
Hypoblast Formation in Bovine Embryos Does Not Depend on NANOG


Abstract
1. Introduction
2. Materials and Methods
2.1. CRISPR/Cas9-Mediated KO of NANOG in Adult Fibroblasts
2.2. Production and Analysis of SCNT and IVP Embryos
2.3. Modulation of Signaling Pathways
2.4. Immunofluorescence Staining and Confocal Laser Scanning Microscopy
2.5. Statistical Analysis
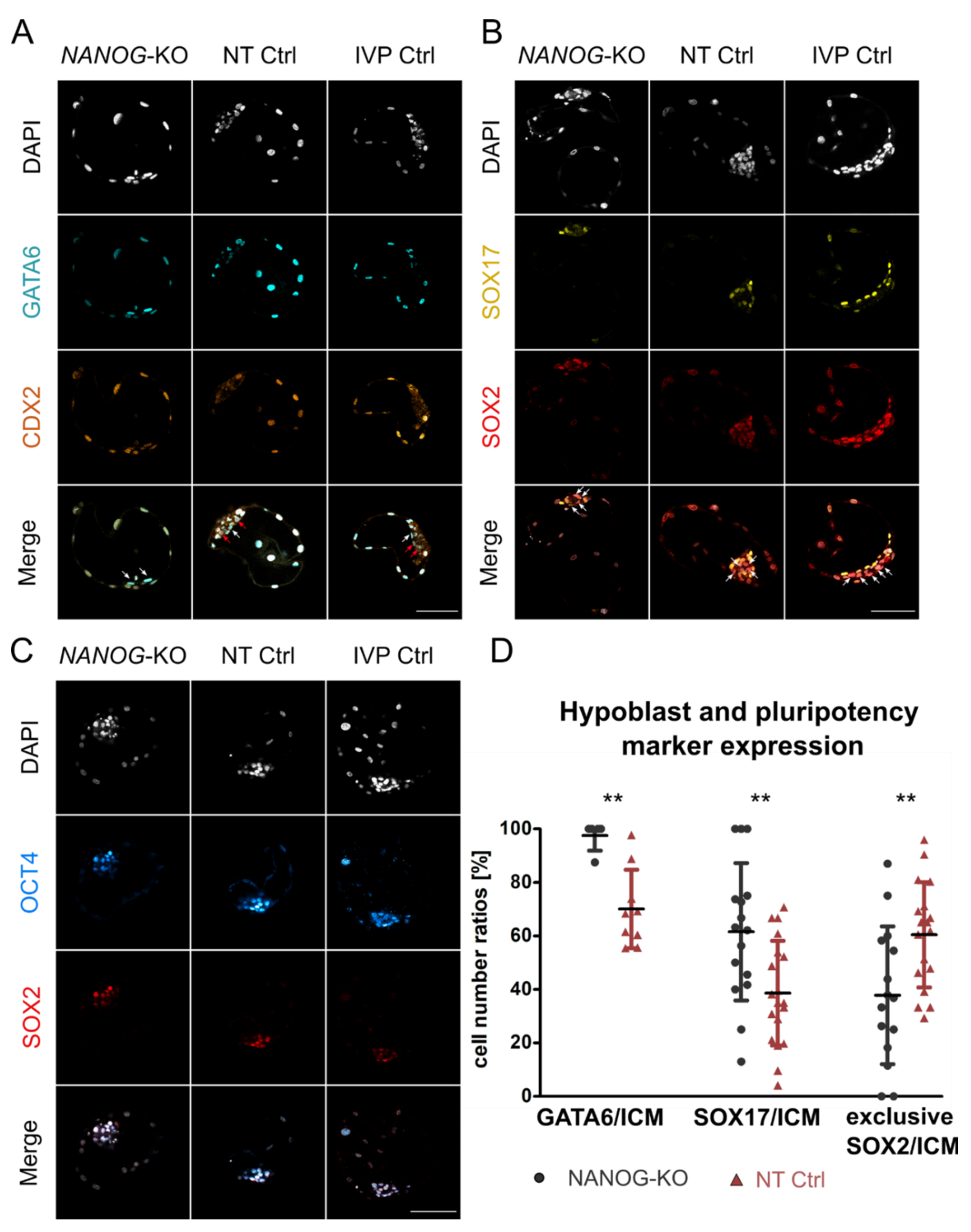
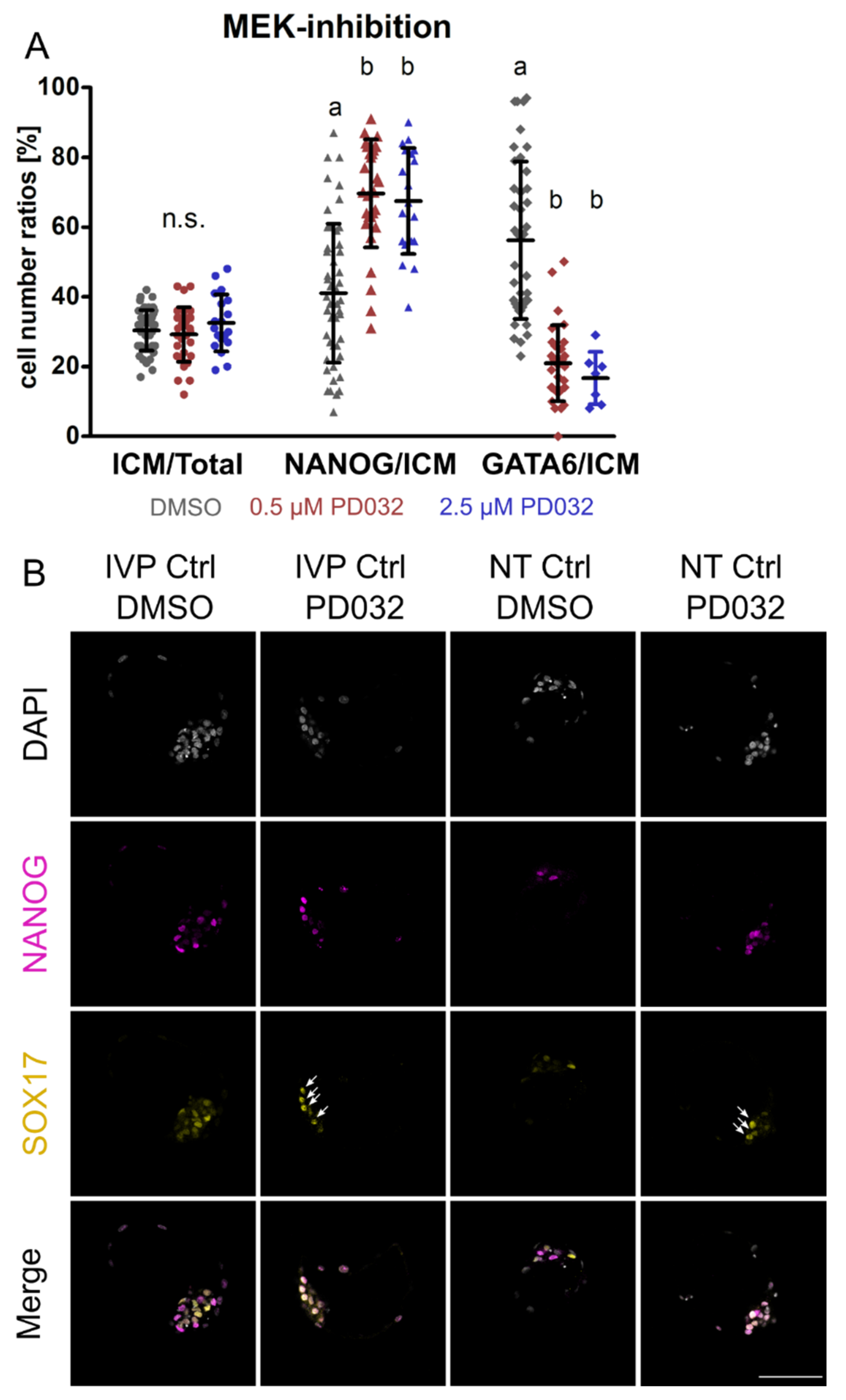
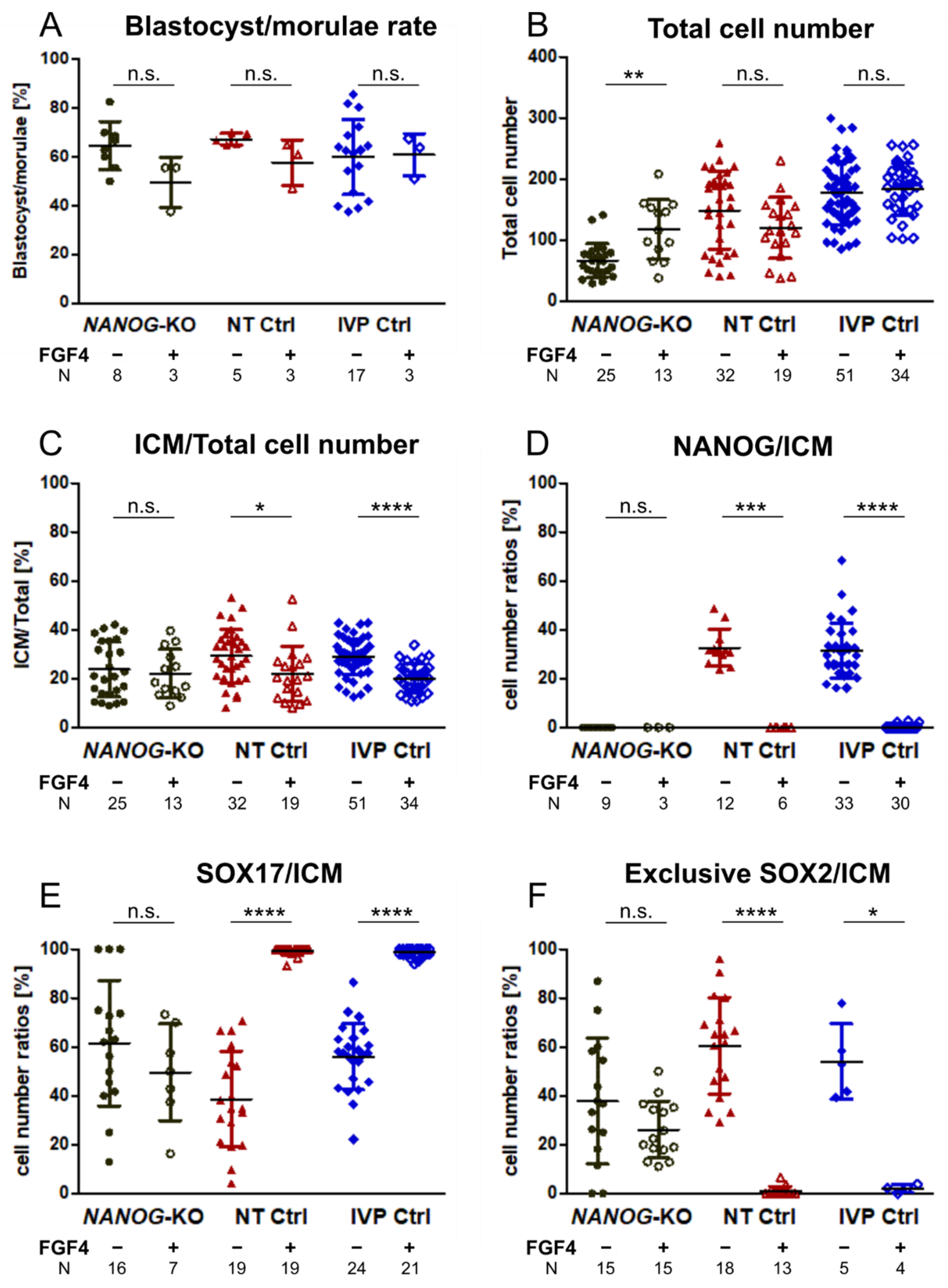
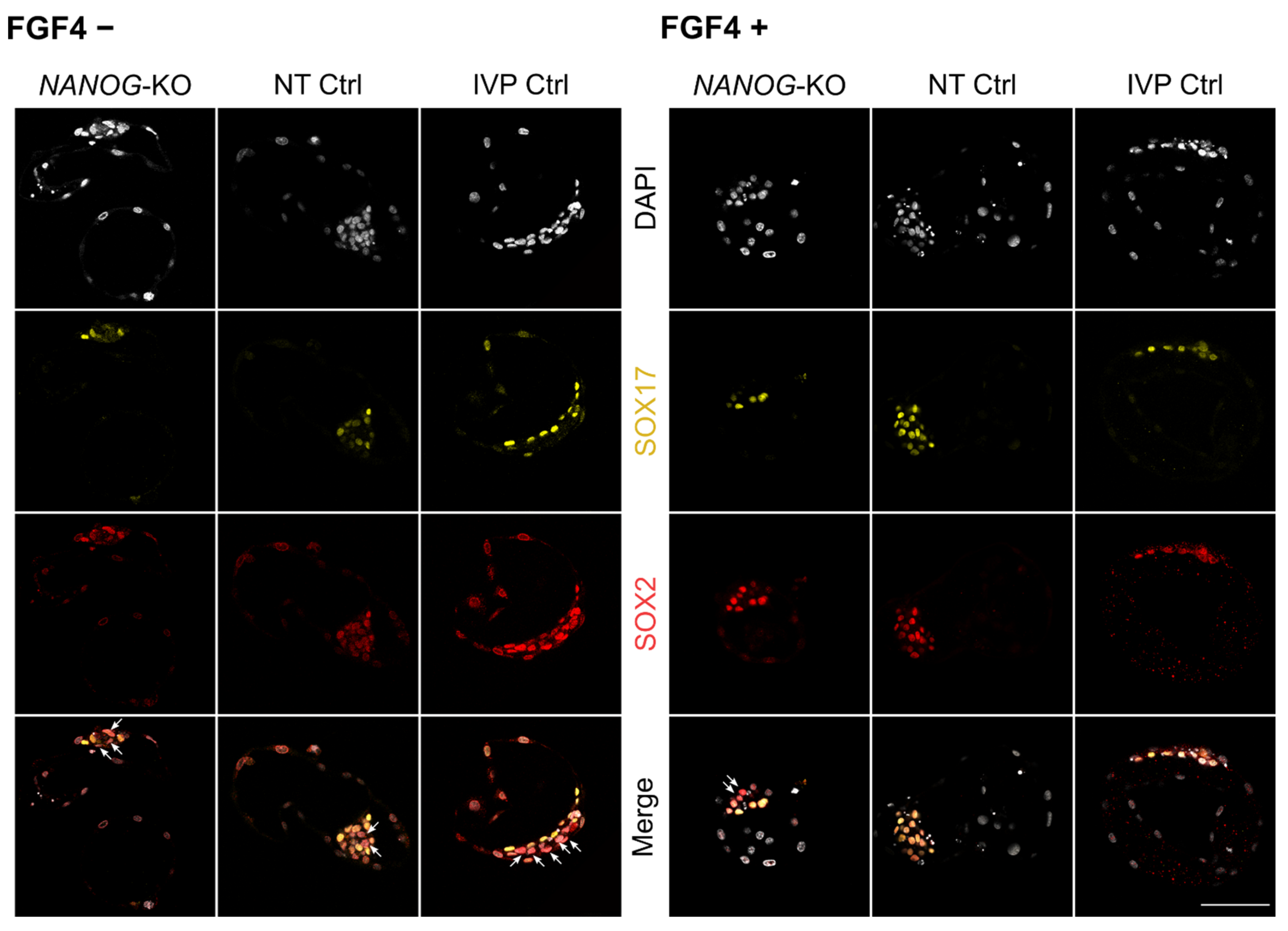
3. Results
3.1. NANOG-KO Has No Effect on Blastocyst Rate but Results in Reduced Total Cell Number
3.2. NANOG Is Dispensable for Expression of Pluripotency Factors and Hypoblast Markers
3.3. Inhibition of MEK Induces Cell Death in NANOG-KO Embryos
3.4. FGF4 in NANOG-KO Embryos Does Not Convert the Entire ICM to Hypoblast Precursor Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BME | Basal Medium Eagle’s amino acids solution |
| CLSM | Confocal laser scanning microscope |
| EPI | Epiblast |
| ESCs | Embryonic stem cells |
| FGFR | FGF-receptor |
| HB | Hypoblast |
| ICM | inner cell mass |
| IVP | In vitro production |
| KO | Knockout |
| MAPK | Mitogen-activated protein kinase |
| MEM | Minimum Essential Medium |
| OCS | Estrous cow serum |
| PD032 | PD0325901 |
| PE | Primitive endoderm |
| RTK | Receptor tyrosine kinase |
| SCNT | Somatic cell nuclear transfer |
| SD | Standard deviation |
| TE | Trophectoderm |
References
- Messerschmidt, D.M.; Kemler, R. Nanog is required for primitive endoderm formation through a non-cell autonomous mechanism. Dev. Biol. 2010, 344, 129–137. [Google Scholar] [CrossRef]
