Transcriptome Analysis of Pistacia vera Inflorescence Buds in Bearing and Non-Bearing Shoots Reveals the Molecular Mechanism Causing Premature Flower Bud Abscission

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, RNA Extraction, Processing and Sequencing
2.2. De Novo Assembly, Evaluation and Annotation
2.3. Differentially Expressed Genes (DEG) between Stages
2.4. Gene Enrichment and Functional Analysis
3. Results
3.1. De Novo Transcriptome Assembly and Annotation
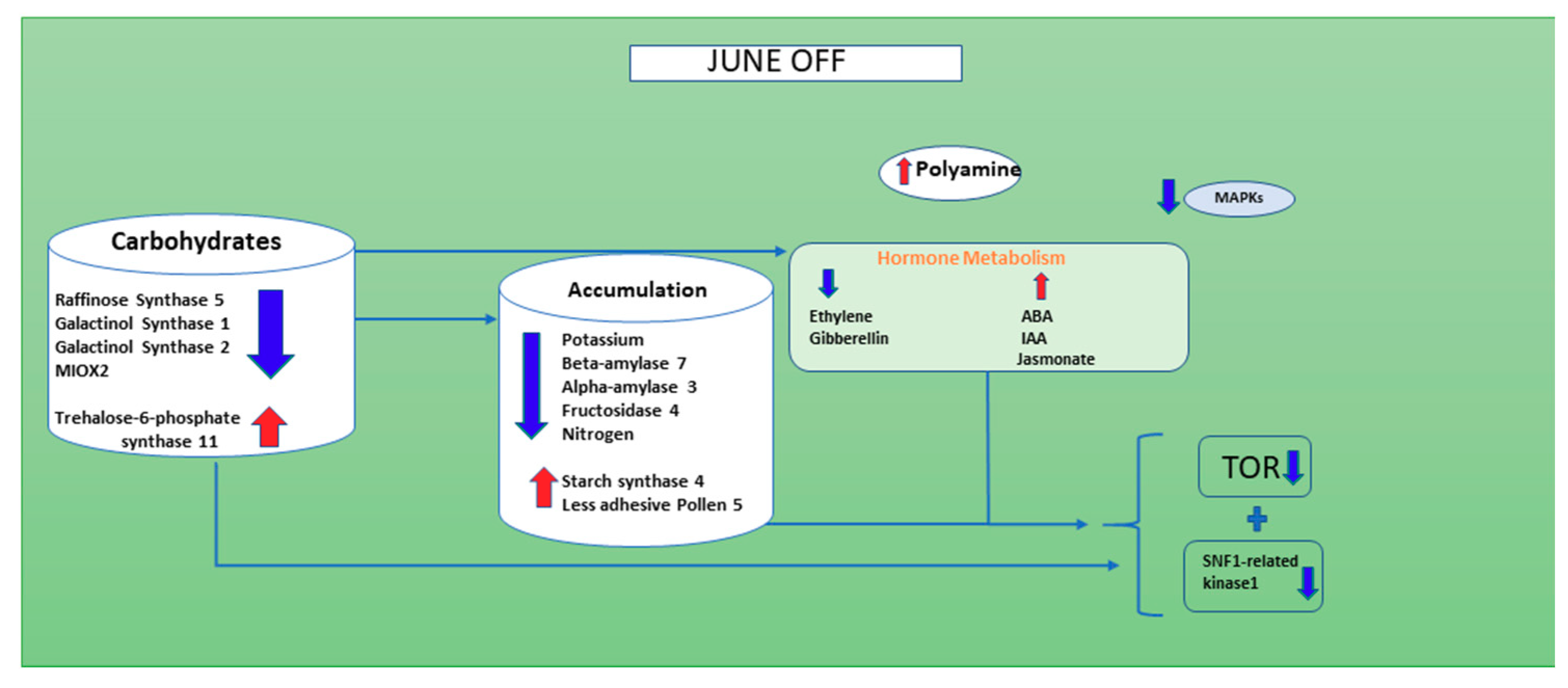
3.2. Effect of Crop Load on Photosynthesis During in June “OFF” vs. June “ON” Inflorescence Buds
3.3. Effect of Crop Load on Starch Metabolism in—June “OFF” vs. June “ON” Inflorescence Buds
3.4. Effect of Crop Load Status on Polyamine and Transcription Factors in Inflorescence Buds June “OFF” vs. June “ON” Shoots
3.5. Gene Set and Pathway Enrichment Analysis During in June “OFF” vs. June “ON” Inflorescence Buds
3.6. Effect of Crop Load on Hormone Metabolism in June “OFF” vs. June “ON” Inflorescence Buds
3.7. Effect of Crop Load on Ubiquitin and Autophagy Dependent Degradation in June “OFF” vs. June “ON” Inflorescence Buds
3.8. Effect of Crop Load on Carbohydrate Metabolism and Mobilization in June “OFF” vs. June “ON” Inflorescence Buds
3.9. Comparison between “ON” and “OFF” Inflorescence Buds of “JUNE” and “JULY”
Effects of Crop Load in July “OFF” vs. July “ON” Inflorescence Buds
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Caruso, T.; Fabbri, A.; Giovannini, D. Inflorescence bud growth, development and abscission in shoots of bearing and disbudded ‘Bianca’ pistachio trees. J. Hortic. Sci. 1995, 70, 857–866. [Google Scholar] [CrossRef]
- Khezri, M.; Heerema, R.; Brar, G.; Ferguson, L. Alternate bearing in pistachio (Pistacia vera L.): A review. Trees 2020. [Google Scholar] [CrossRef]
- Crane, J.C.; Iwakiri, B.T. Reconsideration of the cause of inflorescence bud abscission in Pistachio. Hortic. Sci. 1987, 22, 1315–1316. [Google Scholar]
- Ferguson, L.; Kallsen, C.E. The pistachio tree: Physiology and botany. In Pistachio Production Manual; Ferguson, L., Haviland, D.R., Eds.; University of California, Agricultural and Natural Resources Publication: Richmond, CA, USA, 2016; Volume 3545, pp. 17–19. [Google Scholar]
- Crane, J.C.; Nelson, M.M. Effect of crop load, girdling and auxin application on alternate bearing of the pistachio. J. Am. Soc. Hortic. Sci. 1972, 97, 337–339. [Google Scholar]
- Sparks, D.; Davis, J.T. Alternate bearing relates to carbohydrates. Pecan Q. 1974, 8, 20–28. [Google Scholar]
- Takeda, F.; Crane, J.C. Abscisic-Acid in Pistachio as Related to Inflorescence Bud Abscission. J. Am. Soc. Hortic. Sci. 1980, 105, 573–576. [Google Scholar]
