
Pou5f1 and Nanog Are Reliable Germ Cell-Specific Genes in Gonad of a Protogynous Hermaphroditic Fish, Orange-Spotted Grouper (Epinephelus coioides)

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Ethics
2.2. Cloning of Full-Length cDNA Sequences
2.3. Total RNA Extraction and PCR
2.4. Western Blotting
2.5. In Situ Hybridization (ISH)
2.6. Dual-Label ISH
2.7. Fluorescent Immunostaining
2.8. Statistical Analysis
3. Results
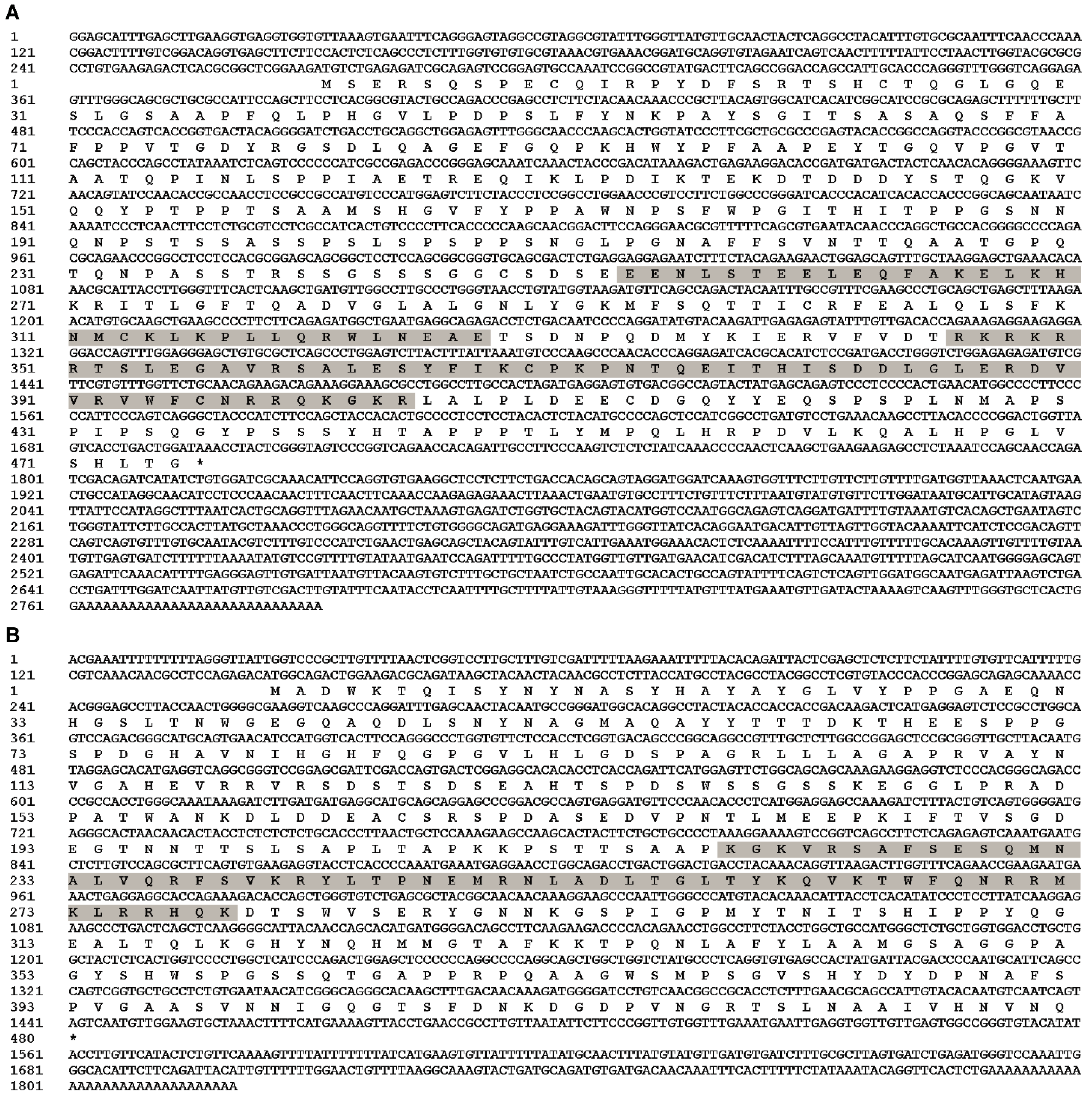
3.1. Identifications of EcPou5f1 and EcNanog
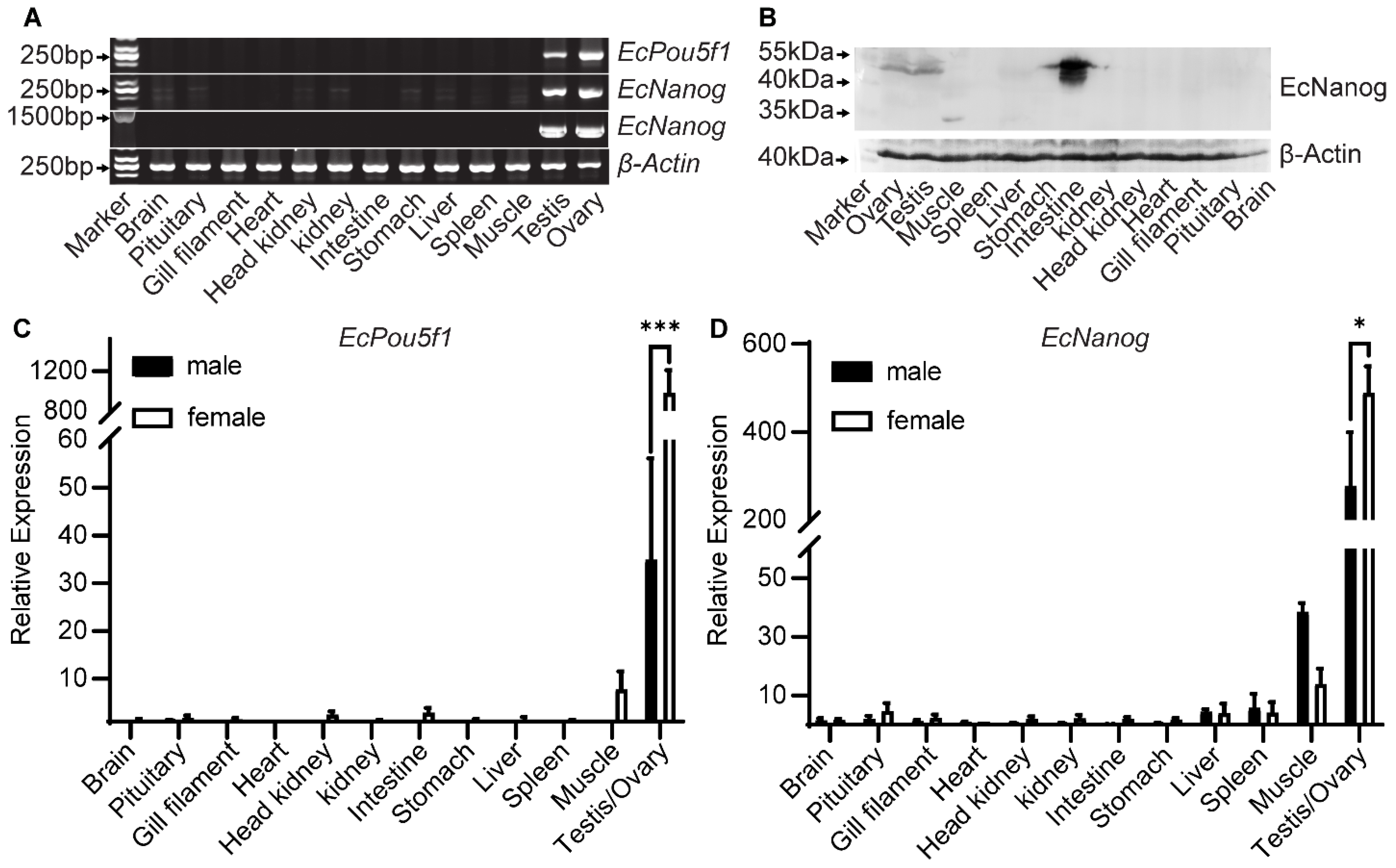
3.2. Tissue Distributions of EcPou5f1 and EcNanog
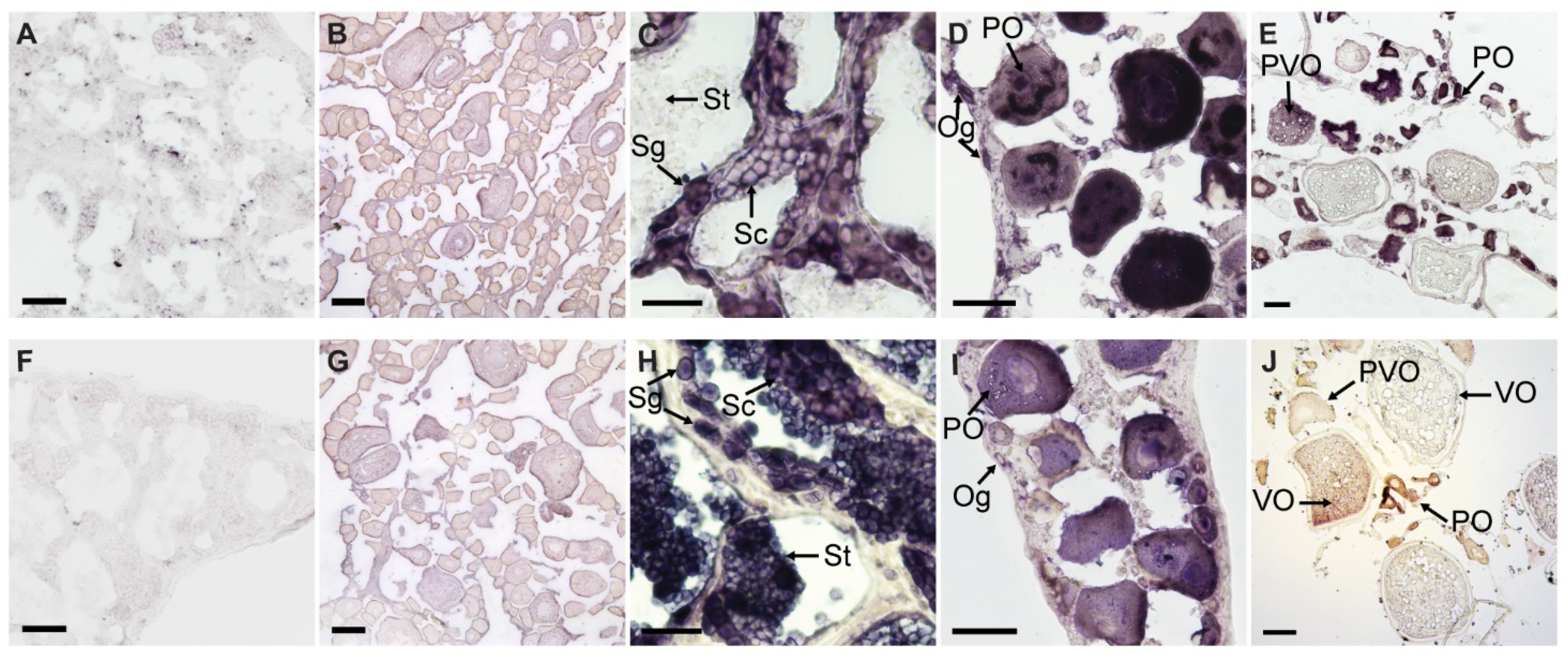
3.3. Chemical ISH of EcPou5f1 and EcNanog in Gonads
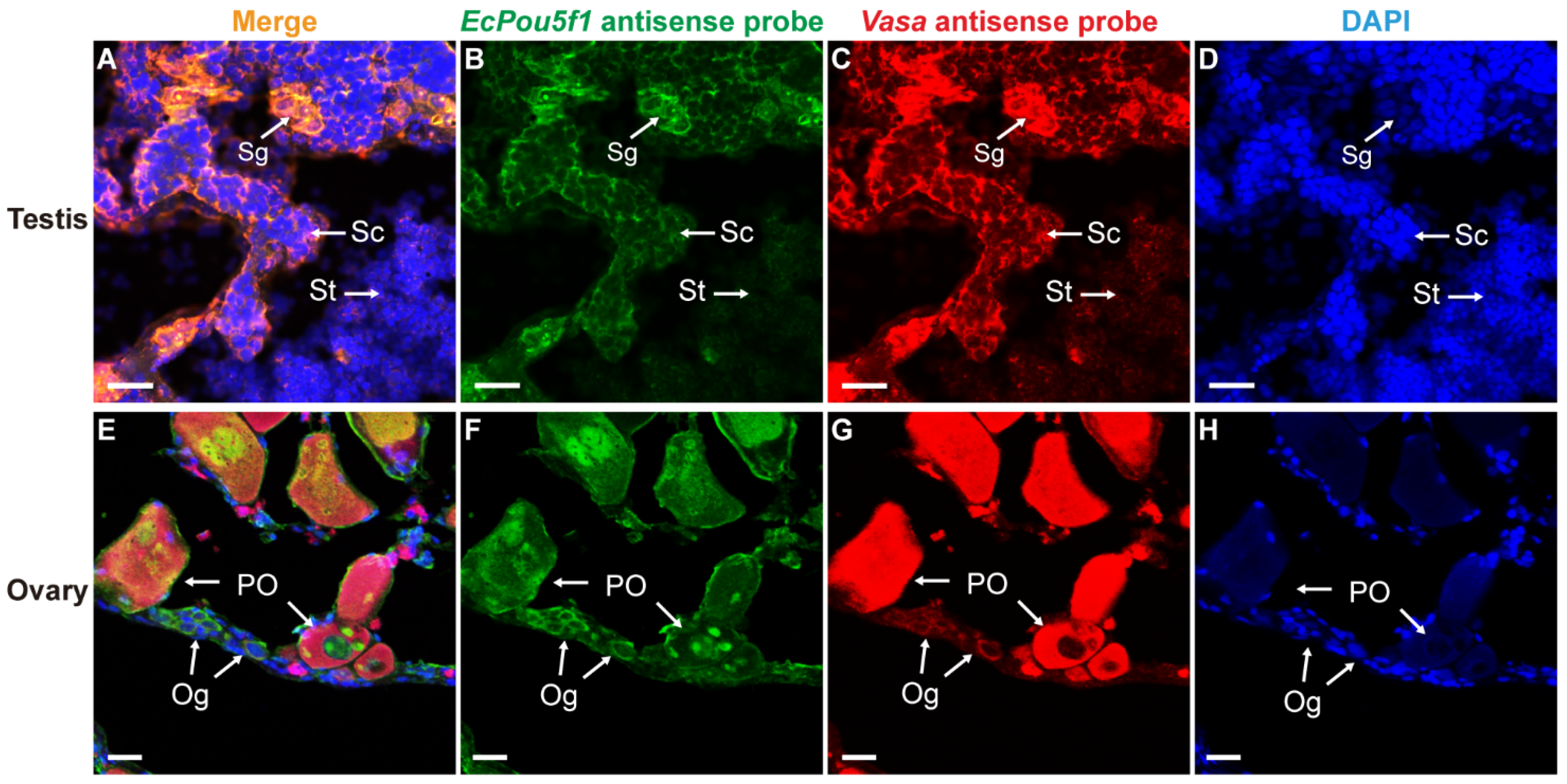
3.4. Dual-Label ISH for EcPou5f1 or EcNanog with Vasa in Gonads
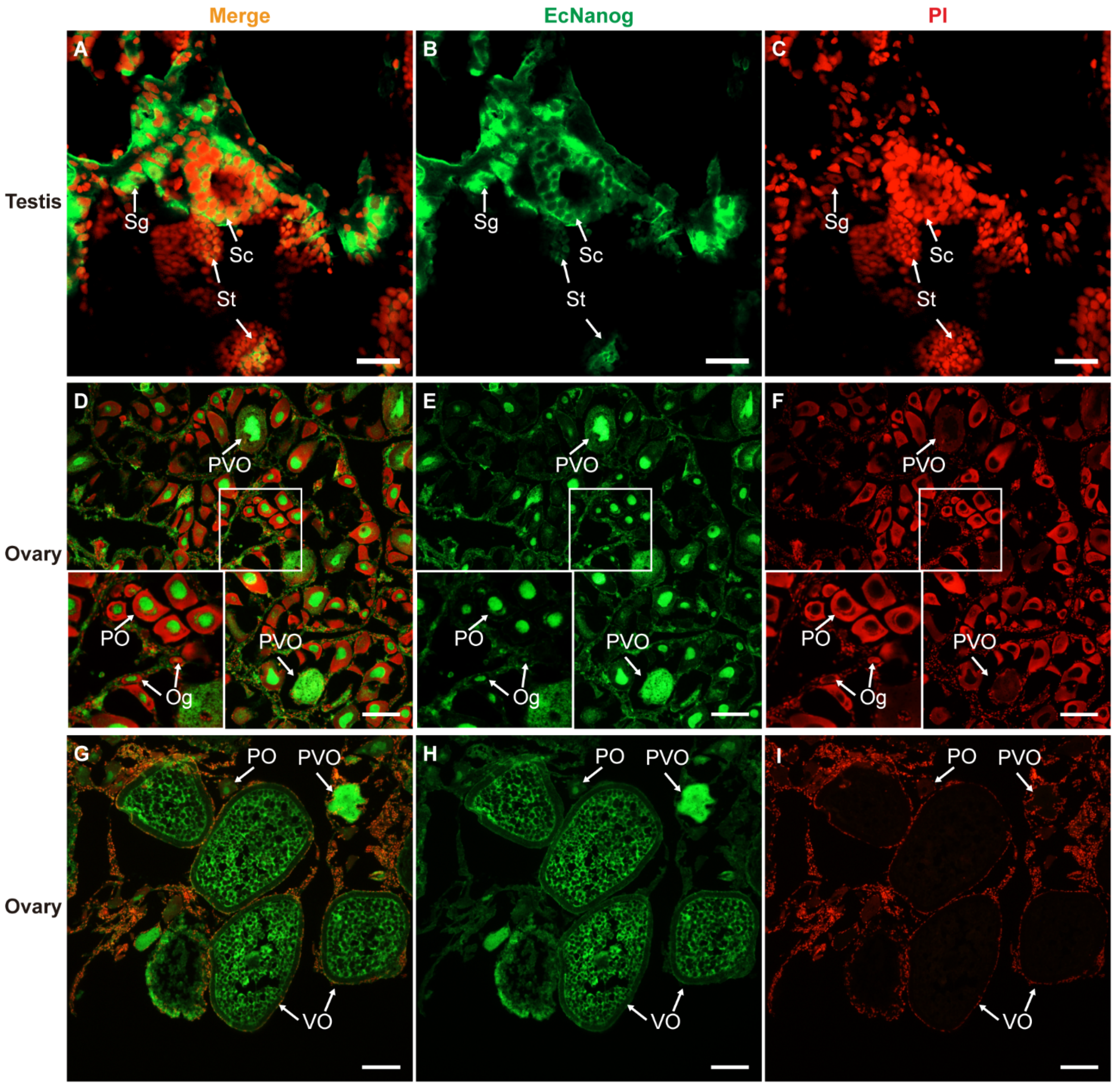
3.5. Immunohistofluorescence of Anti-Nanog Antibody in Gonads
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williamson, A.; Lehmann, R. Germ cell development in Drosophila. Annu. Rev. Cell Dev. Biol. 1996, 12, 365–391. [Google Scholar] [CrossRef]
- Dodson, A.E.; Kennedy, S. Phase Separation in Germ Cells and Development. Dev. Cell 2020, 55, 4–17. [Google Scholar] [CrossRef]
- Robles, V.; Riesco, M.F.; Psenicka, M.; Saito, T.; Valcarce, D.G.; Cabrita, E.; Herraez, P. Biology of teleost primordial germ cells (PGCs) and spermatogonia: Biotechnological applications. Aquaculture 2017, 472, 4–20. [Google Scholar] [CrossRef]
- Sun, Z.H.; Wang, Y.; Lu, W.J.; Li, Z.; Liu, X.C.; Li, S.S.; Zhou, L.; Gui, J.F. Divergent Expression Patterns and Function Implications of Four nanos Genes in a Hermaphroditic Fish, Epinephelus coioides. Int. J. Mol. Sci. 2017, 18, 685. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, Q.H.; Ma, D.Y.; Song, Z.C.; Li, J. A potential germ cell-specific marker in Japanese flounder, Paralichthys olivaceus: Identification and characterization of lymphocyte antigen 75 (Ly75/CD205). J. Oceanol. Limnol. 2018, 36, 1342–1348. [Google Scholar] [CrossRef]
