
BMP7 Functions to Regulate Proliferation of Dermal Papilla Cells in Hu Sheep

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Cell Culture
2.2. Bioinformatics Analysis of BMP7 Gene in Hu Sheep
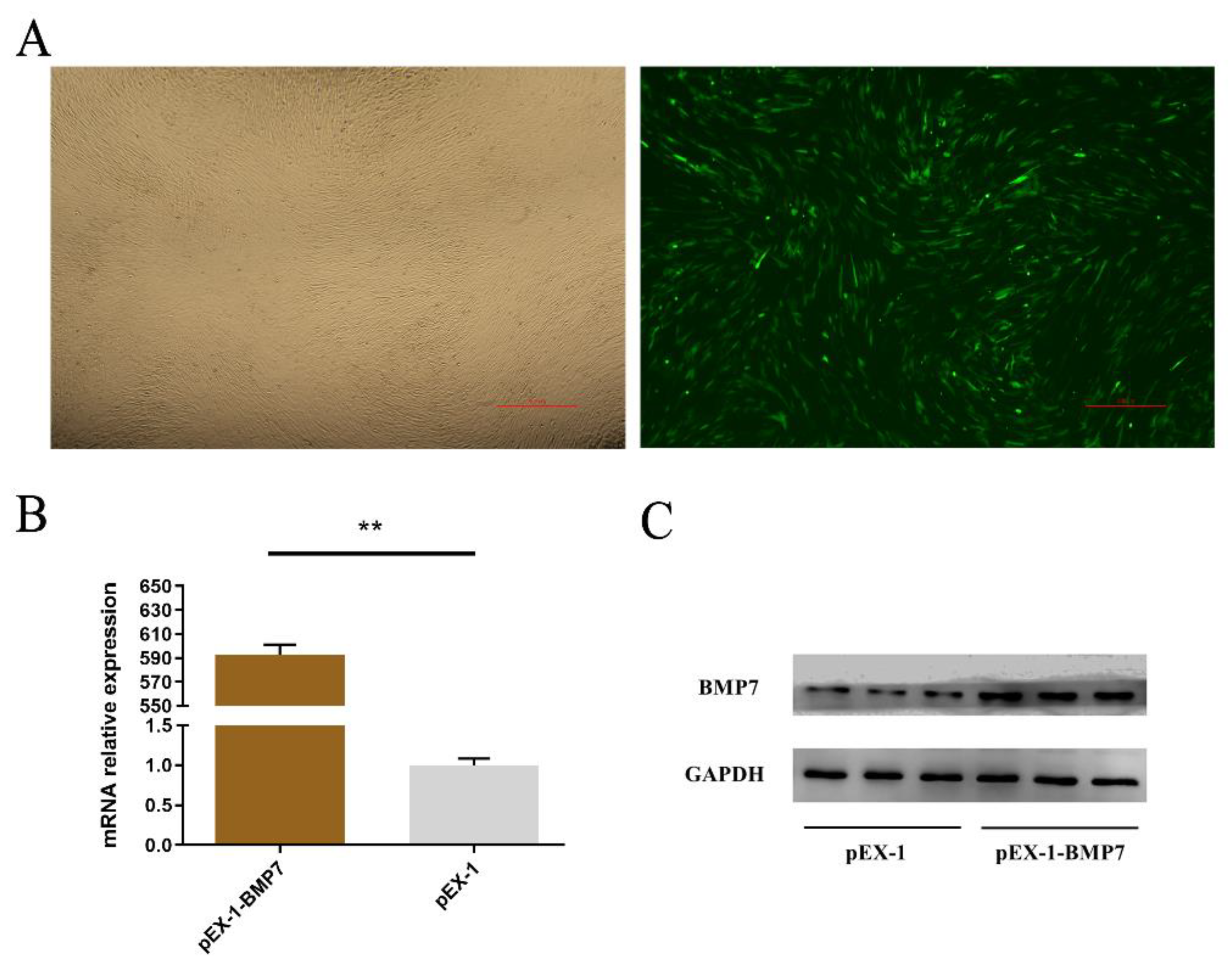
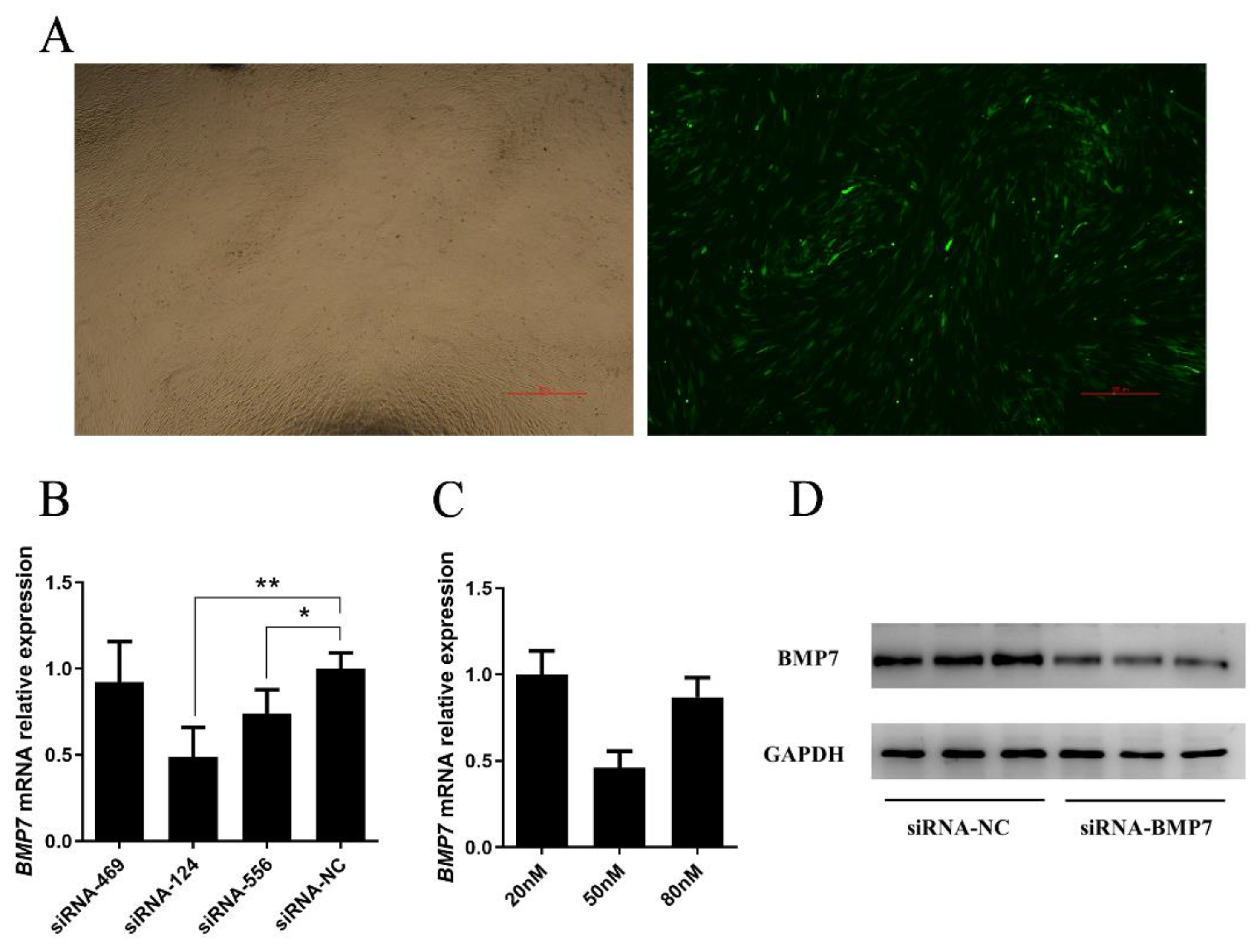
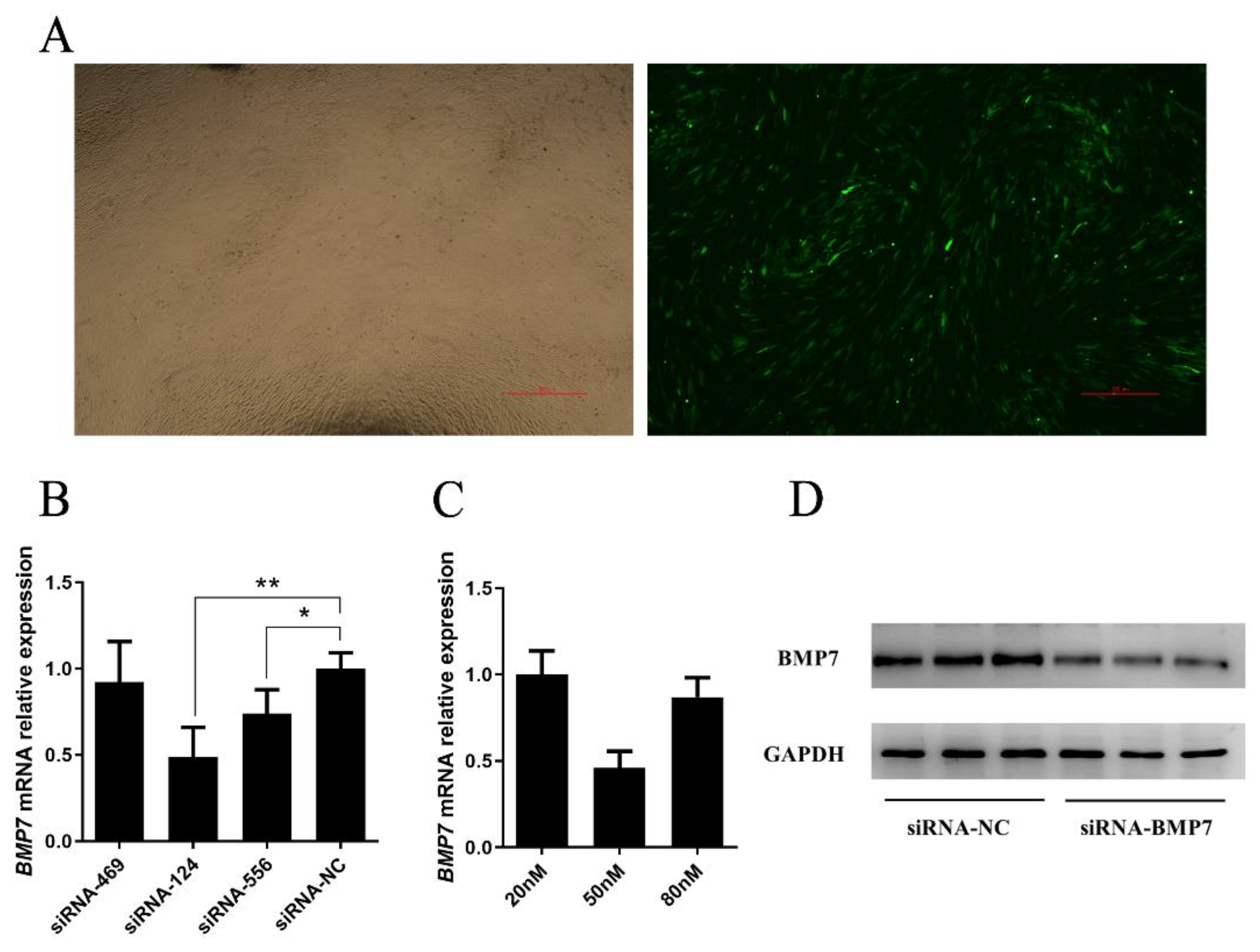
2.3. Over Expression and Inhibition of BMP7
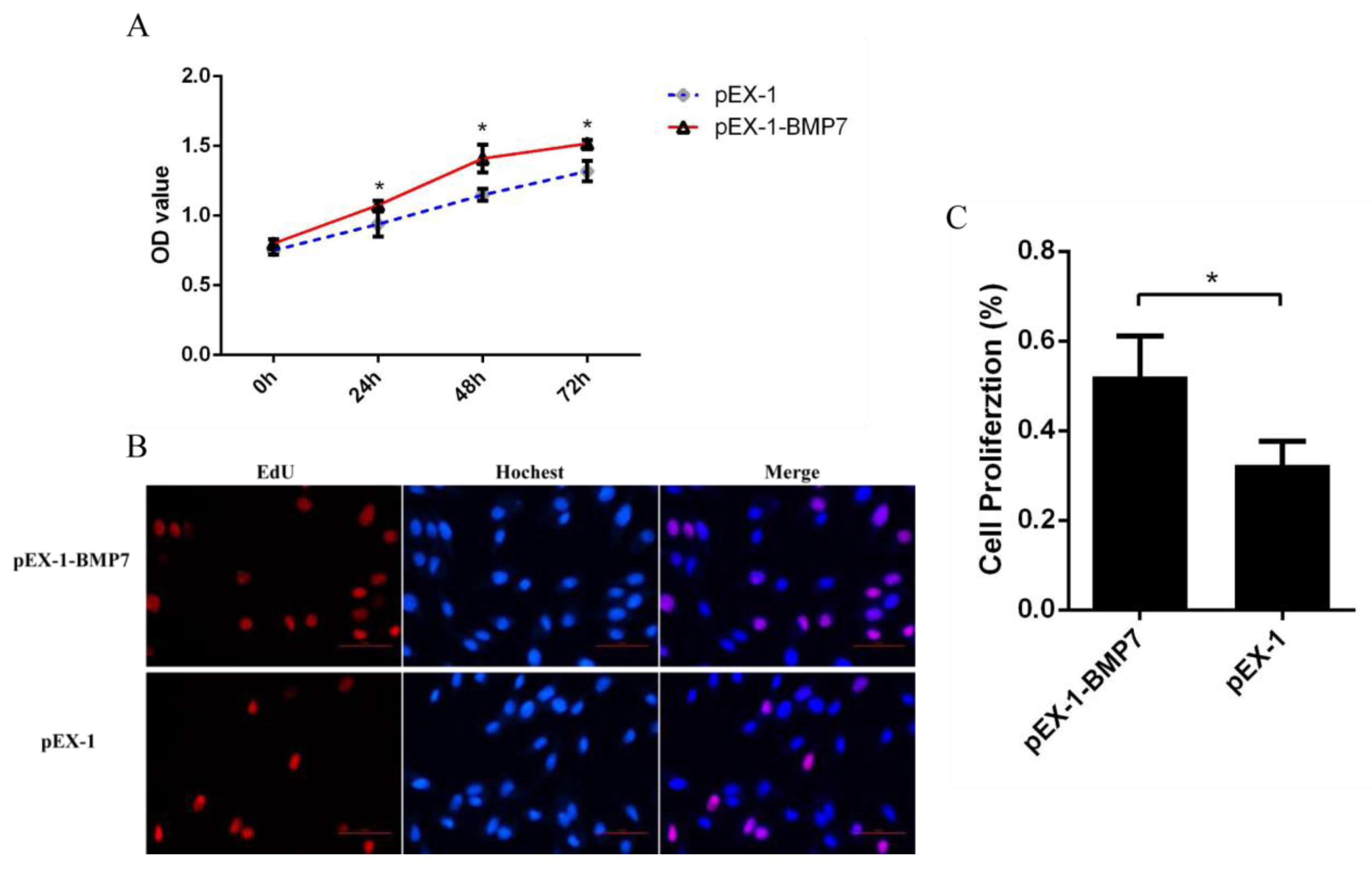
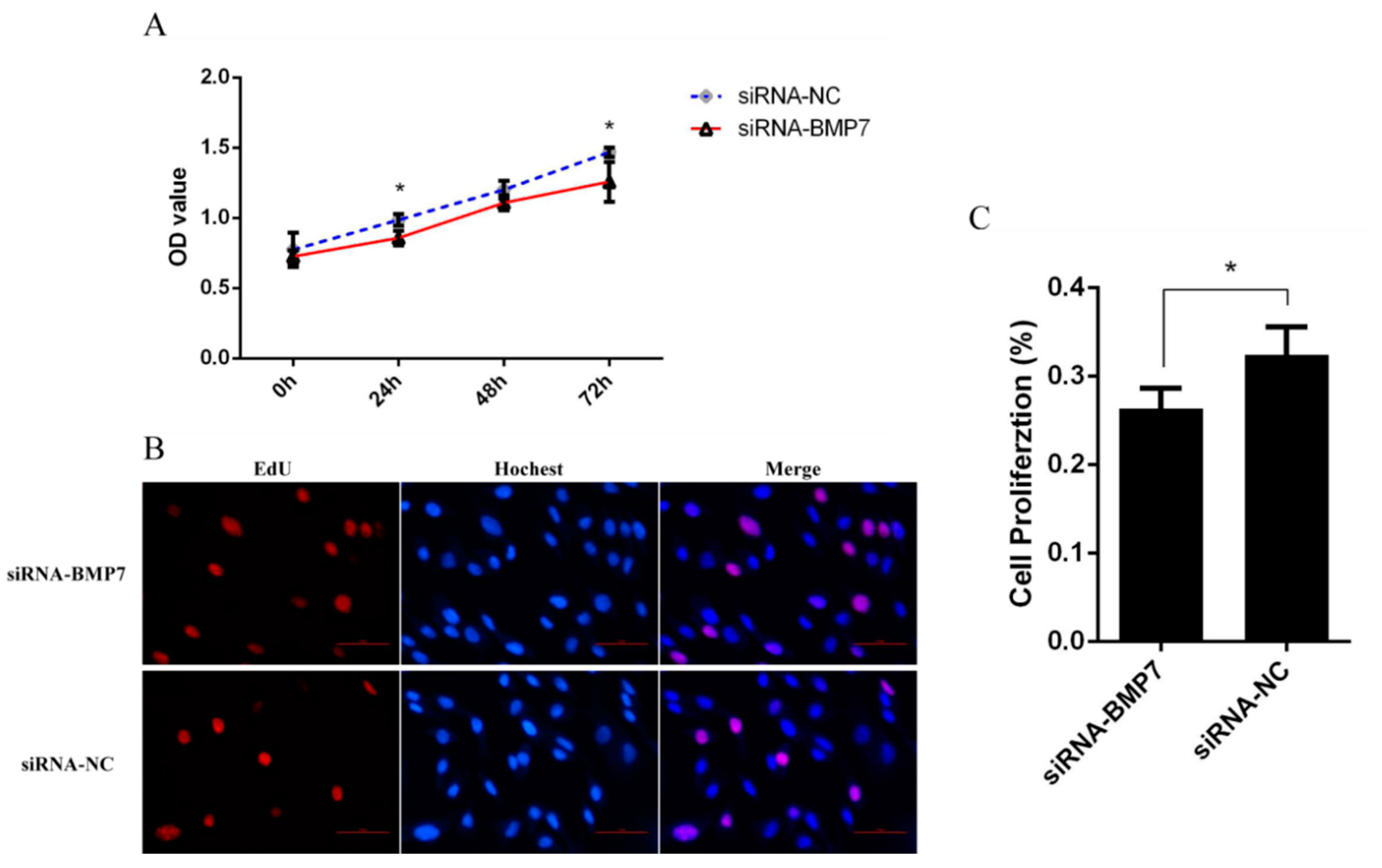
2.4. Cell Proliferation and Cycle Assay
2.5. qRT-PCR
2.6. Western Bolt
2.7. Statistical Analysis
3. Results
3.1. Cloning Hu Sheep BMP7 Full Length cDNA Sequence
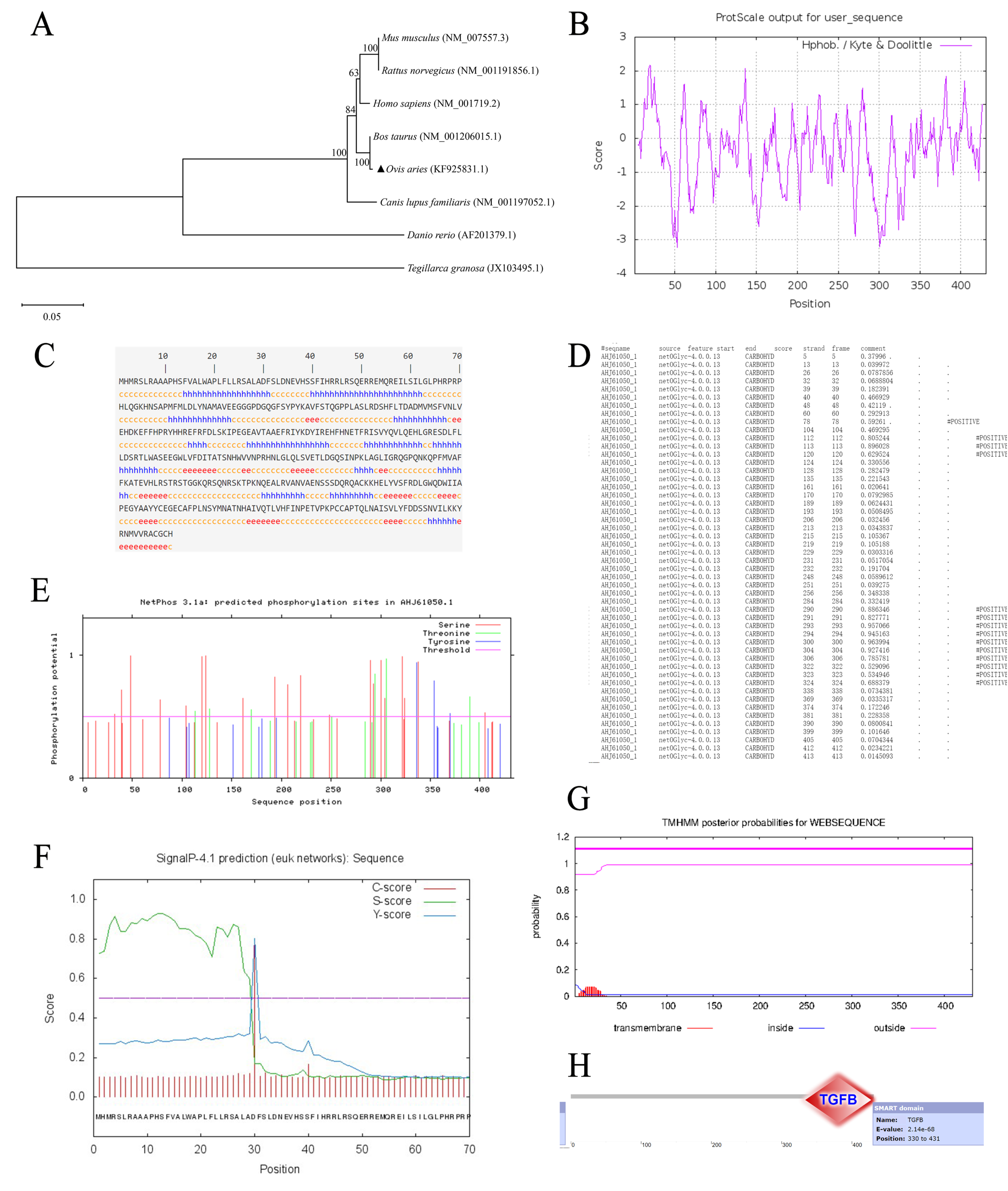
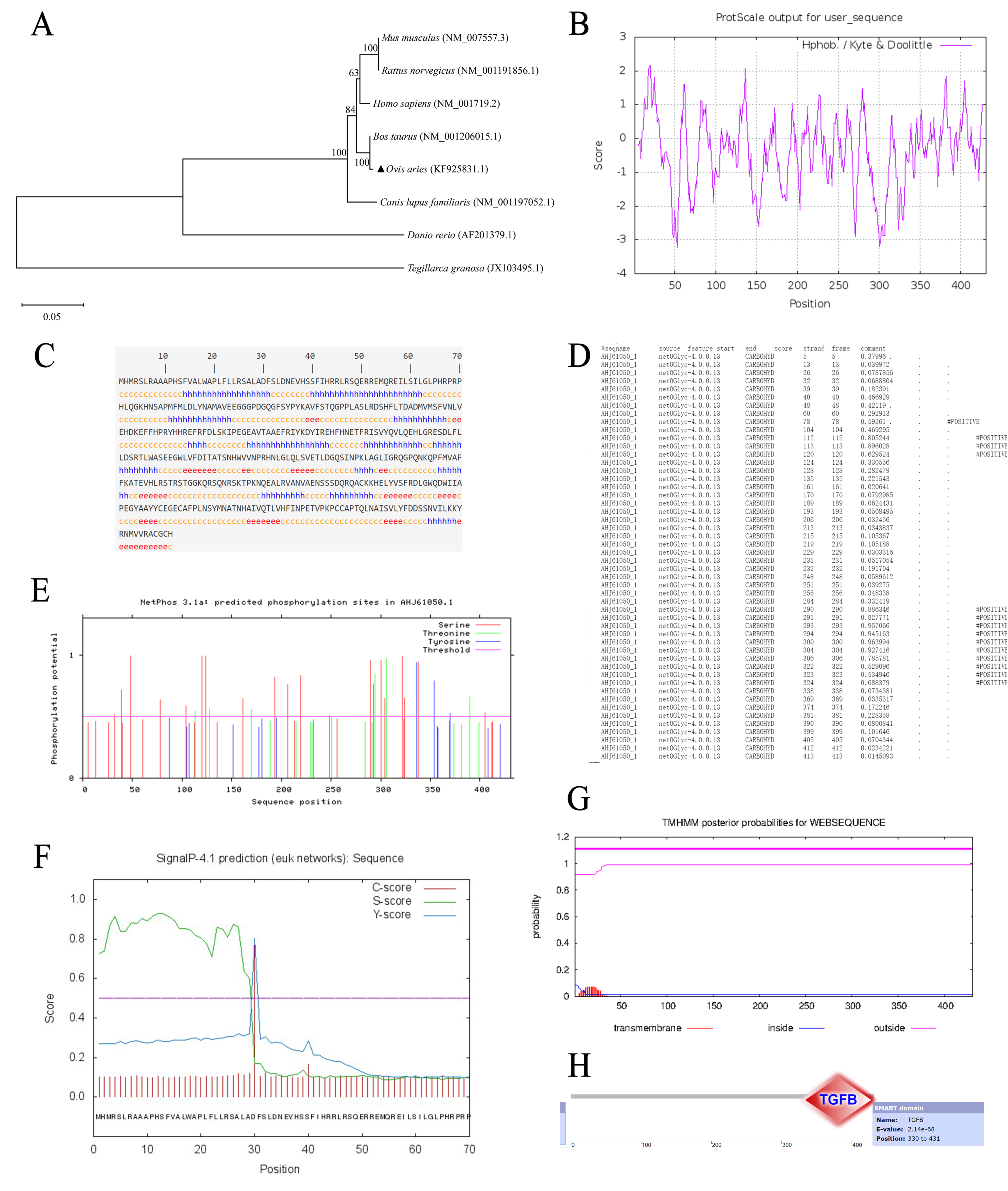
3.2. Bioinformatics Analysis of BMP7 Gene in Hu Sheep
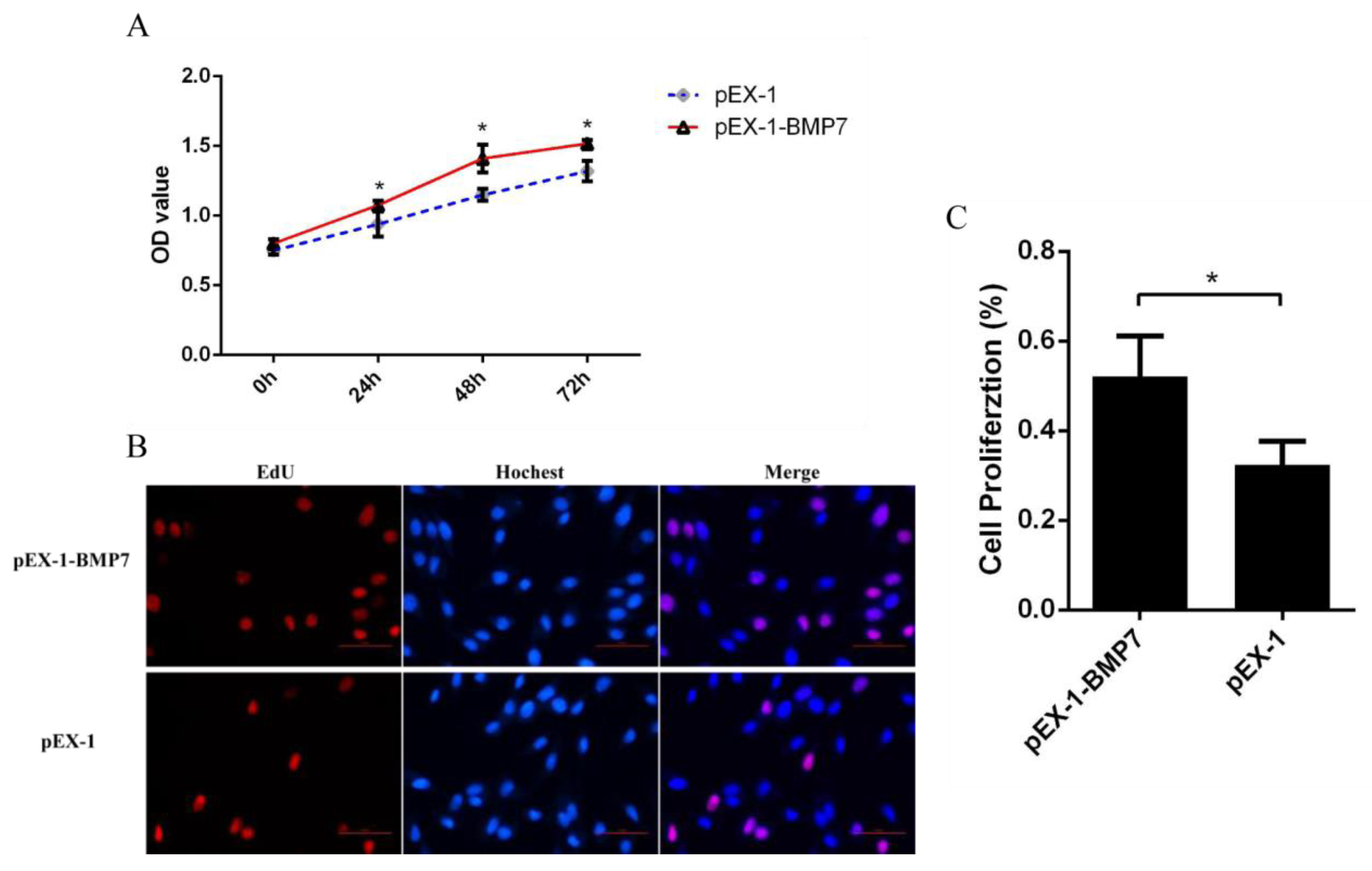
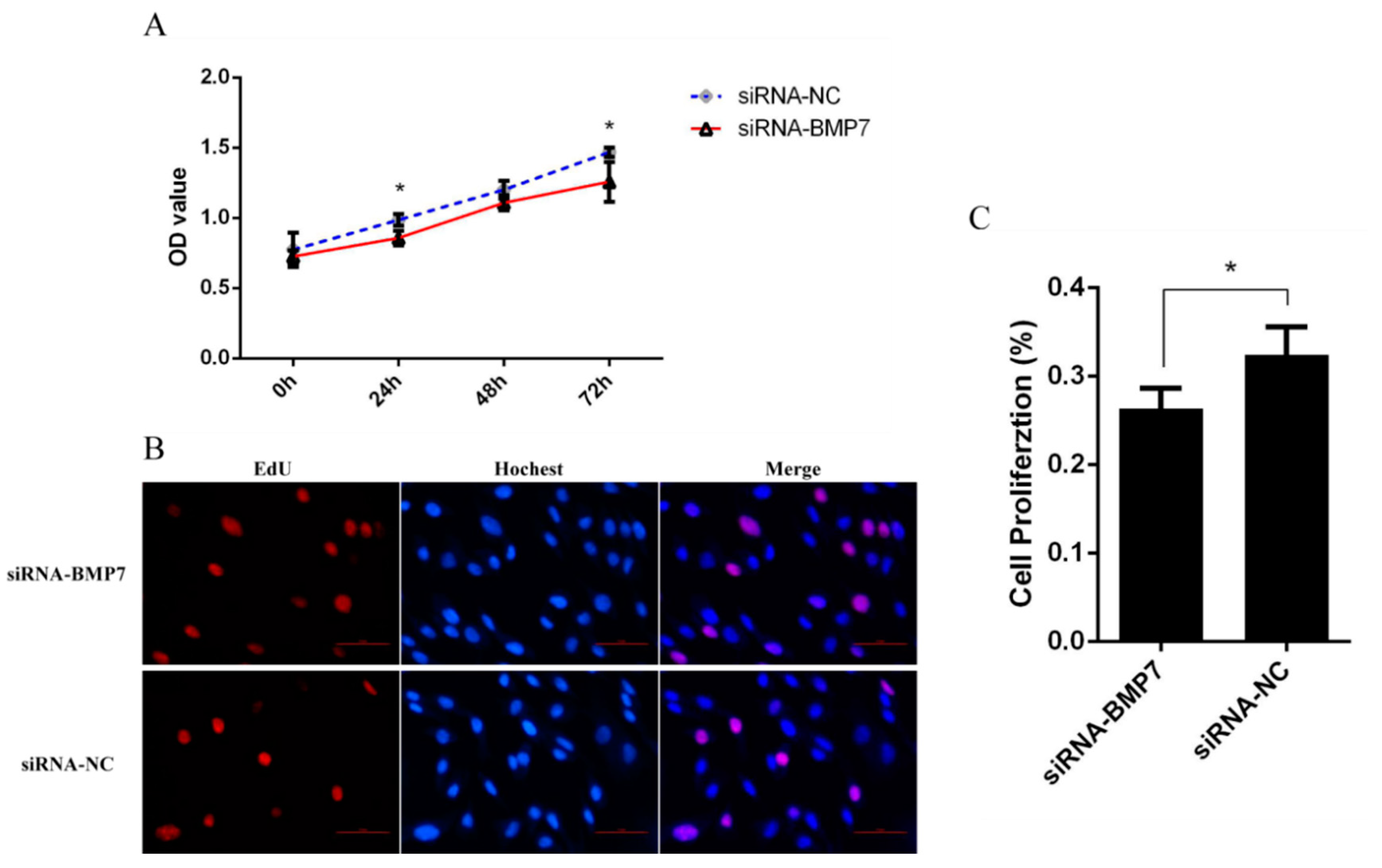
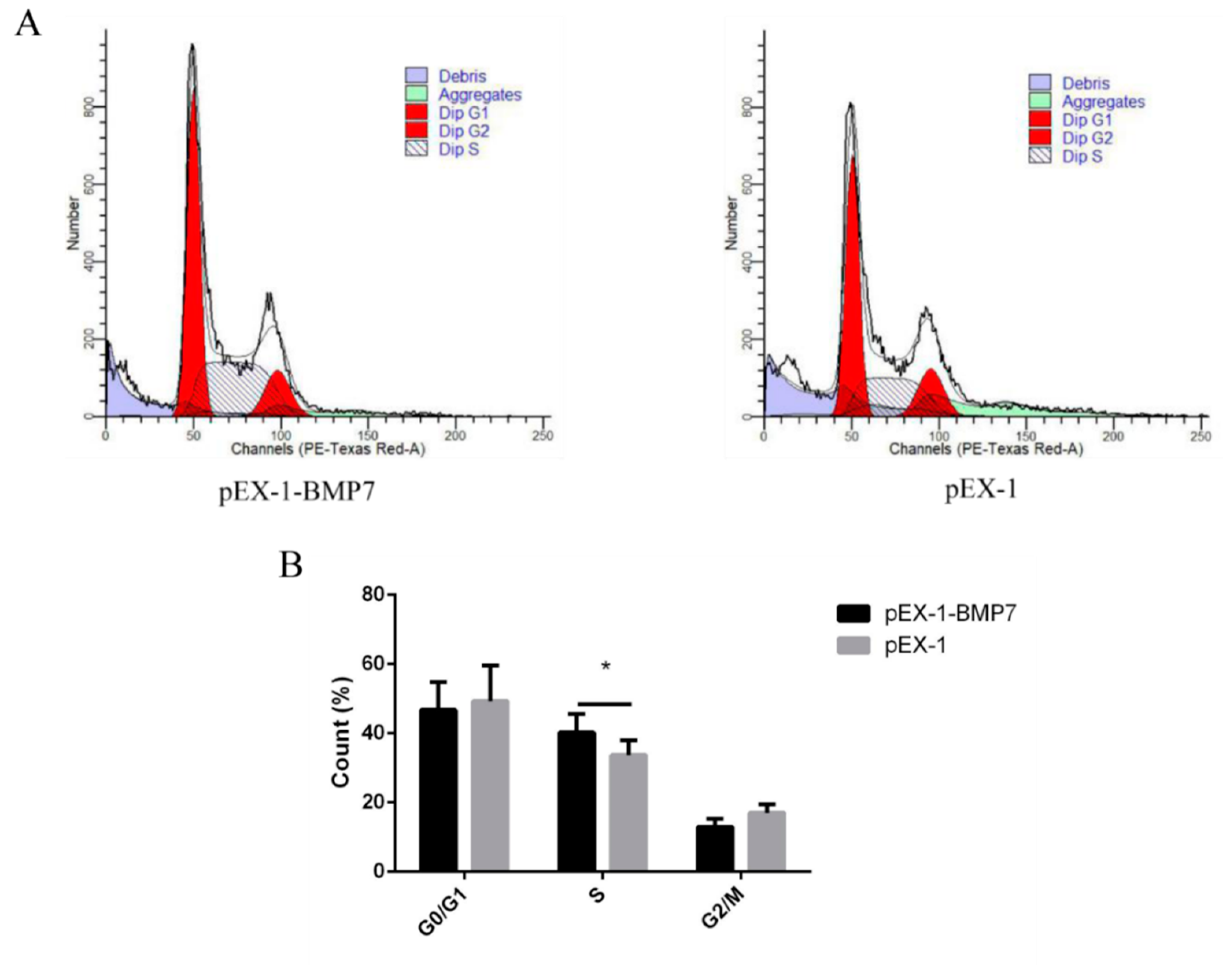
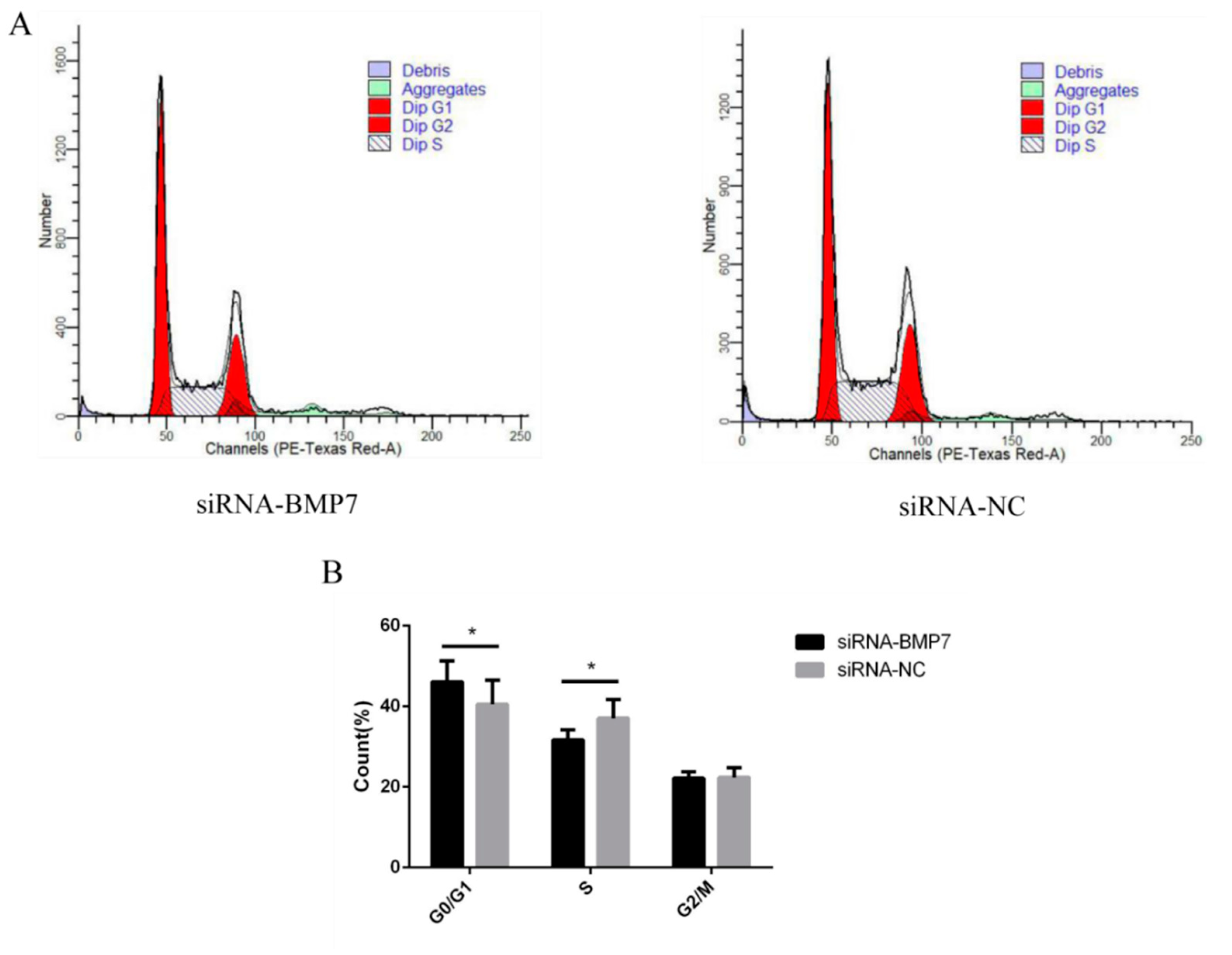
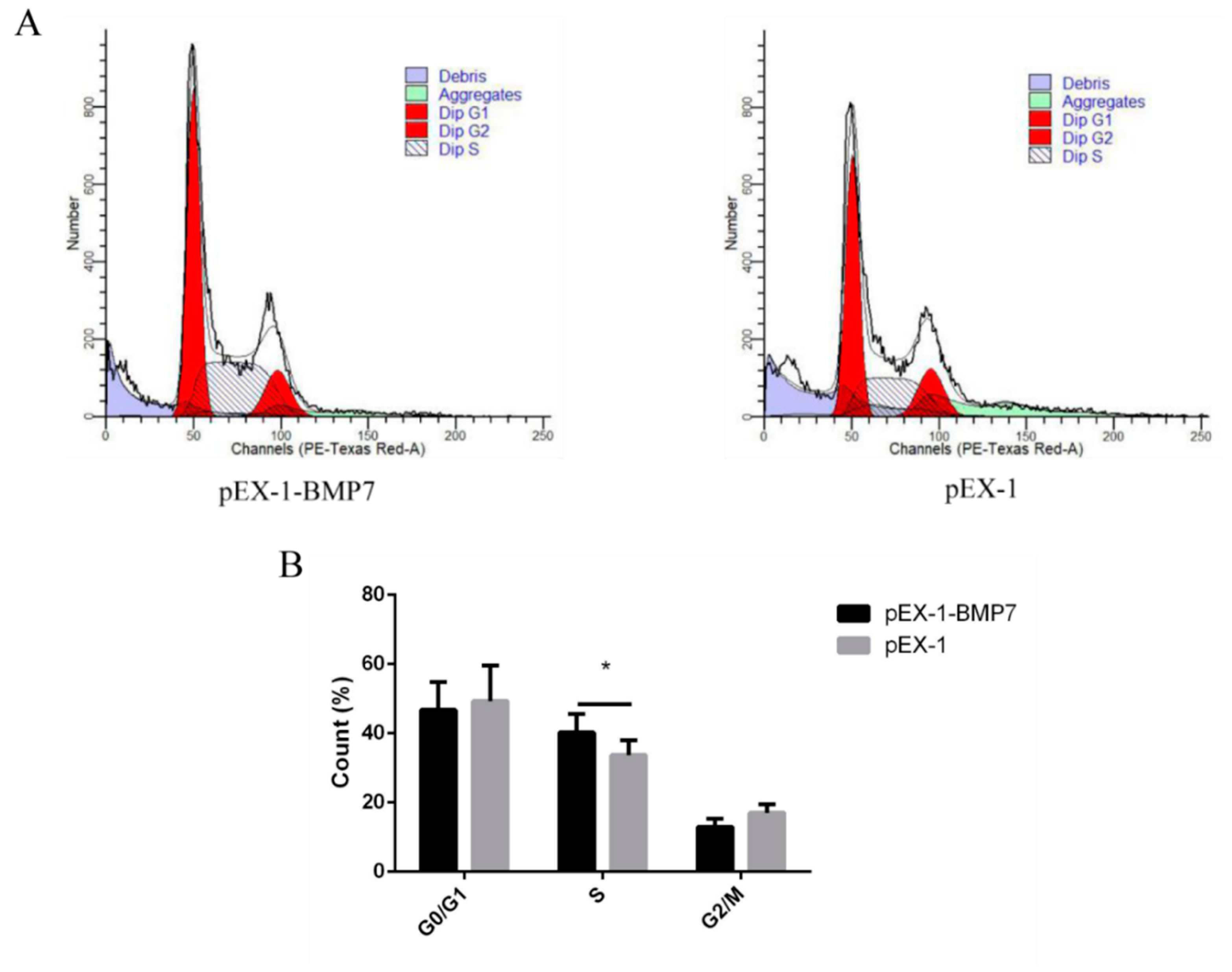
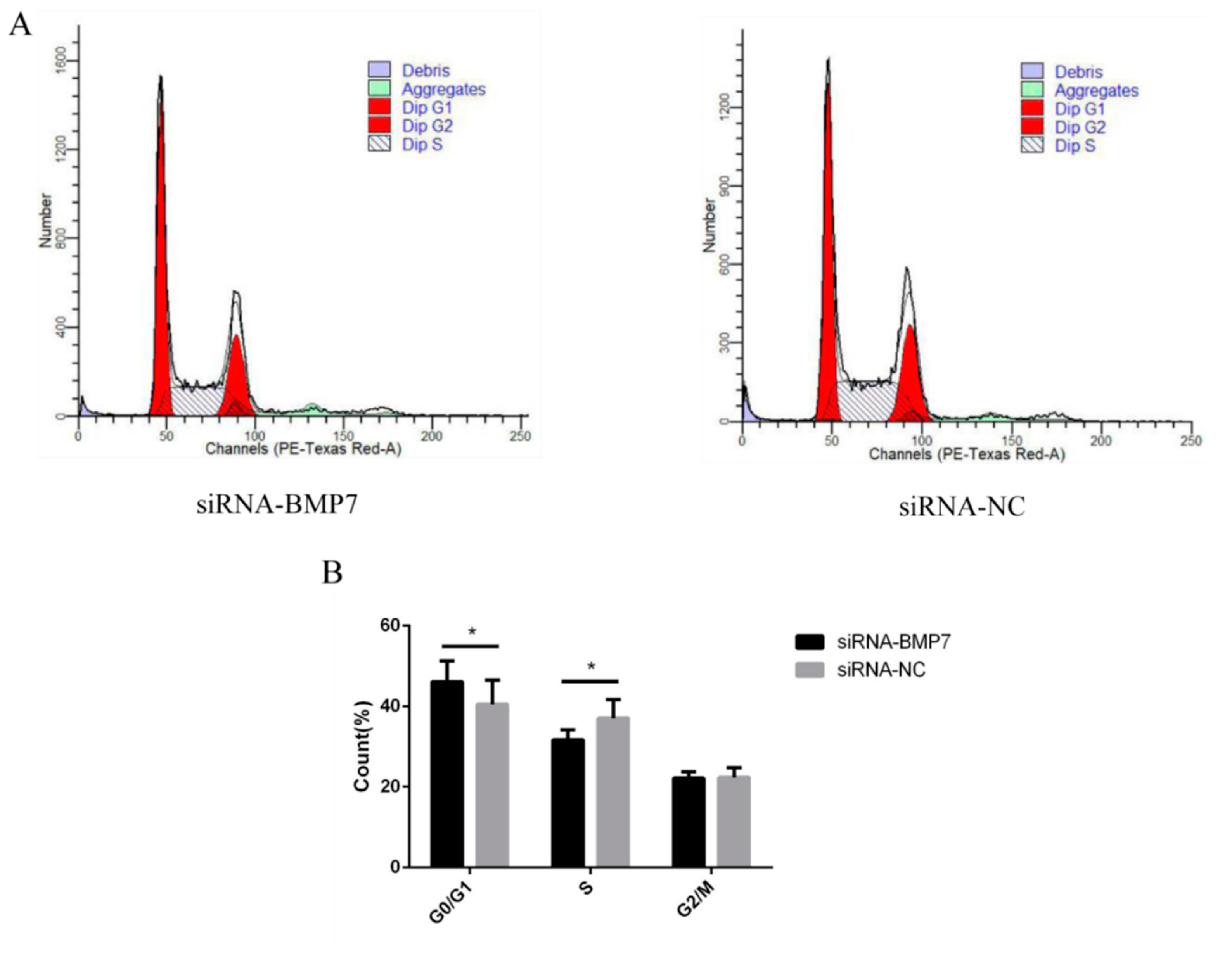
3.3. BMP7 Promoted Proliferation of Hu Sheep DPCs
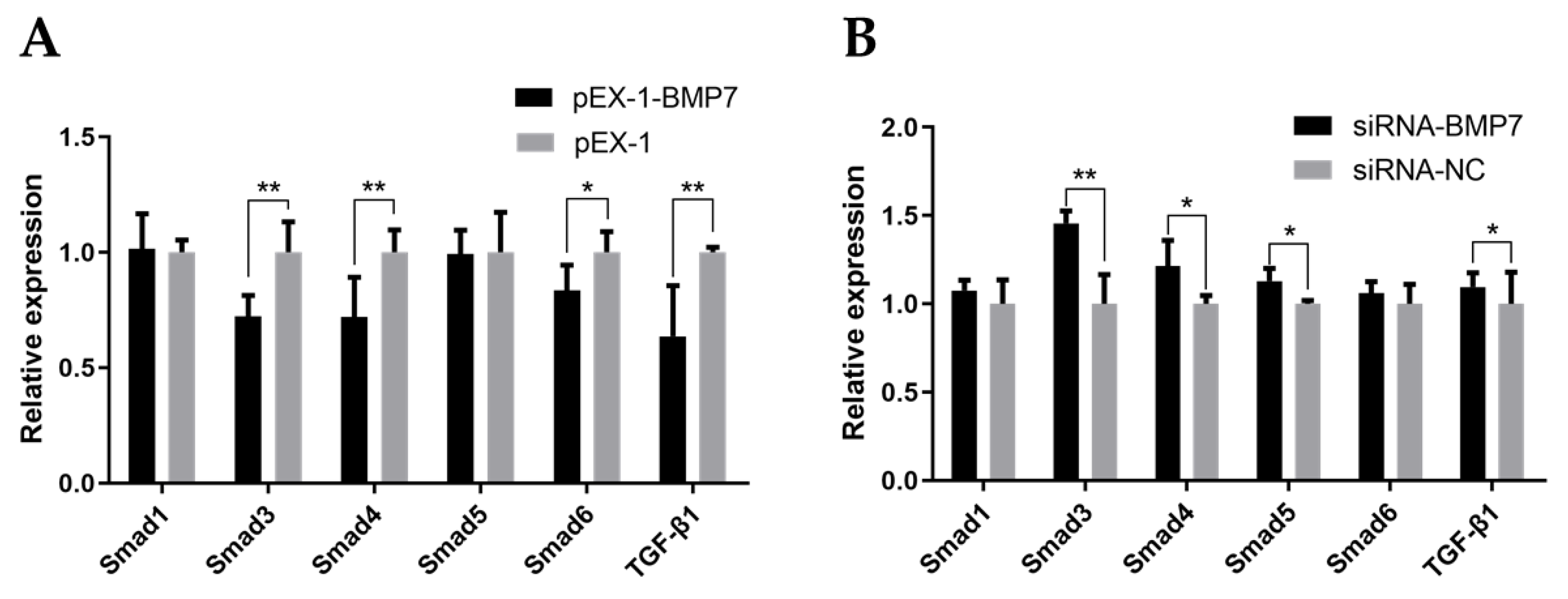
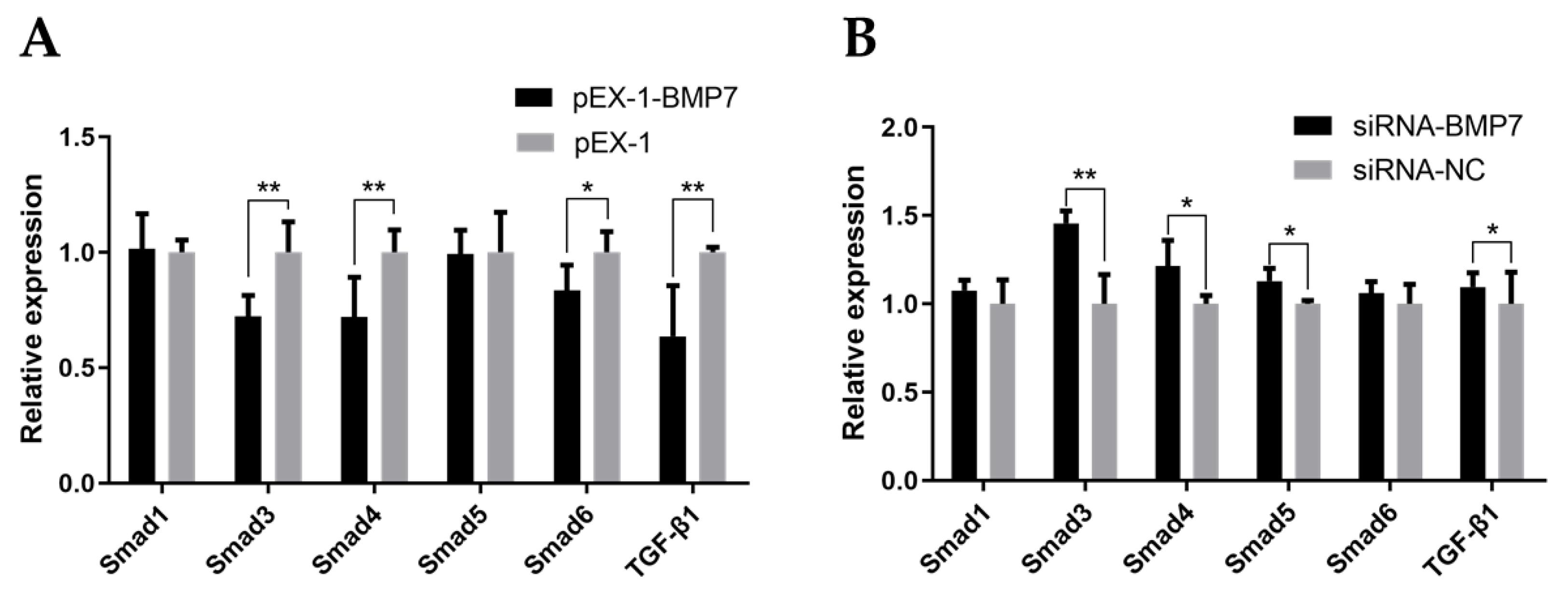
3.4. BMP7 Regulated Key Genes in TGF-β/Smad Signaling Pathway
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lv, X.; Chen, W.; Sun, W.; Hussain, Z.; Wang, S.; Wang, J. Analysis of lncRNAs Expression Profiles in Hair Follicle of Hu Sheep Lambskin. Animals 2020, 10, 1035. [Google Scholar] [CrossRef]
- Chi, W.; Wu, E.; Morgan, B.A. Dermal papilla cell number specifies hair size, shape and cycling and its reduction causes follicular decline. Development 2013, 140, 1676–1683. [Google Scholar] [CrossRef] [Green Version]
