
In Silico and Transcription Analysis of Trehalose-6-phosphate Phosphatase Gene Family of Wheat: Trehalose Synthesis Genes Contribute to Salinity, Drought Stress and Leaf Senescence

 ,
,  ,
,  ,
,  , ,
, ,
Abstract
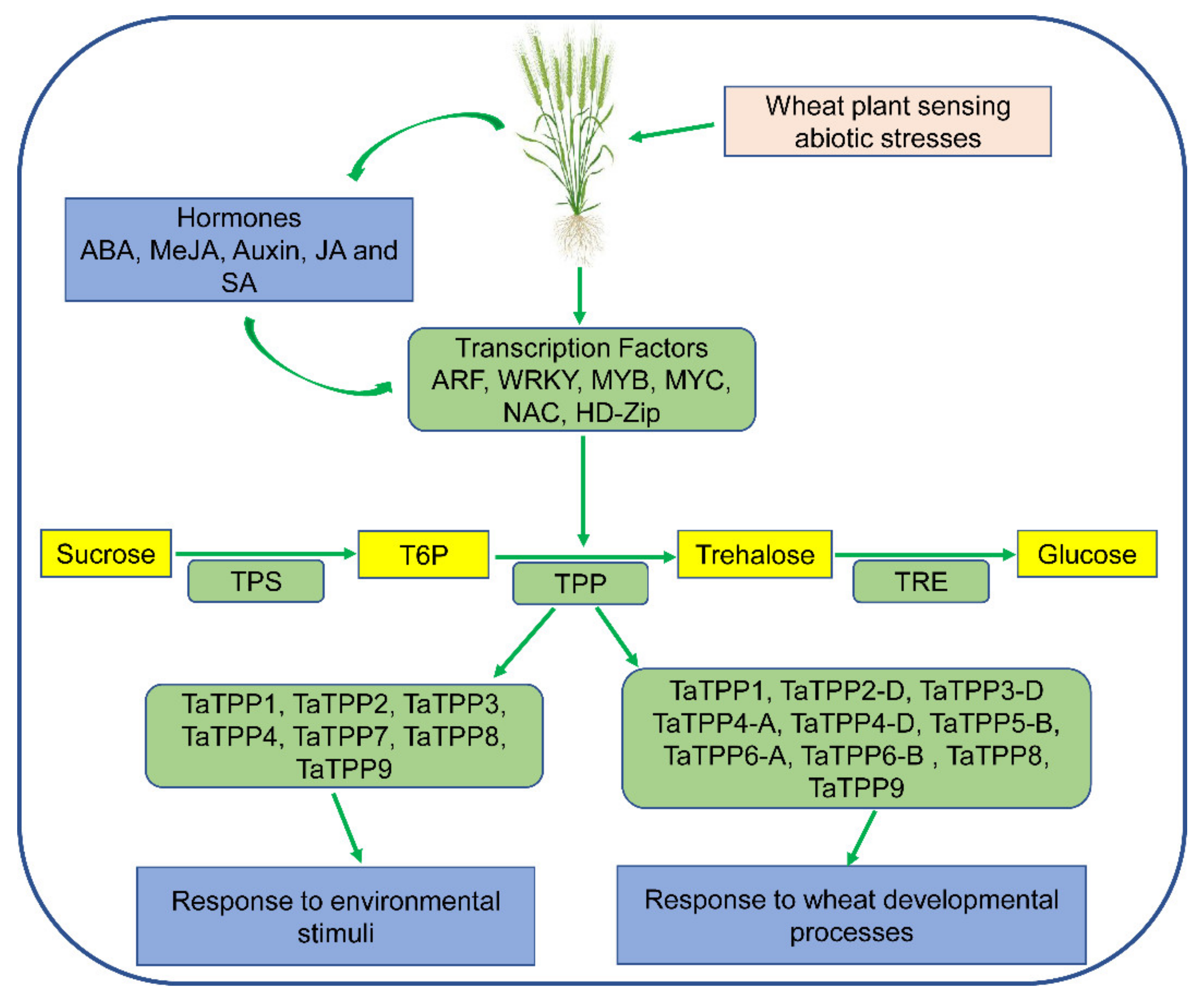
:1. Introduction
2. Materials and Methods
2.1. Identification of Putative TPPs in the Wheat Genome
2.2. Chromosome Localization, Gene Duplication and Synteny Analysis
2.3. Phylogenetic Analysis, Exon-Intron Distribution and 3-D Structure Modeling
2.4. Subcellular Localization Prediction and Protein Domain Analysis
2.5. Analysis of Publicly Accessible Expression Data and Cis-Regulatory Elements (CREs)
2.6. Plant Materials and Treatments
2.7. RNA Extraction, Quantitative Real-Time Reverse Transcription PCR Analysis and Protein Interaction Network
3. Results
3.1. Identification and Annotation of Wheat TPPs
3.2. Subcellular Localization Prediction and Chromosomal Distribution of TaTPPs
3.3. Phylogenetic and Conserved Domain Analyses of TaTPP Proteins
3.4. Gene Structure and Evolution Analyses of TaTPPs
3.5. 3-D Protein Structure Analysis
3.6. Analysis of Cis-Regulatory Elements
3.7. Transcriptional Patterns of TaTPPs in Different Organs and Developmental Stages of Wheat
3.8. Transcriptions of TaTPPs Were Induced in Response to ABA, Abiotic Stresses and Leaf Senescence
3.9. Protein–Protein Interaction Analysis of TaTPPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- USDA. World Agricultural Production. 2019. Available online: https://www.fas.usda.gov/data/world-agricultural-production (accessed on 20 January 2020).