- Spann, T.M.; Beede, R.H.; Dejong, T.M. Seasonal carbohydrate storage and mobilization in bearing and non-bearing pistachio (Pistacia vera) trees. Tree Physiol. 2008, 28, 207–213. [Google Scholar] [CrossRef]
- Vemmos, S.N.; Papagiannopoulou, A.; Coward, S. Effects of shoot girdling on photosynthetic capacity, leaf carbohydrate, and bud abscission in pistachio (Pistacia vera L.). Photosynthetica 2012, 50, 35–48. [Google Scholar] [CrossRef]
- Baninasab, B.; Rahemi, M.; Shariatmadari, H. Seasonal Changes in Mineral Content of Different Organs in the Alternate Bearing of Pistachio Trees. Commun. Soil Sci. Plant Anal. 2007, 38, 241–258. [Google Scholar] [CrossRef]
- Nzima, M.D.; Martin, G.C.; Nishijima, C. Leaf Development, Dry Matter Accumulation, and Distribution within Branches of Alternate-bearing ‘Kerman’ Pistachio Trees. J. Am. Soc. Hortic. Sci. 1997, 122, 31–37. [Google Scholar] [CrossRef]
- Stevenson, M.; Shackel, K.; Ferguson, L. Shoot Length Distribution and Its Relation to Yield of Alternate-bearing Pistachio Trees. J. Am. Soc. Hortic. Sci. 2000, 125, 165–168. [Google Scholar] [CrossRef]
- Marra, F.; Barone, E.; Motisi, A.; Sidari, M.; Caruso, T. Dry Matter Accumulation and Carbohydrate Content within Branches of Fruiting and Deblossomed Pistachio (Pistacia vera L.) Trees. Acta Hortic. 1998, 470, 331–339. [Google Scholar] [CrossRef]
- Marra, F.; Roxas, A.A.; Marino, G.; Caruso, T. Seasonal changes in starch content in pistachio organs as related to crop load. Acta Hortic. 2018, 1229, 171–176. [Google Scholar] [CrossRef]
- Gündeşli, M.A.; Kafkas, S.; Zarifikhosroshahi, M.; Kafkas, N.E. Role of endogenous polyamines in the alternate bearing phenomenon in pistachio. Turk. J. Agric. 2019, 43, 265–274. [Google Scholar] [CrossRef]
- Guitton, B.; Kelner, J.J.; Celton, J.M.; Sabau, X.; Renou, J.P.; Chagné, D.; Costes, E. Analysis of transcripts differentially expressed between fruited and deflowered ‘Gala’ adult trees: A contribution to biennial bearing understanding in apple. BMC Plant Biol. 2016, 16, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Shalom, L.; Samuels, S.; Zur, N.; Shlizerman, L.; Zemach, H.; Weissberg, M.; Ophir, R.; Blumwald, E.; Sadka, A. Alternate Bearing in Citrus: Changes in the Expression of Flowering Control Genes and in Global Gene Expression in ON-versus OFF-Crop Trees. PLoS ONE 2012, 7, e46930. [Google Scholar] [CrossRef]
- Yanik, H.; Turktas, M.; Dundar, E.; Hernandez, P.; Dorado, G.; Unver, T. Genome-wide identification of alternate bearing-associated microRNAs (miRNAs) in olive (Olea europaea L.). BMC Plant Biol. 2013, 13, 10. [Google Scholar] [CrossRef]
