, F.; Feng, D.; Cao, Q.; Ye, H.; Zhang, C.; Xia, W.; Zuo, J. Developmental differences in carcass, meat quality and muscle fibre characteristics between the Landrace and a Chinese native pig. S. Afr. J. Anim. Sci. 2009, 39, 4. [Google Scholar] [CrossRef]
- Panda, S.; Gaur, G.K.; Chauhan, A.; Kar, J.; Mehrotra, A. Accurate assessment of body weights using morphometric measurements in Landlly pigs. Trop. Anim. Health Prod. 2021, 53, 362. [Google Scholar] [CrossRef] [PubMed]
- Li, L.Y.; Xiao, S.J.; Tu, J.M.; Zhang, Z.K.; Zheng, H.; Huang, L.B.; Huang, Z.Y.; Yan, M.; Liu, X.D.; Guo, Y.M. A further survey of the quantitative trait loci affecting swine body size and carcass traits in five related pig populations. Anim. Genet. 2021, 52, 621–632. [Google Scholar] [CrossRef]
- Liu, H.; Song, H.; Jiang, Y.; Jiang, Y.; Zhang, F.; Liu, Y.; Shi, Y.; Ding, X.; Wang, C. A Single-Step Genome Wide Association Study on Body Size Traits Using Imputation-Based Whole-Genome Sequence Data in Yorkshire Pigs. Front. Genet. 2021, 12, 629049. [Google Scholar] [CrossRef]
- Groenen, M.A.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Ye, J.; Dong, L.; Li, Y.; Yan, L.; Cai, G.; Liu, D.; Tan, C.; Wu, Z. Genome-Wide Association Study for Body Length, Body Height, and Total Teat Number in Large White Pigs. Front. Genet. 2021, 12, 650370. [Google Scholar] [CrossRef]
- Johnson, Z.; Nugent, R., III. Heritability of body length and measures of body density and their relationship to backfat thickness and loin muscle area in swine. J. Anim. Sci. 2003, 81, 1943–1949. [Google Scholar] [CrossRef]
- Li, J.; Peng, S.; Zhong, L.; Zhou, L.; Yan, G.; Xiao, S.; Ma, J.; Huang, L. Identification and validation of a regulatory mutation upstream of the BMP2 gene associated with carcass length in pigs. Genet. Sel. Evol. 2021, 53, 94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Grosse-Brinkhaus, C.; Heidt, H.; Uddin, M.J.; Cinar, M.U.; Tesfaye, D.; Tholen, E.; Looft, C.; Schellander, K.; Neuhoff, C. Polymorphisms and expression analysis of SOX-6 in relation to porcine growth, carcass, and meat quality traits. Meat Sci. 2015, 107, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Huang, L.; Yang, M.; Fan, Y.; Li, L.; Fang, S.; Deng, W.; Cui, L.; Zhang, Z.; Ai, H.; et al. Possible introgression of the VRTN mutation increasing vertebral number, carcass length and teat number from Chinese pigs into European pigs. Sci. Rep. 2016, 6, 19240. [Google Scholar] [CrossRef]
- Hartmann, C. Transcriptional networks controlling skeletal development. Curr. Opin Genet. Dev. 2009, 19, 437–443. [Google Scholar] [CrossRef]
- Goodings, C.; Smith, E.; Mathias, E.; Elliott, N.; Cleveland, S.M.; Tripathi, R.M.; Layer, J.H.; Chen, X.; Guo, Y.; Shyr, Y.; et al. Hhex is Required at Multiple Stages of Adult Hematopoietic Stem and Progenitor Cell Differentiation. Stem Cells 2015, 33, 2628–2641. [Google Scholar] [CrossRef]
