
Circulating Mesenchymal Stromal Cells in Patients with Infantile Hemangioma: Evaluation of Their Functional Capacity and Gene Expression Profile

 , , , , , , , , and
, , , , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Clinical Procedures
2.2. In Vitro Cultures Generating CFU-F Cells
2.3. Cell Phenotypes
2.4. Proliferative Capacity
2.5. Differentiation Capacity
2.6. Gene Expression Analysis
2.7. Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) and CCAAT/Enhancer-Binding Protein Alpha (C/EBPα)
2.8. In Vitro Angiogenesis Assay
2.9. Statistical Analysis
3. Results
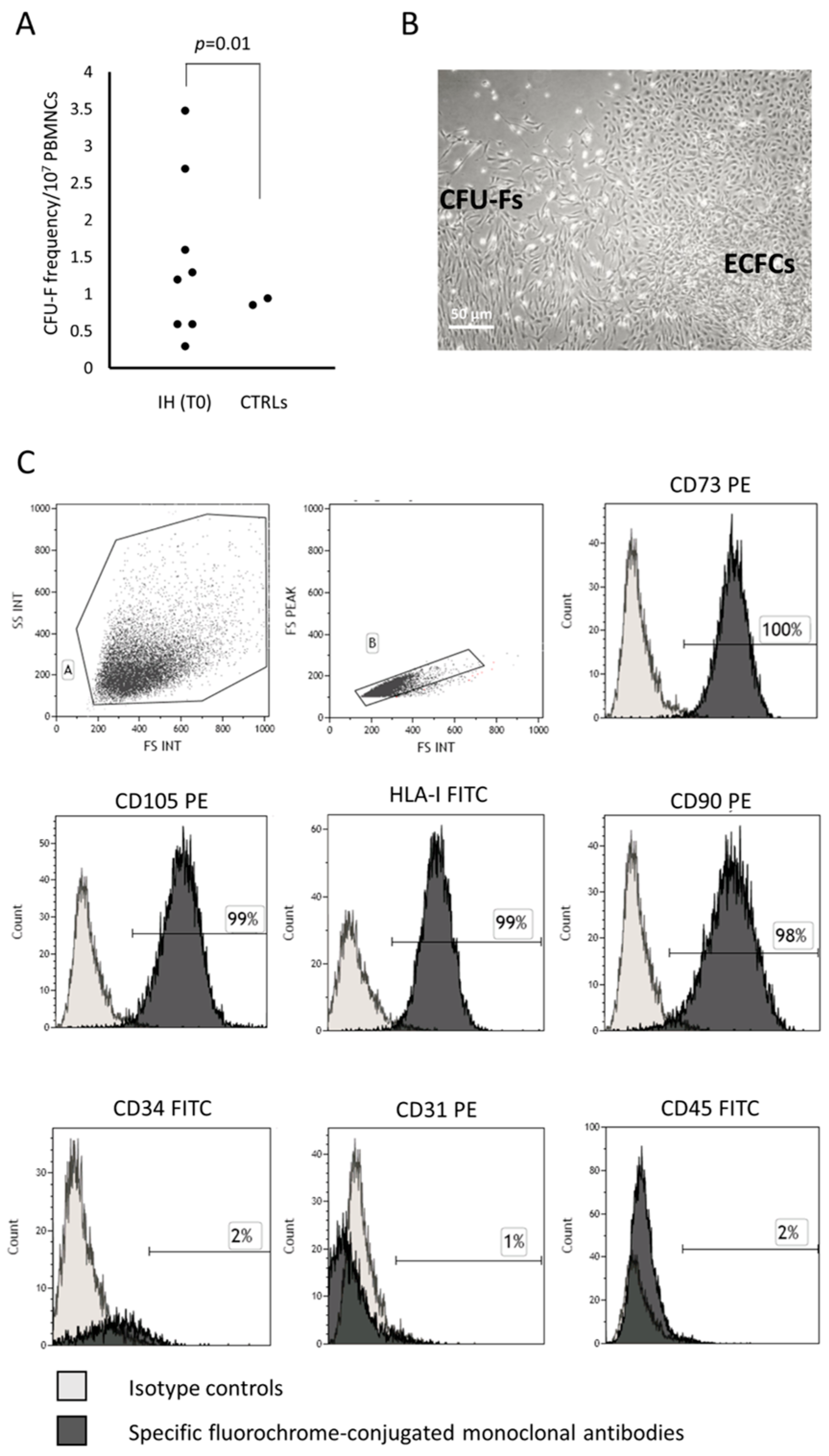
3.1. CFU-Fs Circulate in PB of Patients with IH at T0
3.1.1. CFU-F Characterization
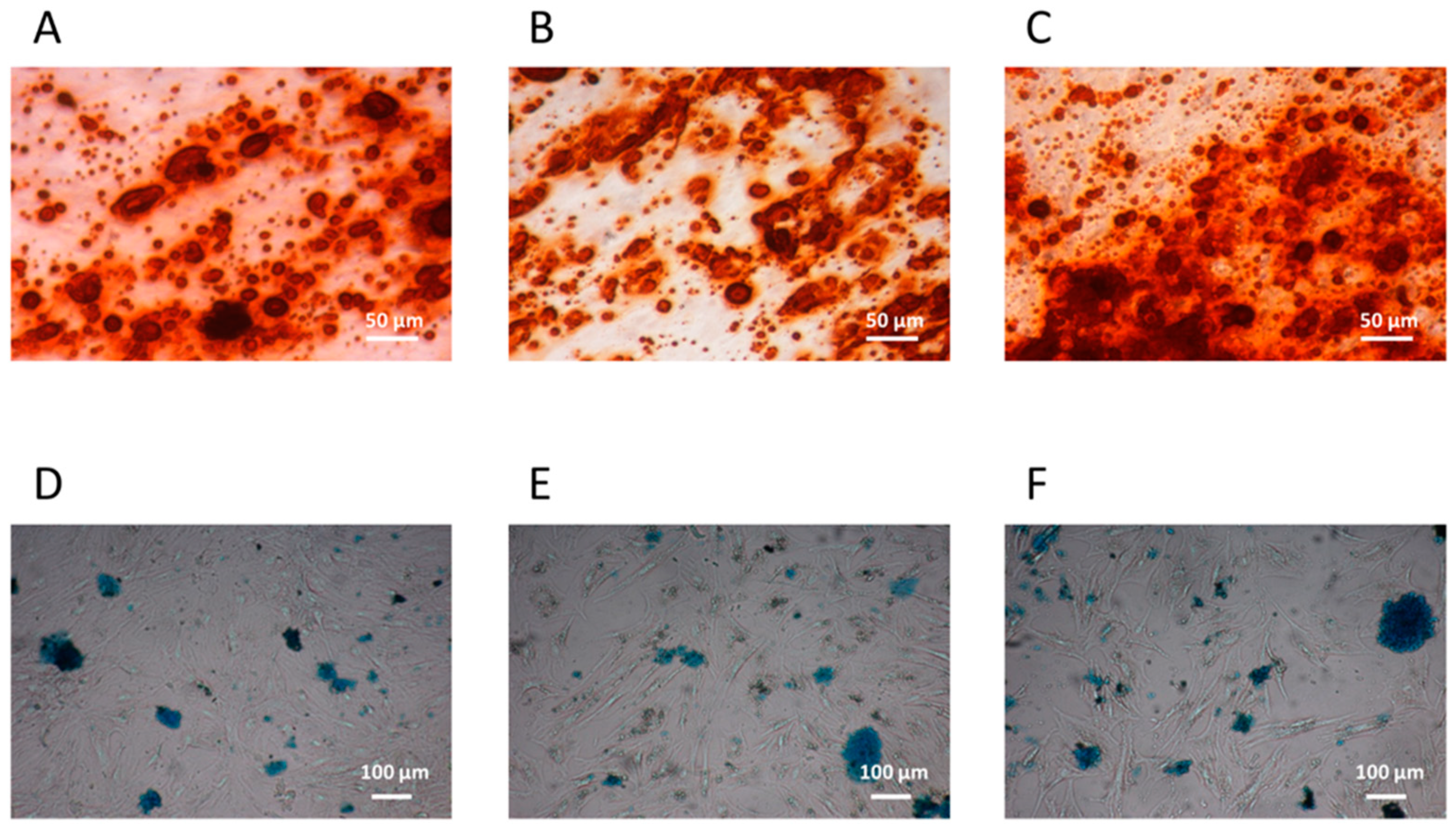
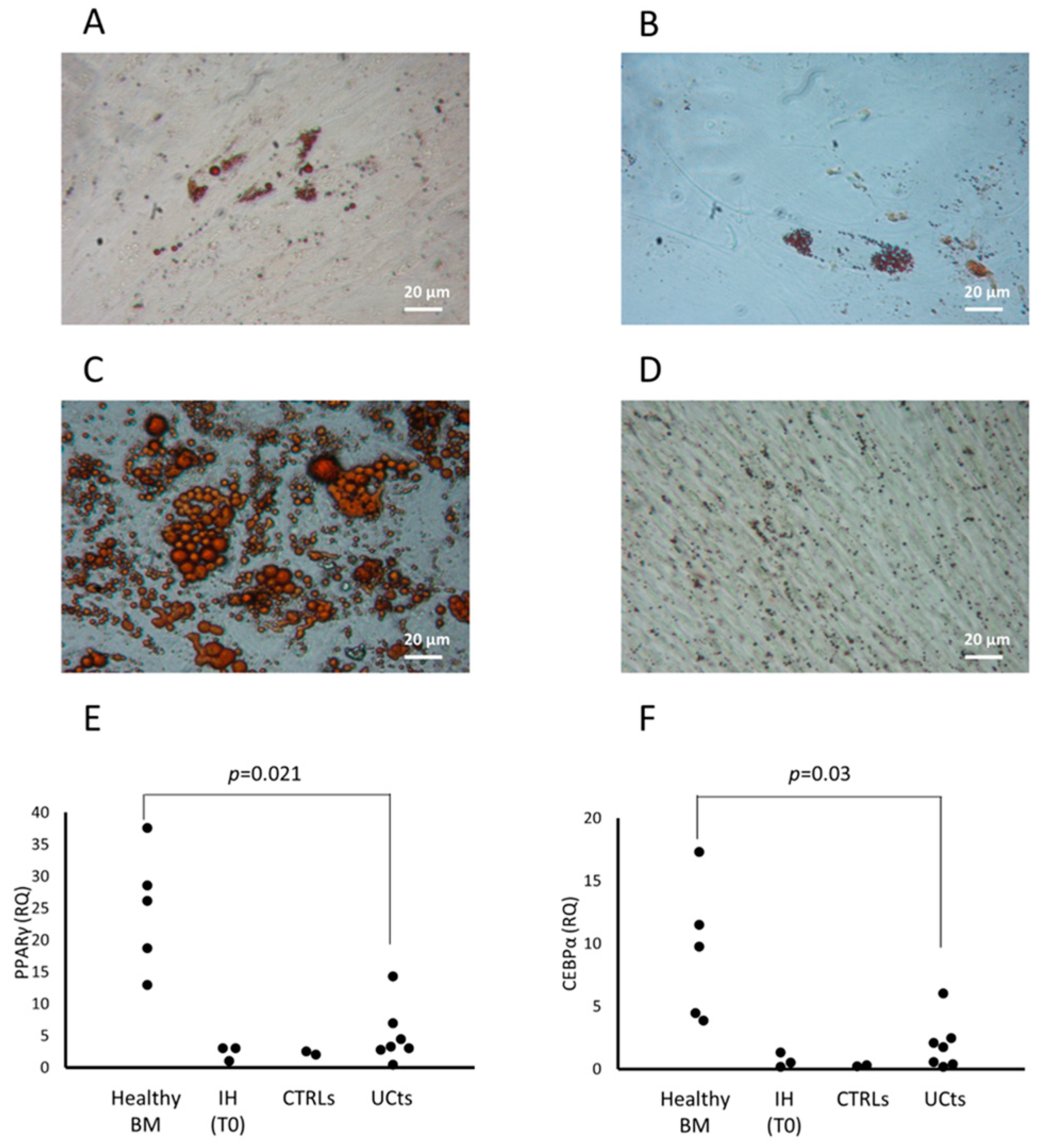
3.1.2. CFU-F Differentiation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yes CFU-Fs (n = 8) | No CFU-Fs (n = 10) | |
|---|---|---|
| Sex (male/total) | 4/8 | 6/10 |
| Gestational age (wk) | 38 ± 3 | 36 ± 2 |
| Weight (g) | 2775 ± 847 | 2005 ± 471 |
| Delivery (Caesarean section/total) | 4/8 | 6/10 |
| Maternal pathology (n/total) | 2/8 | 1/10 |
| Antenatal prednisone (n/total) | 4/8 | 6/10 |
| Cardiac pathologies (n/total) | 1/8 | 2/10 |
| Hemangioma dimension at birth: <5 mm | 6 | 6 |
| >5 mm | 2 | 4 |
| Hemangioma number at sampling: 1 | 6 | 7 |
| >1 | 2 | 3 |
| IH aspect at birth: Surface | 7 | 9 |
| Deep | 2 | 2 |
| Mixed | 0 | 0 |
| Position: Head and neck | 6 | 5 |
| Thorax abdomen | 2 | 4 |
| Upper and lower limbs | 1 | 4 |
| Glutes | 1 | 2 |
| Hemangioma dimension at sampling: <2 cm | 1 | 0 |
| >2 cm | 7 | 10 |
| Colour: Light | 3 | 1 |
| Strong | 5 | 9 |
| Complication (n/total) | 0/8 | 3/10 |
| Complete regression * | 7/7 | 6/10 |
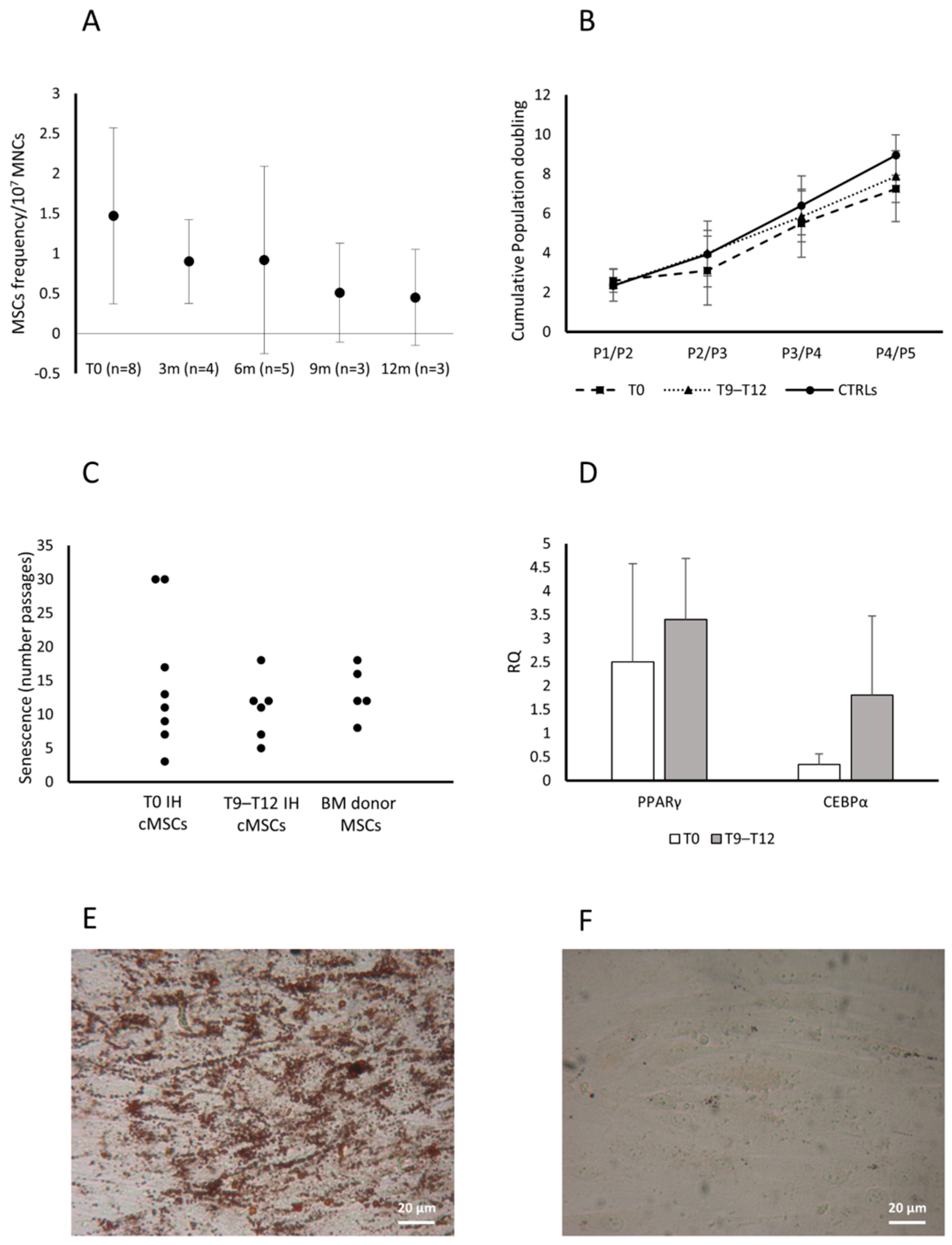
3.2. cMSCs in Patients with IH Receiving Propranolol