- FAO. World Food Situation. 2021. Available online: http://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 10 July 2021).
- Yin, J.-L.; Fang, Z.-W.; Sun, C.; Zhang, P.; Zhang, X.; Lu, C.; Wang, S.-P.; Ma, D.-F.; Zhu, Y.-X. Rapid identification of a stripe rust resistant gene in a space-induced wheat mutant using specific locus amplified fragment (SLAF) sequencing. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Tester, M.; Bacic, A. Abiotic Stress Tolerance in Grasses. From Model Plants to Crop Plants. Plant Physiol. 2005, 137, 791–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafi, M.; Zhang, G.; Bakht, J.; Khan, M.A.; Islam, U.; Khan, M.D.; Raziuddin, G. Effect of cadmium and salinity stresses on root morphology of wheat. Pak. J. Bot. 2010, 42, 2747–2754. [Google Scholar]
- Foolad, M.R. Recent Advances in Genetics of Salt Tolerance in Tomato. Plant Cell Tissue Organ Cult. 2004, 76, 101–119. [Google Scholar] [CrossRef]
- Genc, Y.; Taylor, J.; Lyons, G.; Li, Y.; Cheong, J.; Appelbee, M.; Oldach, K.; Sutton, T. Bread Wheat with High Salinity and Sodicity Tolerance. Front. Plant Sci. 2019, 10, 1280. [Google Scholar] [CrossRef] [Green Version]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.A. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Interact. 2018, 13, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Wingler, A. The function of trehalose biosynthesis in plants. Phytochemistry 2002, 60, 437–440. [Google Scholar] [CrossRef]
- Mostafa, M.R.; Mervat, S.S.; Safaa, R.E.-L.; Ebtihal, M.A.E.; Magdi, T.A. Exogenous α-tocopherol has a beneficial effect on Glycine max(L.) plants irrigated with diluted sea water. J. Hortic. Sci. Biotechnol. 2015, 90, 195–202. [Google Scholar] [CrossRef]
- Patist, A.; Zoerb, H. Preservation mechanisms of trehalose in food and biosystems. Colloids Surf. B Biointerfaces 2005, 40, 107–113. [Google Scholar] [CrossRef]
- Iturriaga, G.; Suárez, R.; Nova-Franco, B. Trehalose Metabolism: From Osmoprotection to Signaling. Int. J. Mol. Sci. 2009, 10, 3793–3810. [Google Scholar] [CrossRef] [Green Version]
- Lunn, J.E.; Delorge, I.; Figueroa, C.M.; Van Dijck, P.; Stitt, M. Trehalose metabolism in plants. Plant J. 2014, 79, 544–567. [Google Scholar] [CrossRef]
- Klähn, S.; Hagemann, M. Compatible solute biosynthesis in cyanobacteria. Environ. Microbiol. 2010, 13, 551–562. [Google Scholar] [CrossRef]
- Chang, S.-W.; Chang, W.-H.; Lee, M.-R.; Yang, T.-J.; Yu, N.-Y.; Chen, C.-S.; Shaw, J.-F. Simultaneous Production of Trehalose, Bioethanol, and High-Protein Product from Rice by an Enzymatic Process. J. Agric. Food Chem. 2010, 58, 2908–2914. [Google Scholar] [CrossRef]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef]
- Lunn, J.E. Gene families and evolution of trehalose metabolism in plants. Funct. Plant Biol. 2007, 34, 550–563. [Google Scholar] [CrossRef]
- Avonce, N.; Mendoza-Vargas, A.; Morett, E.; Iturriaga, G. Insights on the evolution of trehalose biosynthesis. BMC Evol. Biol. 2006, 6, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponnu, J.; Wahl, V.; Schmid, M. Trehalose-6-Phosphate: Connecting Plant Metabolism and Development. Front. Plant Sci. 2011, 2, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunn, J.E.; Feil, R.; Hendriks, J.H.M.; Gibon, Y.; Morcuende, R.; Osuna, D.; Scheible, W.-R.; Carillo, P.; Hajirezaei, M.-R.; Stitt, M. Sugar-induced increases in trehalose 6-phosphate are correlated with redox activation of ADPglucose pyrophosphorylase and higher rates of starch synthesis in Arabidopsis thaliana. Biochem. J. 2006, 397, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meitzel, T.; Radchuk, R.; McAdam, E.L.; Thormählen, I.; Feil, R.; Munz, E.; Hilo, A.; Geigenberger, P.; Ross, J.J.; Lunn, J.E.; et al. Trehalose-6-phosphate promotes seed filling by activating auxin biosynthesis. New Phytol. 2021, 229, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Yadav, U.P.; Ivakov, A.; Feil, R.; Duan, G.Y.; Walther, D.; Giavalisco, P.; Piques, M.; Carillo, P.; Hubberten, H.-M.; Stitt, M.; et al. The sucrose–trehalose 6-phosphate (Tre6P) nexus: Specificity and mechanisms of sucrose signalling by Tre6P. J. Exp. Bot. 2014, 65, 1051–1068. [Google Scholar] [CrossRef] [Green Version]
