
A Polymorphism in C-C Chemokine Receptor 5 (CCR5) Associates with Löfgren’s Syndrome and Alters Receptor Expression as well as Functional Response

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Genotyping
2.3. Flow Cytometry
2.3.1. CCR5 Expression on Peripheral Blood Monocytes
2.3.2. Ca-Influx Assay
2.4. Quantification of Changes in [Ca2+]i after Stimulation
b. fMLP (AUC/time)—Baseline fMLP (AUC/time)
2.5. Statistical Analysis
3. Results
3.1. Genotyping
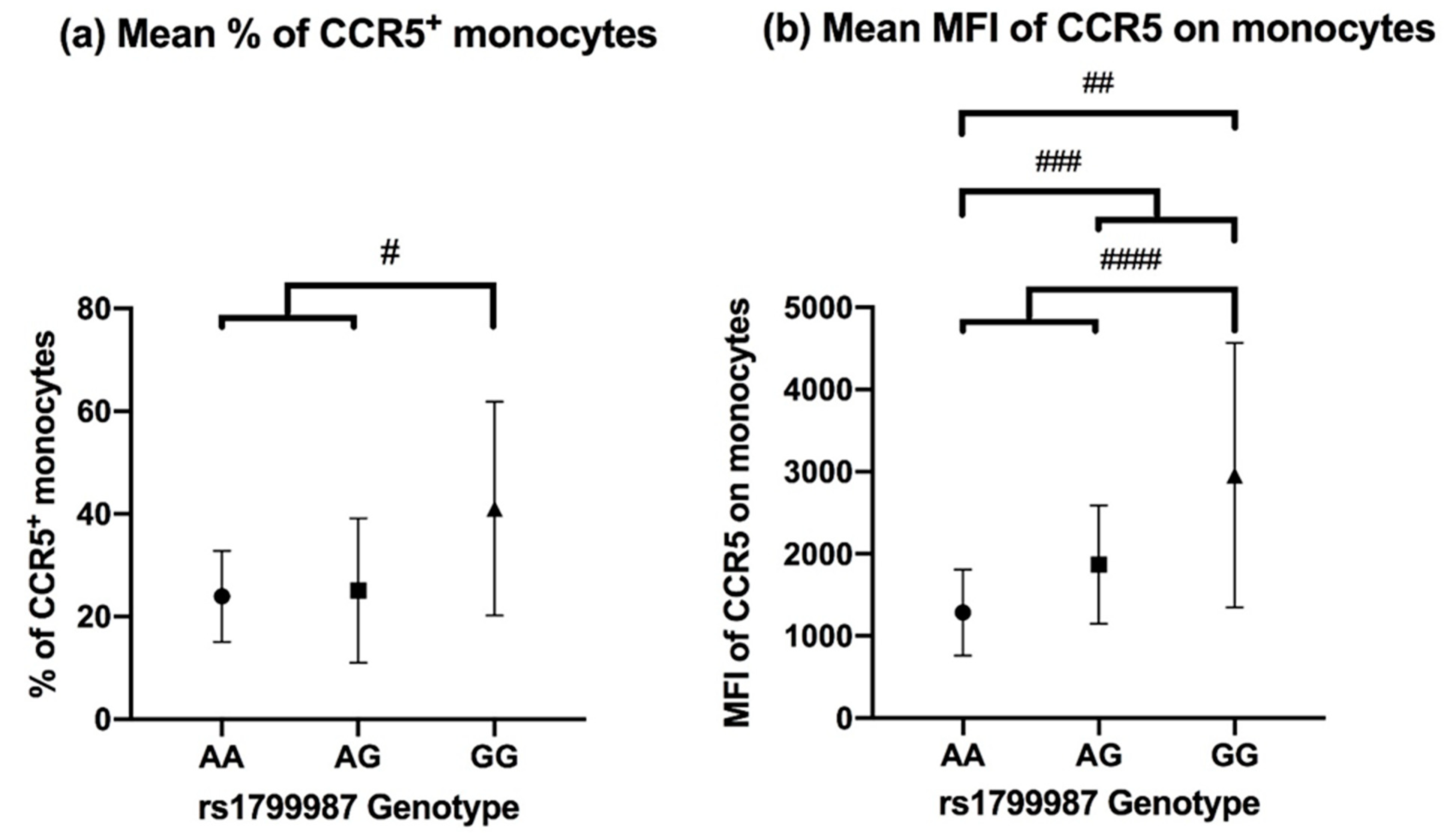
3.2. CCR5 Expression on Peripheral Blood Monocytes
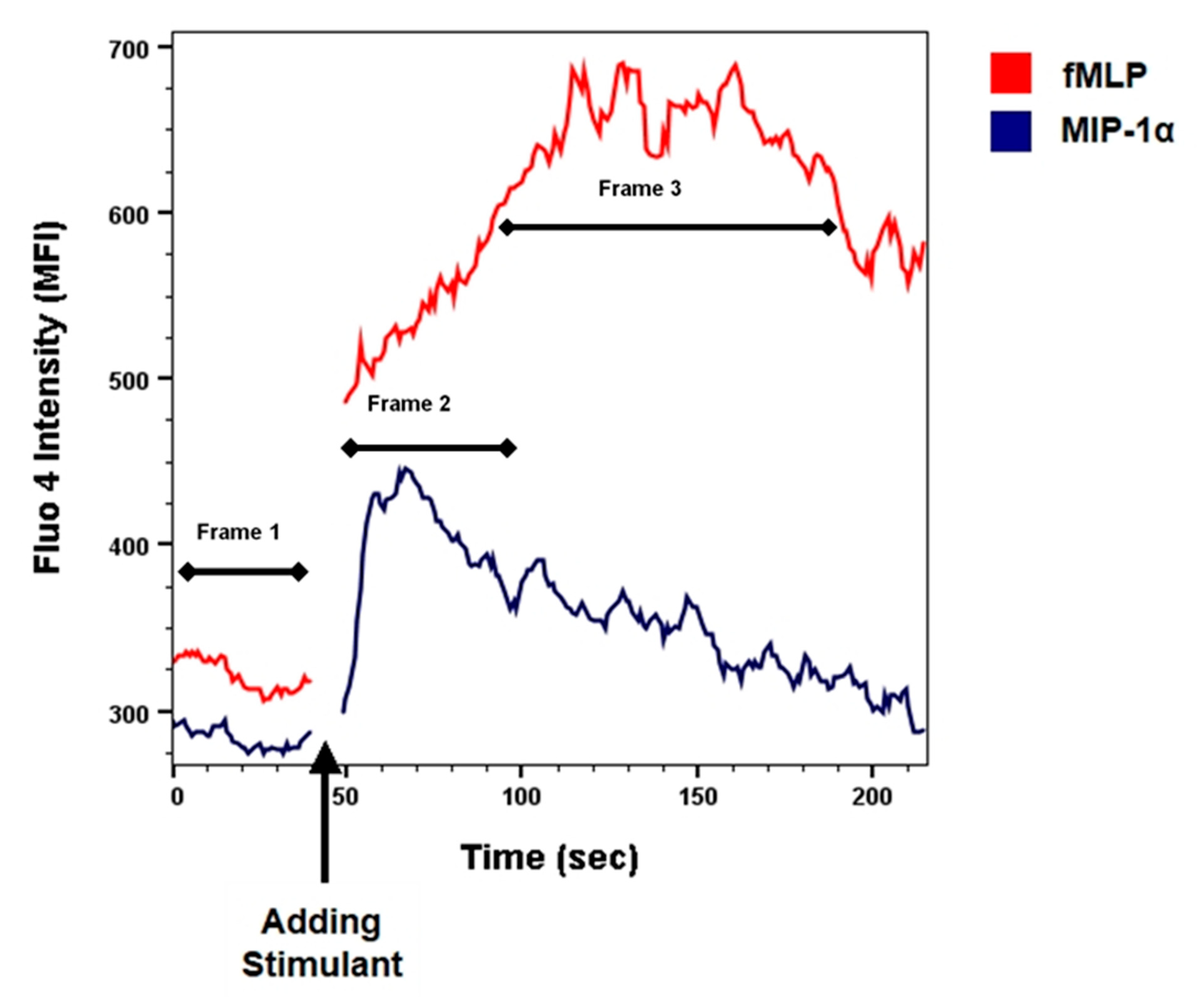
3.3. CCR5 Induced Calcium Mobilization Response in Monocytes
3.4. Kinetic Analysis of Changes in [Ca2+]i after Stimulation
3.5. Quantification of Changes in [Ca2+]i after Stimulation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grunewald, J.; Grutters, J.C.; Arkema, E.V.; Saketkoo, L.A.; Moller, D.R.; Müller-Quernheim, J. Sarcoidosis. Nat. Rev. Dis. Prim. 2019, 5, 1–22. [Google Scholar] [CrossRef]
- Petrek, M.; Pantelidis, P.; Southcott, A.M.; Lympany, P.; Safranek, P.; Black, C.M.; Kolek, V.; Weigl, E.; Du Bois, R.M. The source and role of RANTES in interstitial lung disease. Eur. Respir. J. 1997, 10, 1207–1216. [Google Scholar] [CrossRef]
- Petrek, M. CC chemokine receptor 5 (CCR5) mRNA expression in pulmonary sarcoidosis. Immunol. Lett. 2002, 80, 189–193. [Google Scholar] [CrossRef]
- Bacon, K.; Baggiolini, M.; Broxmeyer, H.; Horuk, R.; Lindley, I.; Mantovani, A.; Matsushima, K.; Murphy, P.; Nomiyama, H.; Oppenheim, J.; et al. Chemokine/chemokine receptor nomenclature. Cytokine 2003, 21, 48–49. [Google Scholar] [CrossRef]
- Lee, Y.H.; Bae, S.C.; Song, G.G. Association between the chemokine receptor 5 delta32 polymorphism and rheumatoid arthritis: A meta-analysis. Mod. Rheumatol. 2013, 23, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, L.; Tagliamonte, M.; Gauzzi, M.C.; Lopalco, L. Dual CCR5/CCR2 targeting: Opportunities for the cure of complex disorders. Cell. Mol. Life Sci. 2019, 76, 4869–4886. [Google Scholar] [CrossRef]
- Qidwai, T. Chemokine genetic polymorphism in human health and disease. Immunol. Lett. 2016, 176, 128–138. [Google Scholar] [CrossRef]
