
Urinary Extracellular Vesicles and Their miRNA Cargo in Patients with Fabry Nephropathy

 , , , and
, , , and
Abstract
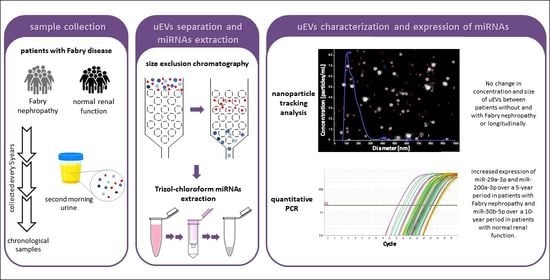
:
1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Small uEVs-Enrichment with Size Exclusion Chromatography (SEC)
2.3. Quantification of uEVs with NTA
2.4. Selection of miRNAs
2.5. Quantification of uEV-Derived miRNAs with qPCR
2.6. Statistical Analysis
3. Results
3.1. Patients Characteristics
3.2. Similar Concentration and Size of uEVs in Patients without and with Fabry Nephropathy
3.3. Positive Correlation between uEV Concentrations and Urinary Creatinine
3.4. No Difference in the Characteristics of uEVs between Chronological Samples
3.5. Similar Expressions of uEV-Derived miRNAs in Patients without and with Fabry Nephropathy
3.6. Changed Expression of miR-29a-3p, miR-200a-3p, and miR-30b-5p in Chronological Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| α-gal A | α-galactosidase A |
| ACEI | angiotensin-converting enzyme inhibitor |
| D-prot | daily proteinuria |
| DKD | diabetic kidney disease |
| DST | disease specific therapy |
| eGFR | estimated glomerular filtration rate |
| EVs | extracellular vesicles |
| FD | Fabry disease |
| GFR | glomerular filtration rate |
| miRNA, miR | microRNA |
| NTA | nanoparticle tracking analysis |
| S-Cr | serum creatinine |
| SEC | size exclusion chromatography |
| UACR | urinary albumin-to-creatinine ratio |
| U-Cr | urinary creatinine |
| uEVs | urinary extracellular vesicles |
| UPCR | urinary protein-to-creatinine ratio |
References
- Brady, R.O.; Gal, A.E.; Bradley, R.M.; Martensson, E.; Warshaw, A.L.; Laster, L. Enzymatic defect in Fabry’s disease: Ceramidetrihexosidase deficiency. N. Engl. J. Med. 1967, 276, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Askari, H.; Kaneski, C.R.; Semino-Mora, C.; Desai, P.; Ang, A.; Kleiner, D.E.; Perlee, L.T.; Quezado, M.; Spollen, L.E.; Wustman, B.A.; et al. Cellular and tissue localization of globotriaosylceramide in Fabry disease. Virchows Arch. 2007, 451, 823–834. [Google Scholar] [CrossRef]
- Germain, D.P. Fabry disease. Orphanet J. Rare Dis. 2010, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Schiffmann, R.; Warnock, D.G.; Banikazemi, M.; Bultas, J.; Linthorst, G.E.; Packman, S.; Sorensen, S.A.; Wilcox, W.R.; Desnick, R.J. Fabry disease: Progression of nephropathy, and prevalence of cardiac and cerebrovascular events before enzyme replacement therapy. Nephrol. Dial. Transplant. 2009, 24, 2102–2111. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.; Oliveira, J.P.; Waldek, S.; Warnock, D.G.; Cianciaruso, B.; Wanner, C. Nephropathy in males and females with Fabry disease: Cross-sectional description of patients before treatment with enzyme replacement therapy. Nephrol. Dial. Transplant. 