- Paz, H.; Lynch, M.R.; Bogue, C.W.; Gasson, J.C. The homeobox gene Hhex regulates the earliest stages of definitive hematopoiesis. Blood 2010, 116, 1254–1262. [Google Scholar] [CrossRef]
- Ruiz-Estevez, M.; Crane, A.T.; Rodriguez-Villamil, P.; Ongaratto, F.L.; You, Y.; Steevens, A.R.; Hill, C.; Goldsmith, T.; Webster, D.A.; Sherry, L.; et al. Liver development is restored by blastocyst complementation of HHEX knockout in mice and pigs. Stem Cell Res. 2021, 12, 292. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.T.; Nasa, C.; Shi, W.; Huntington, N.D.; Bogue, C.W.; Alexander, W.S.; McCormack, M.P. A crucial role for the homeodomain transcription factor Hhex in lymphopoiesis. Blood 2015, 125, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.P.; Wilson, C.M.; Jiang, X.; Cong, R.; Vasavada, H.; Kaestner, K.H.; Bogue, C.W. The homeobox gene Hhex is essential for proper hepatoblast differentiation and bile duct morphogenesis. Dev. Biol. 2007, 308, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Gauvrit, S.; Villasenor, A.; Strilic, B.; Kitchen, P.; Collins, M.M.; Marin-Juez, R.; Guenther, S.; Maischein, H.M.; Fukuda, N.; Canham, M.A.; et al. HHEX is a transcriptional regulator of the VEGFC/FLT4/PROX1 signaling axis during vascular development. Nat. Commun. 2018, 9, 2704. [Google Scholar] [CrossRef]
- Morimoto, R.; Yamamoto, A.; Akimoto, Y.; Obinata, A. Homeoprotein Hex is expressed in mouse developing chondrocytes. J. Biochem. 2011, 150, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, R.; Obinata, A. Overexpression of Hematopoietically Expressed Homeoprotein Induces Nonapoptotic Cell Death in Mouse Prechondrogenic ATDC5 Cells. Biol. Pharm. Bull. 2011, 34, 1589–1595. [Google Scholar] [CrossRef]
- Aghajanian, P.; Mohan, S. The art of building bone: Emerging role of chondrocyte-to-osteoblast transdifferentiation in endochondral ossification. Bone Res. 2018, 6, 19. [Google Scholar] [CrossRef]
- Allen, H.L.; Estrada, K.; Lettre, G.; Berndt, S.; Weedon, M.N.; Rivadeneira, F. Hundreds of variants influence human height and cluster within genomic loci and biological pathways. In The Role of Common Genetic Variation in Model Polygenic Monogenic Traits; University of Exeter: Exeter, UK, 2010; p. 94. [Google Scholar]
- Yang, J.; Benyamin, B.; McEvoy, B.P.; Gordon, S.; Henders, A.K.; Nyholt, D.R.; Madden, P.A.; Heath, A.C.; Martin, N.G.; Montgomery, G.W. Common SNPs explain a large proportion of the heritability for human height. Nat. Genet. 2010, 42, 565–569. [Google Scholar] [CrossRef]
- Kemper, K.E.; Visscher, P.M.; Goddard, M.E. Genetic architecture of body size in mammals. Genome Biol. 2012, 13, 244. [Google Scholar] [CrossRef]
- Struhl, K. Promoters, activator proteins, and the mechanism of transcriptional initiation in yeast. Cell 1987, 49, 295–297. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.T.; Ahmed, M.; Guo, H.; Zhang, Y.; Chen, S.; Soares, F.; Lu, J.; Zhou, S.; Wang, M.; Li, H. Risk SNP-mediated promoter-enhancer switching drives prostate cancer through lncRNA PCAT19. Cell 2018, 174, 564–575.e18. [Google Scholar] [CrossRef] [PubMed]