- Morgan, B.A. The dermal papilla: An instructive niche for epithelial stem and progenitor cells in development and regeneration of the hair follicle. Cold Spring Harb. Perspect Med. 2014, 4, a015180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driskell, R.R.; Giangreco, A.; Jensen, K.B.; Mulder, K.W.; Watt, F.M. Sox2-positive dermal papilla cells specify hair follicle type in mammalian epidermis. Development 2009, 136, 2815–2823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rompolas, P.; Deschene, E.R.; Zito, G.; Gonzalez, D.G.; Saotome, I.; Haberman, A.M.; Greco, V. Live imaging of stem cell and progeny behaviour in physiological hair-follicle regeneration. Nature 2012, 487, 496–499. [Google Scholar] [CrossRef]
- Wang, J.; Cui, K.; Yang, Z.; Li, T.; Hua, G.; Han, D.; Yao, Y.; Chen, J.; Deng, X.; Yang, X.; et al. Transcriptome Analysis of Improved Wool Production in Skin-Specific Transgenic Sheep Overexpressing Ovine β-Catenin. Int. J. Mol. Sci. 2019, 20, 620. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Zhang, Y.; Xing, Y.; Xu, W.; Guo, H.; Deng, F.; Ma, X.; Li, Y. The balance of Bmp6 and Wnt10b regulates the telogen-anagen transition of hair follicles. Cell Commun. Signal 2019, 17, 16. [Google Scholar] [CrossRef] [Green Version]
- Song, L.L.; Cui, Y.; Yu, S.J.; Liu, P.G.; Liu, J.; Yang, X.; He, J.F.; Zhang, Q. Expression characteristics of BMP2, BMPR-IA and Noggin in different stages of hair follicle in yak skin. Gen. Comp. Endocrinol. 2018, 260, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Zhang, J.Y.; Chu, M.X.; Piao, J.; Piao, J.A.; Zhao, F.Q. Cashmere growth control in Liaoning cashmere goat by ovarian carcinoma immunoreactive antigen-like protein 2 and decorin genes. Asian-Australas J. Anim. Sci. 2018, 31, 650–657. [Google Scholar] [CrossRef]
- AlShaibi, H.F.; Ahmed, F.; Buckle, C.; Fowles, A.C.M.; Awlia, J.; Cecchini, M.G.; Eaton, C.L. The BMP antagonist Noggin is produced by osteoblasts in response to the presence of prostate cancer cells. Biotechnol. Appl. Biochem. 2018, 65, 407–418. [Google Scholar] [CrossRef]