- Zimmerman, P.A.; Buckler-White, A.; Alkhatib, G.; Spalding, T.; Kubofcik, J.; Combadiere, C.; Weissman, D.; Cohen, O.; Rubbert, A.; Lam, G.; et al. Inherited resistance to HIV-1 conferred by an inactivating mutation in CC chemokine receptor 5: Studies in populations with contrasting clinical phenotypes, defined racial background, and quantified risk. Mol. Med. 1997, 3, 23–36. [Google Scholar] [CrossRef] [PubMed]
- McDermott, D.H.; Zimmerman, P.A.; Guignard, F.; Kleeberger, C.A.; Leitman, S.F.; Murphy, P.M. CCR5 promoter polymorphism and HIV-1 disease progression. Lancet 1998, 352, 866–870. [Google Scholar] [CrossRef]
- Zissel, G.; Prasse, A.; Müller-Quernheim, J. Sarcoidosis-Immunopathogenetic concepts. Semin. Respir. Crit. Care Med. 2007, 28, 3–14. [Google Scholar] [CrossRef]
- Spagnolo, P.; Renzoni, E.A.; Wells, A.U.; Copley, S.J.; Desai, S.R.; Sato, H.; Grutters, J.C.; Abdallah, A.; Taegtmeyer, A.; Du Bois, R.M.; et al. C-C chemokine receptor 5 gene variants in relation to lung disease in sarcoidosis. Am. J. Respir. Crit. Care Med. 2005, 172, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Valentonyte, R.; Nebel, A.; Nothnagel, M.; Müller-Quernheim, J.; Schürmann, M.; Schreiber, S. Female-specific association of C-C chemokine receptor 5 gene polymorphisms with Löfgren’s syndrome. J. Mol. Med. 2008, 86, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Karakaya, B.; Kaiser, Y.; Grunewald, J.; van Moorsel, C. Löfgren’s Syndrome: Diagnosis, Management, and Disease Pathogenesis. Semin. Respir. Crit. Care Med. 2017, 38, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Achour, L.; Scott, M.G.H.; Shirvani, H.; Thuret, A.; Bismuth, G.; Labbé-Jullié, C.; Marullo, S. CD4-CCR5 interaction in intracellular compartments contributes to receptor expression at the cell surface. Blood 2009, 113, 1938–1947. [Google Scholar] [CrossRef]
- Mehlotra, R.K. CCR5 Promoter Polymorphism -2459G > A: Forgotten or Ignored? Cells 2019, 8, 651. [Google Scholar] [CrossRef]
- Statement on Sarcoidosis. Joint Statement of the American Thoracic Society (ATS), the European Respiratory Society (ERS) and the World Association of Sarcoidosis and Other Granulomatous Disorders (WASOG) adopted by the ATS Board of Directors and by the ER. Am. J. Respir. Crit. Care Med. 1999, 160, 736–755. [Google Scholar] [CrossRef]
- Hijdra, D.; Vorselaars, A.D.M.; Grutters, J.C.; Claessen, A.M.E.; Rijkers, G.T. Phenotypic Characterization of Human Intermediate Monocytes. Front. Immunol. 2013, 4, 4–6. [Google Scholar] [CrossRef]