- Frankenberg, S.; Gerbe, F.; Bessonnard, S.; Belville, C.; Pouchin, P.; Bardot, O.; Chazaud, C. Primitive endoderm differentiates via a three-step mechanism involving Nanog and RTK signaling. Dev. Cell 2011, 21, 1005–1013. [Google Scholar] [CrossRef]
- Cai, K.Q.; Capo-Chichi, D.C.; Rula, M.E.; Yang, D.-H.; Xu, X.-X. Dynamic GATA6 expression in primitive endoderm formation and maturation in early mouse embryogenesis. Dev. Dyn. 2008, 237, 2820–2829. [Google Scholar] [CrossRef]
- Morris, S.A.; Teo, R.T.Y.; Li, H.; Robson, P.; Glover, D.M.; Zernicka-Goetz, M. Origin and formation of the first two distinct cell types of the inner cell mass in the mouse embryo. Proc. Natl. Acad. Sci. USA 2010, 107, 6364–6369. [Google Scholar] [CrossRef]
- Plusa, B.; Piliszek, A.; Frankenberg, S.; Artus, J.; Hadjantonakis, A.-K. Distinct sequential cell behaviours direct primitive endoderm formation in the mouse blastocyst. Development 2008, 135, 3081–3091. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, K.; Tokuzawa, Y.; Itoh, H.; Segawa, K.; Murakami, M.; Takahashi, K.; Maruyama, M.; Maeda, M.; Yamanaka, S. The homeoprotein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell 2003, 113, 631–642. [Google Scholar] [CrossRef]
- Chambers, I.; Silva, J.C.R.; Colby, D.; Nichols, J.; Nijmeijer-Winter, B.; Robertson, M.; Vrana, J.; Jones, K.; Grotewold, L.; Smith, A. Nanog safeguards pluripotency and mediates germline development. Nature 2007, 450, 1230–1234. [Google Scholar] [CrossRef]
