

Crystal Structure of a Classical MHC Class I Molecule in Dogs; Comparison of DLA-88*0 and DLA-88*5 Category Molecules

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Analysis of DLA-88 Sequences
2.2. Synthesis of Viral Peptides
2.3. Protein Preparation
2.4. The Assembly and Purification of the pDLA-88*001:04 Complex
2.5. Thermostability Measurements Using Circular Dichroism Spectroscopy
2.6. Crystallization and Data Collection
2.7. Structure Determination and Refinement
2.8. Analysis and Figure Depictions of Structural Data
3. Results
3.1. DLA-88 Sequences in Dogs and Gray Wolves; the LQW Motif in the α2 Domain That Dsitinguishes Categories DLA-88*0 and DLA-88*5 Is not Restricted to a Single DLA-88 Lineage
3.2. Overall Structure of the pDLA-88*001:04 Complex

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter or Statistic | RTI9/DLA-88*001:04/β2m |
|---|---|
| Data processing | |
| Space group | P212121 |
| Cell parameters (Å) | A = 89.3, b = 93.0, c = 119.4 |
| Resolution range (Å) | 50.00–2.70 (2.75–2.70) |
| Total reflections | 152570 |
| Unique reflections | 50958 |
| Completeness | 0.978 (0.969) |
| Rmerge | 0.201 (0.540) |
| I/σ | 7.308 (2.500) |
| R-value Work | 0.253 |
| R-value Free | 0.282 |
| Bonds (Å) | 0.004 |
| Angles (°) | 0.926 |
| Average B factor | 17.013 |
3.3. The Conformation of the Peptide Binding Groove in pDLA-88*001:04
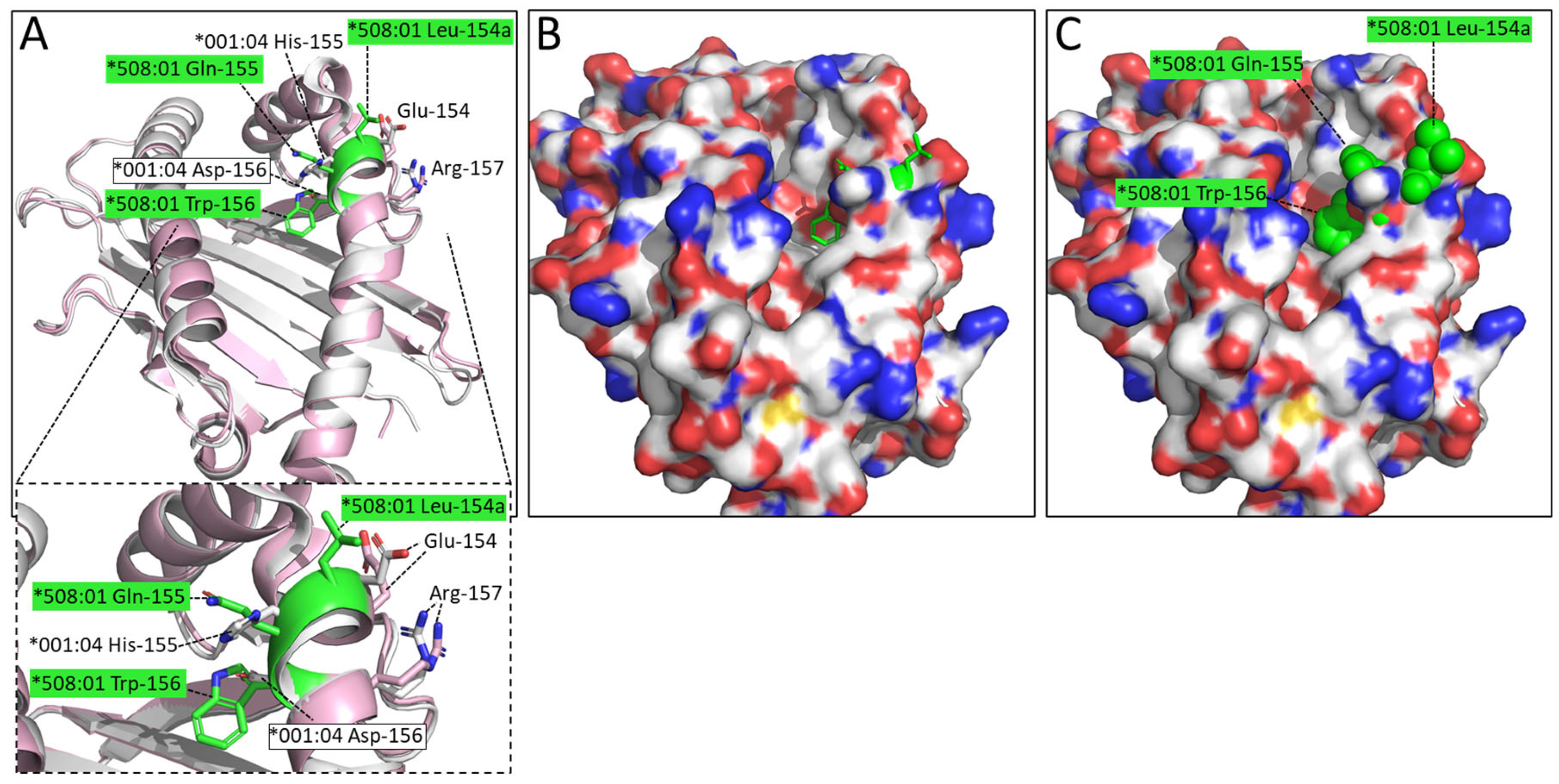
3.4. “Bimodal” Structural Motif Differences between DLA-88 Alleles, Exemplified by pDLA-88*001:04 and pDLA-88*508:01 Structures
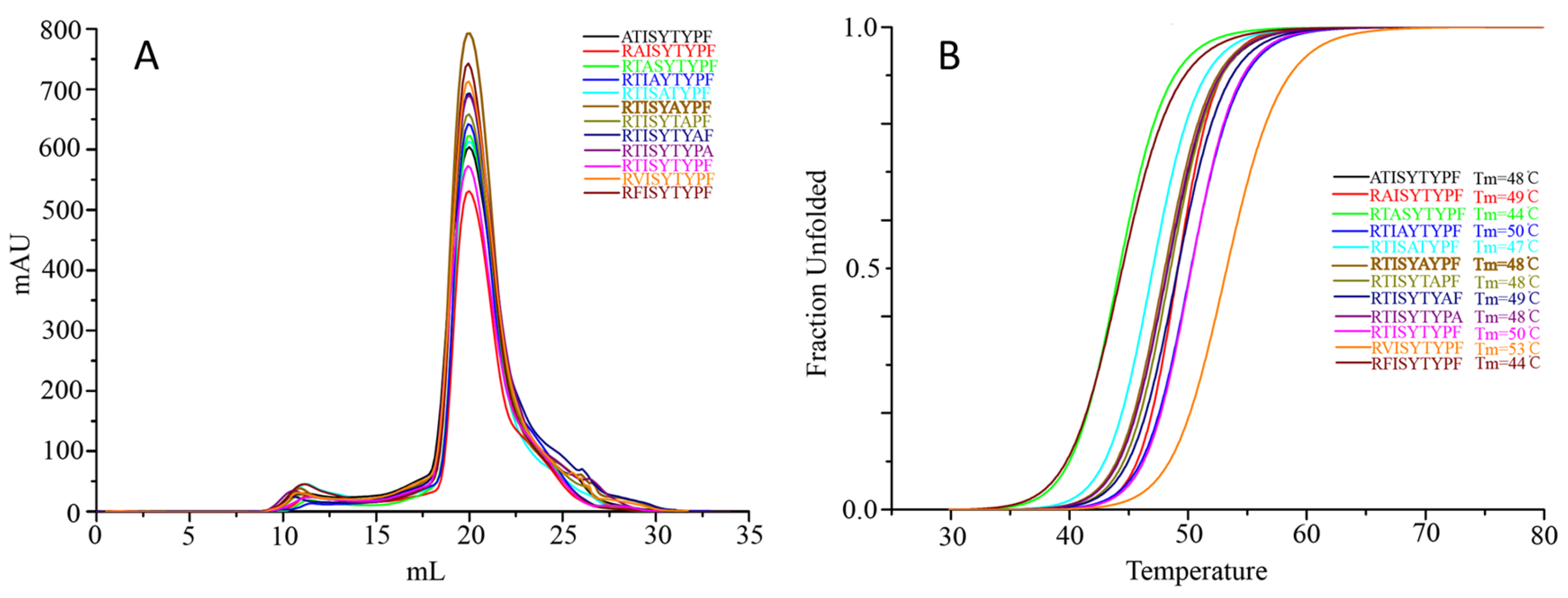
3.5. Amino Acid Substitution Analysis Reveals Anchor Residue Properties at Peptide Ligand Positions P2 and P3
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacHugh, D.E.; Larson, G.; Orlando, L. Taming the Past: Ancient DNA and the Study of Animal Domestication. Annu. Rev. Anim. Biosci. 2017, 5, 329–351. [Google Scholar] [CrossRef] [PubMed]
- Ostrander, E.A.; Wayne, R.K.; Freedman, A.H.; Davis, B.W. Demographic history, selection and functional diversity of the canine genome. Nat. Rev. Genet. 2017, 18, 705–720. [Google Scholar] [CrossRef] [PubMed]
- Knobel, D.L.; Arega, S.; Reininghaus, B.; Simpson, G.J.G.; Gessner, B.D.; Stryhn, H.; Conan, A. Rabies vaccine is associated with decreased all-cause mortality in dogs. Vaccine 2017, 35, 3844–3849. [Google Scholar] [CrossRef] [PubMed]