2008, 23, 1600–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, A.; Cianciaruso, B.; Cizmarik, M.; Germain, D.P.; Mignani, R.; Oliveira, J.P.; Villalobos, J.; Vujkovac, B.; Waldek, S.; Wanner, C.; et al. End-Stage renal disease in patients with Fabry disease: Natural history data from the Fabry registry. Nephrol. Dial. Transplant. 2010, 25, 769–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biegstraaten, M.; Arngrímsson, R.; Barbey, F.; Boks, L.; Cecchi, F.; Deegan, P.B.; Feldt-Rasmussen, U.; Geberhiwot, T.; Germain, D.P.; Hendriksz, C.; et al. Recommendations for initiation and cessation of enzyme replacement therapy in patients with Fabry disease: The European Fabry Working Group consensus document. Orphanet J. Rare Dis. 2015, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Schiffmann, R.; Hughes, D.A.; Linthorst, G.E.; Ortiz, A.; Svarstad, E.; Warnock, D.G.; West, M.L.; Wanner, C. Screening, diagnosis, and management of patients with Fabry disease: Conclusions from a “Kidney Disease: Improving Global Outcomes” (KDIGO) Controversies Conference. Kidney Int. 2017, 91, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Levstek, T.; Vujkovac, B.; Trebusak Podkrajsek, K. Biomarkers of Fabry nephropathy: Review and future perspective. Genes 2020, 11, 1019. [Google Scholar] [CrossRef] [PubMed]
- Verovnik, F.; Benko, D.; Vujkovac, B.; Linthorst, G.E. Remarkable variability in renal disease in a large Slovenian family with Fabry disease. Eur. J. Hum. Genet. 2004, 12, 678–681. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Pisitkun, T.; Shen, R.F.; Knepper, M.A. Identification and proteomic profiling of exosomes in human urine. Proc. Natl. Acad. Sci. USA 2004, 101, 13368–13373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, K.C.; Bond, D.T.; McKee, M.; Skog, J.; Punescu, T.G.; da Silva, N.; Brown, D.; Russo, L.M. Nucleic acids within urinary exosomes/microvesicles are potential biomarkers for renal disease. Kidney Int. 2010, 78, 191–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lennon, R.; Hosawi, S. Glomerular cell crosstalk. Curr. Opin. Nephrol. Hypertens. 2016, 25, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Ståhl, A.; Johansson, K.; Mossberg, M.; Kahn, R.; Karpman, D. Exosomes and microvesicles in normal physiology, pathophysiology, and renal diseases. Pediatr. Nephrol. 2019, 34, 11–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranghino, A.; Dimuccio, V.; Papadimitriou, E.; Bussolati, B. Extracellular vesicles in the urine: Markers and mediators of tissue damage and regeneration. Clin. Kidney J. 2015, 8, 23–30. [Google Scholar] [CrossRef]