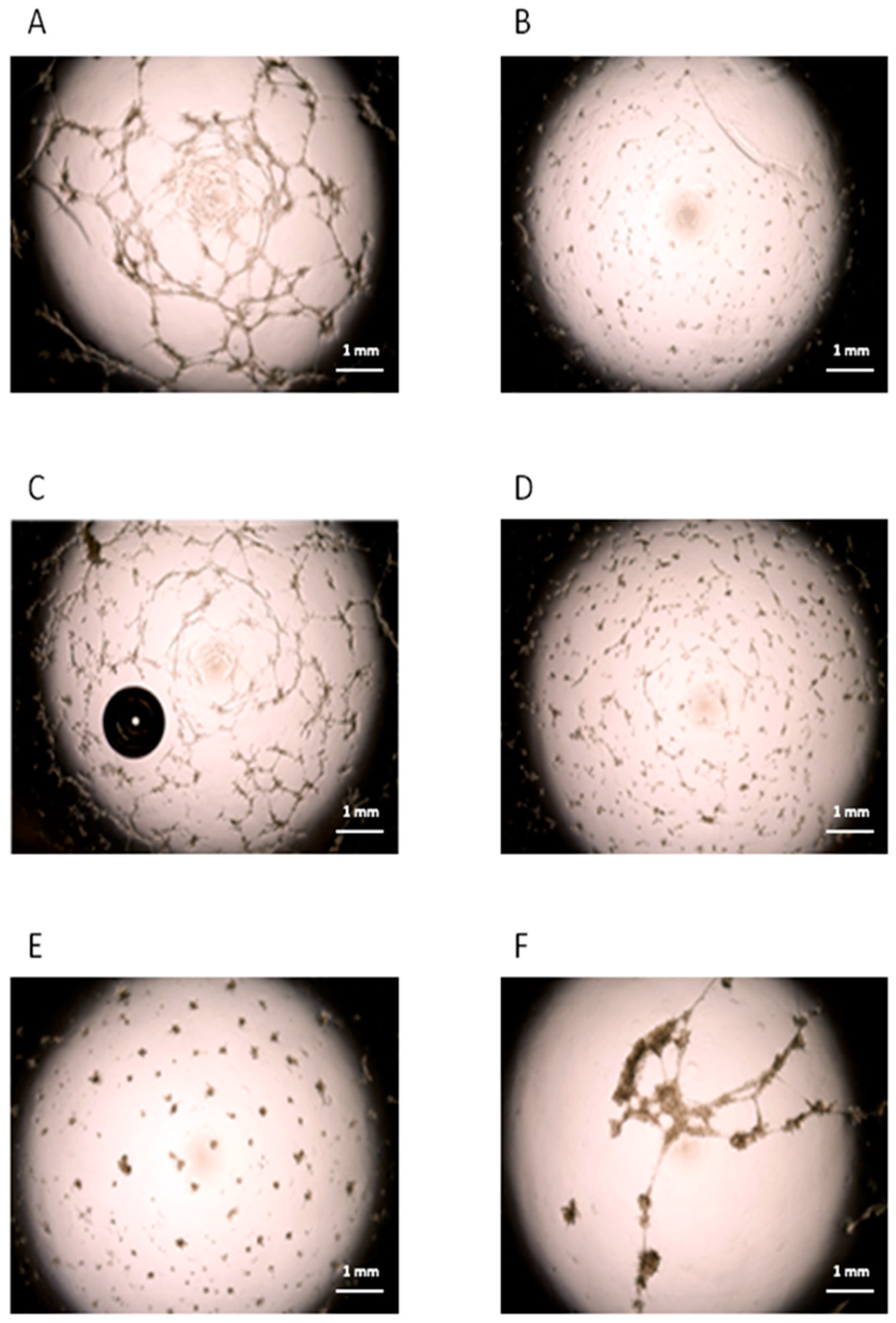
3.3. Role of cMSCs in the In Vitro Angiogenesis Assay
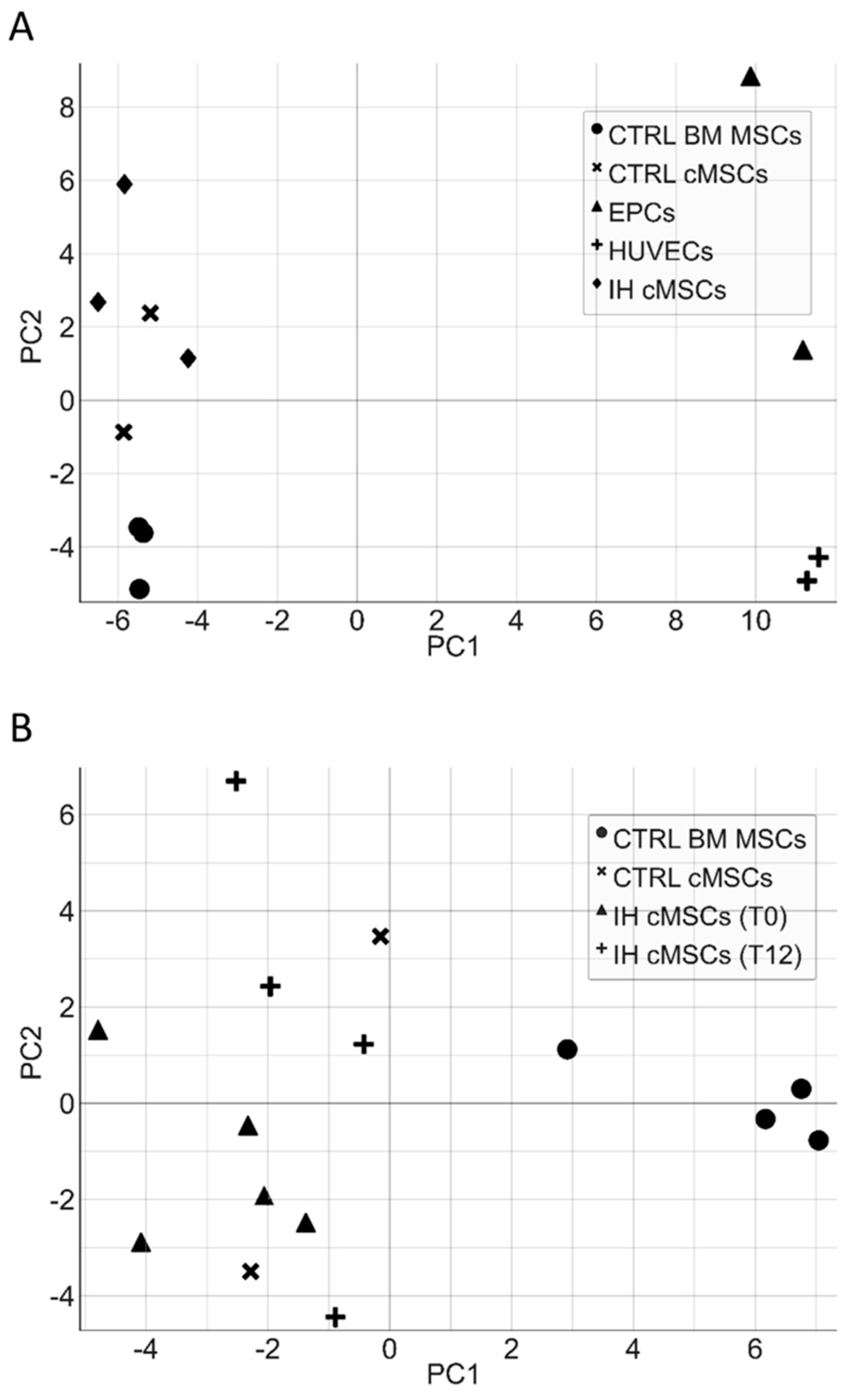
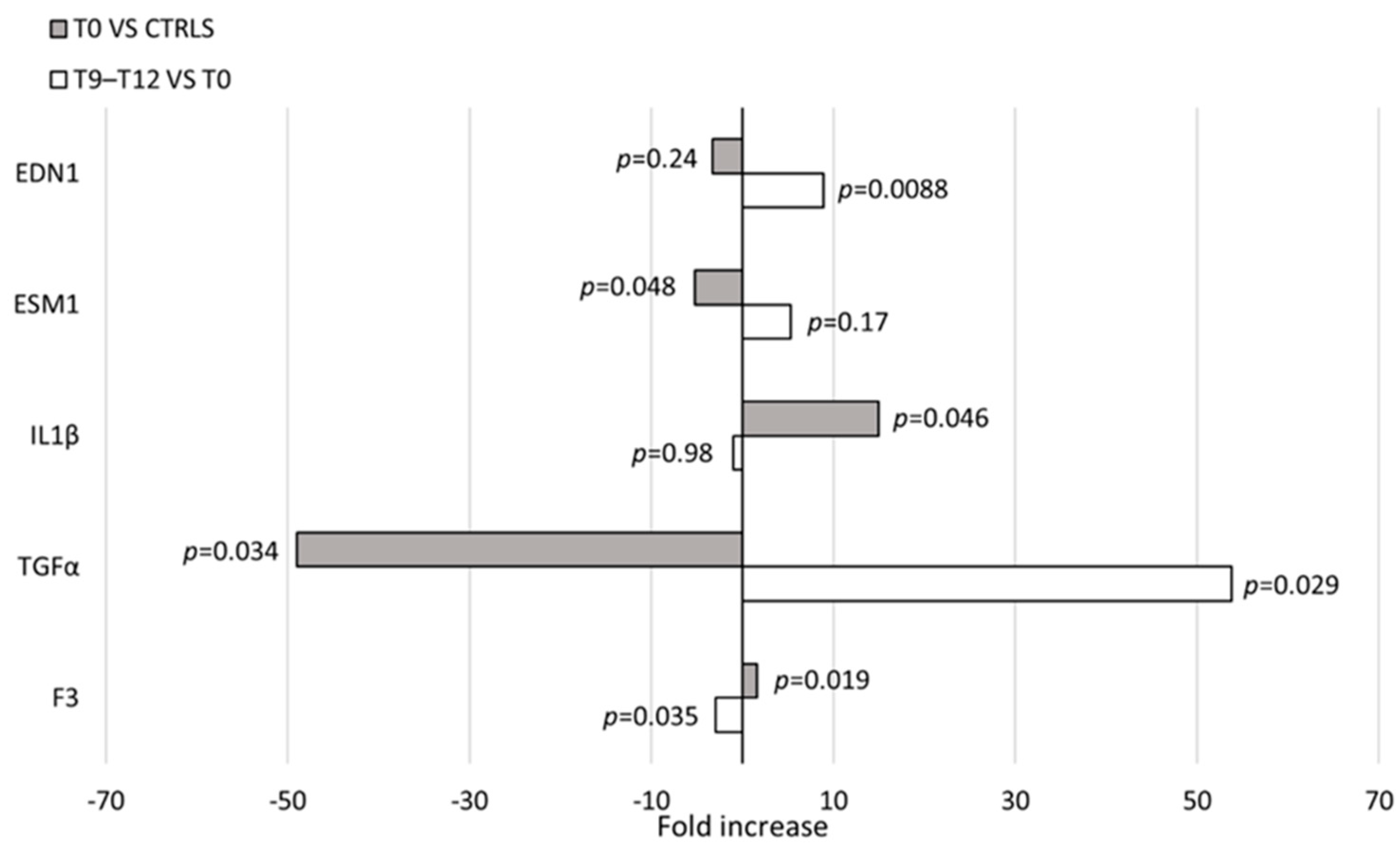
3.4. cMSC Gene Expression Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chang, L.C.; Haggstrom, A.N.; Drolet, B.A.; Baselga, E.; Chamlin, S.L.; Garzon, M.C.; Horii, K.A.; Lucky, A.W.; Mancini, A.J.; Metry, D.W.; et al. Growth characteristics of infantile hemangiomas: Implications for management. Pediatrics 2008, 122, 360–367. [Google Scholar] [CrossRef]
- Bauland, C.G.; Lüning, T.H.; Smit, J.M.; Zeebregts, C.J.; Spauwen, P.H.M. Untreated hemangiomas: Growth pattern and residual lesions. Plast. Reconstr. Surg. 2011, 127, 1643–1648. [Google Scholar] [CrossRef] [PubMed]
- Greenberger, S.; Bischoff, J. Pathogenesis of infantile haemangioma. Br. J. Dermatol. 2013, 169, 12–19. [Google Scholar] [CrossRef]
- Itinteang, T.; Withers, A.H.; Davis, P.F.; Tan, S.T. Biology of infantile hemangioma. Front. Surg. 2014, 1, 38. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, J. Progenitor cells in infantile hemangioma. J. Craniofac. Surg. 2009, 20 (Suppl. S1), 695–697. [Google Scholar] [CrossRef]
- Ritter, M.R.; Butschek, R.A.; Friedlander, M.; Friedlander, S.F. Pathogenesis of infantile haemangioma: New molecular and cellular insights. Expert Rev. Mol. 2007, 9, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Fuhr, J.; Boye, E.; Gyorffy, S.; Soker, S.; Atala, A.; Mulliken, J.B.; Bischoff, J. Mesenchymal stem cells and adipogenesis in hemangioma involution. Stem Cells 2006, 24, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.M.; Chen, R.L.; Shen, W.M.; Chen, H.N.; Zhou, X.J. Mesenchymal stem cells in infantile hemangioma reside in the perivascular region. Pediatr. Dev. Pathol. 2012, 15, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.A.; Boscolo, E.; Picard, A.; Psutka, S.; Melero-Martin, J.M.; Bartch, T.C.; Mulliken, J.B.; Bischoff, J. Multipotential stem cells recapitulate human infantile hemangioma in immunodeficient mice. J. Clin. Investig. 2008, 118, 2592–2599. [Google Scholar] [CrossRef]