- Lopez De Turiso, J.A.; Cortes, E.; Martinez, C.; Ruiz de Ybanez, R.; Simarro, I.; Vela, C.; Casal, I. Recombinant vaccine for canine parvovirus in dogs. J. Virol. 1992, 66, 2748–2753. [Google Scholar] [CrossRef]
- Spibey, N.G. Recombinant Canine Adenovirus 2 (CAV-2). U.S. Patent 5616326, 1 April 1997. [Google Scholar]
- Acree, W.M.; Edwards, B.; Black, J.W. Inactivated Canine Coronavirus Vaccine. U.S. Patent 4567042, 28 January 1986. [Google Scholar]
- Day, M.J.; Horzinek, M.C.; Schultz, R.D.; Squires, R.A. WSAVA Guidelines for the vaccination of dogs and cats. J. Small Anim. Prac. 2016, 57, E1–E45. [Google Scholar] [CrossRef]
- Kapil, S.; Yeary, T.; Johnson, B. Diagnostic investigation of emerging viruses of companion animals. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 755–774. [Google Scholar] [CrossRef]
- Felsburg, P.J. Overview of immune system development in the dog: Comparison with humans. Hum. Exp. Toxicol. 2002, 21, 487–492. [Google Scholar] [CrossRef]
- Arce-Fonseca, M.; Ballinas-Verdugo, M.A.; Zenteno, E.R.; Suárez-Flores, D.; Carrillo-Sánchez, S.C.; Alejandre-Aguilar, R.; Rosales-Encina, J.L.; Reyes, P.A.; Rodríguez-Morales, O. Specific humoral and cellular immunity induced by Trypanosoma cruzi DNA immunization in a canine model. Vet. Res. 2013, 44, 15. [Google Scholar] [CrossRef]
- Hennecke, J.; Wiley, D.C. T cell receptor-MHC interactions up close. Cell 2001, 104, 1–4. [Google Scholar] [CrossRef]
- Wagner, J.L. Molecular organization of the canine major histocompatibility complex. J. Hered. 2003, 94, 23–26. [Google Scholar] [CrossRef][Green Version]
- Yuhki, N.; Beck, T.; Stephens, R.; Neelam, B.; O’Brien, S.J. Comparative genomic structure of human, dog, and cat MHC: HLA, DLA, and FLA. J. Hered. 2007, 98, 390–399. [Google Scholar] [CrossRef]
- Appel, M.J.; Shek, W.R.; Summers, B.A. Lymphocyte-mediated immune cytotoxicity in dogs infected with virulent canine distemper virus. Infect. Immun. 1982, 37, 592–600. [Google Scholar] [CrossRef]
- Venkataraman, G.M.; Geraghty, D.; Fox, J.; Graves, S.S.; Zellmer, E.; Storer, B.E.; Torok-Storb, B.J.; Storb, R. Canine DLA-79 gene: An improved typing method, identification of new alleles and its role in graft rejection and graft-versus-host disease. Tissue Antigens 2013, 81, 204–211. [Google Scholar] [CrossRef]