- Sato, H.; Silveira, L.; Spagnolo, P.; Gillespie, M.; Gottschall, E.B.; Welsh, K.I.; Du Bois, R.M.; Newman, L.S.; Maier, L.A. CC chemokine receptor 5 gene polymorphisms in beryllium disease. Eur. Respir. J. 2010, 36, 331–338. [Google Scholar] [CrossRef]
- Mamtani, M.; Rovin, B.; Brey, R.; Camargo, J.F.; Kulkarni, H.; Herrera, M.; Correa, P.; Holliday, S.; Anaya, J.M.; Ahuja, S.K. CCL3L1 gene-containing segmental duplications and polymorphisms in CCR5 affect risk of systemic lupus erythaematosus. Ann. Rheum. Dis. 2008, 67, 1076–1083. [Google Scholar] [CrossRef]
- Han, S.W.; Sa, K.H.; Kim, S.I.; Lee, S.I.; Park, Y.W.; Lee, S.S.; Yoo, W.H.; Soe, J.S.; Nam, E.J.; Lee, J.; et al. CCR5 gene polymorphism is a genetic risk factor for radiographic severity of rheumatoid arthritis. Tissue Antigens 2012, 80, 416–423. [Google Scholar] [CrossRef]
- Eri, R.; Jonsson, J.R.; Pandeya, N.; Purdie, D.M.; Clouston, A.D.; Martin, N.; Duffy, D.; Powell, E.E.; Fawcett, J.; Florin, T.H.J.; et al. CCR5-Δ32 mutation is strongly associated with primary sclerosing cholangitis. Genes Immun. 2004, 5, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Shieh, B.; Liau, Y.E.; Hsieh, P.S.; Yan, Y.P.; Wang, S.T.; Li, C. Influence of nucleotide polymorphisms in the CCR2 gene and the CCR5 promoter on the expression of cell surface CCR5 and CXCR4. Int. Immunol. 2000, 12, 1311–1318. [Google Scholar] [CrossRef]
- Salkowitz, J.R.; Bruse, S.E.; Meyerson, H.; Valdez, H.; Mosier, D.E.; Harding, C.V.; Zimmerman, P.A.; Lederman, M.M. CCR5 promoter polymorphism determines macrophage CCR5 density and magnitude of HIV-1 propagation in vitro. Clin. Immunol. 2003, 108, 234–240. [Google Scholar] [CrossRef]
- Bleul, C.C.; Wu, L.; Hoxie, J.A.; Springer, T.A.; Mackay, C.R. The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regulated on human T lymphocytes. Proc. Natl. Acad. Sci. USA 1997, 94, 1925–1930. [Google Scholar] [CrossRef]
- Hladik, F.; Liu, H.; Speelmon, E.; Livingston-Rosanoff, D.; Wilson, S.; Sakchalathorn, P.; Hwangbo, Y.; Greene, B.; Zhu, T.; McElrath, M.J. Combined Effect of CCR5-Δ32 Heterozygosity and the CCR5 Promoter Polymorphism −2459 A/G on CCR5 Expression and Resistance to Human Immunodeficiency Virus Type 1 Transmission. J. Virol. 2005, 79, 11677–11684. [Google Scholar] [CrossRef][Green Version]