- Wu, X.; Yang, Y.; Zhong, C.; Guo, Y.; Li, S.; Lin, H.; Liu, X. Transcriptome profiling of laser-captured germ cells and functional characterization of zbtb40 during 17alpha-methyltestosterone-induced spermatogenesis in orange-spotted grouper (Epinephelus coioides). BMC Genom. 2020, 21, 73. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, Y.; Saito, K.; Shinya, M.; Kawasaki, T.; Sakai, N. Evaluation of Sycp3, Plzf and Cyclin B3 expression and suitability as spermatogonia and spermatocyte markers in zebrafish. Gene Expr. Patterns 2011, 11, 309–315. [Google Scholar] [CrossRef]
- Wu, X.; Qu, L.; Li, S.; Guo, Y.; He, J.; Liu, M.; Liu, X.; Lin, H. Molecular characterization and expression patterns of stem-loop binding protein (SLBP) genes in protogynous hermaphroditic grouper, Epinephelus coioides. Gene 2019, 700, 120–130. [Google Scholar] [CrossRef]
- Qu, L.; Wu, X.; Liu, M.; Zhong, C.; Xu, H.; Li, S.; Lin, H.; Liu, X. Identification and characterization of germ cell genes vasa and dazl in a protogynous hermaphrodite fish, orange-spotted grouper (Epinephelus coioides). Gene Expr. Patterns 2020, 35, 119095. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Duan, J.; Cheng, N.; Nagahama, Y. Specific expression of Olpiwi1 and Olpiwi2 in medaka (Oryzias latipes) germ cells. Biochem. Biophys. Res. Commun. 2012, 418, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Pesce, M.; Scholer, H.R. Oct-4: Gatekeeper in the beginnings of mammalian development. Stem Cells 2001, 19, 271–278. [Google Scholar] [CrossRef]
- Chambers, I.; Colby, D.; Robertson, M.; Nichols, J.; Lee, S.; Tweedie, S.; Smith, A. Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell 2003, 113, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Loh, Y.H.; Wu, Q.; Chew, J.L.; Vega, V.B.; Zhang, W.; Chen, X.; Bourque, G.; George, J.; Leong, B.; Liu, J.; et al. The Oct4 and Nanog transcription network regulates pluripotency in mouse embryonic stem cells. Nat. Genet. 2006, 38, 431–440. [Google Scholar] [CrossRef]
- Scholer, H.R.; Balling, R.; Hatzopoulos, A.K.; Suzuki, N.; Gruss, P. Octamer binding proteins confer transcriptional activity in early mouse embryogenesis. EMBO J. 1989, 8, 2551–2557. [Google Scholar] [CrossRef] [Green Version]