- Graumann, M.B.; DeRose, S.A.; Ostrander, E.A.; Storb, R. Polymorphism analysis of four canine MHC class I genes. Tissue Antigens 1998, 51, 374–381. [Google Scholar] [CrossRef]
- Miyamae, J.; Suzuki, S.; Katakura, F.; Uno, S.; Tanaka, M.; Okano, M.; Matsumoto, T.; Kulski, J.K.; Moritomo, T.; Shiina, T. Identification of novel polymorphisms and two distinct haplotype structures in dog leukocyte antigen class I genes: DLA-88, DLA-12 and DLA-64. Immunogenetics 2018, 70, 237–255. [Google Scholar] [CrossRef]
- Miyamae, J.; Okano, M.; Nishiya, K.; Katakura, F.; Kulski, J.K.; Moritomo, T.; Shiina, T. Haplotype structures and polymorphisms of dog leukocyte antigen (DLA) class I loci shaped by intralocus and interlocus recombination events. Immunogenetics 2022, 74, 245–259. [Google Scholar] [CrossRef]
- Kennedy, L.J.; Angles, J.M.; Barnes, A.; Carter, S.D.; Francino, O.; Gerlach, J.A.; Happ, G.M.; Ollier, W.E.; Thomson, W.; Wagner, J.L. Nomenclature for factors of the dog major histocompatibility system (DLA), 2000: Second report of the ISAG DLA Nomenclature Committee. Tissue Antigens 2001, 58, 55–70. [Google Scholar] [CrossRef]
- Xiao, J.; Xiang, W.; Chai, Y.; Haywood, J.; Qi, J.; Ba, L.; Qi, P.; Wang, M.; Liu, J.; Gao, G.F. Diversified Anchoring Features the Peptide Presentation of DLA-88*50801: First Structural Insight into Domestic Dog MHC Class I. J. Immunol. 2016, 197, 2306–2315. [Google Scholar] [CrossRef]
- Robinson, J.; Mistry, K.; McWilliam, H.; Lopez, R.; Marsh, S.G. IPD—The Immuno Polymorphism Database. Nucleic Acids Res. 2010, 38, D863–D869. [Google Scholar] [CrossRef]
- Dijkstra, J.M. A method for making alignments of related protein sequences that share very little similarity; Shark interleukin 2 as an example. Immunogenetics 2021, 73, 35–51. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Zuckerkandl, E.; Pauling, L. Molecules as documents of evolutionary history. J. Theor. Biol. 1965, 8, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, N.; Qu, Z.; Liang, R.; Zhang, L.; Zhang, B.; Meng, G.; Dijkstra, J.M.; Li, S.; Xia, M.C. A Glimpse of the Peptide Profile Presentation by Xenopus laevis MHC Class I: Crystal Structure of pXela-UAA Reveals a Distinct Peptide-Binding Groove. J. Immunol. 2020, 204, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, J.; Fan, S.; Chen, R.; Liu, Y.; Zhang, J.; Yuan, H.; Liang, R.; Zhang, N.; Xia, C. Structural Definition of Duck Major Histocompatibility Complex Class I Molecules That Might Explain Efficient Cytotoxic T Lymphocyte Immunity to Influenza A Virus. J. Virol. 2017, 91, e02511-16. [Google Scholar] [CrossRef]