- Bečanović, K.; Nørremølle, A.; Neal, S.J.; Kay, C.; Collins, J.A.; Arenillas, D.; Lilja, T.; Gaudenzi, G.; Manoharan, S.; Doty, C.N. A SNP in the HTT promoter alters NF-κB binding and is a bidirectional genetic modifier of Huntington disease. Nat. Neurosci. 2015, 18, 807–816. [Google Scholar] [CrossRef] [PubMed]
- De Gobbi, M.; Viprakasit, V.; Hughes, J.R.; Fisher, C.; Buckle, V.J.; Ayyub, H.; Gibbons, R.J.; Vernimmen, D.; Yoshinaga, Y.; De Jong, P. A regulatory SNP causes a human genetic disease by creating a new transcriptional promoter. Science 2006, 312, 1215–1217. [Google Scholar] [CrossRef]
- Shi, Y.; Lee, J.-S.; Galvin, K.M. Everything you have ever wanted to know about Yin Yang 1. Biochim. Biophys. Acta 1997, 1332, F49–F66. [Google Scholar] [CrossRef]
- Gordon, S.; Akopyan, G.; Garban, H.; Bonavida, B. Transcription factor YY1: Structure, function, and therapeutic implications in cancer biology. Oncogene 2006, 25, 1125–1142. [Google Scholar] [CrossRef]
- Aoyama, T.; Okamoto, T.; Fukiage, K.; Otsuka, S.; Furu, M.; Ito, K.; Jin, Y.; Ueda, M.; Nagayama, S.; Nakayama, T. Histone modifiers, YY1 and p300, regulate the expression of cartilage-specific gene, chondromodulin-I, in mesenchymal stem cells. J. Biol. Chem. 2010, 285, 29842–29850. [Google Scholar] [CrossRef]
- Guérit, D.; Brondello, J.-M.; Chuchana, P.; Philipot, D.; Toupet, K.; Bony, C.; Jorgensen, C.; Noël, D. FOXO3A regulation by miRNA-29a Controls chondrogenic differentiation of mesenchymal stem cells and cartilage formation. Stem Cells Dev. 2014, 23, 1195–1205. [Google Scholar] [CrossRef]
- Kurisaki, K.; Kurisaki, A.; Valcourt, U.; Terentiev, A.A.; Pardali, K.; Ten Dijke, P.; Heldin, C.-H.; Ericsson, J.; Moustakas, A. Nuclear factor YY1 inhibits transforming growth factor β-and bone morphogenetic protein-induced cell differentiation. Mol. Cell. Biol. 2003, 23, 4494–4510. [Google Scholar] [CrossRef]
- Ruijter, A.J.D.; Gennip, A.H.V.; Caron, H.N.; Kemp, S.; Kuilenburg, A.B.V. Histone deacetylases (HDACs): Characterization of the classical HDAC family. Biochem. J. 2003, 370, 737–749. [Google Scholar] [CrossRef]
- Hong, S.; Derfoul, A.; Pereira-Mouries, L.; Hall, D.J. A novel domain in histone deacetylase 1 and 2 mediates repression of cartilage—Specific genes in human chondrocytes. FASEB J. 2009, 23, 3539–3552. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Zhang, Z.; Huang, Z.; Chen, W.; Huang, G.; Meng, F.; Kang, Y. MicroRNA-92a-3p regulates the expression of cartilage-specific genes by directly targeting histone deacetylase 2 in chondrogenesis and degradation. Osteoarthr. Cartil. 2017, 25, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-M.; Yao, Y.-L.; Sun, J.