- Jerabek, S.; Merino, F.; Scholer, H.R.; Cojocaru, V. OCT4: Dynamic DNA binding pioneers stem cell pluripotency. Biochim. Biophys. Acta 2014, 1839, 138–154. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsui, K.; Tokuzawa, Y.; Itoh, H.; Segawa, K.; Murakami, M.; Takahashi, K.; Maruyama, M.; Maeda, M.; Yamanaka, S. The homeoprotein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell 2003, 113, 631–642. [Google Scholar] [CrossRef] [Green Version]
- Rodda, D.J.; Chew, J.L.; Lim, L.H.; Loh, Y.H.; Wang, B.; Ng, H.H.; Robson, P. Transcriptional regulation of nanog by OCT4 and SOX2. J. Biol. Chem. 2005, 280, 24731–24737. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Rayner, S.; Katoku-Kikyo, N.; Romanova, L.; Kikyo, N. Successful co-immunoprecipitation of Oct4 and Nanog using cross-linking. BioChem. Biophys. Res. Commun. 2007, 361, 611–614. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.A.; Fulton, N.; Cowan, G.; Coutts, S.; Saunders, P.T. Conserved and divergent patterns of expression of DAZL, VASA and OCT4 in the germ cells of the human fetal ovary and testis. BMC Dev. Biol. 2007, 7, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Deng, B.; Geng, L.; Li, L.; Wu, X. Pluripotency factor NANOG promotes germ cell maintenance in vitro without triggering dedifferentiation of spermatogonial stem cells. Theriogenology 2020, 148, 68–75. [Google Scholar] [CrossRef]
- Liu, R.; Li, M.; Li, Z.; Hong, N.; Xu, H.; Hong, Y. Medaka Oct4 is essential for pluripotency in blastula formation and ES cell derivation. Stem Cell Rev. Rep. 2015, 11, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Zhou, Y.; Xu, W.; Jiang, M.; Li, H.; Long, M.; Liu, W.; Liu, J.; Zhao, X.; Xiao, Y. Generation of Stable Induced Pluripotent Stem-like Cells from Adult Zebra Fish Fibroblasts. Int. J. Biol. Sci. 2019, 15, 2340–2349. [Google Scholar] [CrossRef]
- Sanchez-Sanchez, A.V.; Camp, E.; Garcia-Espana, A.; Leal-Tassias, A.; Mullor, J.L. Medaka Oct4 is expressed during early embryo development, and in primordial germ cells and adult gonads. Dev. Dyn. 2010, 239, 672–679. [Google Scholar] [CrossRef]
- Gao, J.; Wang, J.; Jiang, J.; Fan, L.; Wang, W.; Liu, J.; Zhang, Q.; Wang, X. Identification and characterization of a nanog homolog in Japanese flounder (Paralichthys olivaceus). Gene 2013, 531, 411–421. [Google Scholar] [CrossRef]