- Acevedo Hernández, G.J.; León, P.; Herrera-Estrella, L.R. Sugar and ABA responsiveness of a minimal RBCS light-responsive unit is mediated by direct binding of ABI4. Plant J. 2005, 43, 506–519. [Google Scholar] [CrossRef]
- Gómez, L.D.; Gilday, A.; Feil, R.; Lunn, J.E.; Graham, I.A. AtTPS1-mediated trehalose 6-phosphate synthesis is essential for embryogenic and vegetative growth and responsiveness to ABA in germinating seeds and stomatal guard cells. Plant J. 2010, 64, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fichtner, F.; Olas, J.J.; Feil, R.; Watanabe, M.; Krause, U.; Hoefgen, R.; Stitt, M.; Lunn, J.E. Functional Features of TREHALOSE-6-PHOSPHATE SYNTHASE1, an Essential Enzyme in Arabidopsis [OPEN]. Plant Cell 2020, 32, 1949–1972. [Google Scholar] [CrossRef] [Green Version]
- Fichtner, F.; Barbier, F.F.; Annunziata, M.G.; Feil, R.; Olas, J.J.; Mueller-Roeber, B.; Stitt, M.; Beveridge, C.A.; Lunn, J.E. Regulation of shoot branching in Arabidopsis by trehalose-6-phosphate. New Phytol. 2021, 229, 2135–2151. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Ye, N.; Huang, M.; Feng, L.; Li, H.; Zhang, J. OsTPP1 regulates seed germination through the crosstalk with abscisic acid in rice. New Phytol. 2021, 230, 1925–1939. [Google Scholar] [CrossRef]
- Wang, S.; Ouyang, K.; Wang, K. Genome-Wide Identification, Evolution, and Expression Analysis of TPS and TPP Gene Families in Brachypodium distachyon. Plants 2019, 8, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh-Nagasawa, N.; Nagasawa, N.; Malcomber, S.; Sakai, H.; Jackson, D. A trehalose metabolic enzyme controls inflorescence architecture in maize. Nature 2006, 441, 227–230. [Google Scholar] [CrossRef]
- Pellny, T.K.; Ghannoum, O.; Conroy, J.P.; Schluepmann, H.; Smeekens, S.; Andralojc, J.; Krause, K.P.; Goddijn, O.; Paul, M. Genetic modification of photosynthesis with E. coli genes for trehalose synthesis. Plant Biotechnol. J. 2004, 2, 71–82. [Google Scholar] [CrossRef]
- Miranda, J.A.; Avonce, N.; Suárez, R.; Thevelein, J.M.; Van Dijck, P.; Iturriaga, G. A bifunctional TPS–TPP enzyme from yeast confers tolerance to multiple and extreme abiotic-stress conditions in transgenic Arabidopsis. Planta 2007, 226, 1411–1421. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Yang, J.; Wang, Q.; Zhu, H.; Chen, Z.; Dao, Y.; Wang, K. Overexpression of the trehalose-6-phosphate phosphatase family gene AtTPPF improves the drought tolerance of Arabidopsis thaliana. BMC Plant Biol. 2019, 19, 381. [Google Scholar] [CrossRef] [Green Version]
- Pilon-Smits, E.A.; Terry, N.; Sears, T.; Kim, H.; Zayed, A.; Hwang, S.; van Dun, K.; Voogd, E.; Verwoerd, T.C.; Krutwagen, R.W.; et al. Trehalose-producing transgenic tobacco plants show improved growth performance under drought stress. J. Plant Physiol. 1998, 152, 525–532. [Google Scholar] [CrossRef]
- Wu, B.; Su, X. Identification of drought response genes in Zygophyllum xanthoxylum by suppression subtractive hybridization. J. Plant Biol. 2016, 59, 377–385. [Google Scholar] [CrossRef]
- Garg, A.K.; Kim, J.-K.; Owens, T.G.; Ranwala, A.P.; Choi, Y.D.; Kochian, L.; Wu, R.J. Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc. Natl. Acad. Sci. USA 2002, 99, 15898–15903. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Sun, H.; Wang, X.; Jin, W.; Chen, Q.; Yuan, Z.; Yu, H. Physiological and transcriptomic analyses reveal the molecular networks of responses induced by exogenous trehalose in plant. PLoS ONE 2019, 14, e0217204. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Chen, W.; Gao, J.; Yang, F.; Zhuang, C. Overexpression of the trehalose-6-phosphate phosphatase OsTPP3 increases drought tolerance in rice. Plant Biotechnol. Rep. 2019, 13, 285–292. [Google Scholar] [CrossRef]