- Léauté-Labrèze, C.; Hoeger, P.; Mazereeuw-Hautier, J.; Guibaud, L.; Baselga, E.; Posiunas, G.; Phillips, R.J.; Caceres, H.; Lopez Gutierrez, J.C.; Ballona, R.; et al. A randomized, controlled trial of oral propranolol in infantile hemangioma. N. Engl. J. Med. 2015, 372, 735–746. [Google Scholar] [CrossRef]
- Albiñana, V.; Gallardo-Vara, E.; Casado-Vela, J.; Recio-Poveda, L.; Botella, L.M.; Cuesta, A.M. Propranolol: A "Pick and Roll" Team Player in Benign Tumors and Cancer Therapies. J. Clin. Med. 2022, 11, 4539. [Google Scholar] [CrossRef]
- Li, H.H.; Lou, Y.; Zhang, R.R.; Xie, J.; Cao, D.S. Propranolol Accelerats Hemangioma Stem Cell Transformation into Adipocyte. Ann. Plast. Surg. 2019, 83, e5–e13. [Google Scholar] [CrossRef]
- Campanelli, R.; Codazzi, A.C.; Poletto, V.; Abbà, C.; Catarsi, P.; Fois, G.; Avanzini, M.A.; Brazzelli, V.; Tzialla, C.; De Silvestri, A.; et al. Kinetic and Angiogenic Activity of Circulating Endothelial Colony Forming Cells in Patients with Infantile Haemangioma Receiving Propranolol. Thromb. Haemost. 2019, 119, 274–284. [Google Scholar] [CrossRef]
- Moccia, F.; Zuccolo, E.; Poletto, V.; Cinelli, M.; Bonetti, E.; Guerra, G.; Rosti, V. Endothelial progenitor cells support tumour growth and metastatisation: Implications for the resistance to anti-angiogenic therapy. Tumour Biol. 2015, 36, 6603–6614. [Google Scholar] [CrossRef]
- Moccia, F.; Dragoni, S.; Lodola, F.; Bonetti, E.; Bottino, C.; Guerra, G.; Laforenza, U.; Rosti, V.; Tanzi, F. Store-dependent Ca(2+) entry in endothelial progenitor cells as a perspective tool to enhance cell-based therapy and adverse tumour vascularization. Curr. Med. Chem. 2012, 19, 5802–5818. [Google Scholar] [CrossRef]