- Salih, M.; Zietse, R.; Hoorn, E.J. Urinary extracellular vesicles and the kidney: Biomarkers and beyond. Am. J. Physiol. Renal Physiol. 2014, 306, F1251–F1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seras-Franzoso, J.; Díaz-Riascos, Z.V.; Corchero, J.L.; González, P.; García-Aranda, N.; Mandaña, M.; Riera, R.; Boullosa, A.; Mancilla, S.; Grayston, A.; et al. Extracellular vesicles from recombinant cell factories improve the activity and efficacy of enzymes defective in lysosomal storage disorders. J. Extracell. Vesicles 2021, 10, e12058. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Sun, X.; Scicluna, B.J.; Coleman, B.M.; Hill, A.F. Characterization and deep sequencing analysis of exosomal and non-exosomal miRNA in human urine. Kidney Int. 2014, 86, 433–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekaran, K.; Karolina, D.S.; Sepramaniam, S.; Armugam, A.; Wintour, E.M.; Bertram, J.F.; Jeyaseelan, K. Role of microRNAs in kidney homeostasis and disease. Kidney Int. 2012, 81, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, J.; Wu, Y.; Mai, Y.; Bu, S. Noncoding RNAs in diabetic nephropathy: Pathogenesis, biomarkers, and therapy. J. Diabetes Res. 2020, 2020, 3960857. [Google Scholar] [CrossRef] [PubMed]
- Peters, L.J.F.; Floege, J.; Biessen, E.A.L.; Jankowski, J.; van der Vorst, E.P.C. MicroRNAs in chronic kidney disease: Four candidates for clinical application. Int. J. Mol. Sci. 2020, 21, 6547. [Google Scholar] [CrossRef] [PubMed]
- Levey, A.S.; Stevens, L.A.; Schmid, C.H.; Zhang, Y.; Castro, A.F.; Feldman, H.I.; Kusek, J.W.; Eggers, P.; van Lente, F.; Greene, T.; et al. A new equation to estimate glomerular filtration rate. Ann. Intern. Med. 2009, 150, 604–612. [Google Scholar] [CrossRef]
- Sedej, I.; Tušek Žnidarič, M.; Dolžan, V.; Arnol, M.; Lenassi, M. Extracellular vesicles from urine as biomarkers of kidney alograft injury: Optimization of extracellular vesicle isolation and characterization. Transplantation 2020, 104, 128. [Google Scholar] [CrossRef]
- Lange, T.; Stracke, S.; Rettig, R.; Lendeckel, U.; Kuhn, J.; Schlüter, R.; Rippe, V.; Endlich, K.; Endlich, N. Identification of miR-16 as an endogenous reference gene for the normalization of urinary exosomal miRNA expression data from CKD patients. PLoS ONE 2017, 12, e0183435. [Google Scholar] [CrossRef]
- Beltrami, C.; Simpson, K.; Jesky, M.; Wonnacott, A.; Carrington, C.; Holmans, P.; Newbury, L.; Jenkins, R.; Ashdown, T.; Dayan, C.; et al. Association of elevated urinary miR-126, miR-155 and miR-29b with diabetic kidney disease. Am. J. Pathol. 2018, 188, 1982–1992. [Google Scholar] [CrossRef] [PubMed]
- Zang, J.; Maxwell, A.P.; Simpson, D.A.; McKay, G.J. Differential expression of urinary exosomal microRNAs miR-21-5p and miR-30b-5p in individuals with diabetic kidney disease. Sci. Rep. 2019, 9, 10900. [Google Scholar] [CrossRef] [PubMed]
- Delić, D.; Eisele, C.; Schmid, R.; Baum, P.; Wiech, F.; Gerl, M.; Zimdahl, H.; Pullen, S.S.; Urquhart, R. Urinary exosomal miRNA signature in type II diabetic nephropathy patients. PLoS ONE 2016, 11, e0150154. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ni, W.; Zhang, M.; Liu, S.; Chen, M.; Hong, X.; Ma, Y.; Yu, X.; Wang, W.; Yang, M.; et al. MicroRNA-30/Cx43 axis contributes to podocyte injury by regulating ER stress in diabetic nephropathy. Ann. Transl Med. 2020, 8, 1674. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Zhang, L.; Bai, L.; Chen, S.; Wu, H.; Sun, L.; Wang, X. miR-30b-5p modulate renal epithelial-mesenchymal transition in diabetic nephropathy by directly targeting SNAI1. Biochem. Biophys Res. Commun. 2021, 535, 12–18. [Google Scholar] [CrossRef]
