1. Introduction
Circular RNAs (circRNAs) are produced in all eukaryotic cells [
1]. Absence of free termini confers high stability to circRNAs [
2]. The acknowledged functions of circRNAs include sponging of microRNAs and sequestration of proteins [
3,
4]. CircRNAs can also code for proteins and regulate transcription [
5,
6]. Multiple mechanisms, including backsplicing and circularization of lariat RNAs, are associated with circRNA synthesis [
7]. Most circRNAs in humans are produced by backsplicing in which the downstream 5′ splice site (5′ss) pairs with the upstream 3′ss [
7]. Backsplicing and, hence, circRNA production, are facilitated by inverted Alu repeats present in primate pre-mRNAs due to secondary structure formation [
8,
9]. Protein factors, including DHX9, QKI and SFPQ, have also been implicated in circRNA synthesis [
8,
10,
11]. DHX9 is a DExD/H-box helicase involved in DNA repair, transcription, pre-mRNA splicing and microRNA processing [
12]. QK1 and SFPQ are RNA-binding proteins associated with cell differentiation and neuronal functions, respectively [
13,
14]. Findings that different types of RNA binding proteins are involved in circRNA generation support the diversity of mechanisms by which circRNAs are produced. However, limited attention has been paid to the role of the upstream and/or downstream splicing events in circRNAs formation.
Humans contain two copies of
survival motor neuron (
SMN) genes,
SMN1 and
SMN2 [
15]. Both genes code for SMN, a multifunction protein essential for the survival of all cell types of the animal kingdom [
16]. Mutations or deletions of
SMN1 cause spinal muscular atrophy (SMA), a leading genetic cause of infant mortality [
17,
18].
SMN2 cannot compensate for the loss of
SMN1 due to predominant skipping of exon 7. Exon 7 is a coding exon; thus, its skipping results in production of a truncated and unstable protein [
19]. SMN levels increase through manipulation of
SMN2 exon 7 splicing using antisense oligonucleotide or a small molecule, which constitute FDA-approved therapies for SMA [
20,
21]. Gene therapy has also been used to treat SMA [
22]. Another avenue to increase SMN levels is through enhanced transcription of the
SMN2 gene, which is almost universally present in SMA patients [
23]. With significance to transcription and splicing regulation, the human
SMN genes harbor a remarkably high number of Alu elements [
24]. We and others have recently reported an enormous repertoire of circRNAs generated by the human
SMN genes [
25,
26]. Based on their exon composition, we categorized
SMN circRNAs into four types: Type 1, 2, 3 and 4 [
25]. Type 1 circRNAs harbor early exons and use the 5′ss of exon 4 or upstream exons (
Figure 1). Type 2 circRNAs harbor both upstream and middle exons. Type 3 circRNAs use the 5′ss located downstream of exon 7 and may incorporate all types of exons, including middle and/or downstream exons (
Figure 1). Type 4
SMN circRNAs are formed by trans-splicing encompassing one or more exons from another gene.
C2A-2B-3-4, C2B-3-4 and C3-4 are the most abundant circRNAs produced by the
SMN genes [
25] (
Figure 1). All three of them contain exons 3 and 4 with or without upstream exons 2A and 2B. Depletion of DHX9, an RNA helicase associated with the unwinding of RNA duplexes formed by inverted Alu repeats, upregulates the C3-4 level at the expense of C2A-2B-3-4 and C2B-3-4 [
25]. However, in terms of a complete understanding of the mechanism of
SMN circRNA production, it remains poorly understood. Here, we employed
SMN as a model system to examine the impact of intronic sequences on backsplicing during synthesis of circRNAs with multiple exons. Our results obtained using
SMN circRNA-generating vectors transiently expressed in HeLa cells support that forward splicing of an internal intron joining exons to be included into circRNA favors backsplicing involving a downstream 5′ss. We confirmed our findings employing inducible cell lines stably expressing
SMN circRNAs. Our results support the role of the exon junction complex (EJC) in the generation of exon-only-containing circRNAs. We showed that
SMN circRNAs are preferentially localized in the cytoplasm. These findings bring novel insights into our understanding of circRNA generation. The findings are also significant for uncovering novel functions of the
SMN genes independent of SMN protein functions.