- Martinelli, F.; Marchese, A.; Balan, B.; Giovino, A.; Caruso, T.; Fretto, S.; Marra, F. RNA-Seq analysis to investigate alternate bearing mechanism in Pistacia vera L. Acta Hortic. 2018, 1229, 71–78. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA–Seq data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Smith-Unna, R.; Boursnell, C.; Patro, R.; Hibberd, J.M.; Kelly, S. TransRate: Reference-free quality assessment of de novo transcriptome assemblies. Genome Res. 2016, 26, 1134–1144. [Google Scholar] [CrossRef] [PubMed]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; Mccarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Thimm, O.; Bläsing, O.; Gibon, Y.; Nagel, A.; Meyer, S.; Krüger, P.; Selbig, J.; Müller, L.A.; Rhee, S.Y.; Stitt, M. Mapman: A user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 2004, 37, 914–939. [Google Scholar] [CrossRef]
- Huang, D.; Sherman, B.T.; Tan, Q.; Collins, J.R.; Alvord, W.G.; Roayaei, J.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. The DAVID Gene Functional Classification Tool: A novel biological module-centric algorithm to functionally analyze large gene lists. Genome Biol. 2007, 8, R183. [Google Scholar] [CrossRef] [PubMed]
- Kozlowski, T.T.; Pallardy, S.G. Physiological Regulation of Vegetative Growth. Growth Control Woody Plants 1997, 73–165. [Google Scholar] [CrossRef]
- Marra, F.P.; Barone, E.; Mantia, M.L.; Caruso, T. Toward the definition of a carbon budget model: Seasonal variation and temperature effect on respiration rate of vegetative and reproductive organs of pistachio trees (Pistacia vera). Tree Physiol. 2009, 29, 1095–1103. [Google Scholar] [CrossRef]
- Weinbaum, S.; Picchioni, G.; Muraoka, T.; Ferguson, L.; Brown, P. Fertilizer Nitrogen and Boron Uptake, Storage, and Allocation Vary during the Alternate-bearing Cycle in Pistachio Trees. J. Am. Soc. Hortic. Sci. 1994, 119, 24–31. [Google Scholar] [CrossRef]
- Marino, G.; Ferguson, L.; Caruso, T.; Roxas, A.A.; Marra, F. A carbon budget model to predict branch carbohydrate deficiencies as a function of water stress and crop load in pistachio (Pistacia vera L.). Acta Hortic. 2018, 183–188. [Google Scholar] [CrossRef]
- Lu, C.-A.; Lim, E.-K.; Yu, S.-M. Sugar Response Sequence in the Promoter of a Rice α-Amylase Gene Serves as a Transcriptional Enhancer. J. Biol. Chem. 1998, 273, 10120–10131. [Google Scholar] [CrossRef] [PubMed]
- Zuther, E.; Schulz, E.; Childs, L.H.; Hincha, D.K. Clinal variation in the non-acclimated and cold-acclimated freezing tolerance of Arabidopsis thaliana accessions. Plant Cell Environ. 2012, 35, 1860–1878. [Google Scholar] [CrossRef] [PubMed]
- Valluru, R.; Ende, W.V.D. Myo-inositol and beyond—Emerging networks under stress. Plant Sci. 2011, 181, 387–400. [Google Scholar] [CrossRef]
- Sengupta, S.; Mukherjee, S.; Basak, P.; Majumder, A.L. Significance of galactinol and raffinose family oligosaccharide synthesis in plants. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Couée, I.; Sulmon, C.; Gouesbet, G.; Amrani, A.E. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef]