- Borghesi, A.; Massa, M.; Campanelli, R.; Bollani, L.; Tzialla, C.; Figar, T.A.; Ferrari, G.; Bonetti, E.; Chiesa, G.; de Silvestri, A.; et al. Circulating endothelial progenitor cells in preterm infants with bronchopulmonary dysplasia. Am. J. Respir. Crit. Care Med. 2009, 180, 540–546. [Google Scholar] [CrossRef]
- Marketou, M.E.; Parthenakis, F.I.; Kalyva, A.; Pontikoglou, C.; Maragkoudakis, S.; Kontaraki, J.E.; Zacharis, E.A.; Patrianakos, A.; Chlouverakis, G.; Papadaki, H.A.; et al. Circulating mesenchymal stem cells in patients with hypertrophic cardiomyopathy. Cardiovasc. Pathol. 2015, 24, 149–153. [Google Scholar] [CrossRef]
- Emamnejad, R.; Sahraian, M.; Shakiba, Y.; Salehi, Z.; Masoomi, A.; Imani, D.; Najafi, F.; Laribi, B.; Shirzad, H.; Izad, M. Circulating mesenchymal stem cells, stromal derived factor (SDF)-1 and IP-10 levels increased in clinically active multiple sclerosis patients but not in clinically stable patients treated with beta interferon. Mult. Scler. Relat. Disord. 2019, 35, 233–238. [Google Scholar] [CrossRef]
- Patry, C.; Doniga, T.; Lenz, F.; Viergutz, T.; Weiss, C.; Tönshoff, B.; Kalenka, A.; Yard, B.; Krebs, J.; Schaible, T.; et al. Increased mobilization of mesenchymal stem cells in patients with acute respiratory distress syndrome undergoing extracorporeal membrane oxygenation. PLoS ONE 2020, 15, e0227460. [Google Scholar] [CrossRef]
- Pouryazdanpanah, N.; Vahidi, R.; Dabiri, S.; Derakhshani, A.; Farsinezhad, A. Use of Some Additives for Improving Mesenchymal Stem Cell Isolation Outcomes in Non-Mobilized Peripheral Blood. Arch. Iran. Med. 2018, 21, 362–367. [Google Scholar]