- Chen, G.; Xu, H.; Yao, Y.; Xu, T.; Yuan, M.; Zhang, X.; Lv, Z.; Wu, M. BMP Signaling in the Development and Regeneration of Cranium Bones and Maintenance of Calvarial Stem Cells. Front. Cell Dev. Biol. 2020, 8, 135. [Google Scholar] [CrossRef] [PubMed]
- Jensen, G.S.; Leon-Palmer, N.E.; Townsend, K.L. Bone morphogenetic proteins (BMPs) in the central regulation of energy balance and adult neural plasticity. Metab. Clin. Exp. 2021, 123, 154837. [Google Scholar] [CrossRef]
- Botchkarev, V.A.; Sharov, A.A. BMP signaling in the control of skin development and hair follicle growth. Differentiation 2004, 72, 512–526. [Google Scholar] [CrossRef]
- Botchkarev, V.A. Bone morphogenetic proteins and their antagonists in skin and hair follicle biology. J. Invest. Dermatol. 2003, 120, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Chen, Y.; Chen, M.; Zhang, Y. Overexpression of Fgf8 in the epidermis inhibits hair follicle development. Exp. Dermatol. 2021, 30, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, Y.; Bai, L.; Wang, Y.; Yang, L.; Su, W.; Gao, S. Heat stress decreased hair follicle population in rex rabbits. J. Anim. Physiol. Anim. Nutr. 2019, 103, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Noramly, S.; Morgan, B.A. BMPs mediate lateral inhibition at successive stages in feather tract development. Development 1998, 125, 3775–3787. [Google Scholar] [CrossRef]
- Ceruti, J.M.; Oppenheimer, F.M.; Leirós, G.J.; Balañá, M.E. Androgens downregulate BMP2 impairing the inductive role of dermal papilla cells on hair follicle stem cells differentiation. Mol. Cell Endocrinol. 2021, 520, 111096. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Zheng, Y.; Yan, J.; Wang, J.; Liu, X.; Yin, G. BMP2-mediated PTEN enhancement promotes differentiation of hair follicle stem cells by inducing autophagy. Exp. Cell Res. 2019, 385, 111647. [Google Scholar] [CrossRef] [PubMed]