- Kuijk, E.W.; de Gier, J.; Lopes, S.M.; Chambers, I.; van Pelt, A.M.; Colenbrander, B.; Roelen, B.A. A distinct expression pattern in mammalian testes indicates a conserved role for NANOG in spermatogenesis. PLoS ONE 2010, 5, e10987. [Google Scholar] [CrossRef] [PubMed]
- Zuccotti, M.; Merico, V.; Sacchi, L.; Bellone, M.; Brink, T.C.; Bellazzi, R.; Stefanelli, M.; Redi, C.A.; Garagna, S.; Adjaye, J. Maternal Oct-4 is a potential key regulator of the developmental competence of mouse oocytes. BMC Dev. Biol. 2008, 8, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goel, S.; Fujihara, M.; Minami, N.; Yamada, M.; Imai, H. Expression of NANOG, but not POU5F1, points to the stem cell potential of primitive germ cells in neonatal pig testis. Reproduction 2008, 135, 785–795. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Sanchez, A.V.; Camp, E.; Leal-Tassias, A.; Atkinson, S.P.; Armstrong, L.; Diaz-Llopis, M.; Mullor, J.L. Nanog regulates primordial germ cell migration through Cxcr4b. Stem Cells 2010, 28, 1457–1464. [Google Scholar] [CrossRef]
- Gao, J.; Wang, X.; Zhang, Q. Evolutionary Conservation of pou5f3 Genomic Organization and Its Dynamic Distribution during Embryogenesis and in Adult Gonads in Japanese Flounder Paralichthys olivaceus. Int. J. Mol. Sci. 2017, 18, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Manali, D.; Wang, T.; Bhat, N.; Hong, N.; Li, Z.; Wang, L.; Yan, Y.; Liu, R.; Hong, Y. Identification of pluripotency genes in the fish medaka. Int. J. Biol. Sci. 2011, 7, 440–451. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Han, K.; Chen, S.; Cai, M.; Wang, Y.; Zhang, Z. Molecular cloning and expression of Octamer-binding transcription factor (Oct4) in the large yellow croaker, Larimichthys crocea. Gene Expr. Patterns 2018, 27, 16–30. [Google Scholar] [CrossRef]