2. Materials and Methods
2.1. HeLa Cell Culture
HeLa cells (American Type Culture Collection, ATCC) were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Gibco, Grand Island, NY, USA, Cat No. 11965-092), supplied with 10% fetal bovine serum (FBS, Gibco, Cat No. 26140-079). All cell culture media and reagents were purchased from Life Technologies, Carlsbad, CA, USA. Cells were cultured in CO2 incubator (Thermo Fisher, Waltham, MA, USA, NAPCO Series 8000WJ) at 37 °C and 5% CO2.
2.2. Generation of circRNA Overexpression Constructs and Their Linear Counterparts
Abbreviations used in this publication are listed in
Supplementary Table S1. Expression vectors constructed in this study are listed in
Supplementary Table S2. Primers used in this study are listed in
Supplementary Table S3. The pC2B-3-4 plasmid was constructed by inserting PCR amplified products in pCI vector (Promega, Madison, WI, USA, Cat No. E1731) using cDNA and/or genomic DNA as template, as described in
Supplementary Figure S1. Other expression vectors were generated following the similar protocol using pC2B-3-4 as template (
Figure 2,
Supplementary Figure S2A and
Supplementary Table S2). Briefly, pC2B-3-4 was digested with BsiWI and EcoNI to serve as backbone. Then, PCR products were digested with same enzymes and inserted between BsiWI and EcoNI sites. We used pcDNA5/FRT/TO (Thermo Fisher Scientific, Cat No. V652020) as backbone to make constructs in Flp-In™ system (Thermo Fisher Scientific), To generate expression vectors for stable cell lines, we amplified sequences between XhoI and NotI sites from pCI-based constructs and replaced XhoI with BamHI. Subsequently, inserts and pcDNA5/FRT/TO vector were digested by BamHI and NotI, and ligated through these two sites. The pcDNA5/FRT/TO-based vectors include pTC2A-2B-3-4
Int, pTL2A-2B-3-4
Int, pTC2B-3-4
Int, pTL2B-3-4
Int, pTC3-4
Int and pTL3-4
Int (
Supplementary Table S2).
2.3. Establishment of Inducible Cell Lines
Cell lines established in this study are listed in
Supplementary Table S4. T-REx™-293 cell line (Thermo Fisher Scientific, R71007, hereinafter referred as T-REx) was cultured in DMEM (Gibco, Cat No. 11965-092), supplied with 10% FBS (Gibco, Cat No. 16000-044) and 1× Glutamax (Gibco, Cat No. 35050061). All cell culture media and reagents were purchased from Life Technologies. Antibiotics including zeocin, blasiticidin, hygromycin (Life technologies Cat No. R250-01, R210-01, R220-05) and doxycycline (Dox) (Sigma-Aldrich, St. Louis, MO, USA, Cat No. D9891-10G) were used as instructed. Plasmids pOG44 and pcDNA5/FRT/TO were obtained from Thermo Fisher Scientific (Cat No. V600520 and Cat No. V652020). To establish stable cell lines for inducible expression of circRNAs of interest, the circRNA-expression plasmid was mixed with pOG44 at a 9:1 ratio (
w/w). Then, these two plasmids were co-transfected into the host T-REx cell line by using Lipofectamine 2000 as per the manufacturer’s instructions. Of note, The Flp recombinase was expressed from pOG44, which facilitated integration of circRNA-expression plasmid into the host cell line. To estimate the transfection and integration efficacy, pcDNA5_FRT_mCH was used as a positive control (Addgene, Watertown, MA, USA, Cat No. 127109). Fresh medium was added 6 h (h) post transfection. At 24 h post transfection, cells were trypsinized and seeded at a density of 1.1 × 10
6 cells per 100 mm dish. The concentration of hygromycin used to select for stable transfectants was determined to be 100 µg/mL. Medium containing hygromycin was replaced every 3 days. After 2 weeks of growth with hygromycin, the surviving cell colonies were isolated using cloning disks (Bel-ART, Wayne, NJ, USA, Cat No. 37847-0001) following the manufacturer’s instructions. To induce the expression of circRNA of interest, 0.1 µg/mL Dox was added to cell culture medium, and cells were collected for RNA preparation 24 h post treatment.