- Xue, H.; Chen, X.; Li, G. Involvement of phospholipid signaling in plant growth and hormone effects. Curr. Opin. Plant Biol. 2007, 10, 483–489. [Google Scholar] [CrossRef]
- Thole, J.M.; Nielsen, E. Phosphoinositides in plants: Novel functions in membrane trafficking. Curr. Opin. Plant Biol. 2008, 11, 620–631. [Google Scholar] [CrossRef]
- Okada, M.; Ye, K. Nuclear phosphoinositide signaling regulates messenger RNA export. RNA Biol. 2009, 6, 12–16. [Google Scholar] [CrossRef]
- Kanter, U.; Usadel, B.; Guerineau, F.; Li, Y.; Pauly, M.; Tenhaken, R. The inositol oxygenase gene family of Arabidopsis is involved in the biosynthesis of nucleotide sugar precursors for cell-wall matrix polysaccharides. Planta 2005, 221, 243–254. [Google Scholar] [CrossRef]
- Ponnu, J.; Wahl, V.; Schmid, M. Trehalose-6-Phosphate: Connecting Plant Metabolism and Development. Front. Plant Sci. 2011, 2, 70. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, C.M.; Lunn, J.E. A Tale of Two Sugars: Trehalose 6-Phosphate and Sucrose. Plant Physiol. 2016, 172, 7–27. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.J.; Jhurreea, D.; Zhang, Y.; Primavesi, L.F.; Delatte, T.; Schluepmann, H.; Wingler, A. Up-regulation of biosynthetic processes associated with growth by trehalose 6-phosphate. Plant Signal. Behav. 2010, 5, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Escobar, R.; Moreno, R.; García-Creus, M. Seasonal changes of mineral nutrients in olive leaves during the alternate-bearing cycle. Sci. Hortic. 1999, 82, 25–45. [Google Scholar] [CrossRef]
- Proietti, P.; Nasini, L.; Buono, D.D.; D’Amato, R.; Tedeschini, E.; Businelli, D. Selenium protects olive (Olea europaea L.) from drought stress. Sci. Hortic. 2013, 164, 165–171. [Google Scholar] [CrossRef]
- Syvertsen, J.P.; Goni, C.; Otero, A. Fruit load and canopy shading affect leaf characteristics and net gas exchange of Spring navel orange trees. Tree Physiol. 2003, 23, 899–906. [Google Scholar] [CrossRef]
- Marino, G.; Mantia, M.; Caruso, T.; Marra, F.P. Seasonal dynamics of photosynthesis and total carbon gain in bearing and nonbearing pistachio (Pistacia vera L.) shoots. Photosynthetica 2018, 56, 932–941. [Google Scholar] [CrossRef]
- Butler, D.R.; Landsberg, J.J. Respiration rates of apple trees, estimated by CO2-efflux measurements. Plant Cell Environ. 1981, 4, 153–159. [Google Scholar] [CrossRef]
- Morkunas, I.; Borek, S.; Formela, M.; Ratajczak, L. Plant Responses to Sugar Starvation. Carbohydr. Compr. Stud. Glycobiol. Glycotechnol. 2012. [Google Scholar] [CrossRef]
- Rose, T.L.; Bonneau, L.; Der, C.; Marty-Mazars, D.; Marty, F. Starvation-induced expression of autophagy-related genes in Arabidopsis. Biol. Cell 2006, 98, 53–67. [Google Scholar] [CrossRef]