- Chambers, I.; Colby, D.; Robertson, M.; Nichols, J.; Lee, S.; Tweedie, S.; Smith, A. Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell 2003, 113, 643–655. [Google Scholar] [CrossRef]
- Piliszek, A.; Madeja, Z.E. Pre-implantation development of domestic animals. Curr. Top. Dev. Biol. 2018, 128, 267–294. [Google Scholar] [CrossRef]
- Fogarty, N.M.E.; McCarthy, A.; Snijders, K.E.; Powell, B.E.; Kubikova, N.; Blakeley, P.; Lea, R.; Elder, K.; Wamaitha, S.; Kim, D.; et al. Genome editing reveals a role for OCT4 in human embryogenesis. Nature 2017, 550, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Simmet, K.; Zakhartchenko, V.; Philippou-Massier, J.; Blum, H.; Klymiuk, N.; Wolf, E. OCT4/POU5F1 is required for NANOG expression in bovine blastocysts. Proc. Natl. Acad. Sci. USA 2018, 115, 2770–2775. [Google Scholar] [CrossRef] [PubMed]
- Frum, T.; Halbisen, M.A.; Wang, C.; Amiri, H.; Robson, P.; Ralston, A. Oct4 cell-autonomously promotes primitive endoderm development in the mouse blastocyst. Dev. Cell 2013, 25, 610–622. [Google Scholar] [CrossRef]
- Le Bin, G.C.; Muñoz-Descalzo, S.; Kurowski, A.; Leitch, H.; Lou, X.; Mansfield, W.; Etienne-Dumeau, C.; Grabole, N.; Mulas, C.; Niwa, H.; et al. Oct4 is required for lineage priming in the developing inner cell mass of the mouse blastocyst. Development 2014, 141, 1001–1010. [Google Scholar] [CrossRef]
- Simmet, K.; Zakhartchenko, V.; Wolf, E. Comparative aspects of early lineage specification events in mammalian embryos—Insights from reverse genetics studies. Cell Cycle 2018, 17, 1688–1695. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.; Silva, J.; Roode, M.; Smith, A. Suppression of Erk signalling promotes ground state pluripotency in the mouse embryo. Development 2009, 136, 3215–3222. [Google Scholar] [CrossRef] [PubMed]