- Wu, L.; Paxton, W.A.; Kassam, N.; Ruffing, N.; Rottman, J.B.; Sullivan, N.; Choe, H.; Sodroski, J.; Newman, W.; Koup, R.A.; et al. CCR5 levels and expression pattern correlate with infectability by macrophage-tropic HIV-1, in vitro. J. Exp. Med. 1997, 185, 1681–1691. [Google Scholar] [CrossRef]
- Tuttle, D.L.; Harrison, J.K.; Anders, C.; Sleasman, J.W.; Goodenow, M.M. Expression of CCR5 Increases during Monocyte Differentiation and Directly Mediates Macrophage Susceptibility to Infection by Human Immunodeficiency Virus Type 1. J. Virol. 1998, 72, 4962–4969. [Google Scholar] [CrossRef]
- Petrek, M.; Drabek, J.; Kolek, V.; Zlamal, J.; Welsh, K.I.; Bunce, M.; Weigl, E.; Du Bois, R.M. CC chemokine receptor gene polymorphisms in Czech patients with pulmonary sarcoidosis. Am. J. Respir. Crit. Care Med. 2000, 162, 1000–1003. [Google Scholar] [CrossRef]
- Palchevskiy, V.; Hashemi, N.; Weigt, S.S.; Xue, Y.Y.; Derhovanessian, A.; Keane, M.P.; Strieter, R.M.; Fishbein, M.C.; Deng, J.C.; Lynch, J.P.; et al. Immune response CC chemokines CCL2 and CCL5 are associated with pulmonary sarcoidosis. Fibrogenes. Tissue Repair 2011, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Oshima, M.; Maeda, A.; Ishioka, S.; Hiyama, K.; Yamakido, M. Expression of C-C chemokines in bronchoalveolar lavage cells from patients with granulomatous lung diseases. Lung 1999, 177, 229–240. [Google Scholar] [CrossRef]
- Katchar, K.; Eklund, A.; Grunewald, J. Expression of Th1 markers by lung accumulated T cells in pulmonary sarcoidosis. J. Intern. Med. 2003, 254, 564–571. [Google Scholar] [CrossRef]
- Iida, K.; Kadota, J.I.; Kawakami, K.; Matsubara, Y.; Shirai, R.; Kohno, S. Analysis of t cell subsets and β chemokines in patients with pulmonary sarcoidosis. Thorax 1997, 52, 431–437. [Google Scholar] [CrossRef]
- Molon, B.; Gri, G.; Bettella, M.; Gómez-Moutón, C.; Lanzavecchia, A.; Martínez-A, C.; Mañes, S.; Viola, A. T cell costimulation by chemokine receptors. Nat. Immunol. 2005, 6, 465–471. [Google Scholar] [CrossRef]
- Spagnolo, P.; Renzoni, E.A.; Wells, A.U.; Sato, H.; Grutters, J.C.; Sestini, P.; Abdallah, A.; Gramiccioni, E.; Ruven, H.J.T.; Du Bois, R.M.; et al. C-C Chemokine Receptor 2 and Sarcoidosis: Association with Löfgren’s Syndrome. Am. J. Respir. Crit. Care Med. 2003, 168, 1162–1166. [Google Scholar] [CrossRef]