- Eissa, S.; Matboli, M.; Aboushahba, R.; Bekhet, M.M.; Soliman, Y. Urinary Exosomal microRNA panel unravels novel biomarkers for diagnosis of type 2 diabetic kidney disease. J. Diabetes Complicat. 2016, 30, 1585–1592. [Google Scholar] [CrossRef]
- Liu, Y.; Bi, X.; Xiong, J.; Han, W.; Xiao, T.; Xu, X.; Yang, K.; Liu, C.; Jiang, W.; He, T.; et al. MicroRNA-34a promotes renal fibrosis by downregulation of klotho in tubular epithelial cells. Mol. Ther. 2019, 27, 1051–1065. [Google Scholar] [CrossRef]
- Xu, H.; Sun, F.; Li, X.; Sun, L. Down-regulation of miR-23a inhibits high glucose-induced EMT and renal fibrogenesis by up-regulation of SnoN. Hum. Cell 2018, 31, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Patel, V.; Noureddine, L. MicroRNAs and fibrosis. Curr. Opin. Nephrol. Hypertens. 2012, 21, 410–416. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Harkness, A.; Ring, L.; Augustine, D.X.; Oxborough, D.; Robinson, S.; Sharma, V. Normal reference intervals for cardiac dimensions and function for use in echocardiographic practice: A guideline from the British Society of Echocardiography. Echo Res. Pract. 2020, 7, G1–G18. [Google Scholar] [CrossRef] [PubMed]
- Van Balkom, B.W.M.; Pisitkun, T.; Verhaar, M.C.; Knepper, M.A. Exosomes and the kidney: Prospects for diagnosis and therapy of renal diseases. Kidney Int. 2011, 80, 1138–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De, S.; Kuwahara, S.; Hosojima, M.; Ishikawa, T.; Kaseda, R.; Sarkar, P.; Yoshioka, Y.; Kabasawa, H.; Iida, T.; Goto, S.; et al. Exocytosis-mediated urinary full-length megalin excretion is linked with the pathogenesis of diabetic nephropathy. Diabetes 2017, 66, 1391–1404. [Google Scholar] [CrossRef] [Green Version]
- Lytvyn, Y.; Xiao, F.; Kennedy, C.R.J.; Perkins, B.A.; Reich, H.N.; Scholey, J.W.; Cherney, D.Z.; Burger, D. Assessment of urinary microparticles in normotensive patients with type 1 diabetes. Diabetologia 2017, 60, 581–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Siebenthal, M.; Besic, M.; Gheinani, A.H.; Akshay, A.; Platoni, S.L.; Kunz, N.; Burkhard, F.C.; Monastyrskaya, K. Urinary miRNA profiles discriminate between obstruction-induced bladder dysfunction and healthy controls. Sci. Rep. 2021, 11, 10204. [Google Scholar] [CrossRef] [PubMed]
- Turco, A.E.; Lam, W.; Rule, A.D.; Denic, A.; Lieske, J.C.; Miller, V.M.; Larson, J.J.; Kremers, W.K.; Jayachandran, M. Specific renal parenchymal-derived urinary extracellular vesicles identify age-associated structural changes in living donor kidneys. J. Extracell. Vesicles 2016, 5, 29642. [Google Scholar] [CrossRef] [PubMed]