- Contento, A.L.; Kim, S.-J.; Bassham, D.C. Transcriptome Profiling of the Response of Arabidopsis Suspension Culture Cells to Suc Starvation. Plant Physiol. 2004, 135, 2330–2347. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; Tiburcio, A.F. Polyamine Metabolism and Abiotic Stress Tolerance in Plants. Metab. Adapt. Plants Dur. Abiotic Stress 2018, 191–203. [Google Scholar] [CrossRef]
- Nishikawa, F.; Iwasaki, M.; Fukamachi, H.; Nonaka, K.; Imai, A.; Takishita, F.; Yano, T.; Endo, T. Fruit Bearing Suppresses Citrus FLOWERING LOCUS T Expression in Vegetative Shoots of Satsuma Mandarin (Citrus unshiu Marc.). J. Jpn. Soc. Hortic. Sci. 2012, 81, 48–53. [Google Scholar] [CrossRef]
- Aziz, A. Spermidine and related-metabolic inhibitors modulate sugar and amino acid levels in Vitis vinifera L.: Possible relationships with initial fruitlet abscission. J. Exp. Bot. 2003, 54, 355–363. [Google Scholar] [CrossRef]
- Gomez-Jimenez, M.C.; Paredes, M.A.; Gallardo, M.; Fernandez-Garcia, N.; Olmos, E.; Sanchez-Calle, I.M. Tissue-specific expression of olive S-adenosyl methionine decarboxylase and spermidine synthase genes and polyamine metabolism during flower opening and early fruit development. Planta 2010, 232, 629–647. [Google Scholar] [CrossRef]
- Khezri, M.; Talaie, A.; Javanshah, A.; Hadavi, F. Effect of exogenous application of free polyamines on physiological disorders and yield of ‘Kaleh-Ghoochi’ pistachio shoots (Pistacia vera L.). Sci. Hortic. 2010, 125, 270–276. [Google Scholar] [CrossRef]
- Vemmos, S.; Pontikis, C.; Tolza-Marioli, A. Respiration rate and ethylene production in inflorescence buds of pistachio in relation to alternate bearing. Sci. Hortic. 1994, 57, 165–172. [Google Scholar] [CrossRef]
- Gündeşli, M.A.; Kafkas, S.; Güney, M.; Kafkas, N.E. Changes in endogenous auxin level during flower bud abscission process in Pistachio (Pistacia vera L.). Turk. J. Agric. 2020, 44, 71–82. [Google Scholar] [CrossRef]
- Pontikis, C.A. Effects of 2-Naphthaleneacetic acid on alternate bearing of pistachio. Fruits 1990, 45, 281–285. [Google Scholar]
- Ahn, C.S.; Han, J.-A.; Lee, H.-S.; Lee, S.; Pai, H.-S. The PP2A Regulatory Subunit Tap46, a Component of the TOR Signaling Pathway, Modulates Growth and Metabolism in Plants. Plant Cell 2011, 23, 185–209. [Google Scholar] [CrossRef]
- Marshall, R.S.; Vierstra, R.D. Autophagy: The Master of Bulk and Selective Recycling. Annu. Rev. Plant Biol. 2018, 69, 173–208. [Google Scholar] [CrossRef] [PubMed]