- Esibizione, D.; Cui, C.Y.; Schlessinger, D. Candidate EDA targets revealed by expression profiling of primary keratinocytes from Tabby mutant mice. Gene 2008, 427, 42–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adly, M.A.; Assaf, H.A.; Hussein, M.R. Expression of bone morphogenetic protein-7 in human scalp skin and hair follicles. Br. J. Dermatol. 2006, 154, 551–554. [Google Scholar] [CrossRef] [PubMed]
- Shimasaki, S.; Moore, R.K.; Otsuka, F.; Erickson, G.F. The bone morphogenetic protein system in mammalian reproduction. Endocr. Rev. 2004, 25, 72–101. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.J.; Wu, J.C.; Su, P.; Zhirnov, O.; Miller, W.L. A novel role for bone morphogenetic proteins in the synthesis of follicle-stimulating hormone. Endocrinology 2001, 142, 2275–2283. [Google Scholar] [CrossRef] [PubMed]
- Glister, C.; Kemp, C.F.; Knight, P.G. Bone morphogenetic protein (BMP) ligands and receptors in bovine ovarian follicle cells: Actions of BMP-4, -6 and -7 on granulosa cells and differential modulation of Smad-1 phosphorylation by follistatin. Reproduction 2004, 127, 239–254. [Google Scholar] [CrossRef]
- Erickson, G.F.; Fuqua, L.; Shimasaki, S. Analysis of spatial and temporal expression patterns of bone morphogenetic protein family members in the rat uterus over the estrous cycle. J. Endocrinol. 2004, 182, 203–217. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Ni, R.; Yin, J.F.; Musa, H.H.; Ding, T.; Chen, L. Genome array of hair follicle genes in lambskin with different patterns. PLoS ONE 2013, 8, e68840. [Google Scholar] [CrossRef] [Green Version]