-M.; Davie, J.R.; Seto, E. Isolation and characterization of cDNAs corresponding to an additional member of the human histone deacetylase gene family. J. Biol. Chem. 1997, 272, 28001–28007. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-M.; Inouye, C.; Zeng, Y.; Bearss, D.; Seto, E. Transcriptional repression by YY1 is mediated by interaction with a mammalian homolog of the yeast global regulator RPD3. Proc. Natl. Acad. Sci. USA 1996, 93, 12845–12850. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | Total Number | Genotype, n, Frequency | Allele, n, Frequency | Body Length, cm | χ2, df, p Value a | |||
|---|---|---|---|---|---|---|---|---|
| CC | CT | TT | C | T | ||||
| Wuzhishan pig | 24 | 15 | 9 | 0 | 39 | 9 | 67.08 ± 1.19 | 297.459, 2, p < 0.0001 |
| 0.63 | 0.38 | 0.00 | 0.81 | 0.19 | ||||
| Bama Pig | 22 | 22 | 0 | 0 | 44 | 0 | 74.25 ± 0.25 | |
| 1.00 | 0.00 | 0.00 | 1.00 | 0.00 | ||||
| Tibetan pig | 24 | 13 | 11 | 0 | 37 | 11 | 72.9 ± 0.69 | |
| 0.54 | 0.46 | 0.00 | 0.77 | 0.23 | ||||
| Banna mini-pig | 34 | 24 | 7 | 3 | 55 | 13 | 54.6 ± 0.80 | |
| 0.71 | 0.21 | 0.09 | 0.81 | 0.19 | ||||
| Jiangkou radish pig | 47 | 30 | 15 | 2 | 75 | 19 | 69 | |
| 0.64 | 0.32 | 0.04 | 0.80 | 0.20 | ||||
| Longlin pig | 37 | 22 | 13 | 2 | 57 | 17 | 96.5 ± 3.80 | |
| 0.59 | 0.35 | 0.05 | 0.77 | 0.23 | ||||
| Min pig | 36 | 16 | 20 | 0 | 52 | 20 | 124.5 ± 0.47 | |
| 0.44 | 0.56 | 0.00 | 0.72 | 0.28 | ||||
| Yanan pig | 14 | 12 | 2 | 0 | 26 | 2 | 124 | |
| 0.86 | 0.14 | 0.00 | 0.93 | 0.07 | ||||
| Qianbei black pig | 35 | 21 | 14 | 0 | 56 | 14 | 93.45 | |
| 0.60 | 0.40 | 0.00 | 0.80 | 0.20 | ||||
| Saba pig | 35 | 17 | 18 | 1 | 52 | 20 | 134.38 ± 0.7 | |
| 0.47 | 0.50 | 0.03 | 0.74 | 0.29 | ||||
| Yorkshire pig | 29 | 0 | 0 | 29 | 0 | 58 | 165 ± 2.47 | |
| 0.00 | 0.00 | 1.00 | 0.00 | 1.00 | ||||
| Landrace pig | 30 | 0 | 4 | 26 | 4 | 56 | 160.1 ± 0.73 | |
| 0.00 | 0.13 | 0.87 | 0.07 | 0.93 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Xu, Q.; Xue, M.; Wang, Y.; Yang, X.; Chan, S.; Tang, Q.; Wang, F.; Sun, R.; Chao, Z.; et al. Novel Haplotype in the HHEX Gene Promoter Associated with Body Length in Pigs. Genes 2023, 14, 511. https://doi.org/10.3390/genes14020511
Luo Y, Xu Q, Xue M, Wang Y, Yang X, Chan S, Tang Q, Wang F, Sun R, Chao Z, et al. Novel Haplotype in the HHEX Gene Promoter Associated with Body Length in Pigs. Genes. 2023; 14(2):511. https://doi.org/10.3390/genes14020511
Chicago/Turabian StyleLuo, Yabiao, Qiao Xu, Mingming Xue, Yubei Wang, Xiaoyang Yang, Shuheng Chan, Qiguo Tang, Feng Wang, Ruiping Sun, Zhe Chao, and et al. 2023. "Novel Haplotype in the HHEX Gene Promoter Associated with Body Length in Pigs" Genes 14, no. 2: 511. https://doi.org/10.3390/genes14020511
APA StyleLuo, Y., Xu, Q., Xue, M., Wang, Y., Yang, X., Chan, S., Tang, Q., Wang, F., Sun, R., Chao, Z., & Fang, M. (2023). Novel Haplotype in the HHEX Gene Promoter Associated with Body Length in Pigs. Genes, 14(2), 511. https://doi.org/10.3390/genes14020511