2.4. Transfection
Reverse transfection was used for transient gene overexpression. Unless stated otherwise, for each reverse transfection, cell suspension containing 0.25 × 106 HeLa cells was mixed with pre-formed complexes between the plasmid DNA (0.4 µg) and Lipofectamine 2000 (1 µL) in a total volume of 0.5 mL. The mixture was then placed into a well of a 24-well plate. The medium was changed after 6 h. The cells were collected at 24 h post transcription.
2.5. Cytoplasmic and Nuclear Fractionation
The cytoplasmic and nuclear fraction were prepared using NE-PER™ Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher, Cat No. 78833) following the manufacturer’s instructions. CERI and NER buffers from the kit were supplemented with 1× Halt™ Protease and Phosphatase Inhibitor Cocktail (Thermo Scientific, 100X, Cat No. 78440) and 0.25–1 U/µL RiboLock RNase Inhibitor (Thermo Scientific, Cat No. EO0381). Around 5 × 106–8 × 106 cells were used for each fractionation. Cells were collected by trypsinization, pelleted and washed twice with ice-cold Dulbecco’s phosphate-buffered saline (DPBS, Gibco, Cat No. 14190-144).
2.6. Western Blot
Protein concentration was measured by Bio-Rad protein assay (Bio-Rad, Hercules, CA, USA, Cat No. 5000006). A total amount of 8 µg of protein was loaded per each lane of a 10% SDS-PAGE gel. After electrophoresis, proteins were transferred to PVDF membranes using a Transblot Turbo fast transfer system (Bio-Rad). Membranes were blocked in 5% non-fat milk dissolved in Tris-buffered saline containing 0.05% Tween-20 (TBST). Primary antibody incubation was carried out at 4 °C overnight. Primary antibody dilutions were as follows: mouse anti-α-tubulin 1:4000 (Sigma-Aldrich, Cat No. T6199), rabbit anti-Histone H3 1:4000 (Abcam, Waltham, MA, USA, Cat No. 1791). After incubation with primary antibodies, membranes were washed in TBST three times and incubated with secondary antibodies for 1 h at room temperature. Secondary antibodies and their dilutions were as follows: goat anti-mouse 1:4000 (Jackson ImmunoResearch Laboratories Inc, West Grove, PA, USA, Cat No. 115-035-003), donkey anti-rabbit 1:2000 (Cytiva USA, Marlborough, MA, USA, Cat No. NA934). After secondary antibody incubations, membranes were washed in TBST again and developed using SuperSignal West Femto Maximum Sensitivity Substrate (Thermo Scientific, Cat No. 34094). Bands were then visualized using a UVP Biospectrum AC imaging system.
2.7. RNA Isolation and RNase R Treatment
Total RNA was isolated from cells using TRIzol reagent (Invitrogen, Waltham, MA, USA, Cat No. 15596018). RNA was treated with RQ1 RNase-free DNase (Promega, Cat No. M6101), followed by phenol:chloroform extraction and ethanol precipitation. RNase R treatment was carried out in 10 µL reaction, in which 2 µg of RNA was digested by 0.5 µL (10 U) of RNase R (Applied Biological Material Inc., Richmond, BC, Canada, Cat No. E049). Of note, RNA samples used in the RNase R treatment were treated with DNase first. The treatment was performed for 45 min (min) at 37 °C, followed by the enzyme inactivation at 65 °C for 20 min. To assess the efficiency of digestion, mock reactions in which the enzyme was omitted were performed side by side.
