TRPV1: A Potential Drug Target for Treating Various Diseases

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
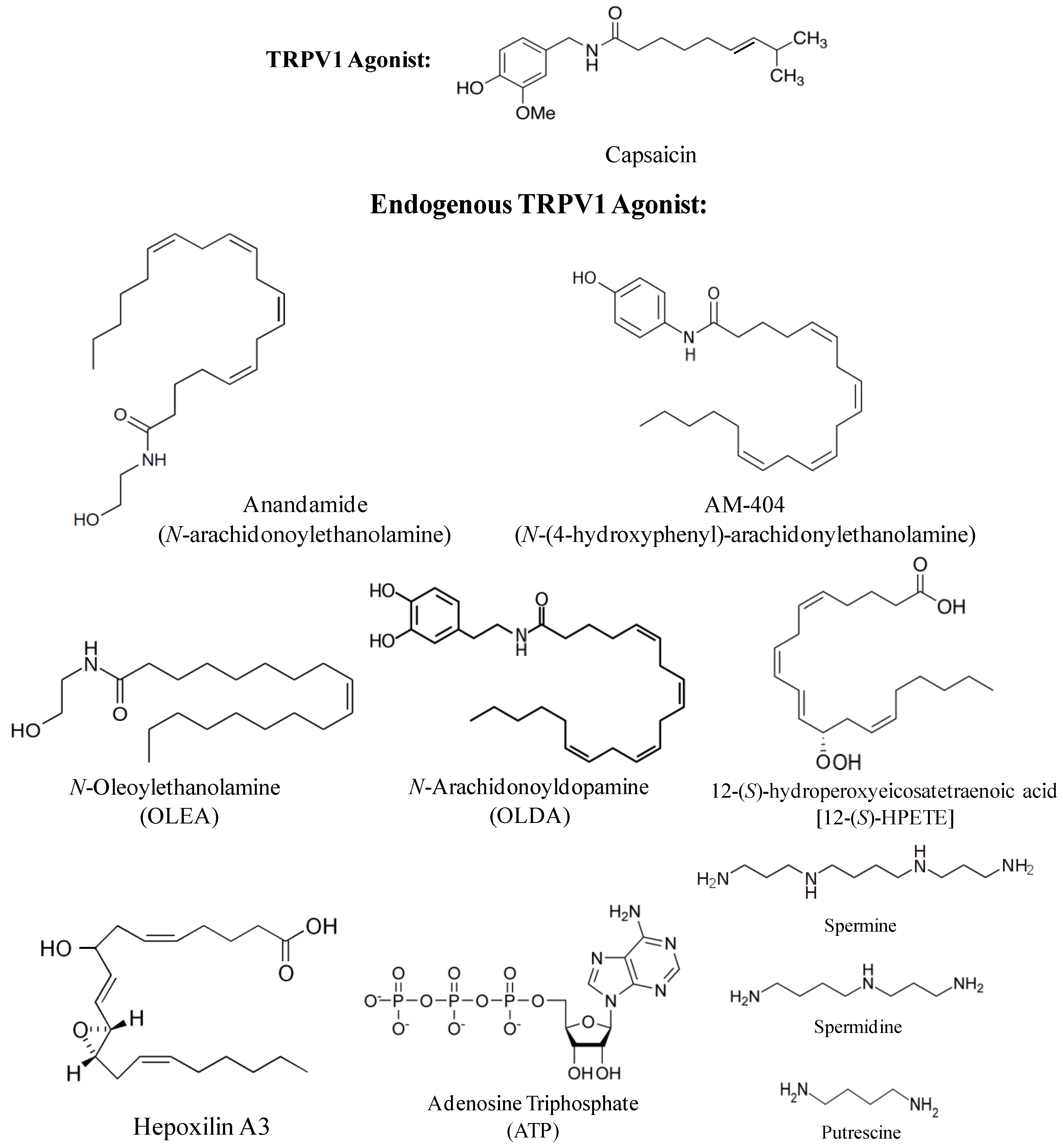
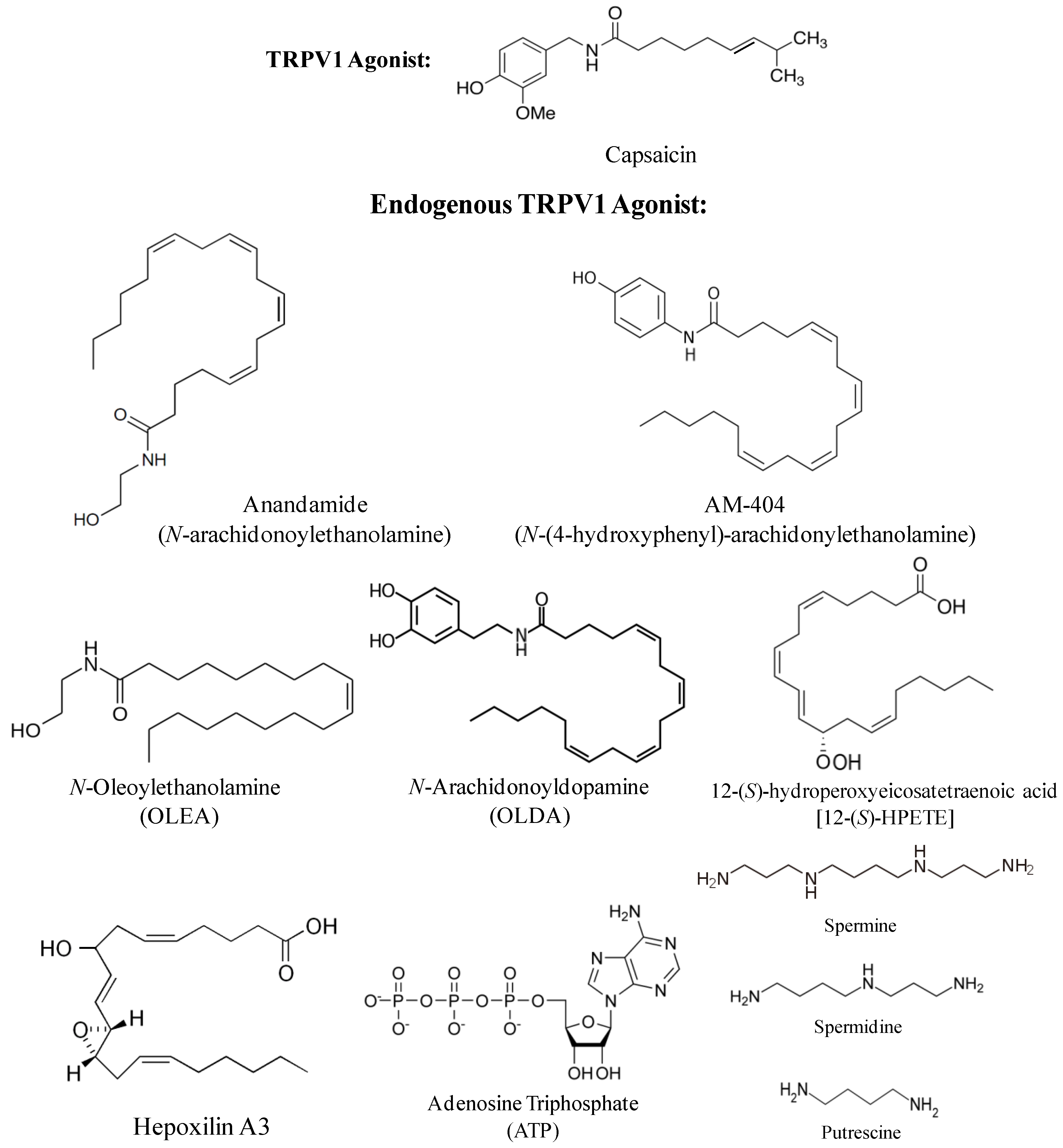
2. Endogenous TRPV1 Agonists
3. Functions of TRPV1 Channel
3.1. Role of TRPV1 in Thermal Sensation
3.2. Role of TRPV1 in Diabetes and Obesity
3.3. TRPV1 in Ototoxicity of Cisplatin and Aminoglycoside Antibiotics
3.4. TRPV1 in the Bladder
3.5. TRPV1 in the Lung
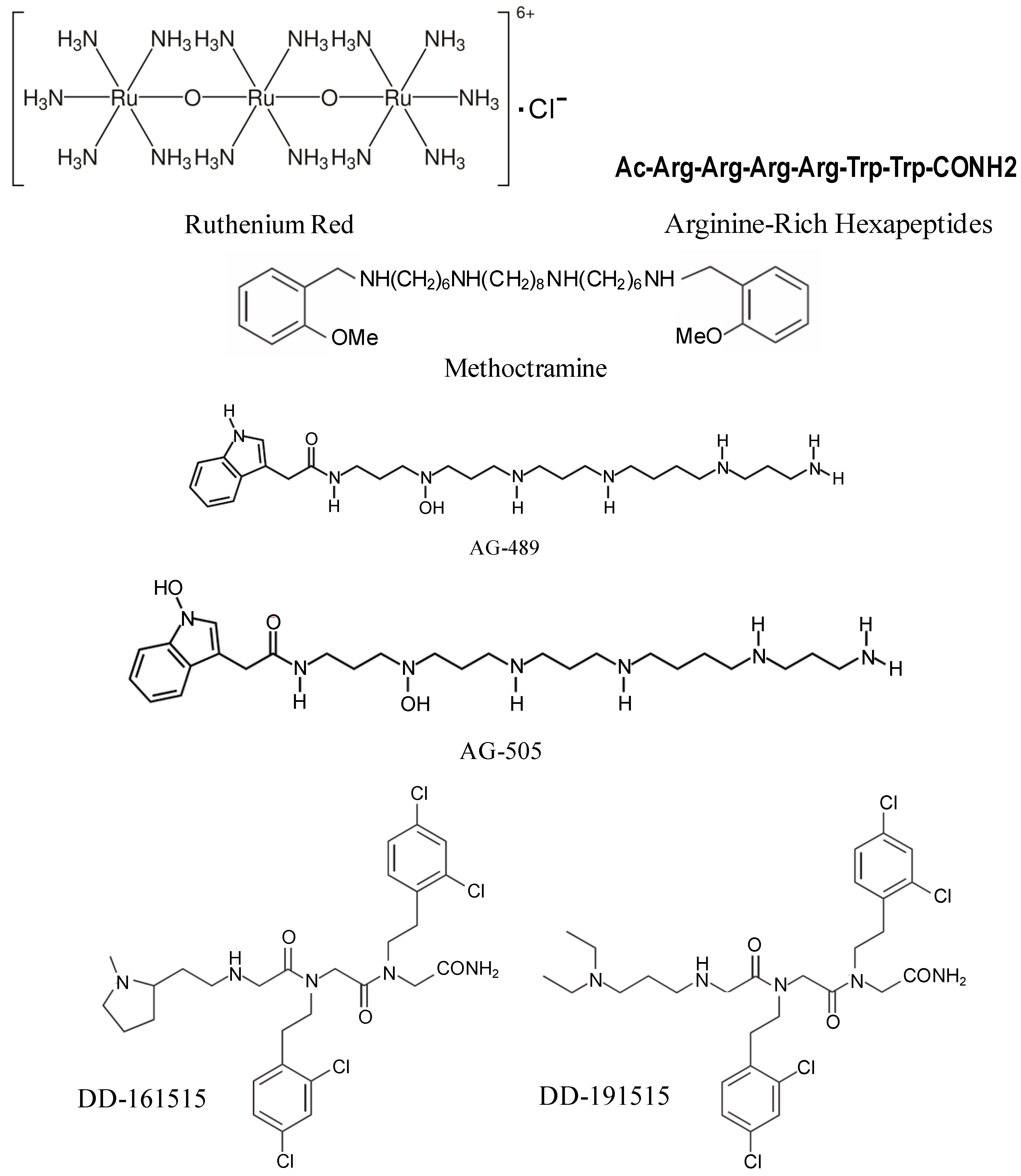
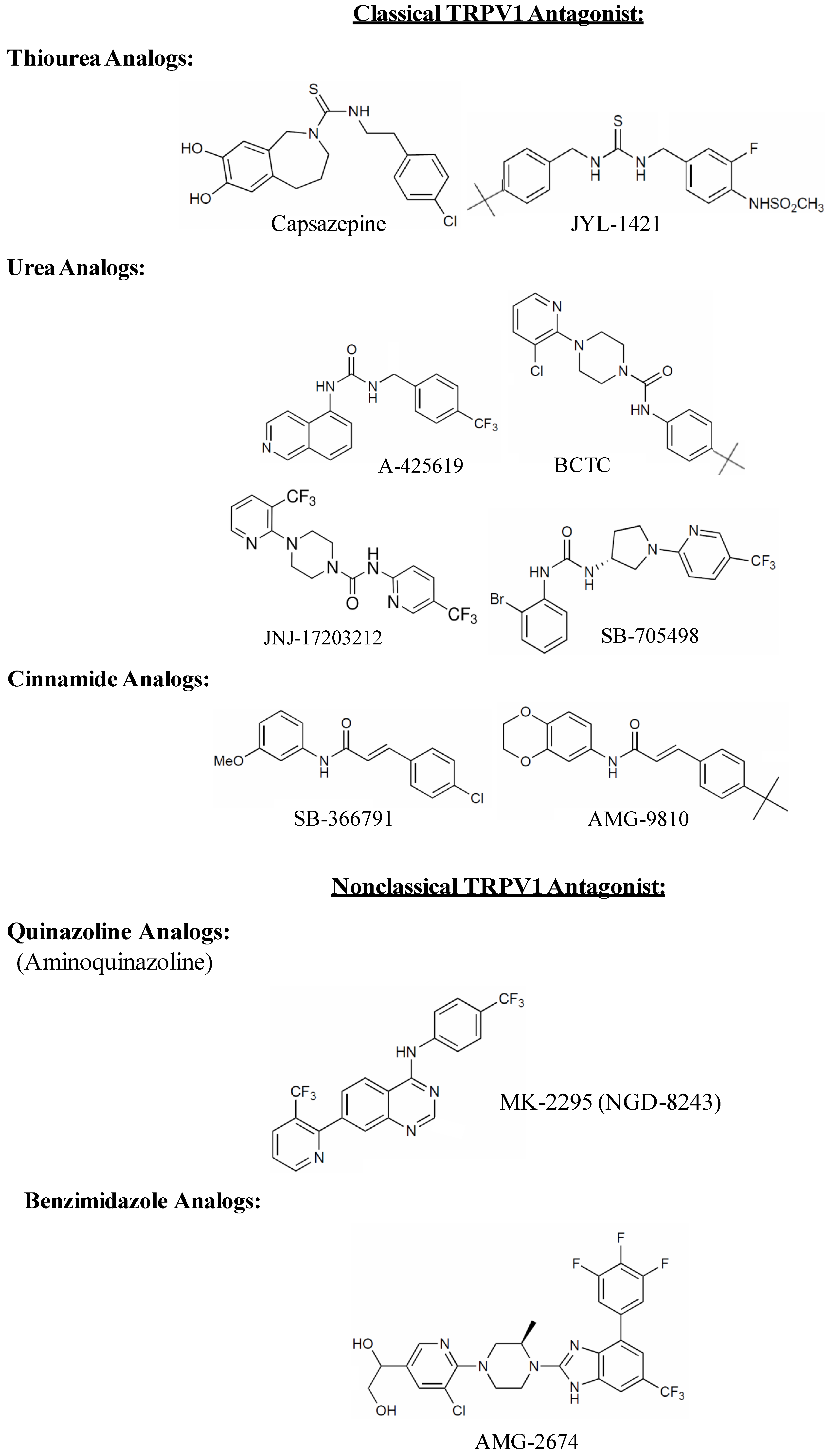
4. TRPV1 Antagonists
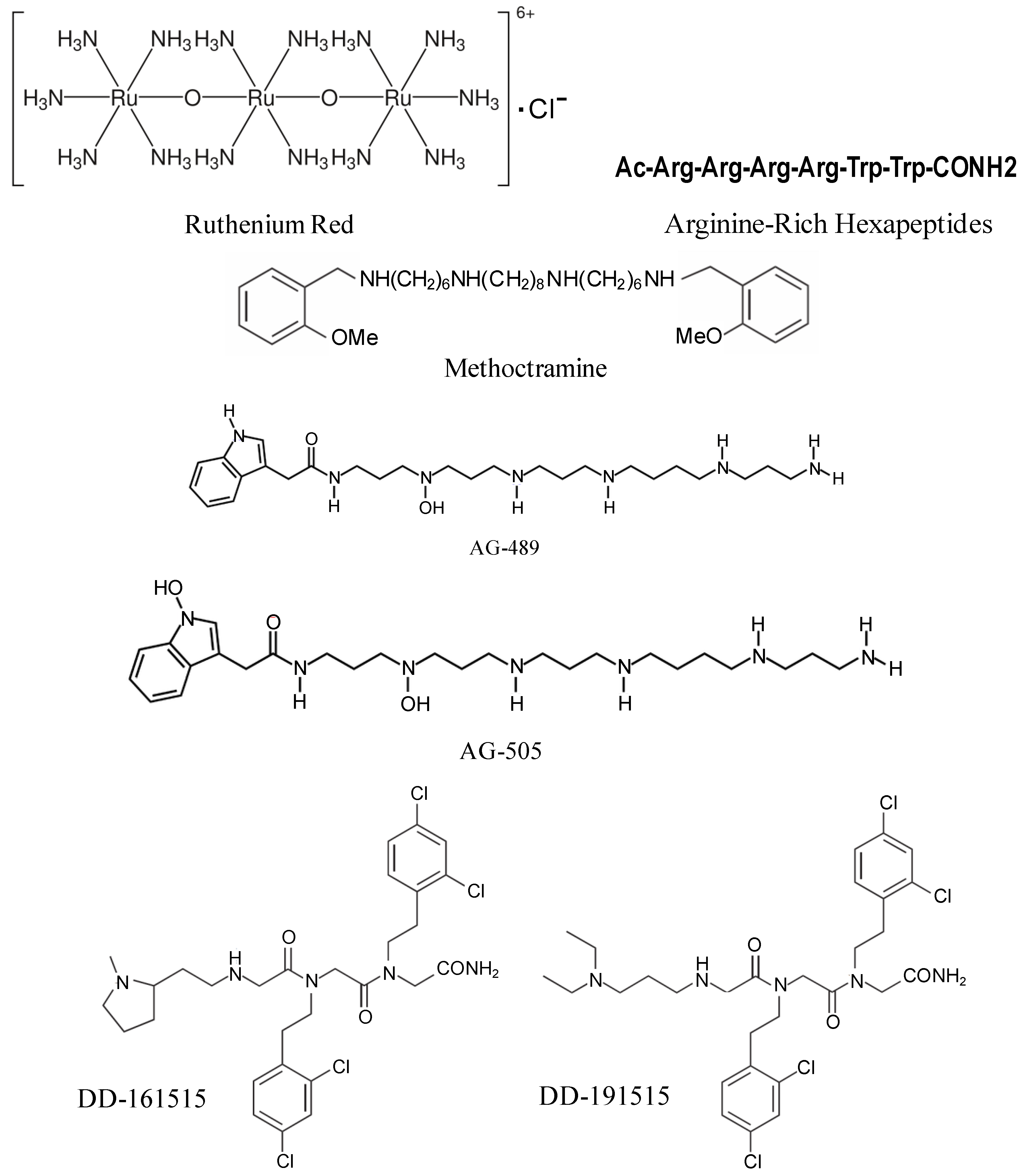
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Caterina, M.J.; Julius, D. The vanilloid receptor: A molecular gateway to the pain pathway. Annu. Rev. Neurosci. 2001, 24, 487–517. [Google Scholar] [CrossRef]
- Holzer, P. Capsaicin: Cellular targets, mechanisms of action, and selectivity for thin sensory neurons. Pharmacol. Rev. 1991, 43, 143–201. [Google Scholar]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Mezey, E.; Toth, Z.E.; Cortright, D.N.; Arzubi, M.K.; Krause, J.E.; Elde, R.; Guo, A.; Blumberg, P.M.; Szallasi, A. Distribution of mRNA for vanilloid receptor subtype 1 (VR1), and VR1-like immunoreactivity, in the central nervous system of the rat and human. Proc. Natl. Acad. Sci. USA 2000, 97, 3655–3660. [Google Scholar] [CrossRef]
- Roberts, J.C.; Davis, J.B.; Benham, C.D. [3H]Resiniferatoxin autoradiography in the CNS of wild-type and TRPV1 null mice defines TRPV1 (VR-1) protein distribution. Brain Res. 2004, 995, 176–183. [Google Scholar] [CrossRef]
- Van der Stelt, M.; di Marzo, V. Endovanilloids. Putative endogenous ligands of transient receptor potential vanilloid 1 channels. Eur. J. Biochem. 2004, 271, 1827–1834. [Google Scholar] [CrossRef]
- Zygmunt, P.M.; Petersson, J.; Andersson, D.A.; Chuang, H.; Sorgard, M.; di Marzo, V.; Julius, D.; Hogestatt, E.D. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 1999, 400, 452–457. [Google Scholar] [CrossRef]
- Ross, R.A. Anandamide and vanilloid TRPV1 receptors. Br. J. Pharmacol. 2003, 140, 790–801. [Google Scholar] [CrossRef]
- McVey, D.C.; Schmid, P.C.; Schmid, H.H.; Vigna, S.R. Endocannabinoids induce ileitis in rats via the capsaicin receptor (VR1). J. Pharmacol. Exp. Ther. 2003, 304, 713–722. [Google Scholar] [CrossRef]
- Lastres-Becker, I.; de Miguel, R.; de Petrocellis, L.; Makriyannis, A.; di Marzo, V.; Fernandez-Ruiz, J. Compounds acting at the endocannabinoid and/or endovanilloid systems reduce hyperkinesia in a rat model of Huntington’s disease. J. Neurochem. 2003, 84, 1097–1109. [Google Scholar] [CrossRef]
- Lastres-Becker, I.; Hansen, H.H.; Berrendero, F.; de Miguel, R.; Perez-Rosado, A.; Manzanares, J.; Ramos, J.A.; Fernandez-Ruiz, J. Alleviation of motor hyperactivity and neurochemical deficits by endocannabinoid uptake inhibition in a rat model of Huntington’s disease. Synapse 2002, 44, 23–35. [Google Scholar] [CrossRef]
- Almasi, R.; Szoke, E.; Bolcskei, K.; Varga, A.; Riedl, Z.; Sandor, Z.; Szolcsanyi, J.; Petho, G. Actions of 3-methyl-N-oleoyldopamine, 4-methyl-N-oleoyldopamine and N-oleoylethanolamide on the rat TRPV1 receptor in vitro and in vivo. Life Sci. 2008, 82, 644–651. [Google Scholar] [CrossRef]
- Wang, X.; Miyares, R.L.; Ahern, G.P. Oleoylethanolamide excites vagal sensory neurones, induces visceral pain and reduces short-term food intake in mice via capsaicin receptor TRPV1. J. Physiol. 2005, 564, 541–547. [Google Scholar] [CrossRef]
- Toth, A.; Kedei, N.; Wang, Y.; Blumberg, P.M. Arachidonyl dopamine as a ligand for the vanilloid receptor VR1 of the rat. Life Sci. 2003, 73, 487–498. [Google Scholar] [CrossRef]
- Huang, S.M.; Bisogno, T.; Trevisani, M.; Al-Hayani, A.; de Petrocellis, L.; Fezza, F.; Tognetto, M.; Petros, T.J.; Krey, J.F.; Chu, C.J.; et al. An endogenous capsaicin-like substance with high potency at recombinant and native vanilloid VR1 receptors. Proc. Natl. Acad. Sci. USA 2002, 99, 8400–8405. [Google Scholar] [CrossRef]
- Harrison, S.; de Petrocellis, L.; Trevisani, M.; Benvenuti, F.; Bifulco, M.; Geppetti, P.; di Marzo, V. Capsaicin-like effects of N-arachidonoyl-dopamine in the isolated guinea pig bronchi and urinary bladder. Eur. J. Pharmacol. 2003, 475, 107–114. [Google Scholar] [CrossRef]
- Chu, C.J.; Huang, S.M.; de Petrocellis, L.; Bisogno, T.; Ewing, S.A.; Miller, J.D.; Zipkin, R.E.; Daddario, N.; Appendino, G.; di Marzo, V.; et al. N-oleoyldopamine, a novel endogenous capsaicin-like lipid that produces hyperalgesia. J. Biol. Chem. 2003, 278, 13633–13639. [Google Scholar] [CrossRef]
- Phillis, J.W.; Horrocks, L.A.; Farooqui, A.A. Cyclooxygenases, lipoxygenases, and epoxygenasesin CNS: Their role and involvement in neurological disorders. Brain Res. Rev. 2006, 52, 201–243. [Google Scholar] [CrossRef]
- Hwang, S.W.; Cho, H.; Kwak, J.; Lee, S.Y.; Kang, C.J.; Jung, J.; Cho, S.; Min, K.H.; Suh, Y.G.; Kim, D.; Oh, U. Direct activation of capsaicin receptors by products of lipoxygenases: Endogenous capsaicin-like substances. Proc. Natl. Acad. Sci. USA 2000, 97, 6155–6160. [Google Scholar] [CrossRef]
- Shin, J.; Cho, H.; Hwang, S.W.; Jung, J.; Shin, C.Y.; Lee, S.Y.; Kim, S.H.; Lee, M.G.; Choi, Y.H.; Kim, J.; et al. Bradykinin-12-lipoxygenase-VR1 signaling pathway for inflammatory hyperalgesia. Proc. Natl. Acad. Sci. USA 2002, 99, 10150–10155. [Google Scholar] [CrossRef]
- Gregus, A.M.; Doolen, S.; Dumlao, D.S.; Buczynski, M.W.; Takasusuki, T.; Fitzsimmons, B.L.; Hua, X.Y.; Taylor, B.K.; Dennis, E.A.; Yaksh, T.L. Spinal 12-lipoxygenase-derived hepoxilin A3 contributes to inflammatory hyperalgesia via activation of TRPV1 and TRPA1 receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 6721–6726. [Google Scholar]
- Morales-Lazaro, S.L.; Simon, S.A.; Rosenbaum, T. The role of endogenous molecules in modulating pain through transient receptor potential vanilloid 1 (TRPV1). J. Physiol. 2013, 591, 3109–3121. [Google Scholar]
- Vriens, J.; Appendino, G.; Nilius, B. Pharmacology of vanilloid transient receptor potential cation channels. Mol. Pharmacol. 2009, 75, 1262–1279. [Google Scholar] [CrossRef]