- Lv, X.; Gao, W.; Jin, C.; Wang, L.; Wang, Y.; Chen, W.; Zou, S.; Huang, S.; Li, Z.; Wang, J.; et al. Preliminary study on microR-148a and microR-10a in dermal papilla cells of Hu sheep. BMC Genet. 2019, 20, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Du, L.; Liu, R.; Di, R.; Zhang, L.; Ma, Y.; Li, Q.; Liu, E.; Chu, M.; Wei, C. MiR-378 and BMP-Smad can influence the proliferation of sheep myoblast. Gene 2018, 674, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hynd, P.I.; Edwards, N.M.; Hebart, M.; McDowall, M.; Clark, S. Wool fibre crimp is determined by mitotic asymmetry and position of final keratinisation and not ortho- and para-cortical cell segmentation. Animal 2009, 3, 838–843. [Google Scholar] [CrossRef]
- Kwack, M.H.; Seo, C.H.; Gangadaran, P.; Ahn, B.C.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Exosomes derived from human dermal papilla cells promote hair growth in cultured human hair follicles and augment the hair-inductive capacity of cultured dermal papilla spheres. Exp. Dermatol. 2019, 28, 854–857. [Google Scholar] [CrossRef]
- Nissimov, J.N.; Das Chaudhuri, A.B. Hair curvature: A natural dialectic and review. Biol. Rev. Camb. Philos. Soc. 2014, 89, 723–766. [Google Scholar] [CrossRef]
- Mishina, Y. Function of bone morphogenetic protein signaling during mouse development. Front. Biosci. 2003, 8, d855–d869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, W.L.; Dang, Y.L.; Wang, J.J.; Yin, R.H.; Wang, Z.Y.; Zhu, Y.B.; Cong, Y.Y.; Xue, H.L.; Deng, L.; Guo, D.; et al. Molecular characterization, expression and methylation status analysis of BMP4 gene in skin tissue of Liaoning cashmere goat during hair follicle cycle. Genetica 2016, 144, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Andl, T.; Ahn, K.; Kairo, A.; Chu, E.Y.; Wine-Lee, L.; Reddy, S.T.; Croft, N.J.; Cebra-Thomas, J.A.; Metzger, D.; Chambon, P.; et al. Epithelial Bmpr1a regulates differentiation and proliferation in postnatal hair follicles and is essential for tooth development. Development 2004, 131, 2257–2268. [Google Scholar] [CrossRef] [Green Version]