2.8. Reverse Transcription and PCR (RT-PCR)
cDNA was synthesized using Superscript III reverse transcriptase (RT) (Invitrogen), following the manufacturer’s instructions. Specifically, 0.5 µg of total RNA was used per 5 µL RT reaction, employing either a gene-specific (
Supplementary Table S3) or a random primer (Promega, Cat No. C1181). For semi-quantitative PCR, 1 µL of cDNA reaction was used as a template in 20 µL reaction. PCR products were separated on 5.3% or 6% native polyacrylamide gels and visualized by ethidium bromide staining. For qPCR, cDNA was first diluted 20 times, and 3 µL was used as a template per 20 µL qPCR reaction containing 1× PowerUp SYBR green master mix (Life Technologies, Cat No. A25742) and the desired pair of primers. qPCR reactions in biological or technical triplicates were performed on a QuantStudio 3 (Thermo Fisher) thermocycler. Relative expression was determined using the 2
-ΔΔCt method using GAPDH as the normalizing gene for gene expression assays. To determine the copy number of circRNA per cell, we employed standard curves that were generated using absolute amounts of the linearized plasmids containing the sequence of the expected PCR products amplified from circRNA of interest. As per published reports, a single HeLa and HEK 293 cell contain ~30 pg and ~16 pg RNA, respectively (
https://www.thermofisher.com/us/en/home/references/ambion-tech-support/rna-tools-and-calculators/macromolecular-components-of-e.html (accessed on 4 January 2022),
https://www.aatbio.com/resources/faq-frequently-asked-questions/What-is-the-total-RNA-content-in-HeLa-cells#:~:text=For%20HeLa%20cells%2C%20the%20total,30%20pg%20per%20HeLa%20cell (accessed on 4 January 2022),
https://www.miltenyibiotec.com/_Resources/Persistent/ca9f513c68ed01981bc4d7aa25b01c90db75e6f5/Average_RNA_yields.pdf (accessed on 4 January 2022)). We confirmed RNA concentration per cell in the range of the reported concentrations. We used 0.015 µg RNA to synthesize cDNA for each qPCR. To obtain the copy number per cell, the quantified total copy number in each qPCR reaction was divided by the cell number. Error bars represent standard error of the mean. Statistical significance: *,
p < 0.05; **,
p < 0.01. All primers were obtained from Integrated DNA Technologies.
2.9. Multi-Exon Skipping Detection Assay (MESDA)
MESDA is a powerful technique developed in 2012 to determine the relative abundance of the alternative spliced transcripts of
SMN [
27]. As many as 16 alternatively spliced variants of
SMN have been detected in a single reaction using MESDA [
28]. While technique does not distinguish between transcripts of
SMN1 and
SMN2, it bridges a critical gap that cannot be reliably filled by available methods, including qPCR. Only exon 7 is differently regulated between
SMN1 and
SMN2 genes, and there is a DdeI digestion method to capture those differences [
27]. However, when it comes to determining the relative abundance of the vast number of alternatively spliced transcripts, as far as
SMN are concerned, MESDA remains the method of choice. Of note, conclusions of MESDA rely upon the comparison of different bands within the same lane. Hence, findings of MESDA cannot be altered by differential loading of samples or changing the nature of control in the neighboring lane. We have confirmed the identity of each band detected by MESDA by sequencing. Method has an inherent advantage to provide useful information even when a single sample is analyzed without any control, particularly when the primary goal is to determine the relative abundance of the alternatively spliced transcripts of
SMN. We performed MESDA using forward and reverse primers at exons 1 and 8, respectively [
28,
29]. The cDNA was prepared as described in the RT-PCR section. To ensure capturing of the low abundant spliced variants of
SMN, we performed MESDA using the 5′-end-P32-radio-labelled primer, 3′ Ex8-25, as reported earlier [
29].
2.10. Identification of Circular and Linear Transcripts
Bands of interest corresponding to RT-PCR products were excised from native polyacrylamide gels, and DNA was eluted using a “crush and soak” method. The recovered DNA was then cloned into a pGEM-T Easy vector (Promega, Cat No. A1360) following the manufacturer’s instructions. After initial screening by blue/white colony selection on indicator plates, colonies were subjected to a colony PCR. Colonies with plasmids that carried inserts of the expected size were propagated, and plasmids purified using the QIAprep Spin Miniprep Kit (Qiagen, Germantown, MD, USA, Cat No. 27014). Insert identities were determined by Sanger sequencing.