- Lishko, P.V.; Procko, E.; Jin, X.; Phelps, C.B.; Gaudet, R. The ankyrin repeats of TRPV1 bind multiple ligands and modulate channel sensitivity. Neuron 2007, 54, 905–918. [Google Scholar] [CrossRef]
- Premkumar, L.S.; Ahern, G.P. Induction of vanilloid receptor channel activity by protein kinase C. Nature 2000, 408, 985–990. [Google Scholar] [CrossRef]
- Tominaga, M.; Wada, M.; Masu, M. Potentiation of capsaicin receptor activity by metabotropic ATP receptors as a possible mechanism for ATP-evoked pain and hyperalgesia. Proc. Natl. Acad. Sci. USA 2001, 98, 6951–6956. [Google Scholar] [CrossRef]
- Julius, D.; Basbaum, A.I. Molecular mechanisms of nociception. Nature 2001, 413, 203–210. [Google Scholar] [CrossRef]
- Dhaka, A.; Uzzell, V.; Dubin, A.E.; Mathur, J.; Petrus, M.; Bandell, M.; Patapoutian, A. TRPV1 is activated by both acidic and basic pH. J. Neurosci. 2009, 29, 153–158. [Google Scholar] [CrossRef]
- Ahern, G.P.; Wang, X.; Miyares, R.L. Polyamines are potent ligands for the capsaicin receptor TRPV1. J. Biol. Chem. 2006, 281, 8991–8995. [Google Scholar] [CrossRef]
- LaMotte, R.H.; Campbell, J.N. Comparison of responses of warm and nociceptive C-fiber afferents in monkey with human judgments of thermal pain. J. Neurophysiol. 1978, 41, 509–528. [Google Scholar]
- Tillman, D.B.; Treede, R.D.; Meyer, R.A.; Campbell, J.N. Response of C fibre nociceptors in the anaesthetized monkey to heat stimuli: Estimates of receptor depth and threshold. J. Physiol. 1995, 485Pt 3, 753–765. [Google Scholar]
- Cesare, P.; McNaughton, P. A novel heat-activated current in nociceptive neurons and its sensitization by bradykinin. Proc. Natl. Acad. Sci. USA 1996, 93, 15435–15439. [Google Scholar] [CrossRef]
- Reichling, D.B.; Levine, J.D. Heat transduction in rat sensory neurons by calcium-dependent activation of a cation channel. Proc. Natl. Acad. Sci. USA 1997, 94, 7006–7011. [Google Scholar] [CrossRef]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef]
- Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with adelta/c-fibers and colocalization with trk receptors. J. Comp. Neurol. 2005, 493, 596–606. [Google Scholar] [CrossRef]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Basbaum, A.I.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Davey, P.T.; Overend, P.; Harries, M.H.; Latcham, J.; Clapham, C.; Atkinson, K.; et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef]
- Walder, R.Y.; Radhakrishnan, R.; Loo, L.; Rasmussen, L.A.; Mohapatra, D.P.; Wilson, S.P.; Sluka, K.A. TRPV1 is important for mechanical and heat sensitivity in uninjured animals and development of heat hypersensitivity after muscle inflammation. Pain 2012, 153, 1664–1672. [Google Scholar] [CrossRef]
- Sudbury, J.R.; Bourque, C.W. Dynamic and permissive roles of TRPV1 and TRPV4 channels for thermosensation in mouse supraoptic magnocellular neurosecretory neurons. J. Neurosci. 2013, 33, 17160–17165. [Google Scholar] [CrossRef]
- Mergler, S.; Garreis, F.; Sahlmuller, M.; Reinach, P.S.; Paulsen, F.; Pleyer, U. Thermosensitive transient receptor potential channels in human corneal epithelial cells. J. Cell Physiol. 2011, 226, 1828–1842. [Google Scholar] [CrossRef]
- Mizumura, K.; Kumazawa, T. Modification of nociceptor responses by inflammatory mediators and second messengers implicated in their action—A study in canine testicular polymodal receptors. Prog. Brain Res. 1996, 113, 115–141. [Google Scholar] [CrossRef]
- Wood, J.N.; Perl, E.R. Pain. Curr. Opin. Genet. Dev. 1999, 9, 328–332. [Google Scholar] [CrossRef]
- Woolf, C.J.; Salter, M.W. Neuronal plasticity: Increasing the gain in pain. Science 2000, 288, 1765–1769. [Google Scholar] [CrossRef]
- Amadesi, S.; Nie, J.; Vergnolle, N.; Cottrell, G.S.; Grady, E.F.; Trevisani, M.; Manni, C.; Geppetti, P.; McRoberts, J.A.; Ennes, H.; et al. Protease-activated receptor 2 sensitizes the capsaicin receptor transient receptor potential vanilloid receptor 1 to induce hyperalgesia. J. Neurosci. 2004, 24, 4300–4312. [Google Scholar] [CrossRef]
- Dai, Y.; Moriyama, T.; Higashi, T.; Togashi, K.; Kobayashi, K.; Yamanaka, H.; Tominaga, M.; Noguchi, K. Proteinase-activated receptor 2-mediated potentiation of transient receptor potential vanilloid subfamily 1 activity reveals a mechanism for proteinase-induced inflammatory pain. J. Neurosci. 2004, 24, 4293–4299. [Google Scholar] [CrossRef]
- Moriyama, T.; Higashi, T.; Togashi, K.; Iida, T.; Segi, E.; Sugimoto, Y.; Tominaga, T.; Narumiya, S.; Tominaga, M. Sensitization of TRPV1 by EP1 and IP reveals peripheral nociceptive mechanism of prostaglandins. Mol. Pain 2005, 1, 3. [Google Scholar] [CrossRef]
- Moriyama, T.; Iida, T.; Kobayashi, K.; Higashi, T.; Fukuoka, T.; Tsumura, H.; Leon, C.; Suzuki, N.; Inoue, K.; Gachet, C.; et al. Possible involvement of P2Y2 metabotropic receptors in ATP-induced transient receptor potential vanilloid receptor 1-mediated thermal hypersensitivity. J. Neurosci. 2003, 23, 6058–6062. [Google Scholar]
- Sugiura, T.; Tominaga, M.; Katsuya, H.; Mizumura, K. Bradykinin lowers the threshold temperature for heat activation of vanilloid receptor 1. J. Neurophysiol. 2002, 88, 544–548. [Google Scholar]
- Bhave, G.; Hu, H.J.; Glauner, K.S.; Zhu, W.; Wang, H.; Brasier, D.J.; Oxford, G.S.; Gereau, R.W.T. Protein kinase C phosphorylation sensitizes but does not activate the capsaicin receptor transient receptor potential vanilloid 1 (TRPV1). Proc. Natl. Acad. Sci. USA 2003, 100, 12480–12485. [Google Scholar]
- Numazaki, M.; Tominaga, T.; Toyooka, H.; Tominaga, M. Direct phosphorylation of capsaicin receptor VR1 by protein kinase Cepsilon and identification of two target serine residues. J. Biol. Chem. 2002, 277, 13375–13378. [Google Scholar] [CrossRef]
- Xing, B.M.; Yang, Y.R.; Du, J.X.; Chen, H.J.; Qi, C.; Huang, Z.H.; Zhang, Y.; Wang, Y. Cyclin-dependent kinase 5 controls TRPV1 membrane trafficking and the heat sensitivity of nociceptors through KIF13B. J. Neurosci. 2012, 32, 14709–14721. [Google Scholar] [CrossRef]
- Cheng, W.; Yang, F.; Liu, S.; Colton, C.K.; Wang, C.; Cui, Y.; Cao, X.; Zhu, M.X.; Sun, C.; Wang, K.; et al. Heteromeric heat-sensitive transient receptor potential channels exhibit distinct temperature and chemical response. J. Biol. Chem. 2012, 287, 7279–7288. [Google Scholar]