- Yu, M.; Xue, T.; Chen, T.; Fang, J.; Pan, Q.; Deng, Y.; Li, L.; Chen, K.; Wang, Y. Maternal inheritance of Nanog ortholog in blunt-snout bream. J. Exp. Zool. B Mol. Dev. Evol. 2017, 328, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Marandel, L.; Labbe, C.; Bobe, J.; Le Bail, P.Y. nanog 5′-upstream sequence, DNA methylation, and expression in gametes and early embryo reveal striking differences between teleosts and mammals. Gene 2012, 492, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; de Mitcheson, Y.S. Gonad development during sexual differentiation in hatchery-produced orange-spotted grouper (Epinephelus coioides) and humpback grouper (Cromileptes altivelis) (Pisces: Serranidae, Epinephelinae). Aquaculture 2009, 287, 191–202. [Google Scholar] [CrossRef]
- Xu, H.; Zhu, X.; Li, W.; Tang, Z.; Zhao, Y.; Wu, X. Isolation and in vitro culture of ovarian stem cells in Chinese soft-shell turtle (Pelodiscus sinensis). J. Cell BioChem. 2018, 119, 7667–7677. [Google Scholar] [CrossRef]
- Wu, X.; Yang, Y.; Zhong, C.; Guo, Y.; Wei, T.; Li, S.; Lin, H.; Liu, X. Integration of ATAC-seq and RNA-seq Unravels Chromatin Accessibility during Sex Reversal in Orange-Spotted Grouper (Epinephelus coioides). Int. J. Mol. Sci. 2020, 21, 2800. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Wang, Q.; Li, G.; He, M.; Tang, H.; Zhang, H.; Yang, X.; Liu, X.; Lin, H. Molecular mechanism of feedback regulation of 17beta-estradiol on two kiss genes in the protogynous orange-spotted grouper (Epinephelus coioides). Mol. Reprod. Dev. 2017, 84, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Theunissen, T.W.; Costa, Y.; Radzisheuskaya, A.; van Oosten, A.L.; Lavial, F.; Pain, B.; Castro, L.F.; Silva, J.C. Reprogramming capacity of Nanog is functionally conserved in vertebrates and resides in a unique homeodomain. Development 2011, 138, 4853–4865. [Google Scholar] [CrossRef] [Green Version]
- Niwa, H.; Sekita, Y.; Tsend-Ayush, E.; Grutzner, F. Platypus Pou5f1 reveals the first steps in the evolution of trophectoderm differentiation and pluripotency in mammals. Evol. Dev. 2008, 10, 671–682. [Google Scholar] [CrossRef]
- Morrison, G.M.; Brickman, J.M. Conserved roles for Oct4 homologues in maintaining multipotency during early vertebrate development. Development 2006, 133, 2011–2022. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, Y.; Ye, D.; Li, J.; Liu, J.; Deng, F. Knockdown of zebrafish Nanog increases primordial germ cells during early embryonic development. Dev. Growth Differ. 