- Ge, L.; Chao, D.-Y.; Shi, M.; Zhu, M.-Z.; Gao, J.-P.; Lin, H.-X. Overexpression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results in the activation of stress responsive genes. Planta 2008, 228, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Pérez, P.; Camacho-Zamora, B.D.; Espinoza-Sánchez, E.A.; Gutiérrez-Soto, G.; Zavala-García, F.; Abraham-Juárez, M.J.; Sinagawa-García, S.R. Characterization of Trehalose-6-phosphate Synthase and Trehalose-6-phosphate Phosphatase Genes and Analysis of its Differential Expression in Maize (Zea mays) Seedlings under Drought Stress. Plants 2020, 9, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Q.; Wang, S.; Dao, Y.; Wang, J.; Wang, K.; Wang, S. Arabidopsis thaliana trehalose-6-phosphate phosphatase gene TPPI enhances drought tolerance by regulating stomatal apertures. J. Exp. Bot. 2020, 71, 4285–4297. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Chen, Q.; Xu, S.; Liu, W.C.; Zhu, X.; Song, C.P. Trehalose-6-phosphate phosphatase E modulates ABA-controlled root growth and stomatal movement in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 1518–1534. [Google Scholar] [CrossRef] [Green Version]
- Wingler, A.; Delatte, T.L.; O’Hara, L.; Primavesi, L.; Jhurreea, D.; Paul, M.; Schluepmann, H. Trehalose 6-Phosphate Is Required for the Onset of Leaf Senescence Associated with High Carbon Availability. Plant Physiol. 2012, 158, 1241–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noodén, L.D.; Guiamét, J.J.; John, I. Senescence mechanisms. Physiol. Plant. 1997, 101, 746–753. [Google Scholar] [CrossRef]
- Pourtau, N.; Jennings, R.; Pelzer, E.; Pallas, J.; Wingler, A. Effect of sugar-induced senescence on gene expression and implications for the regulation of senescence in Arabidopsis. Planta 2006, 224, 556–568. [Google Scholar] [CrossRef] [PubMed]
- Wingler, A.; Purdy, S.; MacLean, J.A.; Pourtau, N. The role of sugars in integrating environmental signals during the regulation of leaf senescence. J. Exp. Bot. 2006, 57, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; He, Z.; Tian, X.; Gao, F.; Xu, D.; Liu, J.; Wen, W.; Fu, L.; Li, G.; Sui, X.; et al. Cloning of TaTPP-6AL1 associated with grain weight in bread wheat and development of functional marker. Mol. Breed. 2017, 37, 78. [Google Scholar] [CrossRef]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Combet, C.; Blanchet, C.; Geourjon, C.; Deléage, G. NPS@: Network Protein Sequence Analysis. Trends Biochem. Sci. 2000, 25, 147–150. [Google Scholar] [CrossRef]
- Izidoro, S.; De Melo-Minardi, R.C.; Pappa, G.L. GASS: Identifying enzyme active sites with genetic algorithms. Bioinformatics 2015, 31, 864–870. [Google Scholar] [CrossRef] [Green Version]
- Lovell, S.C.; Davis, I.W.; Arendall III, W.B.; De Bakker, P.I.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure validation by Cα geometry: ϕ, ψ and Cβ deviation. Proteins 2003, 50, 437–450. [Google Scholar] [CrossRef]
- Moraes, J.P.A.; Pappa, G.L.; Pires, D.E.V.; Izidoro, S.C. GASS-WEB: A web server for identifying enzyme active sites based on genetic algorithms. Nucleic Acids Res. 2017, 45, W315–W319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2017, 27, 293–315. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Wen, L.; Gao, X.; Jin, C.; Xue, Y.; Yao, X. DOG 1.0: Illustrator of protein domain structures. Cell Res. 2009, 19, 271–273. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, w202–w208. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- NetBeans, IDE 8.0. Oracle Co., Redwood City, CA, 2015. Available online: https://netbeans.org (accessed on 25 March 2021).