2.11. Statistical Analysis
All calculations were performed in Excel (Microsoft, Version 16.62, Redmond, WA, USA). Data were expressed as mean ± standard error of the mean (SEM). Statistical analyses were performed using the unpaired Student’s t-test. Unless otherwise mentioned, experiments were performed in triplicate, p values were two-tailed and the level of statistical significance was set as p < 0.05.
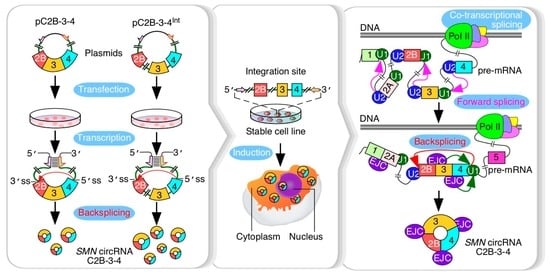
4. Discussion
The human
SMN genes code for a vast repertoire of circRNAs. The most abundant of these circRNAs are generated by the first four internal exons, e.g., exons 2A, 2B, 3 and 4. However, the mechanism of their production remains unknown. We began this study with the construction of a mammalian expression vector that allowed overexpression of C2B-3-4, an
SMN circRNA comprised of exons 2B, 3 and 4. The expression of C2B-3-4 was enabled by a strong RNA:RNA duplex that brought the downstream 5′ss of exon 4 in proximity with the upstream 3′ss of exon 2B, forcing backsplicing between these two splice sites (
Figure 2A). Despite the absence of introns 2B and 3, the expression of C2B-3-4 from the vector was 1000-fold higher than the background level expression in the untransfected cells. Employing a similar approach, we generated two non-
SMN circRNAs containing
eGFP and
HIPK2 sequences. High expression of these non-
SMN circRNAs suggested that backsplicing was primarily driven by the common RNA:RNA duplex that brought the splice sites together. The
SMN genes generate multiple alternatively spliced transcripts due to skipping exons 3, 5 and 7 in different combinations [
31]. We observed no effect of overexpression of any of the circRNAs on the relative abundance of splice isoforms generated from the endogenous
SMN.
Production of circRNAs by backsplicing requires an efficient pairing of a downstream 5′ss with an upstream 3′ss. In the case where multiple exons are located within a circRNA, removal of intron(s) by forward splicing is required so that an RNA circle is fully devoid of an intronic sequence. It is generally difficult to establish if the removal of an intron by forward splicing precedes or follows the backsplicing event. However, it is possible to test the proof of principle that forward splicing of introns within the circularized region modulates the backsplicing event. We tested this proof of principle in the context of the three abundantly expressed exon-only circRNAs of
SMN, C3-4, C2B-3-4 and C2A-2B-3-4, which require removal of 1, 2 and 3 introns, respectively. All three circRNAs use 5′ss of exon 4 but different 3′ss. We observed noticeable upregulation of circRNAs upon insertion of intervening introns, suggesting that factors recruited to the internal introns expedite the backsplicing event. Multiple mechanisms may account for this stimulatory effect, including a better recruitment of U1 snRNP at the 5′ss of exon 4, promoted by cross-exon interactions initiated by factors recruited at the 3′ss of exon 4. Such a mechanism will be consistent with the exon definition model in which recognition of splice sites at each end of an internal exon is promoted by a network of factors enveloping the entire exon [
38]. Transcription is coupled to splicing, and removal of intronic sequences is accompanied by the deposition of the exon junction complex, EJC, which is known to promote splicing of the neighboring introns [
39,
40] (
Figure 7). Moreover, eukaryotic initiation factor eIF4A3, one of the components of EJC, has been implicated in circRNA generation [
41]. Hence, it is possible that components of EJC, including eIF4A3, are regulators of
SMN circRNA generation (
Figure 7). A mutually inclusive hypothesis would be that the intercalating introns increase the size of precursor transcripts, rendering more flexible structures and facilitating backsplicing. Future studies will reveal if these and additional mechanisms that disproportionately confer enhanced stability of circRNAs contribute towards the elevated levels of circRNAs produced by the intron-containing precursors.