- Holzer, P. Acid-sensitive ion channels and receptors. Handb. Exp. Pharmacol. 2009, 194, 283–332. [Google Scholar] [CrossRef]
- Julius, D. TRP channels and pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef]
- Leffler, A.; Monter, B.; Koltzenburg, M. The role of the capsaicin receptor TRPV1 and acid-sensing ion channels (ASICS) in proton sensitivity of subpopulations of primary nociceptive neurons in rats and mice. Neuroscience 2006, 139, 699–709. [Google Scholar] [CrossRef]
- Sugiura, T.; Bielefeldt, K.; Gebhart, G.F. Mouse colon sensory neurons detect extracellular acidosis via TRPV1. Am. J. Physiol. Cell Physiol. 2007, 292, C1768–C1774. [Google Scholar]
- Yagi, J.; Wenk, H.N.; Naves, L.A.; McCleskey, E.W. Sustained currents through ASIC3 ion channels at the modest pH changes that occur during myocardial ischemia. Circ. Res. 2006, 99, 501–509. [Google Scholar] [CrossRef]
- Planells-Cases, R.; Garcia-Sanz, N.; Morenilla-Palao, C.; Ferrer-Montiel, A. Functional aspects and mechanisms of TRPV1 involvement in neurogenic inflammation that leads to thermal hyperalgesia. Pflugers Arch. 2005, 451, 151–159. [Google Scholar] [CrossRef]
- Hudson, L.J.; Bevan, S.; Wotherspoon, G.; Gentry, C.; Fox, A.; Winter, J. VR1 protein expression increases in undamaged DRG neurons after partial nerve injury. Eur. J. Neurosci. 2001, 13, 2105–2114. [Google Scholar] [CrossRef]
- Ji, R.R.; Samad, T.A.; Jin, S.X.; Schmoll, R.; Woolf, C.J. p38 MAPK activation by NGF in primary sensory neurons after inflammation increases TRPV1 levels and maintains heat hyperalgesia. Neuron 2002, 36, 57–68. [Google Scholar] [CrossRef]
- Tisch, R.; Yang, X.D.; Singer, S.M.; Liblau, R.S.; Fugger, L.; McDevitt, H.O. Immune response to glutamic acid decarboxylase correlates with insulitis in non-obese diabetic mice. Nature 1993, 366, 72–75. [Google Scholar] [CrossRef]
- Yoshida, K.; Kikutani, H. Genetic and immunological basis of autoimmune diabetes in the NOD mouse. Rev. Immunogenet. 2000, 2, 140–146. [Google Scholar]
- Winer, S.; Astsaturov, I.; Cheung, R.; Gunaratnam, L.; Kubiak, V.; Cortez, M.A.; Moscarello, M.; O’Connor, P.W.; McKerlie, C.; Becker, D.J.; et al. Type I diabetes and multiple sclerosis patients target islet plus central nervous system autoantigens; nonimmunized nonobese diabetic mice can develop autoimmune encephalitis. J. Immunol. 2001, 166, 2831–2841. [Google Scholar] [CrossRef]
- Winer, S.; Tsui, H.; Lau, A.; Song, A.; Li, X.; Cheung, R.K.; Sampson, A.; Afifiyan, F.; Elford, A.; Jackowski, G.; et al. Autoimmune islet destruction in spontaneous type 1 diabetes is not beta-cell exclusive. Nat. Med. 2003, 9, 198–205. [Google Scholar]
- Jancso, G.; Kiraly, E.; Jancso-Gabor, A. Pharmacologically induced selective degeneration of chemosensitive primary sensory neurones. Nature 1977, 270, 741–743. [Google Scholar] [CrossRef]
- Razavi, R.; Chan, Y.; Afifiyan, F.N.; Liu, X.J.; Wan, X.; Yantha, J.; Tsui, H.; Tang, L.; Tsai, S.; Santamaria, P.; et al. TRPV1+ sensory neurons control beta cell stress and islet inflammation in autoimmune diabetes. Cell 2006, 127, 1123–1135. [Google Scholar] [CrossRef]
- Basu, S.; Srivastava, P. Immunological role of neuronal receptor vanilloid receptor 1 expressed on dendritic cells. Proc. Natl. Acad. Sci. USA 2005, 102, 5120–5125. [Google Scholar] [CrossRef]
- Szollosi, A.G.; Olah, A.; Toth, I.B.; Papp, F.; Czifra, G.; Panyi, G.; Biro, T. Transient receptor potential vanilloid-2 mediates the effects of transient heat shock on endocytosis of human monocyte-derived dendritic cells. FEBS Lett. 2013, 587, 1440–1445. [Google Scholar] [CrossRef]
- Toth, B.I.; Benko, S.; Szollosi, A.G.; Kovacs, L.; Rajnavolgyi, E.; Biro, T. Transient receptor potential vanilloid-1 signaling inhibits differentiation and activation of human dendritic cells. FEBS Lett. 2009, 583, 1619–1624. [Google Scholar] [CrossRef]
- O’Connell, P.J.; Pingle, S.C.; Ahern, G.P. Dendritic cells do not transduce inflammatory stimuli via the capsaicin receptor TRPV1. FEBS Lett. 2005, 579, 5135–5139. [Google Scholar] [CrossRef]
- Akiba, Y.; Kato, S.; Katsube, K.; Nakamura, M.; Takeuchi, K.; Ishii, H.; Hibi, T. Transient receptor potential vanilloid subfamily 1 expressed in pancreatic islet beta cells modulates insulin secretion in rats. Biochem. Biophys. Res. Commun. 2004, 321, 219–225. [Google Scholar] [CrossRef]
- Van Buren, J.J.; Bhat, S.; Rotello, R.; Pauza, M.E.; Premkumar, L.S. Sensitization and translocation of TRPV1 by insulin and IGF-I. Mol. Pain 2005, 1, 17. [Google Scholar] [CrossRef]
- Hong, S.; Wiley, J.W. Early painful diabetic neuropathy is associated with differential changes in the expression and function of vanilloid receptor 1. J. Biol. Chem. 2005, 280, 618–627. [Google Scholar]
- Pabbidi, R.M.; Yu, S.Q.; Peng, S.; Khardori, R.; Pauza, M.E.; Premkumar, L.S. Influence of TRPV1 on diabetes-induced alterations in thermal pain sensitivity. Mol. Pain 2008, 4, 9. [Google Scholar] [CrossRef]
- Wilder-Smith, E.P.; Ong, W.Y.; Guo, Y.; Chow, A.W. Epidermal transient receptor potential vanilloid 1 in idiopathic small nerve fibre disease, diabetic neuropathy and healthy human subjects. Histopathology 2007, 51, 674–680. [Google Scholar] [CrossRef]
- Facer, P.; Casula, M.A.; Smith, G.D.; Benham, C.D.; Chessell, I.P.; Bountra, C.; Sinisi, M.; Birch, R.; Anand, P. Differential expression of the capsaicin receptor TRPV1 and related novel receptors TRPV3, TRPV4 and TRPM8 in normal human tissues and changes in traumatic and diabetic neuropathy. BMC Neurol. 2007, 7, 11. [Google Scholar] [CrossRef]
- Festa, A.; D’Agostino, R., Jr.; Howard, G.; Mykkanen, L.; Tracy, R.P.; Haffner, S.M. Chronic subclinical inflammation as part of the insulin resistance syndrome: The Insulin Resistance Atherosclerosis Study (IRAS). Circulation 2000, 102, 42–47. [Google Scholar] [CrossRef]
- Wu, T.; Dorn, J.P.; Donahue, R.P.; Sempos, C.T.; Trevisan, M. Associations of serum C-reactive protein with fasting insulin, glucose, and glycosylated hemoglobin: The Third National Health and Nutrition Examination Survey, 1988–1994. Am. J. Epidemiol. 2002, 155, 65–71. [Google Scholar] [CrossRef]