2016, 58, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Sharma, R.; George, A.; Singla, S.K.; Palta, P.; Manik, R.; Chauhan, M.S.; Singh, D. Cloning and characterization of buffalo NANOG gene: Alternative transcription start sites, splicing, and polyadenylation in embryonic stem cell-like cells. DNA Cell Biol. 2012, 31, 721–731. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Jena, S.; Levasseur, D.N. Alternative splicing produces Nanog protein variants with different capacities for self-renewal and pluripotency in embryonic stem cells. J. Biol. Chem. 2011, 286, 42690–42703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, H.; Du, H.; Chen, X.H.; Cao, H.; Liu, T.; Li, C.J. Identification of a pou2 ortholog in Chinese sturgeon, Acipenser sinensis and its expression patterns in tissues, immature individuals and during embryogenesis. Fish. Physiol. BioChem. 2012, 38, 929–942. [Google Scholar] [CrossRef]
- Chan, K.K.; Zhang, J.; Chia, N.Y.; Chan, Y.S.; Sim, H.S.; Tan, K.S.; Oh, S.K.; Ng, H.H.; Choo, A.B. KLF4 and PBX1 directly regulate NANOG expression in human embryonic stem cells. Stem Cells 2009, 27, 2114–2125. [Google Scholar] [CrossRef]
- Shanak, S.; Helms, V. DNA methylation and the core pluripotency network. Dev. Biol. 2020, 464, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Santos Nassif Lacerda, S.M.; Costa, G.M.; da Silva Mde, A.; Campos-Junior, P.H.; Segatelli, T.M.; Peixoto, M.T.; Resende, R.R.; de Franca, L.R. Phenotypic characterization and in vitro propagation and transplantation of the Nile tilapia (Oreochromis niloticus) spermatogonial stem cells. Gen. Comp. Endocrinol. 2013, 192, 95–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiaohuan, H.; Yang, Z.; Linyan, L.; Zhenhua, F.; Linyan, Z.; Zhijian, W.; Ling, W.; Deshou, W.; Jing, W. Characterization of the POU5F1 Homologue in Nile Tilapia: From Expression Pattern to Biological Activity. Stem Cells Dev. 2016, 25, 1386–1395. [Google Scholar] [CrossRef]
- Pan, G.; Qin, B.; Liu, N.; Scholer, H.R.; Pei, D. Identification of a nuclear localization signal in OCT4 and generation of a dominant negative mutant by its ablation. J. Biol. Chem. 2004, 279, 37013–37020. [Google Scholar] [CrossRef] [Green Version]