- Goehringer, F.; Tiotiu, A.; Schvoerer, E.; Jaussaud, R.; Hoen, B. An unexpected effect of maraviroc could make it a future treatment for sarcoidosis. J. Investig. Allergol. Clin. Immunol. 2021, 31, 83–84. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Locati, M.; Vecchi, A.; Sozzani, S.; Allavena, P. Decoy receptors: A strategy to regulate inflammatory cytokines and chemokines. Trends Immunol. 2001, 22, 328–336. [Google Scholar] [CrossRef]
- Schepers, E.; Glorieux, G.; Dhondt, A.; Leybaert, L.; Vanholder, R. Flow cytometric calcium flux assay: Evaluation of cytoplasmic calcium kinetics in whole blood leukocytes. J. Immunol. Methods 2009, 348, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Taub, D.D.; Conlon, K.; Lloyd, A.R.; Oppenheim, J.J.; Kelvin, D.J. Preferential migration of activated CD4+ and CD8+ T cells in response to MIP-1α and MIP-1β. Science 1993, 260, 355–358. [Google Scholar] [CrossRef]
- Castellino, F.; Huang, A.Y.; Altan-Bonnet, G.; Stoll, S.; Scheinecker, C.; Germain, R.N. Chemokines enhance immunity by guiding naive CD8+ T cells to sites of CD4+ T cell-dendritic cell interaction. Nature 2006, 440, 890–895. [Google Scholar] [CrossRef]
- Capelli, A.; Stefano, A.D.I.; Lusuardi, M.; Gnemmi, I.; Donner, C.F. Increased macrophage inflammatory protein-1α and macrophage inflammatory protein-1β levels in bronchoalveolar lavage fluid of patients affected by different stages of pulmonary sarcoidosis. Am. J. Respir. Crit. Care Med. 2002, 165, 236–241. [Google Scholar] [CrossRef]
- Ziegenhagen, M.W.; Schrum, S.; Zissel, G.; Zipfel, P.F.; Schlaak, M.; Müller-Quernheim, J. Increased expression of proinflammatory chemokines in bronchoalveolar lavage cells of patients with progressing idiopathic pulmonary fibrosis and sarcoidosis. J. Investig. Med. 1998, 46, 223–231. [Google Scholar]
- Standiford, T.J.; Rolfe, M.W.; Kunkel, S.L.; Lynch, J.P.; Burdick, M.D.; Gilbert, A.R.; Orringer, M.B.; Whyte, R.I.; Strieter, R.M. Macrophage inflammatory protein-1 alpha expression in interstitial lung disease. J. Immunol. 1993, 151, 2852–2863. [Google Scholar]