- Najafian, B.; Svarstad, E.; Bostad, L.; Gubler, M.C.; Tøndel, C.; Whitley, C.; Mauer, M. Progressive podocyte injury and globotriaosylceramide (GL-3) accumulation in young patients with Fabry disease. Kidney Int. 2011, 79, 663–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burger, D.; Thibodeau, J.F.; Holterman, C.E.; Burns, K.D.; Touyz, R.M.; Kennedy, C.R.J. Urinary podocyte microparticles identify prealbuminuric diabetic glomerular injury. J. Am. Soc. Nephrol. 2014, 25, 1401–1407. [Google Scholar] [CrossRef] [Green Version]
- Dragovic, R.A.; Gardiner, C.; Brooks, A.S.; Tannetta, D.S.; Ferguson, D.J.P.; Hole, P.; Carr, B.; Redman, C.W.G.; Harris, A.L.; Dobson, P.J.; et al. Sizing and phenotyping of cellular vesicles using nanoparticle tracking analysis. Nanomedicine 2011, 7, 780–788. [Google Scholar] [CrossRef] [Green Version]
- Kamińska, A.; Platt, M.; Kasprzyk, J.; Kuśnierz-Cabala, B.; Gala-Błądzińska, A.; Woźnicka, O.; Jany, B.R.; Krok, F.; Piekoszewski, W.; Kuźniewski, M.; et al. Urinary extracellular vesicles: Potential biomarkers of renal function in diabetic patients. J. Diabetes Res. 2016, 2016, 5741518. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Zieren, R.C.; Horie, K.; Kim, C.J.; Mallick, E.; Jing, Y.; Feng, M.; Kuczler, M.D.; Green, J.; Amend, S.R.; et al. Comprehensive evaluation of methods for small extracellular vesicles separation from human plasma, urine and cell culture medium. J. Extracell. Vesicles 2020, 10, e12044. [Google Scholar] [CrossRef]
- Jaurretche, S.; Venera, G.; Antongiovanni, N.; Perretta, F.; Perez, G.R. Urinary excretion of microRNAs in young Fabry disease patients with mild or absent nephropathy. Open J. Nephrol. 2018, 8, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Jaurretche, S.; Perez, G.R.; Venera, G. High lyso-Gb3 plasma levels associated with decreased miR-29 and miR-200 urinary Excretion in young non-albuminuric male patient with classic Fabry disease. Case Rep. Nephrol. 2019, 2019, 4980942. [Google Scholar] [CrossRef] [Green Version]
- Pezzolesi, M.G.; Satake, E.; McDonnell, K.P.; Major, M.; Smiles, A.M.; Krolewski, A.S. Circulating TGF-Β1-regulated miRNAs and the risk of rapid progression to ESRD in type 1 diabetes. Diabetes 2015, 64, 3285–3293. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Jiang, X.; Duan, W.; Wang, R.; Wang, I.; Zheng, G.; Yan, K.; Wang, L.; Li, J.; Zhang, X.; et al. Cell-free microRNA expression signatures in urine serve as novel noninvasive biomarkers for diagnosis and recurrence prediction of bladder cancer. Oncotarget 2017, 8, 40832–40842. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, K.; Dwivedi, O.P.; Leparc, G.; Rolser, M.; Delic, D.; Forsblom, C.; Groop, P.H.; Groop, L.; Huber, T.B.; Puhka, M.; et al. Comparison of urinary extracellular vesicle isolation methods for transcriptomic biomarker research in diabetic kidney disease. J. Extracell. Vesicles 2020, 10, e12038. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Zhang, Q.; Hu, X.-M.; Mi, T.-Y.; Yu, H.-Y.; Liu, S.-S.; Zhang, B.; Tang, M.; Huang, J.-F.; Xiong, K. How does temperature play a role in the storage of extracellular vesicles? J. Cell. Physiol 2020, 235, 7663–7680. [Google Scholar] [CrossRef]