- Zvaifler, N.J.; Marinova-Mutafchieva, L.; Adams, G.; Edwards, C.J.; Moss, J.; Burger, J.A.; Maini, R.N. Mesenchymal precursor cells in the blood of normal individuals. Arthritis Res. 2000, 2, 477–488. [Google Scholar] [CrossRef]
- Coccini, T.; De Simone, U.; Roccio, M.; Croce, S.; Lenta, E.; Zecca, M.; Spinillo, A.; Avanzini, M.A. In vitro toxicity screening of magnetite nanoparticles by applying mesenchymal stem cells derived from human umbilical cord lining. J. Appl. Toxicol. 2019, 39, 1320–1336. [Google Scholar] [CrossRef]
- Mantelli, M.; Avanzini, M.A.; Rosti, V.; Ingo, D.M.; Conforti, A.; Novara, F.; Arrigo, G.; Boni, M.; Zappatore, R.; Lenta, E.; et al. Comprehensive characterization of mesenchymal stromal cells from patients with Fanconi anaemia. Br. J. Haematol. 2015, 170, 826–836. [Google Scholar] [CrossRef]
- Bernardo, M.E.; Avanzini, M.A.; Perotti, C.; Cometa, A.M.; Moretta, A.; Lenta, E.; Del Fante, C.; Novara, F.; de Silvestri, A.; Amendola, G.; et al. Optimization of in vitro expansion of human multipotent mesenchymal stromal cells for cell-therapy approaches: Further insights in the search for a fetal calf serum substitute. J. Cell Physiol. 2007, 211, 121–130. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Borghesi, A.; Avanzini, M.A.; Novara, F.; Mantelli, M.; Lenta, E.; Achille, V.; Cerbo, R.M.; Tzialla, C.; Longo, S.; De Silvestri, A.; et al. Genomic alterations in human umbilical cord-derived mesenchymal stromal cells call for stringent quality control before any possible therapeutic approach. Cytotherapy 2013, 15, 1362–1373. [Google Scholar] [CrossRef]
- Mastrolia, I.; Foppiani, E.M.; Murgia, A.; Candini, O.; Samarelli, A.V.; Grisendi, G.; Veronesi, E.; Horwitz, E.M.; Dominici, M. Challenges in Clinical Development of Mesenchymal Stromal/Stem Cells: Concise Review. Stem Cells Trans. Med. 2019, 8, 1135–1148. [Google Scholar] [CrossRef]