- Liang, R.; Sun, Y.; Liu, Y.; Wang, J.; Wu, Y.; Li, Z.; Ma, L.; Zhang, N.; Zhang, L.; Wei, X.; et al. Major Histocompatibility Complex Class I (FLA-E*01801) Molecular Structure in Domestic Cats Demonstrates Species-Specific Characteristics in Presenting Viral Antigen Peptides. J. Virol. 2018, 92, e01631-17. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, L.; Zhang, L.; Li, X.; Xia, C. Crystal structure of the giant panda MHC class I complex: First insights into the viral peptide presentation profile in the bear family. Protein Sci. 2020, 29, 2468–2481. [Google Scholar] [CrossRef]
- Li, X.; Liu, J.; Qi, J.; Gao, F.; Li, Q.; Li, X.; Zhang, N.; Xia, C.; Gao, G.F. Two distinct conformations of a rinderpest virus epitope presented by bovine major histocompatibility complex class I N*01801: A host strategy to present featured peptides. J. Virol. 2011, 85, 6038–6048. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar]
- Adams, P.D.; Grosse-Kunstleve, R.W.; Hung, L.W.; Ioerger, T.R.; McCoy, A.J.; Moriarty, N.W.; Read, R.J.; Sacchettini, J.C.; Sauter, N.K.; Terwilliger, T.C. PHENIX: Building new software for automated crystallographic structure determination. Acta Crystallogr. D Biol. Crystallogr. 2002, 58, 1948–1954. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Moss, D.S.; Thornton, J.M. Main-chain bond lengths and bond angles in protein structures. J. Mol. Biol. 1993, 231, 1049–1067. [Google Scholar] [CrossRef]
- Bailey, S. The CCP4 suite: Programs for protein crystallography. Acta Crystallogr. D Biol. Crystallogr. 1994, 50, 760–763. [Google Scholar]
- Wu, Y.; Zhang, N.; Wei, X.; Lu, S.; Li, S.; Hashimoto, K.; Dijkstra, J.M.; Xia, C. The Structure of a Peptide-Loaded Shark MHC Class I Molecule Reveals Features of the Binding between β2-Microglobulin and H Chain Conserved in Evolution. J. Immunol. 2021, 207, 308–321. [Google Scholar] [CrossRef]