- Hruz, T.; Laule, O.; Szabo, G.; Wessendorp, F.; Bleuler, S.; Oertle, L.; Widmayer, P.; Gruissem, W.; Zimmermann, P. Genevestigator V3: A Reference Expression Database for the Meta-Analysis of Transcriptomes. Adv. Bioinform. 2008, 2008, 1–5. [Google Scholar] [CrossRef]
- Howe, E.; Holton, K.; Nair, S.; Schlauch, D.; Sinha, R.; Quackenbush, J. MeV: Multi Experiment Viewer. In Biomedical Informatics for Cancer Research; Springer: Berlin/Heidelberg, Germany, 2010; pp. 267–277. [Google Scholar]
- Duan, Y.-H.; Guo, J.; Ding, K.; Wang, S.-J.; Zhang, H.; Dai, X.-W.; Chen, Y.-Y.; Govers, F.; Huang, L.-L.; Kang, Z.-S. Characterization of a wheat HSP70 gene and its expression in response to stripe rust infection and abiotic stresses. Mol. Biol. Rep. 2010, 38, 301–307. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Paolacci, A.R.; Tanzarella, O.A.; Porceddu, E.; Ciaffi, M. Identification and validation of reference genes for quantitative RT-PCR normalization in wheat. BMC Mol. Biol. 2009, 10, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleury, D.; Jefferies, S.; Kuchel, H.; Langridge, P. Genetic and genomic tools to improve drought tolerance in wheat. J. Exp. Bot. 2010, 61, 3211–3222. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, M.; Hota, A.; Kar, A.; Chini, D.S.; Malick, R.C.; Patra, B.C.; Das, B.K. In silico structural and functional modelling of Antifreeze protein (AFP) sequences of Ocean pout (Zoarces americanus, Bloch & Schneider 1801). J. Genet. Eng. Biotechnol. 2018, 16, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Yaqoob, U.; Kaul, T.; Nawchoo, I.A. In-Silico Analysis, Structural Modelling and Phylogenetic Analysis of Acetohydroxyacid Synthase Gene of Oryza sativa. Med. Aromat. Plants 2016, 5, 2167-0412. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Lv, W.; Jiang, S.; Zhang, D.; Cai, G.; Pan, J.; Li, D. Genome-wide identification and expression analysis of calcium-dependent protein kinase in maize. BMC Genom. 2013, 14, 433. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Cavalcanti, A.; Chen, F.-C.; Bouman, P.; Li, W.-H. Extent of Gene Duplication in the Genomes of Drosophila, Nematode, and Yeast. Mol. Biol. Evol. 2002, 19, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Bordoli, L.; Schwede, T. Automated Protein Structure Modeling with SWISS-MODEL Workspace and the Protein Model Portal. In Homology Modeling; Springer: Berlin/Heidelberg, Germany, 2011; Volume 857, pp. 107–136. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Rahman, M.M.; Eom, J.S.; Jeon, J.S. Genome-wide identification, expression profiling and promoter analysis of trehalose-6-phosphate phosphatase gene family in rice. J. Plant Biol. 2021, 64, 55–71. [Google Scholar] [CrossRef]
- Grace, M.L.; Chandrasekharan, M.; Hall, T.C.; Crowe, A.J. Sequence and Spacing of TATA Box Elements Are Critical for Accurate Initiation from the β-Phaseolin Promoter. J. Biol. Chem. 2004, 279, 8102–8110. [Google Scholar] [CrossRef] [Green Version]
- Vogel, G.; Aeschbacher, R.A.; Müller, J.; Boller, T.; Wiemken, A. Trehalose-6-phosphate phosphatases from Arabidopsis thaliana: Identification by functional complementation of the yeast tps2 mutant. Plant J. 1998, 13, 673–683. [Google Scholar] [CrossRef]
- Svanström, Å.; van Leeuwen, M.R.; Dijksterhuis, J.; Melin, P. Trehalose synthesis in Aspergillus niger: Characterization of six homologous genes, all with conserved orthologs in related species. BMC Microbiol. 2014, 14, 90. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-J.; Hao, Y.-J.; Zhang, Z.-G.; Chen, T.; Zhang, J.-S.; Chen, S.-Y. Isolation of trehalose-6-phosphate phosphatase gene from tobacco and its functional analysis in yeast cells. J. Plant Physiol. 2005, 162, 215–223. [Google Scholar] [CrossRef]