- Volonté, M.; Codazzi, A.C.; Davidovich, S.; Apicella, A.; Isoletta, E.; Barruscotti, S.; Massa, M.; De Silvestri, A.; Marseglia, G.L.; Brazzelli, V. Propranolol for the treatment of infantile hemangiomas: A nine-year monocentric experience from a tertiary hospital. Eur. J. Dermatol. EJD 2023, 33, 265–269. [Google Scholar] [CrossRef]
- Hall, B.; Andreeff, M.; Marini, F. The participation of mesenchymal stem cells in tumor stroma formation and their application as targeted-gene delivery vehicles. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar] [CrossRef]
- Bui, K.C.; Weems, M.; Biniwale, M.; George, A.A.; Zielinska, E.; Azen, C.G.; Durand, M.; Abdel-Azim, H. Circulating hematopoietic and endothelial progenitor cells in newborn infants: Effects of gestational age, postnatal age and clinical stress in the first 3 weeks of life. Early Hum. Dev. 2013, 89, 411–418. [Google Scholar] [CrossRef]
- Zhang, L.; Mai, H.M.; Zheng, J.; Zheng, J.W.; Wang, Y.A.; Qin, Z.P.; Li, K.L. Propranolol inhibits angiogenesis via down-regulating the expression of vascular endothelial growth factor in hemangioma derived stem cell. Int. J. Clin. Exp. Pathol. 2014, 7, 48–55. [Google Scholar]
- Ma, X.; Zhao, T.; Ouyang, T.; Xin, S.; Ma, Y.; Chang, M. Propranolol enhanced adipogenesis instead of induction of apoptosis of hemangiomas stem cells. Int. J. Clin. Exp. Pathol. 2014, 7, 3809–3817. [Google Scholar]
- England, R.W.; Hardy, K.L.; Kitajewski, A.M.; Wong, A.; Kitajewski, J.K.; Shawber, C.J.; Wu, J.K. Propranolol promotes accelerated and dysregulated adipogenesis in hemangioma stem cells. Ann. Plast. Surg. 2014, 73 (Suppl. S1), S119–S124. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.; Hardy, K.L.; Kitajewski, A.M.; Shawber, C.J.; Kitajewski, J.K.; Wu, J.K. Propranolol accelerates adipogenesis in hemangioma stem cells and causes apoptosis of hemangioma endothelial cells. Plast. Reconstr. Surg. 2012, 130, 1012–1021. [Google Scholar] [CrossRef]