- Chazaud, C.; Yamanaka, Y.; Pawson, T.; Rossant, J. Early lineage segregation between epiblast and primitive endoderm in mouse blastocysts through the Grb2-MAPK pathway. Dev. Cell 2006, 10, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, Y.; Lanner, F.; Rossant, J. FGF signal-dependent segregation of primitive endoderm and epiblast in the mouse blastocyst. Development 2010, 137, 715–724. [Google Scholar] [CrossRef]
- Kuijk, E.W.; van Tol, L.T.A.; Van de Velde, H.; Wubbolts, R.; Welling, M.; Geijsen, N.; Roelen, B.A.J. The roles of FGF and MAP kinase signaling in the segregation of the epiblast and hypoblast cell lineages in bovine and human embryos. Development 2012, 139, 871–882. [Google Scholar] [CrossRef]
- McLean, Z.; Meng, F.; Henderson, H.; Turner, P.; Oback, B. Increased MAP kinase inhibition enhances epiblast-specific gene expression in bovine blastocysts. Biol. Reprod. 2014, 91, 1–10. [Google Scholar] [CrossRef][Green Version]
- Warzych, E.; Pawlak, P.; Lechniak, D.; Madeja, Z.E. WNT signalling supported by MEK/ERK inhibition is essential to maintain pluripotency in bovine preimplantation embryo. Dev. Biol. 2020, 463, 63–76. [Google Scholar] [CrossRef]
- Roode, M.; Blair, K.; Snell, P.; Elder, K.; Marchant, S.; Smith, A.; Nichols, J. Human hypoblast formation is not dependent on FGF signalling. Dev. Biol. 2012, 361, 358–363. [Google Scholar] [CrossRef]