- Streeter, J.; Gomez, M. Three enzymes for trehalose synthesis in Bradyrhizobium cultured bacteria and in bacteroids from soybean nodules. Appl. Environ. Microbiol. 2006, 72, 4250–4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, C.; Bledsoe, S.W.; Siekman, A.; Kollman, A.; Waters, B.; Feil, R.; Stitt, M.; Lagrimini, L.M. The trehalose pathway in maize: Conservation and gene regulation in response to the diurnal cycle and extended darkness. J. Exp. Bot. 2014, 65, 5959–5973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schluepmann, H.; van Dijken, A.; Aghdasi, M.; Wobbes, B.; Paul, M.; Smeekens, S. Trehalose Mediated Growth Inhibition of Arabidopsis Seedlings Is Due to Trehalose-6-Phosphate Accumulation. Plant Physiol. 2004, 135, 879–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyman, B.; Van Dijck, P.; Thevelein, J. An unexpected plethora of trehalose biosynthesis genes in Arabidopsis thaliana. Trends Plant Sci. 2001, 6, 510–513. [Google Scholar] [CrossRef]
- Lawton-Rauh, A. Evolutionary dynamics of duplicated genes in plants. Mol. Phylogenetics Evol. 2003, 29, 396–409. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [Green Version]
- Holub, E.B. The arms race is ancient history in Arabidopsis, the wildflower. Nat. Rev. Genet. 2001, 2, 516–527. [Google Scholar] [CrossRef]
- Miyakawa, T.; Miyazono, K.-I.; Sawano, Y.; Hatano, K.-I.; Tanokura, M. Crystal structure of ginkbilobin-2 with homology to the extracellular domain of plant cysteine-rich receptor-like kinases. Proteins Struct. Funct. Bioinform. 2009, 77, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tian, L.-H.; Zhao, J.-F.; Song, Y.; Zhang, C.-J.; Guo, Y. Identification of an Apoplastic Protein Involved in the Initial Phase of Salt Stress Response in Rice Root by Two-Dimensional Electrophoresis. Plant Physiol. 2009, 149, 916–928. [Google Scholar] [CrossRef] [Green Version]
- Krasensky-Wrzaczek, J.; Broyart, C.; Rabanal, F.; Jonak, C. The Redox-Sensitive Chloroplast Trehalose-6-Phosphate Phosphatase AtTPPD Regulates Salt Stress Tolerance. Antioxid. Redox Signal. 2014, 21, 1289–1304. [Google Scholar] [CrossRef] [Green Version]
- Cowan, J.A. Metal Activation of Enzymes in Nucleic Acid Biochemistry. Chem. Rev. 1998, 98, 1067–1088. [Google Scholar] [CrossRef] [PubMed]
- Bertini, G.; Gray, H.B.; Gray, H.; Valentine, J.S.; Stiefel, E.I.; Stiefel, E. Biological Inorganic Chemistry: Structure and Reactivity; University Science Books: Sausalito, CA, USA, 2007. [Google Scholar]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.-I.; Hong, J.-H.; Ha, J.-O.; Kang, J.-Y.; Kim, S.Y. ABFs, a Family of ABA-responsive Element Binding Factors. J. Biol. Chem. 2000, 275, 1723–1730. [Google Scholar] [CrossRef] [Green Version]
- Hobo, T.; Asada, M.; Kowyama, Y.; Hattori, T. ACGT-containing abscisic acid response element (ABRE) and coupling element 3 (CE3) are functionally equivalent. Plant J. 1999, 19, 679–689. [Google Scholar] [CrossRef]
- Guiltinan, M.J.; Marcotte, W.R.; Quatrano, R.S. A Plant Leucine Zipper Protein That Recognizes an Abscisic Acid Response Element. Science 1990, 250, 267–271. [Google Scholar] [CrossRef] [Green Version]
- Marcotte, W.R., Jr.; Russell, S.H.; Quatrano, R.S. Abscisic acid-responsive sequences from the em gene of wheat. Plant Cell 1989, 1, 969–976. [Google Scholar]
- Narusaka, Y.; Nakashima, K.; Shinwari, Z.K.; Sakuma, Y.; Furihata, T.; Abe, H.; Narusaka, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Interaction between two cis-acting elements, ABRE and DRE, in ABA-dependent expression of Arabidopsis rd29A gene in response to dehydration and high-salinity stresses. Plant J. 2003, 34, 137–148. [Google Scholar] [CrossRef]
- Shen, Q.; Zhang, P.; Ho, T. Modular nature of abscisic acid (ABA) response complexes: Composite promoter units that are necessary and sufficient for ABA induction of gene expression in barley. Plant Cell 1996, 8, 1107–1119. [Google Scholar] [PubMed] [Green Version]