- Noce, T.; Okamoto-Ito, S.; Tsunekawa, N. Vasa homolog genes in mammalian germ cell development. Cell Struct. Funct. 2001, 26, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Raz, E. The function and regulation of vasa-like genes in germ-cell development. Genome Biol. 2000, 1, REVIEWS1017. [Google Scholar] [CrossRef] [Green Version]
- Shinomiya, A.; Tanaka, M.; Kobayashi, T.; Nagahama, Y.; Hamaguchi, S. The vasa-like gene, olvas, identifies the migration path of primordial germ cells during embryonic body formation stage in the medaka, Oryzias latipes. Dev. Growth Differ. 2000, 42, 317–326. [Google Scholar] [CrossRef]
- Toyooka, Y.; Tsunekawa, N.; Takahashi, Y.; Matsui, Y.; Satoh, M.; Noce, T. Expression and intracellular localization of mouse Vasa-homologue protein during germ cell development. Mech Dev. 2000, 93, 139–149. [Google Scholar] [CrossRef]
- Maki, C.B.; Pacchiarotti, J.; Ramos, T.; Pascual, M.; Pham, J.; Kinjo, J.; Anorve, S.; Izadyar, F. Phenotypic and molecular characterization of spermatogonial stem cells in adult primate testes. Hum. Reprod. 2009, 24, 1480–1491. [Google Scholar] [CrossRef]
- Ventela, S.; Makela, J.A.; Kulmala, J.; Westermarck, J.; Toppari, J. Identification and regulation of a stage-specific stem cell niche enriched by Nanog-positive spermatogonial stem cells in the mouse testis. Stem Cells 2012, 30, 1008–1020. [Google Scholar] [CrossRef]
- Khosravi-Farsani, S.; Amidi, F.; Habibi Roudkenar, M.; Sobhani, A. Isolation and enrichment of mouse female germ line stem cells. Cell J. 2015, 16, 406–415. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′ to 3′) | Purpose |
|---|---|---|
| Vasa-F | GAGCCTGAGACCATCATC 1 | ISH |
| Vasa-R | AGGACTCTTCACACTGTTG 1 | ISH |
| Pou5f1-F | TCTACAACAAACCCGCTTACAGT | ISH |
| Pou5f1-R | GCAGAACCAAACACGAACGAC | ISH |
| Nanog-F | ATGGCAGACTGGAAGACGCAGATAA | ISH, RT-PCR |
| Nanog-R | CTACTGATTGACATTGTGTACAATG | ISH, RT-PCR |
| Pou5f1-F1 | TGCGTCCTCGCCATCACTG | RACE |
| Pou5f1-F2 | AACGCATTACCTTGGGTTTCAC | RACE |
| Pou5f1-R1 | GCTCCCTCCAAACTGGTCCTC | RACE |
| Pou5f1-R2 | TGTTGAGTAGTCATCATCGGTGTCC | RACE |