- Baena-González, E.; Rolland, F.; Thevelein, J.M.; Sheen, J. A central integrator of transcription networks in plant stress and energy signalling. Nature 2007, 448, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Wurzinger, B.; Nukarinen, E.; Nägele, T.; Weckwerth, W.; Teige, M. The SnRK1 Kinase as Central Mediator of Energy Signaling between Different Organelles. Plant Physiol. 2018, 176, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Arenas-Huertero, F.; Arroyo, A.; Zhou, L.; Sheen, J.; León, P. Analysis of Arabidopsis glucose insensitive mutants, gin5 and gin6, reveals a central role of the plant hormone ABA in the regulation of plant vegetative development by sugar. Genes Dev. 2000, 14, 2085–2096. [Google Scholar] [PubMed]
- Guo, H.; Ecker, J.R. The ethylene signaling pathway: New insights. Curr. Opin. Plant Biol. 2004, 7, 40–49. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Tawaratsumida, T.; Kondo, K.; Kasa, S.; Sakamoto, M.; Aoki, N.; Zheng, S.-H.; Yuasa, T.; Iwaya-Inoue, M. Reactive Oxygen Species Are Involved in Gibberellin/Abscisic Acid Signaling in Barley Aleurone Cells. Plant Physiol. 2012, 158, 1705–1714. [Google Scholar] [CrossRef]
- Takács, Z.; Poór, P.; Tari, I. Comparison of polyamine metabolism in tomato plants exposed to different concentrations of salicylic acid under light or dark conditions. Plant Physiol. Biochem. 2016, 108, 266–278. [Google Scholar] [CrossRef]
- Moschou, P.N.; Wu, J.; Cona, A.; Tavladoraki, P.; Angelini, R.; Roubelakis-Angelakis, K.A. The polyamines and their catabolic products are significant players in the turnover of nitrogenous molecules in plants. J. Exp. Bot. 2012, 63, 5003–5015. [Google Scholar] [CrossRef]
- Corpas, F.J.; Río, L.A.D.; Palma, J.M. Plant peroxisomes at the crossroad of NO and H2O2 metabolism. J. Integr. Plant Biol. 2019. [Google Scholar] [CrossRef]
- Moschou, P.N.; Paschalidis, K.A.; Delis, I.D.; Andriopoulou, A.H.; Lagiotis, G.D.; Yakoumakis, D.I.; Roubelakis-Angelakis, K.A. Spermidine Exodus and Oxidation in the Apoplast Induced by Abiotic Stress Is Responsible for H2O2 Signatures That Direct Tolerance Responses in Tobacco. Plant Cell 2008, 20, 1708–1724. [Google Scholar] [CrossRef]
- Yang, H.; Kim, H.-J.; Chen, H.; Lü, Y.; Lu, X.; Wang, C.; Zhou, B. Reactive oxygen species and nitric oxide induce senescence of rudimentary leaves and the expression profiles of the related genes in Litchi chinensis. Hortic. Res. 2018, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Pitino, M.; Armstrong, C.M.; Duan, Y. Molecular mechanisms behind the accumulation of ATP and H2O2 in citrus plants in response to ‘Candidatus Liberibacter asiaticus’ infection. Hortic. Res. 2017, 4, 1–8. [Google Scholar] [CrossRef]
- López-Huertas, E.; Corpas, F.J.; Sandalio, L.M.; Río, L.A.D. Characterization of membrane polypeptides from pea leaf peroxisomes involved in superoxide radical generation. Biochem. J. 1999, 337, 531. [Google Scholar] [CrossRef]
- Matthiadis, A.; Long, T.A. Further insight into BRUTUS domain composition and functionality. Plant Signal. Behav. 2016, 11, e1204508. [Google Scholar] [CrossRef]