- Xiao, K.; Lu, D.; Hoepfner, J.; Santer, L.; Gupta, S.; Pfanne, A.; Thum, S.; Lenders, M.; Brand, E.; Nordbeck, P.; et al. Circulating microRNAs in Fabry disease. Sci Rep. 2019, 9, 15277. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| miRNA | Sequence | Accession Number |
|---|---|---|
| hsa-miR-16-5p | 5’UAGCAGCACGUAAAUAUUGGCG | MIMAT0000069 |
| hsa-miR-23a-3p | 5’UGGCUCAGUUCAGCAGGAACAG | MIMAT0000078 |
| hsa-miR-29a-3p | 5’UAGCACCAUCUGAAAUCGGUUA | MIMAT0000086 |
| hsa-miR-30b-5p | 5’UGUAAACAUCCUACACUCAGCU | MIMAT0000420 |
| hsa-miR-34a-5p | 5’UGGCAGUGUCUUAGCUGGUUGU | MIMAT0000255 |
| hsa-miR-191-5p | 5’CAACGGAAUCCCAAAAGCAGCUG | MIMAT0000440 |
| hsa-miR-200a-3p | 5’UAACACUGUCUGGUAACGAUGU | MIMAT0000682 |
| Patient | Genetic Variant | Nephropathy | HCMP | Stroke | Characteristic FD Symptoms | |
|---|---|---|---|---|---|---|
| 1 | p.Arg363Pro | c.1088G>C | Yes | Yes | No | Yes |
| 2 | p.Asn272Ser | c.815A>G | No | Yes | No | Yes |
| 3 | p.Asn272Ser | c.815A>G | No | No | No | Yes |
| 4 | p.Asn272Ser | c.815A>G | Yes | Yes | No | Yes |
| 5 | p.Asn272Ser | c.815A>G | Yes | Yes | No | Yes |
| 6 | p.Asn272Ser | c.815A>G | No | Yes | No | Yes |
| 7 | p.Asn272Ser | c.815A>G | No | No | No | Yes |
| 8 | p.Asn272Ser | c.815A>G | No | No | No | No |
| 9 | p.Asn272Ser | c.815A>G | No | No | No | Yes |
| 10 | p.Glu358del | c.1072_1074delGAG | Yes | Yes | Yes | Yes |
| 11 | p.Glu87Asp | c.261_278del18 | Yes | No | No | Yes |
| 12 | p.Arg342Gln | c.1025G>A | Yes | Yes | No | Yes |
| 13 | p.Ile270Met | c.810T>G | Yes | Yes | Yes | Yes |
| 14 | p.Ile270Met | c.810T>G | Yes | Yes | No | Yes |
| 15 | p.Arg227Ter | c.679C>T | No | No | No | Yes |
| 16 | p.Leu180Phe | c.540G>C | Yes | Yes | Yes | Yes |
| 17 | p.Leu180Phe | c.540G>C | No | No | No | No |
| 18 | p.Gly261ValfsTer8 | c.789delG | Yes | Yes | Yes | Yes |
| 19 | p.Gly261ValfsTer8 | c.789delG | No | No | No | Yes |
| 20 | p.Tyr152His | c.454T>C | Yes | Yes | No | No |
| Parameter | Absolute Frequency (%) | p Value | ||
|---|---|---|---|---|
| No Nephropathy | Nephropathy | |||
| Gender | Male | 4 (20) | 4 (20) | 0.714 |
| Female | 5 (25) | 7 (35) | ||
| DST | No | 4 (20) | 2 (10) | 0.336 |
| Yes | 5 (25) | 9 (45) | ||
| ACEI | No | 8 (40) | 1 (5) | 0.001 |
| Yes | 1 (5) | 10 (50) | ||
| Diabetes | No | 8 (40) | 8 (40) | 0.591 |
| Yes | 1 (5) | 3 (15) | ||
| Parameter | Median (25–75%) | p Value | ||
| No Nephropathy | Nephropathy | |||
| Age (years) | 32.0 (27.3–48.8) | 58.9 (47.9–65.8) | 0.001 | |
| UPCR (g/mol) | 9.60 (8.25–10.9) | 32.2 (12.2–95.2) * | 0.001 | |