- Banerjee, J.; Sahoo, D.K.; Dey, N.; Houtz, R.L.; Maiti, I.B. An Intergenic Region Shared by At4g35985 and At4g35987 in Arabidopsis thaliana Is a Tissue Specific and Stress Inducible Bidirectional Promoter Analyzed in Transgenic Arabidopsis and Tobacco Plants. PLoS ONE 2013, 8, e79622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretzschmar, T.; Pelayo, M.A.F.; Trijatmiko, K.; Gabunada, L.F.M.; Alam, R.; Jimenez, R.; Mendioro, M.S.; Slamet-Loedin, I.; Sreenivasulu, N.; Bailey-Serres, J.; et al. A trehalose-6-phosphate phosphatase enhances anaerobic germination tolerance in rice. Nat. Plants 2015, 1, 15124. [Google Scholar] [CrossRef]
- Shima, S.; Matsui, H.; Tahara, S.; Imai, R. Biochemical characterization of rice trehalose-6-phosphate phosphatases supports distinctive functions of these plant enzymes. FEBS J. 2007, 274, 1192–1201. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Splice Variant | PC | ORF | Chromosome Location | Introns | Exons | Length (aa) | M.W. (KDa) | PI | Instability Index | Aliphatic Index | GRAVY | SL Prediction | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chr | Strand | Start | End | Chr Length | ||||||||||||||
| TaTPP1-A | TraesCS1A02G210400.1 | 1 | VII | 1146 | 1A | reverse | 372639121 | 372643307 | 594102056 | 9 | 10 | 381 | 42.56 | 5.61 | 46.35 | 83.91 | −0.314 | chloroplast |
| TaTPP1-B | TraesCS1B02G224300.2 | 2 | VII | 1317 | 1B | reverse | 402147526 | 402150391 | 689851870 | 12 | 13 | 438 | 49.46 | 6.09 | 47.92 | 86.32 | −0.238 | mitochondrion |
| TaTPP1-D | TraesCS1D02G213700.1 | 1 | VII | 1146 | 1D | reverse | 298692275 | 298696295 | 495453186 | 9 | 10 | 380 | 42.54 | 5.53 | 45.29 | 84.13 | −0.315 | chloroplast |
| TaTPP2-A | TraesCS2A02G161000.1 | 1 | II | 1074 | 2A | forward | 111921839 | 111925164 | 780798557 | 8 | 9 | 357 | 39.43 | 6.11 | 34.73 | 79.3 | −0.254 | chloroplast |
| TaTPP2-B | TraesCS2B02G187000.1 | 1 | II | 1077 | 2B | forward | 161722263 | 161725704 | 801256715 | 8 | 9 | 358 | 39.62 | 6.04 | 35.64 | 82.6 | −0.216 | chloroplast |
| TaTPP2-D | TraesCS2D02G168100.1 | 1 | II | 1077 | 2D | forward | 111588533 | 111591881 | 651852609 | 8 | 9 | 358 | 39.66 | 5.77 | 35.52 | 80.42 | −0.249 | chloroplast |
| TaTPP3-A | TraesCS2A02G161100.1 | 1 | II | 1077 | 2A | forward | 112744717 | 112747766 | 780798557 | 8 | 9 | 358 | 39.63 | 7.14 | 35.67 | 79.61 | −0.282 | chloroplast |
| TaTPP3-B | TraesCS2B02G187100.1 | 1 | II | 1077 | 2B | forward | 162007594 | 162010969 | 801256715 | 8 | 9 | 358 | 39.61 | 6.57 | 33.2 | 79.61 | −0.264 | chloroplast |
| TaTPP3-D | TraesCS2D02G168200.1 | 1 | II | 1077 | 2D | forward | 112099169 | 112102442 | 651852609 | 8 | 9 | 358 | 39.50 | 6.84 | 34.77 | 79.08 | −0.283 | chloroplast |
| TaTPP4-A | TraesCS2A02G161200.1 | 1 | II | 1077 | 2A | reverse | 113309228 | 113312068 | 780798557 | 8 | 9 | 358 | 39.63 | 6.77 | 35.05 | 79.86 | −0.281 | chloroplast |
| TaTPP4-B | TraesCS2B02G187200.1 | 1 | II | 1077 | 2B | reverse | 162445597 | 162448929 | 801256715 | 8 | 9 | 358 | 39.65 | 7.12 | 32.59 | 79.86 | −0.257 | chloroplast |
| TaTPP4-D | TraesCS2D02G168300.1 | 2 | II | 1179 | 2D | reverse | 112177996 | 112181078 | 651852609 | 7 | 8 | 392 | 43.58 | 6.44 | 39.59 | 82.65 | −0.190 | chloroplast |
| TaTPP5-A | TraesCS2A02G167100.1 | 1 | V | 750 | 2A | reverse | 119307539 | 119314162 | 780798557 | 8 | 9 | 249 | 28.67 | 8.28 | 43.34 | 75.9 | −0.700 | other |
| TaTPP5-B | TraesCS2B02G193300.1 | 1 | V | 1680 | 2B | reverse | 168831609 | 168853380 | 801256715 | 10 | 11 | 559 | 62.28 | 8.61 | 45.14 | 85.64 | −0.230 | secreted |
| TaTPP6-A | TraesCS2A02G412100.1 | 1 | VI | 1113 | 2A | forward | 669749666 | 669753186 | 780798557 | 9 | 10 | 370 | 41.23 | 5.70 | 49.54 | 83.46 | −0.307 | chloroplast |
| TaTPP6-B | TraesCS2B02G430700.1 | 1 | VI | 1116 | 2B | forward | 619679522 | 619682850 | 801256715 | 9 | 10 | 371 | 41.19 | 5.88 | 51.27 | 82.21 | −0.297 | chloroplast |
| TaTPP6-D | TraesCS2D02G409300.1 | 1 | VI | 1113 | 2D | forward | 524105415 | 524108680 | 651852609 | 9 | 10 | 370 | 41.11 | 5.58 | 55.74 | 84.27 | −0.232 | chloroplast |