- Canizo, J.R.; Rivolta, A.E.Y.; Echegaray, C.V.; Suvá, M.; Alberio, V.; Aller, J.F.; Guberman, A.S.; Salamone, D.F.; Alberio, R.H.; Alberio, R. A dose-dependent response to MEK inhibition determines hypoblast fate in bovine embryos. BMC Dev. Biol. 2019, 19, 13. [Google Scholar] [CrossRef] [PubMed]
- Springer, C.; Wolf, E.; Simmet, K. A new toolbox in experimental embryology—Alternative model organisms for studying preimplantation development. J. Dev. Biol. 2021, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Schöler, H.R. Role of Oct4 in the early embryo development. Cell Regen. 2014, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Nichols, J.; Theunissen, T.W.; Guo, G.; van Oosten, A.L.; Barrandon, O.; Wray, J.; Yamanaka, S.; Chambers, I.; Smith, A. Nanog is the gateway to the pluripotent ground state. Cell 2009, 138, 722–737. [Google Scholar] [CrossRef] [PubMed]
- Hastreiter, S.; Skylaki, S.; Loeffler, D.; Reimann, A.; Hilsenbeck, O.; Hoppe, P.S.; Coutu, D.L.; Kokkaliaris, K.D.; Schwarzfischer, M.; Anastassiadis, K.; et al. Inductive and selective effects of GSK3 and MEK inhibition on Nanog heterogeneity in embryonic stem cells. Stem Cell Rep. 2018, 11, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Oron, E.; Nelson, B.; Razis, S.; Ivanova, N. Distinct lineage specification roles for NANOG, OCT4, and SOX2 in human embryonic stem cells. Cell Stem Cell 2012, 10, 440–454. [Google Scholar] [CrossRef] [PubMed]
- Madeja, Z.E.; Sosnowski, J.; Hryniewicz, K.; Warzych, E.; Pawlak, P.; Rozwadowska, N.; Plusa, B.; Lechniak, D. Changes in sub-cellular localisation of trophoblast and inner cell mass specific transcription factors during bovine preimplantation development. BMC Dev. Biol. 2013, 13, 32. [Google Scholar] [CrossRef]