| UACR (g/mol) | 0.85 (0.60–1.55) * | 7.80 (2.35–81.5) ** | 0.001 | |
| D-prot (g/day) | 0.11 (0.09–0.16) | 0.20 (0.14–1.41) | 0.067 | |
| S-Cr (µmol/L) | 63.0 (55.5–72.5) | 80.0 (58.0–87.0) | 0.175 | |
| U-Cr (µmol/L) | 5489 (3180–9601) * | 5441 (3247–10,802) | 0.657 | |
| eGFR (mL/min/1.73 m2) | 116.0 (99.5–119.5) | 81.0 (64.0–95.0) | 0.001 | |
| GFR slope (mL/min/year) | −0.71 (−2.30–1.87) | −2.17 (−4.34–−1.28) | 0.080 | |
| Parameter | Median (25–75%) | p Value | |
|---|---|---|---|
| No Nephropathy | Nephropathy | ||
| Concentration (× 1011/mL) | 3.26 (1.23–11.7) | 1.89 (1.27–9.72) | 0.766 |
| Mean Diameter (nm) | 166.4 (140.5–173.0) | 161.8 (140.5–176.5) | 0.882 |
| Modal Diameter (nm) | 121.3 (109.4–146.0) | 113.8 (96.1–129.6) | 0.370 |
| Median Diameter (nm) | 150.8 (126.3–157.5) | 145.5 (129.8–161.4) | 0.941 |
| Parameter | Median (25–75%) | p Value | |
|---|---|---|---|
| 5 Years Ago | Last Follow-up | ||
| Concentration (× 1011/mL) | 4.34 (1.90–7.77) | 1.96 (1.26–9.42) | 0.215 |
| Mean Diameter (nm) | 158.1 (148.1–165.3) | 164.1 (140.5–173.7) | 0.446 |
| Modal Diameter (nm) | 120.9 (113.2–132.2) | 115.3 (105.9–136.3) | 0.777 |
| Median Diameter (nm) | 143.3 (132.5–153.0) | 148.2 (129.4–156.8) | 0.528 |
| Parameter | Median (25–75%) | p Value | |
| 10 Years Ago | Last Follow-up | ||
| Concentration (× 1011/mL) | 7.21 (5.43–17.6) | 2.63 (1.26–9.42) | 0.096 |
| Mean Diameter (nm) | 152.8 (143.8–162.4) | 160.6 (139.1–172.8) | 0.754 |
| Modal Diameter (nm) | 113.7 (107.8–120.3) | 115.3 (105.9–130.8) | 0.900 |
| Median Diameter (nm) | 135.3 (129.4–147.4) | 142.8 (127.8–156.2) | 0.551 |
| Parameter | Median (25–75%) | p Value | |
|---|---|---|---|
| No Nephropathy | Nephropathy | ||
| hsa-miR-200a-3p | 2.73 (1.45–3.82) | 2.35 (1.79–3.34) * | 0.905 |
| hsa-miR-29a-3p | 1.05 (0.91–2.58) | 1.06 (0.85–2.88) | 0.766 |
| hsa-miR-30b-5p | 7.11 (6.42–10.7) | 7.81 (5.39–10.4) | 0.603 |
| hsa-miR-23a-3p | 1.66 (0.76–2.19) | 1.30 (0.96–1.68) | 0.766 |
| hsa-miR-34a-5p | 0.14 (0.10–0.19) | 0.16 (0.06–0.23) | 0.941 |
| (a) All Patients | Parameter | Median (25–75%) | p Value | |
| 5 Years Ago | Last Follow-up | |||
| hsa-miR-200a-3p | 1.96 (1.48–2.25) | 2.73 (1.71–3.69) * | 0.035 | |
| hsa-miR-29a-3p | 0.81 (0.55–1.10) | 1.06 (0.92–2.76) | 0.016 | |
| hsa-miR-30b-5p | 7.62 (4.88–12.1) | 7.47 (6.48–10.4) | 0.711 | |
| hsa-miR-23a-3p | 2.10 (1.45–2.81) | 1.38 (0.92–2.17) | 0.184 | |
| hsa-miR-34a-5p | 0.13 (0.10–0.19) | 0.14 (0.09–0.20) | 0.777 | |
| (b) Without Nephropathy | Parameter | Median (25–75%) | p Value | |
| 5 Years Ago | Last Follow-up | |||
| hsa-miR-200a-3p | 2.02 (1.15–2.71) | 2.77 (1.63–4.09) | 0.263 | |
| hsa-miR-29a-3p | 0.92 (0.82–1.82) | 1.04 (0.90–2.61) | 0.401 | |
| hsa-miR-30b-5p | 9.67 (6.44–14.9) | 7.04 (6.36–10.3) | 0.484 | |
| hsa-miR-23a-3p | 2.43 (1.79–3.38) | 1.76 (0.86–2.20) | 0.484 | |
| hsa-miR-34a-5p | 0.11 (0.08–0.23) | 0.13 (0.10–0.18) | 1.000 | |