- Tajiri, Y.; Takei, R.; Mimura, K.; Umeda, F. Indicators for the efficacy of pioglitazone before and during treatment in Japanese patients with type 2 diabetes. Diabetes Technol. Ther. 2007, 9, 429–437. [Google Scholar] [CrossRef]
- Gram, D.X.; Hansen, A.J.; Wilken, M.; Elm, T.; Svendsen, O.; Carr, R.D.; Ahren, B.; Brand, C.L. Plasma calcitonin gene-related peptide is increased prior to obesity, and sensory nerve desensitization by capsaicin improves oral glucose tolerance in obese Zucker rats. Eur. J. Endocrinol. 2005, 153, 963–969. [Google Scholar] [CrossRef]
- Melnyk, A.; Himms-Hagen, J. Resistance to aging-associated obesity in capsaicin-desensitized rats one year after treatment. Obes. Res. 1995, 3, 337–344. [Google Scholar] [CrossRef]
- Pettersson, M.; Ahren, B.; Bottcher, G.; Sundler, F. Calcitonin gene-related peptide: Occurrence in pancreatic islets in the mouse and the rat and inhibition of insulin secretion in the mouse. Endocrinology 1986, 119, 865–869. [Google Scholar] [CrossRef]
- Gram, D.X.; Ahren, B.; Nagy, I.; Olsen, U.B.; Brand, C.L.; Sundler, F.; Tabanera, R.; Svendsen, O.; Carr, R.D.; Santha, P.; et al. Capsaicin-sensitive sensory fibers in the islets of Langerhans contribute to defective insulin secretion in Zucker diabetic rat, an animal model for some aspects of human type 2 diabetes. Eur. J. Neurosci. 2007, 25, 213–223. [Google Scholar] [CrossRef]
- Westerterp-Plantenga, M.S.; Smeets, A.; Lejeune, M.P. Sensory and gastrointestinal satiety effects of capsaicin on food intake. Int. J. Obes. 2005, 29, 682–688. [Google Scholar] [CrossRef]
- Belza, A.; Frandsen, E.; Kondrup, J. Body fat loss achieved by stimulation of thermogenesis by a combination of bioactive food ingredients: A placebo-controlled, double-blind 8-week intervention in obese subjects. Int. J. Obes. 2007, 31, 121–130. [Google Scholar] [CrossRef]
- Rybak, L.P.; Ramkumar, V. Ototoxicity. Kidney Int. 2007, 72, 931–935. [Google Scholar] [CrossRef]
- Rybak, L.P.; Whitworth, C.; Somani, S. Application of antioxidants and other agents to prevent cisplatin ototoxicity. Laryngoscope 1999, 109, 1740–1744. [Google Scholar] [CrossRef]
- Kopke, R.D.; Liu, W.; Gabaizadeh, R.; Jacono, A.; Feghali, J.; Spray, D.; Garcia, P.; Steinman, H.; Malgrange, B.; Ruben, R.J.; et al. Use of organotypic cultures of Corti’s organ to study the protective effects of antioxidant molecules on cisplatin-induced damage of auditory hair cells. Am. J. Otol. 1997, 18, 559–571. [Google Scholar]
- Zheng, J.; Dai, C.; Steyger, P.S.; Kim, Y.; Vass, Z.; Ren, T.; Nuttall, A.L. Vanilloid receptors in hearing: Altered cochlear sensitivity by vanilloids and expression of TRPV1 in the organ of corti. J. Neurophysiol. 2003, 90, 444–455. [Google Scholar] [CrossRef]
- Vass, Z.; Dai, C.F.; Steyger, P.S.; Jancso, G.; Trune, D.R.; Nuttall, A.L. Co-localization of the vanilloid capsaicin receptor and substance P in sensory nerve fibers innervating cochlear and vertebro-basilar arteries. Neuroscience 2004, 124, 919–927. [Google Scholar] [CrossRef]
- Wu, T.; Song, L.; Shi, X.; Jiang, Z.; Santos-Sacchi, J.; Nuttall, A.L. Effect of capsaicin on potassium conductance and electromotility of the guinea pig outer hair cell. Hear. Res. 2011, 272, 117–124. [Google Scholar] [CrossRef]
- Zhou, J.; Balaban, C.; Durrant, J.D. Effect of intracochlear perfusion of vanilloids on cochlear neural activity in the guinea pig. Hear. Res. 2006, 218, 43–49. [Google Scholar]
- Meyers, J.R.; MacDonald, R.B.; Duggan, A.; Lenzi, D.; Standaert, D.G.; Corwin, J.T.; Corey, D.P. Lighting up the senses: FM1–43 loading of sensory cells through nonselective ion channels. J. Neurosci. 2003, 23, 4054–4065. [Google Scholar]
- Myrdal, S.E.; Steyger, P.S. TRPV1 regulators mediate gentamicin penetration of cultured kidney cells. Hear. Res. 2005, 204, 170–182. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, C.; Kim, S.J.; Kim, H.J.; Oh, G.S.; Shen, A.; So, H.S.; Park, R. Different uptake of gentamicin through TRPV1 and TRPV4 channels determines cochlear hair cell vulnerability. Exp. Mol. Med. 2013, 45, e12. [Google Scholar] [CrossRef]
- Mukherjea, D.; Jajoo, S.; Whitworth, C.; Bunch, J.R.; Turner, J.G.; Rybak, L.P.; Ramkumar, V. Short interfering RNA against transient receptor potential vanilloid 1 attenuates cisplatin-induced hearing loss in the rat. J. Neurosci. 2008, 28, 13056–13065. [Google Scholar]
- Mukherjea, D.; Jajoo, S.; Sheehan, K.; Kaur, T.; Sheth, S.; Bunch, J.; Perro, C.; Rybak, L.P.; Ramkumar, V. NOX3 NADPH oxidase couples transient receptor potential vanilloid 1 to signal transducer and activator of transcription 1-mediated inflammation and hearing loss. Antioxid Redox Signal. 2011, 14, 999–1010. [Google Scholar] [CrossRef]
- Kitahara, T.; Li, H.S.; Balaban, C.D. Changes in transient receptor potential cation channel superfamily V (TRPV) mRNA expression in the mouse inner ear ganglia after kanamycin challenge. Hear. Res. 2005, 201, 132–144. [Google Scholar] [CrossRef]
- Ishibashi, T.; Takumida, M.; Akagi, N.; Hirakawa, K.; Anniko, M. Changes in transient receptor potential vanilloid (TRPV) 1, 2, 3 and 4 expression in mouse inner ear following gentamicin challenge. Acta Oto-Laryngol. 2009, 129, 116–126. [Google Scholar] [CrossRef]
- Kizawa, K.; Kitahara, T.; Horii, A.; Maekawa, C.; Kuramasu, T.; Kawashima, T.; Nishiike, S.; Doi, K.; Inohara, H. Behavioral assessment and identification of a molecular marker in a salicylate-induced tinnitus in rats. Neuroscience 2010, 165, 1323–1332. [Google Scholar] [CrossRef]
- Bauer, C.A.; Brozoski, T.J.; Myers, K.S. Acoustic injury and TRPV1 expression in the cochlear spiral ganglion. Int. Tinnitus J. 2007, 13, 21–28. [Google Scholar]
- Kaur, T.; Mukherjea, D.; Sheehan, K.; Jajoo, S.; Rybak, L.P.; Ramkumar, V. Short interfering RNA against STAT1 attenuates cisplatin-induced ototoxicity in the rat by suppressing inflammation. Cell Death. Dis. 2011, 2, e180. [Google Scholar] [CrossRef]