- Jamora, C.; DasGupta, R.; Kocieniewski, P.; Fuchs, E. Links between signal transduction, transcription and adhesion in epithelial bud development. Nature 2003, 422, 317–322. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Ikeda, T. Transcripts for two members of the transforming growth factor-beta superfamily BMP-3 and BMP-7 are expressed in developing rat embryos. Dev. Dyn. 1996, 207, 439–449. [Google Scholar] [CrossRef]
- Zouvelou, V.; Luder, H.U.; Mitsiadis, T.A.; Graf, D. Deletion of BMP7 affects the development of bones, teeth, and other ectodermal appendages of the orofacial complex. J. Exp. Zool. B Mol. Dev. Evol. 2009, 312B, 361–374. [Google Scholar] [CrossRef]
- Song, D.; Liu, X.; Liu, R.; Yang, L.; Zuo, J.; Liu, W. Connexin 43 hemichannel regulates H9c2 cell proliferation by modulating intracellular ATP and [Ca2+]. Acta Biochim. Biophys. Sin. 2010, 42, 472–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Zhang, P.J.; Chen, T.; Yum, S.W.; Pasha, T.; Furth, E.E. Connexin43 Expression Increases in the Epithelium and Stroma along the Colonic Neoplastic Progression Pathway: Implications for Its Oncogenic Role. Gastroenterol. Res. Pract. 2011, 2011, 561719. [Google Scholar] [CrossRef] [Green Version]
- Medland, S.E.; Nyholt, D.R.; Painter, J.N.; McEvoy, B.P.; McRae, A.F.; Zhu, G.; Gordon, S.D.; Ferreira, M.A.; Wright, M.J.; Henders, A.K.; et al. Common variants in the trichohyalin gene are associated with straight hair in Europeans. Am. J. Hum. Genet. 2009, 85, 750–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, T.; Gambardella, L.; Horcher, M.; Tschanz, S.; Capol, J.; Bertram, P.; Jochum, W.; Barrandon, Y.; Busslinger, M. The transcriptional repressor CDP (Cutl1) is essential for epithelial cell differentiation of the lung and the hair follicle. Genes Dev. 2001, 15, 2307–2319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeisberg, M.; Hanai, J.; Sugimoto, H.; Mammoto, T.; Charytan, D.; Strutz, F.; Kalluri, R. BMP-7 counteracts TGF-beta1-induced epithelial-to-mesenchymal transition and reverses chronic renal injury. Nat. Med. 2003, 9, 964–968. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene name | Primer Name | Sequences (5′→3′) | Length (bp) |
|---|---|---|---|
| BMP7 | 1F | TAGAGCCGGCGCGATGCAC | 478 |
| 1R | ACGCTACCACCACCGGGAG | ||
| 3O | CCTATCCCTACAAGGCCGTCT | 1466 | |
| 3I | ATCACAGCCACCAGCAACCAC | 1095 |
| Name | Sequences (5′→3′) |
|---|---|
| siRNA-124 | sense: GCACUCGAGCUUCAUCCAUTT |
| antisense: AUGGAUGAAGCUCGAGUGCTT | |
| siRNA-469 | sense: CCACCCGGGAGUUCCGGUUUTT |
| antisense: AAACCGGAACUCCCGGUGGTT | |
| siRNA-556 | sense: CCGGGAACACUUCCACAAUTT |
| antisense: AUUGUGGAAGUGUUCCCGGTT |
| Gene Name | Sequences(5′→3′) (F: Forward R: Reverse) | Length (bp) | Tm (°C) | Accession No. |
|---|---|---|---|---|
| BMP7 | F: TGAGTTCCGCATTTACAAGG | 177 | 60 | KF925831.1 |
| R: GTGGCTGTGATGTCAAAAAC | ||||
| Smad1 | F: GAAAGCCCCGTTCTTCCTCC | 150 | 60 | AY035385.1 |
| R: GTTGGGCTGCTGGAAAGAAT | ||||
| Smad3 | F: GGACGACTACAGCCATTCCA | 172 | 60 | AF508024.1 |
| R: ATTCGGGGAGAGGTTTGGAG | ||||
| Smad4 | F: CTTCAGGTGGCTGGTCGG | 177 | 60 | NM_001267886.1 |
| R: TCCAGGTGATACAACTCGTTCA | ||||
| Smad5 | F: CCAGTATATCCAGCAGAGATGTT | 102 | 60 | AF508027.1 |
| R: AAGCTTCCCCAACACGATTG | ||||
| Smad6 | F: CTGCTCGGACGCCTCTTC | 105 | 60 | XM_004010255.5 |
| R: GGGTGGCGGTGATTCTGG | ||||
| TGF-β1 | F: GGTGGAATACGGCAACAAAATC | 162 | 60 | NM_001009400.2 |
| R: TGCTGCTCCACTTTTAACTTGA | ||||
| GAPDH | F: GTCGGAGTGAACGGATTTGG | 196 | 60 | NM_001190390.1 |
| R: CATTGATGACGAGCTTCCCG |
| Species | Accession No. | Nucleotide (%) | Amino Acid (%) |
|---|---|---|---|
| Homo sapiens | NM_001719.2 | 92.7% | 98.7% |