- Makkeyah, S.M.; Elseedawy, M.E.; Abdel-Kader, H.M.; Mokhtar, G.M.; Ragab, I.A. Vascular endothelial growth factor response with propranolol therapy in patients with infantile hemangioma. Pediatr. Hematol. Oncol. 2022, 39, 215–224. [Google Scholar] [CrossRef]
- Yuan, W.; Wang, X. Propranolol Participates in the Treatment of Infantile Hemangioma by Inhibiting HUVECs Proliferation, Migration, Invasion, and Tube Formation. Biomed. Res. Int. 2021, 2021, 6636891. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shen, Y.W.; Zhang, L.J.; Chen, J.J.; Bian, H.T.; Gu, W.J.; Chen, H.Z.; Zhang, W.D.; Luan, X. Targeting Endothelial Cell-Specific Molecule 1 Protein in Cancer: A Promising Therapeutic Approach. Front. Oncol. 2021, 11, 687120. [Google Scholar] [CrossRef]
- Peña, E.; de la Torre, R.; Arderiu, G.; Slevin, M.; Badimon, L. mCRP triggers angiogenesis by inducing F3 transcription and TF signalling in microvascular endothelial cells. Thromb. Haemost. 2017, 117, 357–370. [Google Scholar] [CrossRef]
- Lee, H.-J.; Shi, H.; Brönneke, H.S.; Jin, B.-Y.; Choi, S.-H.; Seeley, R.J.; Kim, D.-H. Vascular reactivity contributes to adipose tissue remodeling in obesity. J. Endocrinol. 2021, 251, 195–206. [Google Scholar] [CrossRef]
- Sakata, M.; Kunimoto, K.; Kawaguchi, A.; Inaba, Y.; Kaminaka, C.; Yamamoto, Y.; Kakimoto, N.; Suenaga, T.; Tokuhara, D.; Jinnin, M. Analysis of cytokine profiles in sera of single and multiple infantile hemangioma. J. Dermatol. 2023, 50, 906–911. [Google Scholar] [CrossRef]
- Gornostaeva, A.N.; Buravkova, L.B. Changes Induced by Inflammatory-Activated Immune Cell Microenvironment in the Paracrine Profile of MSC. Bull. Exp. Biol. Med. 2023, 174, 544–548. [Google Scholar] [CrossRef]