- Lecci, A.; Maggi, C.A. Tachykinins as modulators of the micturition reflex in the central and peripheral nervous system. Regul. Pept. 2001, 101, 1–18. [Google Scholar] [CrossRef]
- Yiangou, Y.; Facer, P.; Ford, A.; Brady, C.; Wiseman, O.; Fowler, C.J.; Anand, P. Capsaicin receptor VR1 and ATP-gated ion channel P2X3 in human urinary bladder. BJU Int. 2001, 87, 774–779. [Google Scholar]
- Maggi, C.A.; Barbanti, G.; Santicioli, P.; Beneforti, P.; Misuri, D.; Meli, A.; Turini, D. Cystometric evidence that capsaicin-sensitive nerves modulate the afferent branch of micturition reflex in humans. J. Urol. 1989, 142, 150–154. [Google Scholar]
- Maggi, C.A. The dual function of capsaicin-sensitive sensory nerves in the bladder and urethra. Ciba. Found. Symp. 1990, 151, 77–90, 77–83; discussion 83–90. [Google Scholar]
- Szallasi, A.; Conte, B.; Goso, C.; Blumberg, P.M.; Manzini, S. Characterization of a peripheral vanilloid (capsaicin) receptor in the urinary bladder of the rat. Life Sci. 1993, 52, PL221–PL226. [Google Scholar]
- Avelino, A.; Cruz, C.; Nagy, I.; Cruz, F. Vanilloid receptor 1 expression in the rat urinary tract. Neuroscience 2002, 109, 787–798. [Google Scholar] [CrossRef]
- Birder, L.A.; Kanai, A.J.; de Groat, W.C.; Kiss, S.; Nealen, M.L.; Burke, N.E.; Dineley, K.E.; Watkins, S.; Reynolds, I.J.; Caterina, M.J. Vanilloid receptor expression suggests a sensory role for urinary bladder epithelial cells. Proc. Natl. Acad. Sci. USA 2001, 98, 13396–13401. [Google Scholar] [CrossRef]
- Apostolidis, A.; Brady, C.M.; Yiangou, Y.; Davis, J.; Fowler, C.J.; Anand, P. Capsaicin receptor TRPV1 in urothelium of neurogenic human bladders and effect of intravesical resiniferatoxin. Urology 2005, 65, 400–405. [Google Scholar] [CrossRef]
- Kobayashi, H.; Yoshiyama, M.; Zakoji, H.; Takeda, M.; Araki, I. Sex differences in the expression profile of acid-sensing ion channels in the mouse urinary bladder: A possible involvement in irritative bladder symptoms. BJU Int. 2009, 104, 1746–1751. [Google Scholar] [CrossRef]
- Lazzeri, M.; Vannucchi, M.G.; Spinelli, M.; Bizzoco, E.; Beneforti, P.; Turini, D.; Faussone-Pellegrini, M.S. Transient receptor potential vanilloid type 1 (TRPV1) expression changes from normal urothelium to transitional cell carcinoma of human bladder. Eur. Urol. 2005, 48, 691–698. [Google Scholar] [CrossRef]
- Shabir, S.; Cross, W.; Kirkwood, L.A.; Pearson, J.F.; Appleby, P.A.; Walker, D.; Eardley, I.; Southgate, J. Functional expression of purinergic P2 receptors and transient receptor potential channels by the human urothelium. Am. J. Physiol. Renal Physiol. 2013, 305, F396–F406. [Google Scholar] [CrossRef]
- Yu, W.; Hill, W.G.; Apodaca, G.; Zeidel, M.L. Expression and distribution of transient receptor potential (TRP) channels in bladder epithelium. Am. J. Physiol. Renal Physiol. 2011, 300, F49–F59. [Google Scholar] [CrossRef]
- Ost, D.; Roskams, T.; van der Aa, F.; de Ridder, D. Topography of the vanilloid receptor in the human bladder: More than just the nerve fibers. J. Urol. 2002, 168, 293–297. [Google Scholar] [CrossRef]
- Sui, G.P.; Wu, C.; Fry, C.H. Electrical characteristics of suburothelial cells isolated from the human bladder. J. Urol. 2004, 171, 938–943. [Google Scholar] [CrossRef]
- Everaerts, W.; Vriens, J.; Owsianik, G.; Appendino, G.; Voets, T.; de Ridder, D.; Nilius, B. Functional characterization of transient receptor potential channels in mouse urothelial cells. Am. J. Physiol. Renal Physiol. 2010, 298, F692–F701. [Google Scholar] [CrossRef]
- Yamada, T.; Ugawa, S.; Ueda, T.; Ishida, Y.; Kajita, K.; Shimada, S. Differential localizations of the transient receptor potential channels TRPV4 and TRPV1 in the mouse urinary bladder. J. Histochem. Cytochem. 2009, 57, 277–287. [Google Scholar]
- Daly, D.; Rong, W.; Chess-Williams, R.; Chapple, C.; Grundy, D. Bladder afferent sensitivity in wild-type and TRPV1 knockout mice. J. Physiol. 2007, 583, 663–674. [Google Scholar] [CrossRef]
- Cefalu, J.S.; Guillon, M.A.; Burbach, L.R.; Zhu, Q.M.; Hu, D.Q.; Ho, M.J.; Ford, A.P.; Nunn, P.A.; Cockayne, D.A. Selective pharmacological blockade of the TRPV1 receptor suppresses sensory reflexes of the rodent bladder. J. Urol. 2009, 182, 776–785. [Google Scholar] [CrossRef]
- Cheng, C.L.; Liu, J.C.; Chang, S.Y.; Ma, C.P.; de Groat, W.C. Effect of capsaicin on the micturition reflex in normal and chronic spinal cord-injured cats. Am. J. Physiol. 1999, 277, R786–R794. [Google Scholar]
- Cheng, C.L.; Ma, C.P.; de Groat, W.C. Effect of capsaicin on micturition and associated reflexes in chronic spinal rats. Brain Res. 1995, 678, 40–48. [Google Scholar] [CrossRef]
- Santos-Silva, A.; Charrua, A.; Cruz, C.D.; Gharat, L.; Avelino, A.; Cruz, F. Rat detrusor overactivity induced by chronic spinalization can be abolished by a transient receptor potential vanilloid 1 (TRPV1) antagonist. Autono. Neurosci. 2012, 166, 35–38. [Google Scholar] [CrossRef]
- Chancellor, M.B.; de Groat, W.C. Intravesical capsaicin and resiniferatoxin therapy: Spicing up the ways to treat the overactive bladder. J. Urol. 1999, 162, 3–11. [Google Scholar] [CrossRef]
- Birder, L.A.; Wolf-Johnston, A.S.; Sun, Y.; Chai, T.C. Alteration in TRPV1 and Muscarinic (M3) receptor expression and function in idiopathic overactive bladder urothelial cells. Acta Physiol. (Oxf.) 2013, 207, 123–129. [Google Scholar] [CrossRef]
- Dornelles, F.N.; Andrade, E.L.; Campos, M.M.; Calixto, J.B. Role of CXCR2 and TRPV1 in functional, inflammatory and behavioural changes in the rat model of cyclophosphamide-induced haemorrhagic cystitis. Br. J. Pharmacol. 2014, 171, 452–467. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Wang, P.; Merriam, F.V.; Bjorling, D.E. Lack of TRPV1 inhibits cystitis-induced increased mechanical sensitivity in mice. Pain 2008, 139, 158–167. [Google Scholar] [CrossRef]
- Dang, K.; Bielefeldt, K.; Gebhart, G.F. Cyclophosphamide-induced cystitis reduces ASIC channel but enhances TRPV1 receptor function in rat bladder sensory neurons. J. Neurophysiol. 2013, 110, 408–417. [Google Scholar] [CrossRef]
- Homma, Y.; Nomiya, A.; Tagaya, M.; Oyama, T.; Takagaki, K.; Nishimatsu, H.; Igawa, Y. Increased mRNA expression of genes involved in pronociceptive inflammatory reactions in bladder tissue of interstitial cystitis. J. Urol. 2013, 190, 1925–1931. [Google Scholar] [CrossRef]