We established the inducible stable cell lines for overexpression of C3-4, C2B-3-4 and C2A-2B-3-4 upon induction with Dox. Unlike transient transfections in which multiple copies of the expressing vector could be present within a single cell, stable cell lines incorporated a single transgene of interest at a specific site within the genome. All these stable cell lines designed to express
SMN circRNAs harbored internal introns. We observed robust expression of
SMN circRNAs upon induction with Dox. As in the case of transient transfections, we observed no effect of the overexpressed
SMN circRNAs on splicing of the endogenous
SMN in any of the stable cell lines. These results support that factors associated with the removal of internal introns 2A, 2B and 3 are not limiting due to overexpression of
SMN circRNAs. All three circRNAs of
SMN we examined utilize 5′ss of exon 4 for backsplicing. A single G-to-C substitution at the invariant first position of the 5′ss of intron 4 completely suppressed backsplicing events. Interestingly, G-to-C substitution appeared to suppress skipping of exon 3 in the context of the linear transcripts generated in HeLa cells transiently transfected with vectors expressing L2A-2B-3-4 and L2B-3-4 (
Supplementary Figure S2D).
We have previously reported circRNAs encompassing individual
SMN exons 2A, 2B, 3 and 4, although they were generated at insignificant levels [
25]. G-to-C substitution abrogating the 5′ss of exon 4 could, in principle, favor a circularization event involving individual exons 2A, 2B and 3 due to the steric advantage conferred by the protracted stem formed by inverted repeats present in the overexpressed precursor RNA. Yet, we did not detect circRNAs comprised of single exons. These results suggest that the structural constraints imposed by the RNA:RNA duplex, which pairs the 5′ss of exon 4 with the 3′ss of an upstream exon, are not conducive for the usage of the alternative (internal) 5′ss generating small circRNAs.
Our results of fractionation studies confirm the overwhelming cytosolic localization of
SMN circRNAs. We have previously reported potential binding sites of several microRNAs within C2A-2B-3-4, C2B-3-4 and C3-4 [
42]. Considering microRNAs suppress translation, sponging of microRNAs by
SMN circRNAs localized in the cytoplasm is likely to impact translation of multiple mRNAs. Some of these microRNAs, including miR-130b-5p, miR-15b-3p and miR-6813-5p, are predicted to interact with the unique backsplice junction of C2A-2B-3-4. Hence, C2A-2B-3-4 has the potential to regulate the expression of microRNA targets with high specificity. Based on the analysis of CLIP data, several proteins, including AGO1, ASF/SF2, SRSF3, U2AF2 and YTHDC1, are predicted to interact with C2A-2B-3-4, C2B-3-4 and C3-4 [
42]. Additional proteins, including FUS and TAF, have potential interaction sites within C2A-2B-3-4 and C2B-3-4 [
42]. Sequestration of these proteins is likely to impact essential processes, including transcription, pre-mRNA splicing, translation and stress granule formation. Despite the absence of Alu elements, the mouse
Smn gene expresses mC2A-2B-3-4 [
25]. Of note, we used prefix ‘m’ to denote mouse circRNAs for clarity. Hence, functions associated with C2A-2B-3-4 are likely to be conserved between two distantly related organisms. Interestingly, C2B-3-4 and C3-4 were not detected in mice, suggesting their origins after Alu elements were inserted in primates [
25]. Future studies will reveal if some of the human-specific functions of
SMN genes are linked in part to
SMN circRNAs. Now that we have generated stable cell lines expressing three abundant circRNAs of
SMN, the stage is set to uncover novel
SMN gene functions independent of the SMN protein.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