| Nanog-F1 | CTAACAACACTACCTCTC | RACE |
| Nanog-F2 | GTGGACCTGCTGGCTACT | RACE |
| Nanog-R1 | CTTGTCGGTGGTGGTGTA | RACE |
| Nanog-R2 | TATCTGCGTCTTCCAGTCTGCCAT | RACE |
| UPM long | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT 2 | RACE |
| UPM short | CTAATACGACTCACTATAGGGC 2 | RACE |
| NUP | AAGCAGTGGTAACAACGCAGAGT 2 | RACE |
| Pou5f1-F3 | AACGCATTACCTTGGGTTTCACT | RT-PCR |
| Pou5f1-R3 | GGTCCTCCTCTTCCTCTTTCTGG | RT-PCR |
| Nanog-F3 | AACTGAGGAGGCACCAGAAAGAC | RT-PCR |
| Nanog-R3 | CAGCAGGTCCACCAGCAGAG | RT-PCR |
| β-Actin-F | TTCACCACCACAGCCGAGA | RT-PCR |
| β-Actin-R | TGGTCTCGTGGATTCCGCAG | RT-PCR |
| Pou5f1-F4 | TGTCCCAAGCCCAACACCCA | RT-qPCR |
| Pou5f1-R4 | CCAGGCGCTTTCCTTTCTGTC | RT-qPCR |
| Nanog-F4 | GGAGGCACCAGAAAGACACCA | RT-qPCR |
| Nanog-R4 | CTGAGTCAGGGCTTCTCCTTGATA | RT-qPCR |
| β-Actin-F1 | AAATCGCCGCACTGGTTGTT | RT-qPCR |
| β-Actin-R1 | CCCTCTTGCTCTGGGCTTCAT | RT-qPCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, C.; Liu, M.; Tao, Y.; Wu, X.; Yang, Y.; Wang, T.; Meng, Z.; Xu, H.; Liu, X. Pou5f1 and Nanog Are Reliable Germ Cell-Specific Genes in Gonad of a Protogynous Hermaphroditic Fish, Orange-Spotted Grouper (Epinephelus coioides). Genes 2022, 13, 79. https://doi.org/10.3390/genes13010079
Zhong C, Liu M, Tao Y, Wu X, Yang Y, Wang T, Meng Z, Xu H, Liu X. Pou5f1 and Nanog Are Reliable Germ Cell-Specific Genes in Gonad of a Protogynous Hermaphroditic Fish, Orange-Spotted Grouper (Epinephelus coioides). Genes. 2022; 13(1):79. https://doi.org/10.3390/genes13010079
Chicago/Turabian StyleZhong, Chaoyue, Meifeng Liu, Yuhao Tao, Xi Wu, Yang Yang, Tong Wang, Zining Meng, Hongyan Xu, and Xiaochun Liu. 2022. "Pou5f1 and Nanog Are Reliable Germ Cell-Specific Genes in Gonad of a Protogynous Hermaphroditic Fish, Orange-Spotted Grouper (Epinephelus coioides)" Genes 13, no. 1: 79. https://doi.org/10.3390/genes13010079
APA StyleZhong, C., Liu, M., Tao, Y., Wu, X., Yang, Y., Wang, T., Meng, Z., Xu, H., & Liu, X. (2022). Pou5f1 and Nanog Are Reliable Germ Cell-Specific Genes in Gonad of a Protogynous Hermaphroditic Fish, Orange-Spotted Grouper (Epinephelus coioides). Genes, 13(1), 79. https://doi.org/10.3390/genes13010079