- Miyamae, J.; Okano, M.; Katakura, F.; Kulski, J.K.; Moritomo, T.; Shiina, T. Large-Scale Polymorphism Analysis of Dog Leukocyte Antigen Class I and Class II Genes (DLA-88, DLA-12/88L and DLA-DRB1) and Comparison of the Haplotype Diversity between Breeds in Japan. Cells 2023, 12, 809. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, H.; Sun, G.; Zhao, C.; Shang, S.; Gao, X.; Xia, T.; Yang, X. Characterization of the peripheral blood transcriptome and adaptive evolution of the MHC I and TLR gene families in the wolf (Canis lupus). BMC Genom. 2017, 18, 584. [Google Scholar] [CrossRef]
- Murgia, C.; Pritchard, J.K.; Kim, S.Y.; Fassati, A.; Weiss, R.A. Clonal origin and evolution of a transmissible cancer. Cell 2006, 126, 477–487. [Google Scholar] [CrossRef]
- Saper, M.A.; Bjorkman, P.J.; Wiley, D.C. Refined structure of the human histocompatibility antigen HLA-A2 at 2.6 A resolution. J. Mol. Biol. 1991, 219, 277–319. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, N.; Hashimoto, K.; Xia, C.; Dijkstra, J.M. Structural Comparison Between MHC Classes I and II; in Evolution, a Class-II-Like Molecule Probably Came First. Front. Immunol. 2021, 12, 621153. [Google Scholar] [CrossRef]
- Koch, M.; Camp, S.; Collen, T.; Avila, D.; Salomonsen, J.; Wallny, H.J.; van Hateren, A.; Hunt, L.; Jacob, J.P.; Johnston, F.; et al. Structures of an MHC class I molecule from B21 chickens illustrate promiscuous peptide binding. Immunity 2007, 27, 885–899. [Google Scholar] [CrossRef]
- Madden, D.R.; Garboczi, D.N.; Wiley, D.C. The antigenic identity of peptide-MHC complexes: A comparison of the conformations of five viral peptides presented by HLA-A2. Cell 1993, 75, 693–708. [Google Scholar] [CrossRef] [PubMed]
- Rammensee, H.G.; Friede, T.; Stevanoviíc, S. MHC ligands and peptide motifs: First listing. Immunogenetics 1995, 41, 178–228. [Google Scholar] [CrossRef] [PubMed]
- Yeager, M.; Hughes, A.L. Evolution of the mammalian MHC: Natural selection, recombination, and convergent evolution. Immunol. Rev. 1999, 167, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Aoyagi, K.; Dijkstra, J.M.; Xia, C.; Denda, I.; Ototake, M.; Hashimoto, K.; Nakanishi, T. Classical MHC class I genes composed of highly divergent sequence lineages share a single locus in rainbow trout (Oncorhynchus mykiss). J. Immunol. 2002, 168, 260–273. [Google Scholar] [CrossRef]