- Kuijk, E.W.; Du Puy, L.; Van Tol, H.T.A.; Oei, C.H.Y.; Haagsman, H.P.; Colenbrander, B.; Roelen, B.A.J. Differences in early lineage segregation between mammals. Dev. Dyn. 2008, 237, 918–927. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.S.; Kelleher, A.M.; O’Neil, E.; Benne, J.; Cecil, R.; Spencer, T.E. NANOG is required to form the epiblast and maintain pluripotency in the bovine embryo. Mol. Reprod. Dev. 2020, 87, 152–160. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed]
- Vochozkova, P.; Simmet, K.; Jemiller, E.-M.; Wünsch, A.; Klymiuk, N. Gene Editing in Primary Cells of Cattle and Pig. Breast Cancer 2019, 1961, 271–289. [Google Scholar] [CrossRef]
- Bauersachs, S.; Ulbrich, S.E.; Zakhartchenko, V.; Minten, M.; Reichenbach, M.; Reichenbach, H.-D.; Blum, H.; Spencer, T.; Wolf, E. The endometrium responds differently to cloned versus fertilized embryos. Proc. Natl. Acad. Sci. USA 2009, 106, 5681–5686. [Google Scholar] [CrossRef]
- Messinger, S.M.; Albertini, D.F. Centrosome and microtubule dynamics during meiotic progression in the mouse oocyte. J. Cell Sci. 1991, 100, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Wuensch, A.; Habermann, F.A.; Kurosaka, S.; Klose, R.; Zakhartchenko, V.; Reichenbach, H.-D.; Sinowatz, F.; McLaughlin, K.J.; Wolf, E. Quantitative Monitoring of Pluripotency Gene Activation after Somatic Cloning in Cattle1. Biol. Reprod. 2007, 76, 983–991. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Peng, C. Automated quantification and analysis of cell counting procedures using ImageJ plugins. J. Vis. Exp. 2016, 117, e54719. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Kunath, T.; Hadjantonakis, A.-K.; Nagy, A.; Rossant, J. Promotion of trophoblast stem cell proliferation by FGF4. Science 1998, 282, 2072–2075. [Google Scholar] [CrossRef]
- Nichols, J.; Zevnik, B.; Anastassiadis, K.; Niwa, H.; Klewe-Nebenius, D.; Chambers, I.; Schöler, H.; Smith, A. Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell 1998, 95, 379–391. [Google Scholar] [CrossRef]
- Molè, M.A.; Coorens, T.H.H.; Shahbazi, M.N.; Weberling, A.; Weatherbee, B.A.T.; Gantner, C.W.; Sancho-Serra, C.; Richardson, L.; Drinkwater, A.; Syed, N.; et al. A single cell characterisation of human embryogenesis identifies pluripotency transitions and putative anterior hypoblast centre. Nat. Commun. 2021, 12, 3679. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.K.; Smith, C.S.; Pearton, D.J.; Wells, D.N.; Broadhurst, R.; Donnison, M.; Pfeffer, P.L. Trophectoderm lineage determination in cattle. Dev. Cell 2011, 20, 244–255. [Google Scholar] [CrossRef]
- Meng, F.; Forrester-Gauntlett, B.; Turner, P.; Henderson, H.; Oback, B. Signal inhibition reveals JAK/STAT3 pathway as critical for bovine inner cell mass development. Biol. Reprod. 2015, 93, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Meilhac, S.M.; Adams, R.J.; Morris, S.A.; Danckaert, A.; Le Garrec, J.-F.; Zernicka-Goetz, M. Active cell movements coupled to positional induction are involved in lineage segregation in the mouse blastocyst. Dev. Biol. 2009, 331, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Piliszek, A.; Artus, J.; Hadjantonakis, A.