- Watanabe, N.; Horie, S.; Michael, G.J.; Keir, S.; Spina, D.; Page, C.P.; Priestley, J.V. Immunohistochemical co-localization of transient receptor potential vanilloid (TRPV)1 and sensory neuropeptides in the guinea-pig respiratory system. Neuroscience 2006, 141, 1533–1543. [Google Scholar] [CrossRef]
- Jang, Y.; Lee, Y.; Kim, S.M.; Yang, Y.D.; Jung, J.; Oh, U. Quantitative analysis of TRP channel genes in mouse organs. Arch. Pharm. Res. 2012, 35, 1823–1830. [Google Scholar] [CrossRef]
- Kunert-Keil, C.; Bisping, F.; Kruger, J.; Brinkmeier, H. Tissue-specific expression of TRP channel genes in the mouse and its variation in three different mouse strains. BMC Genomics 2006, 7, 159. [Google Scholar] [CrossRef]
- Baxter, L.A.; Birrell, M.A.; Belvisi, M.G. The role of TRPV1 in tobacco smoke induced airway inflammatiom. Am. J. Respir. Crit. Care Med. 2012, 185, A6410. [Google Scholar]
- Smit, L.A.; Kogevinas, M.; Anto, J.M.; Bouzigon, E.; Gonzalez, J.R.; le Moual, N.; Kromhout, H.; Carsin, A.E.; Pin, I.; Jarvis, D.; et al. Transient receptor potential genes, smoking, occupational exposures and cough in adults. Respir. Res. 2012, 13, 26. [Google Scholar] [CrossRef]
- Cantero-Recasens, G.; Gonzalez, J.R.; Fandos, C.; Duran-Tauleria, E.; Smit, L.A.; Kauffmann, F.; Anto, J.M.; Valverde, M.A. Loss of function of transient receptor potential vanilloid 1 (TRPV1) genetic variant is associated with lower risk of active childhood asthma. J. Biol. Chem. 2010, 285, 27532–27535. [Google Scholar] [CrossRef]
- McLeod, R.L.; Fernandez, X.; Correll, C.C.; Phelps, T.P.; Jia, Y.; Wang, X.; Hey, J.A. TRPV1 antagonists attenuate antigen-provoked cough in ovalbumin sensitized guinea pigs. Cough 2006, 2, 10. [Google Scholar] [CrossRef]
- Trevisani, M.; Milan, A.; Gatti, R.; Zanasi, A.; Harrison, S.; Fontana, G.; Morice, A.H.; Geppetti, P. Antitussive activity of iodo-resiniferatoxin in guinea pigs. Thorax 2004, 59, 769–772. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Scott, B.P.; Nasser, N.; Ao, H.; Maher, M.P.; Dubin, A.E.; Swanson, D.M.; Shankley, N.P.; Wickenden, A.D.; Chaplan, S.R. Pharmacology and antitussive efficacy of 4-(3-trifluoromethyl-pyridin-2-yl)-piperazine-1-carboxylic acid (5-trifluoromethyl-pyridin-2-yl)-amide (JNJ17203212), a transient receptor potential vanilloid 1 antagonist in guinea pigs. J. Pharmacol. Exp. Ther. 2007, 323, 665–674. [Google Scholar] [CrossRef]
- Lalloo, U.G.; Fox, A.J.; Belvisi, M.G.; Chung, K.F.; Barnes, P.J. Capsazepine inhibits cough induced by capsaicin and citric acid but not by hypertonic saline in guinea pigs. J. Appl. Physiol. 1995, 79, 1082–1087. [Google Scholar]
- Doherty, M.J.; Mister, R.; Pearson, M.G.; Calverley, P.M. Capsaicin responsiveness and cough in asthma and chronic obstructive pulmonary disease. Thorax 2000, 55, 643–649. [Google Scholar]
- Nakajima, T.; Nishimura, Y.; Nishiuma, T.; Kotani, Y.; Nakata, H.; Yokoyama, M. Cough sensitivity in pure cough variant asthma elicited using continuous capsaicin inhalation. Allergol. Int. 2006, 55, 149–155. [Google Scholar] [CrossRef]
- O’Connell, F.; Thomas, V.E.; Studham, J.M.; Pride, N.B.; Fuller, R.W. Capsaicin cough sensitivity increases during upper respiratory infection. Respir. Med. 1996, 90, 279–286. [Google Scholar] [CrossRef]
- Pecova, R.; Zucha, J.; Pec, M.; Neuschlova, M.; Hanzel, P.; Tatar, M. Cough reflex sensitivity testing in in seasonal allergic rhinitis patients and healthy volunteers. J. Physiol. Pharmacol. 2008, 59 (Suppl. 6), 557–564. [Google Scholar]
- Plevkova, J.; Varechova, S.; Brozmanova, M.; Tatar, M. Testing of cough reflex sensitivity in children suffering from allergic rhinitis and common cold. J. Physiol. Pharmacol. 2006, 57 (Suppl. 4), 289–296. [Google Scholar]
- Groneberg, D.A.; Niimi, A.; Dinh, Q.T.; Cosio, B.; Hew, M.; Fischer, A.; Chung, K.F. Increased expression of transient receptor potential vanilloid-1 in airway nerves of chronic cough. Am. J. Respir. Crit. Care Med. 2004, 170, 1276–1280. [Google Scholar] [CrossRef]
- Mitchell, J.E.; Campbell, A.P.; New, N.E.; Sadofsky, L.R.; Kastelik, J.A.; Mulrennan, S.A.; Compton, S.J.; Morice, A.H. Expression and characterization of the intracellular vanilloid receptor (TRPV1) in bronchi from patients with chronic cough. Exp. Lung Res. 2005, 31, 295–306. [Google Scholar]
- Carr, M.J.; Hunter, D.D.; Jacoby, D.B.; Undem, B.J. Expression of tachykinins in nonnociceptive vagal afferent neurons during respiratory viral infection in guinea pigs. Am. J. Respir. Crit. Care Med. 2002, 165, 1071–1075. [Google Scholar] [CrossRef]
- Lewis, C.A.; Ambrose, C.; Banner, K.; Battram, C.; Butler, K.; Giddings, J.; Mok, J.; Nasra, J.; Winny, C.; Poll, C. Animal models of cough: Literature review and presentation of a novel cigarette smoke-enhanced cough model in the guinea-pig. Pulm. Pharmacol. Ther. 2007, 20, 325–333. [Google Scholar] [CrossRef]
- Lieu, T.M.; Myers, A.C.; Meeker, S.; Undem, B.J. TRPV1 induction in airway vagal low-threshold mechanosensory neurons by allergen challenge and neurotrophic factors. Am. J. Physiol. Lung. Cell Mol. Physiol. 2012, 302, L941–L948. [Google Scholar] [CrossRef]
- Myers, A.C.; Kajekar, R.; Undem, B.J. Allergic inflammation-induced neuropeptide production in rapidly adapting afferent nerves in guinea pig airways. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 282, L775–L781. [Google Scholar] [CrossRef]
- Ye, X.M.; Zhong, N.S.; Liu, C.L.; Chen, R.C. Cough reflex sensitivity is increased in guinea pigs with parainfluenza virus infection. Exp. Lung Res. 2011, 37, 186–194. [Google Scholar] [CrossRef]
- Zhang, G.; Lin, R.L.; Wiggers, M.; Snow, D.M.; Lee, L.Y. Altered expression of TRPV1 and sensitivity to capsaicin in pulmonary myelinated afferents following chronic airway inflammation in the rat. J. Physiol. 2008, 586, 5771–5786. [Google Scholar] [CrossRef]
- Chong, M.S.; Hester, J. Diabetic painful neuropathy: Current and future treatment options. Drugs 2007, 67, 569–585. [Google Scholar] [CrossRef]