- Buslepp, J.; Wang, H.; Biddison, W.E.; Appella, E.; Collins, E.J. A correlation between TCR Valpha docking on MHC and CD8 dependence: Implications for T cell selection. Immunity 2003, 19, 595–606. [Google Scholar] [CrossRef]
- Varani, L.; Bankovich, A.J.; Liu, C.W.; Colf, L.A.; Jones, L.L.; Kranz, D.M.; Puglisi, J.D.; Garcia, K.C. Solution mapping of T cell receptor docking footprints on peptide-MHC. Proc. Natl. Acad. Sci. USA 2007, 104, 13080–13085. [Google Scholar] [CrossRef]
- VonHoldt, B.M.; Ostrander, E.A. The singular history of a canine transmissible tumor. Cell 2006, 126, 445–447. [Google Scholar] [CrossRef][Green Version]
- Mello Martins, M.I.; Ferreira de Souza, F.; Gobello, C. The Canine Transmissible Venereal Tumor: Etiology, Pathology, Diagnosis and Treatment. In Recent Advances in Small Animal Reproduction; Concannon, P.W., England, G., Verstegen, J., III, Linde-Forsberg, C., Eds.; International Veterinary Information Service: Ithaca, NY, USA, 2005; p. A1233.0405. [Google Scholar]
- Li, X.C.; Raghavan, M. Structure and function of major histocompatibility complex class I antigens. Curr. Opin. Organ Transplant. 2010, 15, 499–504. [Google Scholar] [CrossRef]
- Siddle, H.V.; Kreiss, A.; Eldridge, M.D.; Noonan, E.; Clarke, C.J.; Pyecroft, S.; Woods, G.M.; Belov, K. Transmission of a fatal clonal tumor by biting occurs due to depleted MHC diversity in a threatened carnivorous marsupial. Proc. Natl. Acad. Sci. USA 2007, 104, 16221–16226. [Google Scholar] [CrossRef]
- Radwan, J.; Babik, W.; Kaufman, J.; Lenz, T.L.; Winternitz, J. Advances in the Evolutionary Understanding of MHC Polymorphism. Trends Genet. 2020, 36, 298–311. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Dijkstra, J.M. Major Histocompatibility Complex (MHC) Genes and Disease Resistance in Fish. Cells 2019, 8, 378. [Google Scholar] [CrossRef]
- Barth, S.M.; Schreitmüller, C.M.; Proehl, F.; Oehl, K.; Lumpp, L.M.; Kowalewski, D.J.; Di Marco, M.; Sturm, T.; Backert, L.; Schuster, H.; et al. Characterization of the Canine MHC Class I DLA-88*50101 Peptide Binding Motif as a Prerequisite for Canine T Cell Immunotherapy. PLoS ONE 2016, 11, e0167017. [Google Scholar] [CrossRef]
- Ross, P.; Nemec, P.S.; Kapatos, A.; Miller, K.R.; Holmes, J.C.; Suter, S.E.; Buntzman, A.S.; Soderblom, E.J.; Collins, E.J.; Hess, P.R. The canine MHC class Ia allele DLA-88*508, 01 presents diverse self- and canine distemper virus-origin peptides of varying length that have a conserved binding motif. Vet. Immunol. Immunopathol. 2018, 197, 76–86. [Google Scholar] [CrossRef]