- Barra, J.; Cerda-Infante, J.; Sandoval, L.; Gajardo-Meneses, P.; Henriquez, J.F.; Labarca, M.; Metz, C.; Venegas, J.; Retamal, C.; Oyanadel, C.; et al. D-Propranolol Impairs EGFR Trafficking and Destabilizes Mutant p53 Counteracting AKT Signaling and Tumor Malignancy. Cancers 2021, 13, 3622. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbà, C.; Croce, S.; Valsecchi, C.; Lenta, E.; Campanelli, R.; Codazzi, A.C.; Brazzelli, V.; Carolei, A.; Catarsi, P.; Acquafredda, G.; et al. Circulating Mesenchymal Stromal Cells in Patients with Infantile Hemangioma: Evaluation of Their Functional Capacity and Gene Expression Profile. Cells 2024, 13, 254. https://doi.org/10.3390/cells13030254
Abbà C, Croce S, Valsecchi C, Lenta E, Campanelli R, Codazzi AC, Brazzelli V, Carolei A, Catarsi P, Acquafredda G, et al. Circulating Mesenchymal Stromal Cells in Patients with Infantile Hemangioma: Evaluation of Their Functional Capacity and Gene Expression Profile. Cells. 2024; 13(3):254. https://doi.org/10.3390/cells13030254
Chicago/Turabian StyleAbbà, Carlotta, Stefania Croce, Chiara Valsecchi, Elisa Lenta, Rita Campanelli, Alessia C. Codazzi, Valeria Brazzelli, Adriana Carolei, Paolo Catarsi, Gloria Acquafredda, and et al. 2024. "Circulating Mesenchymal Stromal Cells in Patients with Infantile Hemangioma: Evaluation of Their Functional Capacity and Gene Expression Profile" Cells 13, no. 3: 254. https://doi.org/10.3390/cells13030254
APA StyleAbbà, C., Croce, S., Valsecchi, C., Lenta, E., Campanelli, R., Codazzi, A. C., Brazzelli, V., Carolei, A., Catarsi, P., Acquafredda, G., Apicella, A., Caliogna, L., Berni, M., Mannarino, S., Avanzini, M. A., Rosti, V., & Massa, M. (2024). Circulating Mesenchymal Stromal Cells in Patients with Infantile Hemangioma: Evaluation of Their Functional Capacity and Gene Expression Profile. Cells, 13(3), 254. https://doi.org/10.3390/cells13030254