| (c) With Nephropathy | Parameter | Median (25–75%) | p Value | |
| 5 Years Ago | Last Follow-up | |||
| hsa-miR-200a-3p | 1.96 (1.48–2.16) | 2.58 (1.71–3.59) * | 0.051 | |
| hsa-miR-29a-3p | 0.60 (0.48–0.89) | 1.11 (0.94–2.94) | 0.017 | |
| hsa-miR-30b-5p | 6.40 (4.50–8.65) | 8.37 (6.26–10.5) | 0.333 | |
| hsa-miR-23a-3p | 1.87 (1.39–2.45) | 1.29 (0.89–1.86) | 0.169 | |
| hsa-miR-34a-5p | 0.14 (0.11–0.19) | 0.15 (0.06–0.25) | 0.721 | |
| (a) All Patients | Parameter | Median (25–75%) | p Value | |
| 10 Years Ago | Last Follow-up | |||
| hsa-miR-200a-3p | 1.49 (1.36–2.02) | 2.81 (1.99–4.13) * | 0.001 | |
| hsa-miR-29a-3p | 0.66 (0.34–0.84) | 1.79 (0.98–2.88) | 0.001 | |
| hsa-miR-30b-5p | 5.19 (3.59–11.2) | 9.29 (6.48–10.6) | 0.048 | |
| hsa-miR-23a-3p | 1.55 (1.05–1.93) | 1.29 (0.79–1.78) | 0.510 | |
| hsa-miR-34a-5p | 0.17 (0.10–0.27) | 0.12 (0.07–0.19) | 0.826 | |
| (b) Without Nephropathy | Parameter | Median (25–75%) | p Value | |
| 10 Years Ago | Last Follow-up | |||
| hsa-miR-200a-3p | 1.54 (1.33–1.95) | 3.05 (2.21–4.76) | 0.028 | |
| hsa-miR-29a-3p | 0.71 (0.25–1.00) | 1.78 (1.00–2.70) | 0.028 | |
| hsa-miR-30b-5p | 7.07 (3.35–10.0) | 8.31 (6.49–11.7) | 0.028 | |
| hsa-miR-23a-3p | 1.58 (0.93–1.70) | 1.34 (0.79–2.17) | 0.917 | |
| hsa-miR-34a-5p | 0.12 (0.08–0.27) | 0.12 (0.09–0.22) | 0.600 | |
| (c) With Nephropathy | Parameter | Median (25–75%) | p Value | |
| 10 Years Ago | Last Follow-up | |||
| hsa-miR-200a-3p | 1.49 (1.35–2.18) | 2.58 (1.86–4.09) * | 0.018 | |
| hsa-miR-29a-3p | 0.60 (0.41–0.88) | 1.89 (0.97–3.08) | 0.012 | |
| hsa-miR-30b-5p | 5.02 (3.88–13.0) | 9.63 (5.68–10.7) | 0.401 | |
| hsa-miR-23a-3p | 1.32 (1.07–2.09) | 1.29 (0.76–1.58) | 0.161 | |
| hsa-miR-34a-5p | 0.21 (0.10–0.35) | 0.12 (0.05–0.22) | 0.575 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levstek, T.; Mlinšek, T.; Holcar, M.; Goričar, K.; Lenassi, M.; Dolžan, V.; Vujkovac, B.; Trebušak Podkrajšek, K. Urinary Extracellular Vesicles and Their miRNA Cargo in Patients with Fabry Nephropathy. Genes 2021, 12, 1057. https://doi.org/10.3390/genes12071057
Levstek T, Mlinšek T, Holcar M, Goričar K, Lenassi M, Dolžan V, Vujkovac B, Trebušak Podkrajšek K. Urinary Extracellular Vesicles and Their miRNA Cargo in Patients with Fabry Nephropathy. Genes. 2021; 12(7):1057. https://doi.org/10.3390/genes12071057
Chicago/Turabian StyleLevstek, Tina, Teo Mlinšek, Marija Holcar, Katja Goričar, Metka Lenassi, Vita Dolžan, Bojan Vujkovac, and Katarina Trebušak Podkrajšek. 2021. "Urinary Extracellular Vesicles and Their miRNA Cargo in Patients with Fabry Nephropathy" Genes 12, no. 7: 1057. https://doi.org/10.3390/genes12071057
APA StyleLevstek, T., Mlinšek, T., Holcar, M., Goričar, K., Lenassi, M., Dolžan, V., Vujkovac, B., & Trebušak Podkrajšek, K. (2021). Urinary Extracellular Vesicles and Their miRNA Cargo in Patients with Fabry Nephropathy. Genes, 12(7), 1057. https://doi.org/10.3390/genes12071057