| Bos taurus | NM_001206015.1 | 98.5% | 99.4% |
| Mus musculus | NM_007557.3 | 89% | 98.9% |
| Rattus norvegicus | NM_001191856.1 | 88% | 98.9% |
| Danio rerio | AF201379.1 | 68% | 88.4% |
| Tegillarca granosa | JX103495.1 | 48.7% | 79.5% |
| Canis lupus familiaris | NM_001197052.1 | 92.1% | 98.5% |
| Name | Length | mTP | SP | Other |
|---|---|---|---|---|
| AHJ61050.1 | 431 | 0.327 | 0.858 | 0.015 |
| Functional Category | Odds |
|---|---|
| Amino acid biosynthesis | 0.500 |
| Biosynthesis of cofactors | 2.917 |
| Cell envelope | 0.541 |
| Cellular processes | 0.411 |
| Central intermediary metabolism | 0.762 |
| Energy metabolism | 0.389 |
| Purines and pyrimidines | 1.308 |
| Fatty acid metabolism | 1.362 |
| Replication and transcription | 0.211 |
| Regulatory functions | 0.075 |
| Translation | 1.614 |
| Transport and binding | 1.885 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Lv, X.; Wang, S.; Cao, X.; Yuan, Z.; Getachew, T.; Mwacharo, J.M.; Haile, A.; Sun, W. BMP7 Functions to Regulate Proliferation of Dermal Papilla Cells in Hu Sheep. Genes 2022, 13, 201. https://doi.org/10.3390/genes13020201
Li Y, Lv X, Wang S, Cao X, Yuan Z, Getachew T, Mwacharo JM, Haile A, Sun W. BMP7 Functions to Regulate Proliferation of Dermal Papilla Cells in Hu Sheep. Genes. 2022; 13(2):201. https://doi.org/10.3390/genes13020201
Chicago/Turabian StyleLi, Yue, Xiaoyang Lv, Shanhe Wang, Xiukai Cao, Zehu Yuan, Tesfaye Getachew, Joram M. Mwacharo, Aynalem Haile, and Wei Sun. 2022. "BMP7 Functions to Regulate Proliferation of Dermal Papilla Cells in Hu Sheep" Genes 13, no. 2: 201. https://doi.org/10.3390/genes13020201
APA StyleLi, Y., Lv, X., Wang, S., Cao, X., Yuan, Z., Getachew, T., Mwacharo, J. M., Haile, A., & Sun, W. (2022). BMP7 Functions to Regulate Proliferation of Dermal Papilla Cells in Hu Sheep. Genes, 13(2), 201. https://doi.org/10.3390/genes13020201