- Agarwal, P.; Baranwal, V.K.; Khurana, P. Genome-wide Analysis of bZIP Transcription Factors in wheat and Functional Characterization of a TabZIP under Abiotic Stress. Sci. Rep. 2019, 9, 4608. [Google Scholar] [CrossRef]
- Sharoni, A.M.; Nuruzzaman, M.; Satoh, K.; Shimizu, T.; Kondoh, H.; Sasaya, T.; Choi, I.-R.; Omura, T.; Kikuchi, S. Gene Structures, Classification and Expression Models of the AP2/EREBP Transcription Factor Family in Rice. Plant Cell Physiol. 2010, 52, 344–360. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Mccormack, M.; Li, L.; Hall, Q.; Xiang, C.; Sheen, J. Glucose–TOR signalling reprograms the transcriptome and activates meristems. Nature 2013, 496, 181–186. [Google Scholar] [CrossRef]
- Kamiab, F.; Heidari Salehabad, M.; Zamanibahramabadi, E. Evaluation the effects of foliar treatments of polyamines and some organic acids on quantitative and qualitative traits in some pistachio cultivars. J. Nuts 2015, 6, 131–142. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison (Inflorescence Bud) | Differentially Expressed Genes | Up-Regulated | Down-Regulated |
|---|---|---|---|
| July “OFF” vs. July “ON” | 1087 | 247 | 840 |
| June “OFF” vs. July “OFF” | 2299 | 976 | 1323 |
| June “ON” vs. July “OFF” | 2450 | 591 | 1859 |
| June “OFF” vs. July “ON” | 2768 | 820 | 1948 |
| June “ON” vs. July “ON” | 3882 | 712 | 3170 |
| June “OFF” vs. June “ON” | 1844 | 1409 | 435 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benny, J.; Marra, F.P.; Giovino, A.; Balan, B.; Caruso, T.; Martinelli, F.; Marchese, A. Transcriptome Analysis of Pistacia vera Inflorescence Buds in Bearing and Non-Bearing Shoots Reveals the Molecular Mechanism Causing Premature Flower Bud Abscission. Genes 2020, 11, 851. https://doi.org/10.3390/genes11080851
Benny J, Marra FP, Giovino A, Balan B, Caruso T, Martinelli F, Marchese A. Transcriptome Analysis of Pistacia vera Inflorescence Buds in Bearing and Non-Bearing Shoots Reveals the Molecular Mechanism Causing Premature Flower Bud Abscission. Genes. 2020; 11(8):851. https://doi.org/10.3390/genes11080851
Chicago/Turabian StyleBenny, Jubina, Francesco Paolo Marra, Antonio Giovino, Bipin Balan, Tiziano Caruso, Federico Martinelli, and Annalisa Marchese. 2020. "Transcriptome Analysis of Pistacia vera Inflorescence Buds in Bearing and Non-Bearing Shoots Reveals the Molecular Mechanism Causing Premature Flower Bud Abscission" Genes 11, no. 8: 851. https://doi.org/10.3390/genes11080851
APA StyleBenny, J., Marra, F. P., Giovino, A., Balan, B., Caruso, T., Martinelli, F., & Marchese, A. (2020). Transcriptome Analysis of Pistacia vera Inflorescence Buds in Bearing and Non-Bearing Shoots Reveals the Molecular Mechanism Causing Premature Flower Bud Abscission. Genes, 11(8), 851. https://doi.org/10.3390/genes11080851