-K. FGF4 is required for lineage restriction and salt-and-pepper distribution of primitive endoderm factors but not their initial expression in the mouse. Development 2013, 140, 267–279. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | NANOG-KO | NT Ctrl | IVP Ctrl |
|---|---|---|---|
| No. of experiments | 8 | 5 | 17 |
| Blastocysts/zygotes [%] | 29.0 ± 12.9 | 36.1 ± 9.9 | 28.3 ± 4.2 |
| No. of analyzed embryos | 25 | 32 | 51 |
| Total cell number | 66.4 ± 27.3 a | 148.6 ± 65.6 b | 177.4 ± 52.2 b |
| ICM/total cell number [%] | 23.8 ± 11.3 | 29.2 ± 11.0 | 29.0 ± 7.3 |
| ICM number | 15.8 ± 10.5 a | 43.5 ± 24.5 b | 51.3 ± 20.0 b |
| TE cell number | 50.6 ± 22.6 a | 105.1 ± 47.2 b | 126.1 ± 40.5 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Springer, C.; Zakhartchenko, V.; Wolf, E.; Simmet, K. Hypoblast Formation in Bovine Embryos Does Not Depend on NANOG. Cells 2021, 10, 2232. https://doi.org/10.3390/cells10092232
Springer C, Zakhartchenko V, Wolf E, Simmet K. Hypoblast Formation in Bovine Embryos Does Not Depend on NANOG. Cells. 2021; 10(9):2232. https://doi.org/10.3390/cells10092232
Chicago/Turabian StyleSpringer, Claudia, Valeri Zakhartchenko, Eckhard Wolf, and Kilian Simmet. 2021. "Hypoblast Formation in Bovine Embryos Does Not Depend on NANOG" Cells 10, no. 9: 2232. https://doi.org/10.3390/cells10092232
APA StyleSpringer, C., Zakhartchenko, V., Wolf, E., & Simmet, K. (2021). Hypoblast Formation in Bovine Embryos Does Not Depend on NANOG. Cells, 10(9), 2232. https://doi.org/10.3390/cells10092232