- Hashimoto, S.; Nakayama, T.; Gon, Y.; Hata, N.; Koura, T.; Maruoka, S.; Matsumoto, K.; Hayashi, S.; Abe, Y.; Horie, T. Correlation of plasma monocyte chemoattractant protein-1 (MCP-1) and monocyte inflammatory protein-1α (MIP-1α) levels with disease activity and clinical course of sarcoidosis. Clin. Exp. Immunol. 1998, 111, 604–610. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| CCR5 rs1799987 | Controls (n = 257) | Löfgren’s Syndrome (n = 106) * |
|---|---|---|
| A (%) | 299 (58%) | 96 (45%) |
| G (%) | 215 (42%) | 116 (55%) |
| AA (%) | 92 (36%) | 21 (20%) |
| AG (%) | 115 (45%) | 54 (51%) |
| GG (%) | 50 (19%) | 31 (29%) |
| Stimulation with MIP-1α | ||
|---|---|---|
| Genotype | Increase in [Ca2+]i * | No Increase in [Ca2+]i |
| AA | 6 | 1 |
| AG | 4 | 3 |
| GG | 1 | 6 |
| Total | 11 | 10 |
| Genotype | MIP-1α * | MIP-1α/fMLP ** |
|---|---|---|
| AA | 23.25 | 0.38 |
| AG | 16.35 | 0.24 |
| GG | 5.35 | 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karakaya, B.; van Moorsel, C.H.M.; Veltkamp, M.; Roodenburg-Benschop, C.; Kazemier, K.M.; van der Helm-van Mil, A.H.M.; Huizinga, T.W.J.; Grutters, J.C.; Rijkers, G.T. A Polymorphism in C-C Chemokine Receptor 5 (CCR5) Associates with Löfgren’s Syndrome and Alters Receptor Expression as well as Functional Response. Cells 2021, 10, 1967. https://doi.org/10.3390/cells10081967
Karakaya B, van Moorsel CHM, Veltkamp M, Roodenburg-Benschop C, Kazemier KM, van der Helm-van Mil AHM, Huizinga TWJ, Grutters JC, Rijkers GT. A Polymorphism in C-C Chemokine Receptor 5 (CCR5) Associates with Löfgren’s Syndrome and Alters Receptor Expression as well as Functional Response. Cells. 2021; 10(8):1967. https://doi.org/10.3390/cells10081967
Chicago/Turabian StyleKarakaya, Bekir, Coline H. M. van Moorsel, Marcel Veltkamp, Claudia Roodenburg-Benschop, Karin M. Kazemier, Annette H. M. van der Helm-van Mil, Tom W. J. Huizinga, Jan C. Grutters, and Ger T. Rijkers. 2021. "A Polymorphism in C-C Chemokine Receptor 5 (CCR5) Associates with Löfgren’s Syndrome and Alters Receptor Expression as well as Functional Response" Cells 10, no. 8: 1967. https://doi.org/10.3390/cells10081967
APA StyleKarakaya, B., van Moorsel, C. H. M., Veltkamp, M., Roodenburg-Benschop, C., Kazemier, K. M., van der Helm-van Mil, A. H. M., Huizinga, T. W. J., Grutters, J. C., & Rijkers, G. T. (2021). A Polymorphism in C-C Chemokine Receptor 5 (CCR5) Associates with Löfgren’s Syndrome and Alters Receptor Expression as well as Functional Response. Cells, 10(8), 1967. https://doi.org/10.3390/cells10081967