| TaTPP7-A | TraesCS3A02G085700.1 | 1 | V | 1662 | 3A | forward | 55223622 | 55255982 | 750843639 | 11 | 12 | 553 | 61.55 | 8.06 | 36.82 | 83.67 | −0.291 | secreted |
| TaTPP7-D | TraesCS3D02G085800.1 | 1 | V | 1755 | 3D | forward | 43259299 | 43283531 | 615552423 | 13 | 14 | 584 | 65.02 | 8.86 | 32.8 | 77.4 | −0.400 | mitochondrion |
| TaTPP8-A | TraesCS5A02G190000.1 | 1 | I | 1122 | 5A | reverse | 394181080 | 394183400 | 709773743 | 5 | 6 | 373 | 40.85 | 8.95 | 41.99 | 79.81 | −0.144 | chloroplast |
| TaTPP8-B | TraesCS5B02G193100.1 | 3 | I | 1248 | 5B | reverse | 348448002 | 348450302 | 713149757 | 4 | 5 | 373 | 40.93 | 8.97 | 41.35 | 79.81 | −0.145 | chloroplast |
| TaTPP8-D | TraesCS5D02G200800.1 | 1 | I | 1122 | 5D | reverse | 303758772 | 303761166 | 566080677 | 5 | 6 | 373 | 40.86 | 8.96 | 42.04 | 79.81 | −0.142 | chloroplast |
| TaTPP9-A | TraesCS6A02G248400.1 | 1 | VI | 1119 | 6A | forward | 461143866 | 461147635 | 618079260 | 8 | 9 | 372 | 41.11 | 5.68 | 48.98 | 85.65 | −0.218 | chloroplast |
| TaTPP9-B | TraesCS6B02G276300.1 | 1 | VI | 1119 | 6B | reverse | 500209451 | 500213722 | 720988478 | 8 | 9 | 372 | 41.28 | 5.89 | 49.51 | 85.89 | −0.232 | chloroplast |
| TaTPP9-D | TraesCS6D02G230500.1 | 1 | VI | 1119 | 6D | forward | 323712099 | 323716021 | 473592718 | 8 | 9 | 372 | 41.05 | 5.56 | 49.53 | 86.42 | −0.192 | chloroplast |
| TaTPP10-A | TraesCS6A02G301800.1 | 1 | I | 1251 | 6A | reverse | 535151913 | 535154867 | 618079260 | 8 | 9 | 416 | 45.37 | 9.26 | 44.89 | 72.28 | −0.305 | other |
| TaTPP10-B | TraesCS6B02G330900.1 | 3 | I | 1224 | 6B | reverse | 581079293 | 581082545 | 720988478 | 7 | 8 | 407 | 44.29 | 8.64 | 42.6 | 77.69 | −0.147 | mitochondrion |
| TaTPP10-D | TraesCS6D02G281100.1 | 1 | I | 1110 | 6D | reverse | 388537685 | 388540648 | 473592718 | 8 | 9 | 369 | 40.25 | 8.79 | 37.81 | 76.99 | −0.164 | other |
| TaTPP11-A | TraesCS7A02G180800.1 | 1 | I | 1086 | 7A | reverse | 135006112 | 135008690 | 736706236 | 9 | 10 | 361 | 39.54 | 8.41 | 50.69 | 81.39 | −0.219 | chloroplast |
| TaTPP11-B | TraesCS7B02G085800.1 | 1 | I | 1095 | 7B | reverse | 97972425 | 97975038 | 750620385 | 9 | 10 | 364 | 40.16 | 8.11 | 50.16 | 79.09 | −0.274 | chloroplast |
| TaTPP11-D | TraesCS7D02G182600.1 | 1 | I | 1092 | 7D | reverse | 136013159 | 136015620 | 638686055 | 9 | 10 | 363 | 39.91 | 8.60 | 51.06 | 79.56 | −0.268 | chloroplast |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.A.; Rahman, M.M.; Rahman, M.M.; Jin, X.; Sun, L.; Zhao, K.; Wang, S.; Sikdar, A.; Noor, H.; Jeon, J.-S.; et al. In Silico and Transcription Analysis of Trehalose-6-phosphate Phosphatase Gene Family of Wheat: Trehalose Synthesis Genes Contribute to Salinity, Drought Stress and Leaf Senescence. Genes 2021, 12, 1652. https://doi.org/10.3390/genes12111652
Islam MA, Rahman MM, Rahman MM, Jin X, Sun L, Zhao K, Wang S, Sikdar A, Noor H, Jeon J-S, et al. In Silico and Transcription Analysis of Trehalose-6-phosphate Phosphatase Gene Family of Wheat: Trehalose Synthesis Genes Contribute to Salinity, Drought Stress and Leaf Senescence. Genes. 2021; 12(11):1652. https://doi.org/10.3390/genes12111652
Chicago/Turabian StyleIslam, Md Ashraful, Md Mustafizur Rahman, Md Mizanor Rahman, Xiujuan Jin, Lili Sun, Kai Zhao, Shuguang Wang, Ashim Sikdar, Hafeez Noor, Jong-Seong Jeon, and et al. 2021. "In Silico and Transcription Analysis of Trehalose-6-phosphate Phosphatase Gene Family of Wheat: Trehalose Synthesis Genes Contribute to Salinity, Drought Stress and Leaf Senescence" Genes 12, no. 11: 1652. https://doi.org/10.3390/genes12111652
APA StyleIslam, M. A., Rahman, M. M., Rahman, M. M., Jin, X., Sun, L., Zhao, K., Wang, S., Sikdar, A., Noor, H., Jeon, J.-S., Zhang, W., & Sun, D. (2021). In Silico and Transcription Analysis of Trehalose-6-phosphate Phosphatase Gene Family of Wheat: Trehalose Synthesis Genes Contribute to Salinity, Drought Stress and Leaf Senescence. Genes, 12(11), 1652. https://doi.org/10.3390/genes12111652