- Jensen, P.G.; Larson, J.R. Management of painful diabetic neuropathy. Drugs Aging 2001, 18, 737–749. [Google Scholar] [CrossRef]
- Zin, C.S.; Nissen, L.M.; Smith, M.T.; O’Callaghan, J.P.; Moore, B.J. An update on the pharmacological management of post-herpetic neuralgia and painful diabetic neuropathy. CNS Drugs 2008, 22, 417–442. [Google Scholar] [CrossRef]
- Galluzzi, K.E. Management strategies for herpes zoster and postherpetic neuralgia. J. Am. Osteopath. Assoc. 2007, 107, S8–S13. [Google Scholar]
- Broad, L.M.; Keding, S.J.; Blanco, M.J. Recent progress in the development of selective TRPV1 antagonists for pain. Curr. Top. Med. Chem. 2008, 8, 1431–1441. [Google Scholar] [CrossRef]
- Pal, M.; Angaru, S.; Kodimuthali, A.; Dhingra, N. Vanilloid receptor antagonists: Emerging class of novel anti-inflammatory agents for pain management. Curr. Pharm. Des. 2009, 15, 1008–1026. [Google Scholar] [CrossRef]
- Wong, G.Y.; Gavva, N.R. Therapeutic potential of vanilloid receptor TRPV1 agonists and antagonists as analgesics: Recent advances and setbacks. Brain Res. Rev. 2009, 60, 267–277. [Google Scholar] [CrossRef]
- Walpole, C.S.; Bevan, S.; Bovermann, G.; Boelsterli, J.J.; Breckenridge, R.; Davies, J.W.; Hughes, G.A.; James, I.; Oberer, L.; Winter, J.; et al. The discovery of capsazepine, the first competitive antagonist of the sensory neuron excitants capsaicin and resiniferatoxin. J. Med. Chem. 1994, 37, 1942–1954. [Google Scholar] [CrossRef]
- Bevan, S.; Hothi, S.; Hughes, G.; James, I.F.; Rang, H.P.; Shah, K.; Walpole, C.S.; Yeats, J.C. Capsazepine: A competitive antagonist of the sensory neurone excitant capsaicin. Br. J. Pharmacol. 1992, 107, 544–552. [Google Scholar] [CrossRef]
- Liu, L.; Simon, S.A. Capsazepine, a vanilloid receptor antagonist, inhibits nicotinic acetylcholine receptors in rat trigeminal ganglia. Neurosci. Lett. 1997, 228, 29–32. [Google Scholar] [CrossRef]
- Docherty, R.J.; Yeats, J.C.; Piper, A.S. Capsazepine block of voltage-activated calcium channels in adult rat dorsal root ganglion neurones in culture. Br. J. Pharmacol. 1997, 121, 1461–1467. [Google Scholar] [CrossRef]
- Weil, A.; Moore, S.E.; Waite, N.J.; Randall, A.; Gunthorpe, M.J. Conservation of functional and pharmacological properties in the distantly related temperature sensors TRVP1 and TRPM8. Mol. Pharmacol. 2005, 68, 518–527. [Google Scholar]
- Walker, K.M.; Urban, L.; Medhurst, S.J.; Patel, S.; Panesar, M.; Fox, A.J.; McIntyre, P. The VR1 antagonist capsazepine reverses mechanical hyperalgesia in models of inflammatory and neuropathic pain. J. Pharmacol. Exp. Ther. 2003, 304, 56–62. [Google Scholar] [CrossRef]
- Chung, J.U.; Kim, S.Y.; Lim, J.O.; Choi, H.K.; Kang, S.U.; Yoon, H.S.; Ryu, H.; Kang, D.W.; Lee, J.; Kang, B.; et al. Alpha-substituted N-(4-tert-butylbenzyl)-N'-[4-(methylsulfonylamino) benzyl]thiourea analogues as potent and stereospecific TRPV1 antagonists. Bioorg. Med. Chem. 2007, 15, 6043–6053. [Google Scholar] [CrossRef]
- Suh, Y.G.; Lee, Y.S.; Min, K.H.; Park, O.H.; Kim, J.K.; Seung, H.S.; Seo, S.Y.; Lee, B.Y.; Nam, Y.H.; Lee, K.O.; et al. Novel potent antagonists of transient receptor potential channel, vanilloid subfamily member 1: Structure-activity relationship of 1,3-diarylalkyl thioureas possessing new vanilloid equivalents. J. Med. Chem. 2005, 48, 5823–5836. [Google Scholar] [CrossRef]
- Kym, P.R.; Kort, M.E.; Hutchins, C.W. Analgesic potential of TRPV1 antagonists. Biochem. Pharmacol. 2009, 78, 211–216. [Google Scholar] [CrossRef]
- Szallasi, A.; Cortright, D.N.; Blum, C.A.; Eid, S.R. The vanilloid receptor TRPV1: 10 years from channel cloning to antagonist proof-of-concept. Nat. Rev. Drug Discov. 2007, 6, 357–372. [Google Scholar]
- Wang, Y.; Szabo, T.; Welter, J.D.; Toth, A.; Tran, R.; Lee, J.; Kang, S.U.; Suh, Y.G.; Blumberg, P.M. High affinity antagonists of the vanilloid receptor. Mol. Pharmacol. 2002, 62, 947–956. [Google Scholar] [CrossRef]
- El Kouhen, R.; Surowy, C.S.; Bianchi, B.R.; Neelands, T.R.; McDonald, H.A.; Niforatos, W.; Gomtsyan, A.; Lee, C.H.; Honore, P.; Sullivan, J.P.; et al. A-425619 [1-isoquinolin-5-yl-3-(4-trifluoromethyl-benzyl)-urea], a novel and selective transient receptor potential type V1 receptor antagonist, blocks channel activation by vanilloids, heat, and acid. J. Pharmacol. Exp. Ther. 2005, 314, 400–409. [Google Scholar]
- Valenzano, K.J.; Grant, E.R.; Wu, G.; Hachicha, M.; Schmid, L.; Tafesse, L.; Sun, Q.; Rotshteyn, Y.; Francis, J.; Limberis, J.; et al. N-(4-tertiarybutylphenyl)-4-(3-chloropyridin-2-yl) tetrahydropyrazine -1(2H)-carbox-amide (BCTC), a novel, orally effective vanilloid receptor 1 antagonist with analgesic properties: I. In vitro characterization and pharmacokinetic properties. J. Pharmacol. Exp. Ther. 2003, 306, 377–386. [Google Scholar] [CrossRef]
- Swanson, D.M.; Dubin, A.E.; Shah, C.; Nasser, N.; Chang, L.; Dax, S.L.; Jetter, M.; Breitenbucher, J.G.; Liu, C.; Mazur, C.; et al. Identification and biological evaluation of 4-(3-trifluoromethylpyridin-2-yl)piperazine-1-carboxylic acid (5-trifluoromethylpyridin-2-yl)amide, a high affinity TRPV1 (VR1) vanilloid receptor antagonist. J. Med. Chem. 2005, 48, 1857–1872. [Google Scholar] [CrossRef]
- Rami, H.K.; Thompson, M.; Stemp, G.; Fell, S.; Jerman, J.C.; Stevens, A.J.; Smart, D.; Sargent, B.; Sanderson, D.; Randall, A.D.; et al. Discovery of SB-705498: A potent, selective and orally bioavailable TRPV1 antagonist suitable for clinical development. Bioorg. Med. Chem. Lett. 2006, 16, 3287–3291. [Google Scholar] [CrossRef]
- Gunthorpe, M.J.; Rami, H.K.; Jerman, J.C.; Smart, D.; Gill, C.H.; Soffin, E.M.; Luis Hannan, S.; Lappin, S.C.; Egerton, J.; Smith, G.D.; et al. Identification and characterisation of SB-366791, a potent and selective vanilloid receptor (VR1/TRPV1) antagonist. Neuropharmacology 2004, 46, 133–149. [Google Scholar] [CrossRef]
- Gavva, N.R.; Tamir, R.; Qu, Y.; Klionsky, L.; Zhang, T.J.; Immke, D.; Wang, J.; Zhu, D.; Vanderah, T.W.; Porreca, F.; et al. AMG 9810 [(E)-3-(4-t-butylphenyl)-N-(2,3-dihydrobenzo [b][1,4] dioxin-6-yl)acrylamide], a novel vanilloid receptor 1 (TRPV1) antagonist with antihyperalgesic properties. J. Pharmacol. Exp. Ther. 2005, 313, 474–484. [Google Scholar]