- Nemec, P.S.; Kapatos, A.; Holmes, J.C.; Hess, P.R. The prevalent Boxer MHC class Ia allotype dog leukocyte antigen (DLA)-88*034, 01 preferentially binds nonamer peptides with a defined motif. HLA 2018, 92, 403–407. [Google Scholar] [CrossRef]







| Hydrogen Bonds and Salt Bridges | ||||
|---|---|---|---|---|
| Peptide | H Chain | |||
| Residue | Atom | Residue | Atom | Van der Waals Forces |
| P1-R | O | Y159 | OH | Y7, E63, Y159, T163, W167 |
| N | Y7 | OH | ||
| N | Y171 | OH | ||
| P2-T | N | E63 | OE1 | Y7, Y9, M45, E63, T66, Y99, Y159 |
| N | E63 | OE2 | ||
| OG1 | T66 | OG1 | ||
| OG1 | E63 | OE1 | ||
| OG1 | E63 | OE2 | ||
| P3-I | N | Y99 | OH | T66, Y99, H155, D156, Y159 |
| P4-S | O | R73 | NE2 | R62, T66, E69 |
| P5-Y | R73, H155 | |||
| P6-T | OG1 | R73 | NE | T70, R73, Y74 |
| OE1 | R73 | NH2 | ||
| P7-Y | O | W147 | NE1 | W147, A150, G151, T152, H155 |
| P8-P | O | W147 | NE1 | D77, K146 |
| P9-F | OXT | Y84 | OH | D77, T80, L81, Y84, R95, D116, Y123, I124, T143, W147 |
| OXT | T143 | OG1 | ||
| N | D77 | OD1 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Ma, L.; Li, S.; Wang, Y.; Xiao, R.; Yang, J.; Dijkstra, J.M.; Xia, C. Crystal Structure of a Classical MHC Class I Molecule in Dogs; Comparison of DLA-88*0 and DLA-88*5 Category Molecules. Cells 2023, 12, 1097. https://doi.org/10.3390/cells12071097
Sun Y, Ma L, Li S, Wang Y, Xiao R, Yang J, Dijkstra JM, Xia C. Crystal Structure of a Classical MHC Class I Molecule in Dogs; Comparison of DLA-88*0 and DLA-88*5 Category Molecules. Cells. 2023; 12(7):1097. https://doi.org/10.3390/cells12071097
Chicago/Turabian StyleSun, Yujiao, Lizhen Ma, Shen Li, Yawen Wang, Ruiqi Xiao, Junqi Yang, Johannes M. Dijkstra, and Chun Xia. 2023. "Crystal Structure of a Classical MHC Class I Molecule in Dogs; Comparison of DLA-88*0 and DLA-88*5 Category Molecules" Cells 12, no. 7: 1097. https://doi.org/10.3390/cells12071097
APA StyleSun, Y., Ma, L., Li, S., Wang, Y., Xiao, R., Yang, J., Dijkstra, J. M., & Xia, C. (2023). Crystal Structure of a Classical MHC Class I Molecule in Dogs; Comparison of DLA-88*0 and DLA-88*5 Category Molecules. Cells, 12(7), 1097. https://doi.org/10.3390/cells12071097