- Zheng, X.; Hodgetts, K.J.; Brielmann, H.; Hutchison, A.; Burkamp, F.; Brian Jones, A.; Blurton, P.; Clarkson, R.; Chandrasekhar, J.; Bakthavatchalam, R.; et al. From arylureas to biarylamides to aminoquinazolines: Discovery of a novel, potent TRPV1 antagonist. Bioorg. Med. Chem. Lett. 2006, 16, 5217–5221. [Google Scholar] [CrossRef]
- Ognyanov, V.I.; Balan, C.; Bannon, A.W.; Bo, Y.; Dominguez, C.; Fotsch, C.; Gore, V.K.; Klionsky, L.; Ma, V.V.; Qian, Y.X.; et al. Design of potent, orally available antagonists of the transient receptor potential vanilloid 1. Structure-activity relationships of 2-piperazin-1-yl-1H-benzimidazoles. J. Med. Chem. 2006, 49, 3719–3742. [Google Scholar] [CrossRef]
- Messeguer, A.; Planells-Cases, R.; Ferrer-Montiel, A. Physiology and pharmacology of the vanilloid receptor. Curr. Neuropharmacol. 2006, 4, 1–15. [Google Scholar]
- Garcia-Martinez, C.; Morenilla-Palao, C.; Planells-Cases, R.; Merino, J.M.; Ferrer-Montiel, A. Identification of an aspartic residue in the P-loop of the vanilloid receptor that modulates pore properties. J. Biol. Chem. 2000, 275, 32552–32558. [Google Scholar]
- Himmel, H.M.; Kiss, T.; Borvendeg, S.J.; Gillen, C.; Illes, P. The arginine-rich hexapeptide R4W2 is a stereoselective antagonist at the vanilloid receptor 1: A Ca2+ imaging study in adult rat dorsal root ganglion neurons. J. Pharmacol. Exp. Ther. 2002, 301, 981–986. [Google Scholar] [CrossRef]
- Mellor, I.R.; Ogilvie, J.; Pluteanu, F.; Clothier, R.H.; Parker, T.L.; Rosini, M.; Minarini, A.; Tumiatti, V.; Melchiorre, C. Methoctramine analogues inhibit responses to capsaicin and protons in rat dorsal root ganglion neurons. Eur. J. Pharmacol. 2004, 505, 37–50. [Google Scholar] [CrossRef]
- Kitaguchi, T.; Swartz, K.J. An inhibitor of TRPV1 channels isolated from funnel Web spider venom. Biochemistry 2005, 44, 15544–15549. [Google Scholar] [CrossRef]
- Garcia-Martinez, C.; Humet, M.; Planells-Cases, R.; Gomis, A.; Caprini, M.; Viana, F.; de la Pena, E.; Sanchez-Baeza, F.; Carbonell, T.; de Felipe, C.; et al. Attenuation of thermal nociception and hyperalgesia by VR1 blockers. Proc. Natl. Acad. Sci. USA 2002, 99, 2374–2379. [Google Scholar]
- McIntyre, P.; McLatchie, L.M.; Chambers, A.; Phillips, E.; Clarke, M.; Savidge, J.; Toms, C.; Peacock, M.; Shah, K.; Winter, J.; et al. Pharmacological differences between the human and rat vanilloid receptor 1 (VR1). Br. J. Pharmacol. 2001, 132, 1084–1094. [Google Scholar] [CrossRef]
- Honore, P.; Wismer, C.T.; Mikusa, J.; Zhu, C.Z.; Zhong, C.; Gauvin, D.M.; Gomtsyan, A.; el Kouhen, R.; Lee, C.H.; Marsh, K.; et al. A-425619 [1-isoquinolin-5-yl-3-(4-trifluoromethyl-benzyl)-urea], a novel transient receptor potential type V1 receptor antagonist, relieves pathophysiological pain associated with inflammation and tissue injury in rats. J. Pharmacol. Exp. Ther. 2005, 314, 410–421. [Google Scholar] [CrossRef]
- Pomonis, J.D.; Harrison, J.E.; Mark, L.; Bristol, D.R.; Valenzano, K.J.; Walker, K. N-(4-Tertiarybutylphenyl)-4-(3-cholorphyridin-2-yl)tetrahydropyrazine-1(2H)-carbox-amide (BCTC), a novel, orally effective vanilloid receptor 1 antagonist with analgesic properties: I. in vivo characterization in rat models of inflammatory and neuropathic pain. J. Pharmacol. Exp. Ther. 2003, 306, 387–393. [Google Scholar] [CrossRef]
- Cui, M.; Honore, P.; Zhong, C.; Gauvin, D.; Mikusa, J.; Hernandez, G.; Chandran, P.; Gomtsyan, A.; Brown, B.; Bayburt, E.K.; et al. TRPV1 receptors in the CNS play a key role in broad-spectrum analgesia of TRPV1 antagonists. J. Neurosci. 2006, 26, 9385–9393. [Google Scholar] [CrossRef]
- Hori, T. Capsaicin and central control of thermoregulation. Pharmacol. Ther. 1984, 26, 389–416. [Google Scholar] [CrossRef]
- Gavva, N.R.; Bannon, A.W.; Hovland, D.N., Jr.; Lehto, S.G.; Klionsky, L.; Surapaneni, S.; Immke, D.C.; Henley, C.; Arik, L.; Bak, A.; et al. Repeated administration of vanilloid receptor TRPV1 antagonists attenuates hyperthermia elicited by TRPV1 blockade. J. Pharmacol. Exp. Ther. 2007, 323, 128–137. [Google Scholar] [CrossRef]
- Gavva, N.R.; Bannon, A.W.; Surapaneni, S.; Hovland, D.N., Jr.; Lehto, S.G.; Gore, A.; Juan, T.; Deng, H.; Han, B.; Klionsky, L.; et al. The vanilloid receptor TRPV1 is tonically activated in vivo and involved in body temperature regulation. J. Neurosci. 2007, 27, 3366–3374. [Google Scholar] [CrossRef]
- Steiner, A.A.; Turek, V.F.; Almeida, M.C.; Burmeister, J.J.; Oliveira, D.L.; Roberts, J.L.; Bannon, A.W.; Norman, M.H.; Louis, J.C.; Treanor, J.J.; et al. Nonthermal activation of transient receptor potential vanilloid-1 channels in abdominal viscera tonically inhibits autonomic cold-defense effectors. J. Neurosci. 2007, 27, 7459–7468. [Google Scholar] [CrossRef]
- Chizh, B.A.; O’Donnell, M.B.; Napolitano, A.; Wang, J.; Brooke, A.C.; Aylott, M.C.; Bullman, J.N.; Gray, E.J.; Lai, R.Y.; Williams, P.M.; et al. The effects of the TRPV1 antagonist SB-705498 on TRPV1 receptor-mediated activity and inflammatory hyperalgesia in humans. Pain 2007, 132, 132–141. [Google Scholar] [CrossRef]
- Vay, L.; Gu, C.; McNaughton, P.A. The thermo-TRP ion channel family: Properties and therapeutic implications. Br. J. Pharmacol. 2012, 165, 787–801. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Brito, R.; Sheth, S.; Mukherjea, D.; Rybak, L.P.; Ramkumar, V. TRPV1: A Potential Drug Target for Treating Various Diseases. Cells 2014, 3, 517-545. https://doi.org/10.3390/cells3020517
Brito R, Sheth S, Mukherjea D, Rybak LP, Ramkumar V. TRPV1: A Potential Drug Target for Treating Various Diseases. Cells. 2014; 3(2):517-545. https://doi.org/10.3390/cells3020517
Chicago/Turabian StyleBrito, Rafael, Sandeep Sheth, Debashree Mukherjea, Leonard P. Rybak, and Vickram Ramkumar. 2014. "TRPV1: A Potential Drug Target for Treating Various Diseases" Cells 3, no. 2: 517-545. https://doi.org/10.3390/cells3020517
APA StyleBrito, R., Sheth, S., Mukherjea, D., Rybak, L. P., & Ramkumar, V. (2014). TRPV1: A Potential Drug Target for Treating Various Diseases. Cells, 3(2), 517-545. https://doi.org/10.3390/cells3020517