
Lysine 53 Acetylation of Cytochrome c in Prostate Cancer: Warburg Metabolism and Evasion of Apoptosis

 , ,
, ,  ,
, 
Abstract
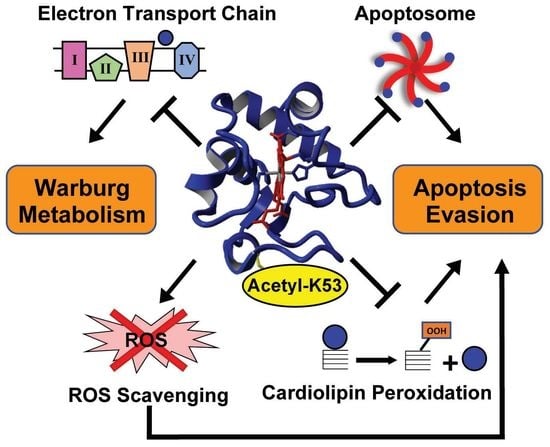
1. Introduction
2. Methods
2.1. Prostate Cancer Xenografts and Mass Spectrometry
2.2. Immunoprecipitation and Western Blotting
2.3. Generation of Cytc Variants
2.4. Bacterial Overexpression and Purification of Cytc Variants
2.5. Determination of the Concentration of Cytc Variants
2.6. COX Activity Measurement
2.7. Caspase-3 Activity Measurement
2.8. Rate of Oxidation Measurement
2.9. Rate of Reduction by Superoxide
2.10. Cardiolipin Peroxidase Activity Measurement
2.11. Redox Potential Measurement
2.12. Crystallization of K53Q Cytc
2.13. Statistical Analyses
2.14. Molecular Dynamics
3. Results
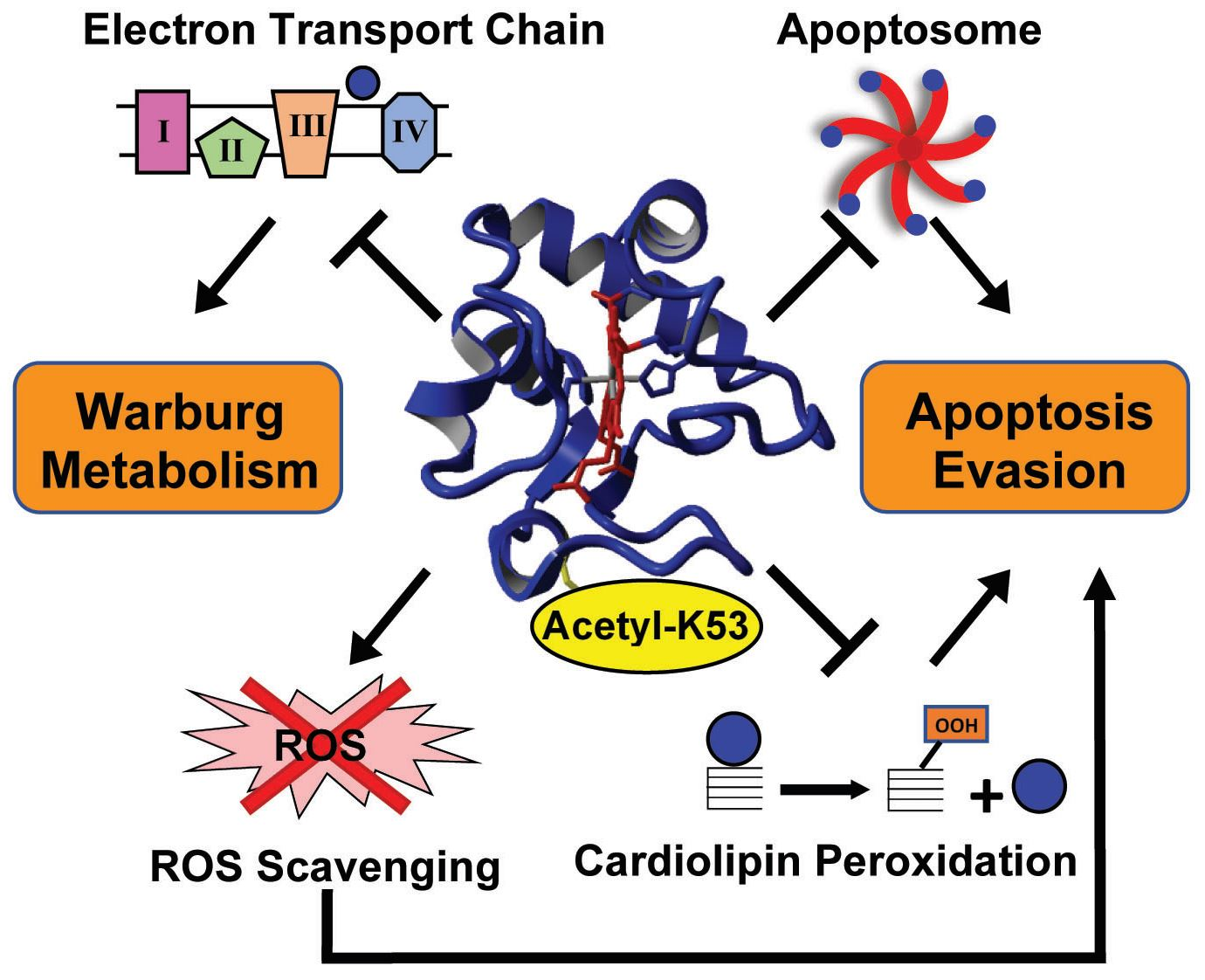
3.1. Cytc Is Acetylated on Lysine 53 in Castrate-Resistant and Castrate-Sensitive Human Prostate Cancer Xenografts
3.2. Purification of WT, Acetylmimetic, and Additional Cytc Control Variants
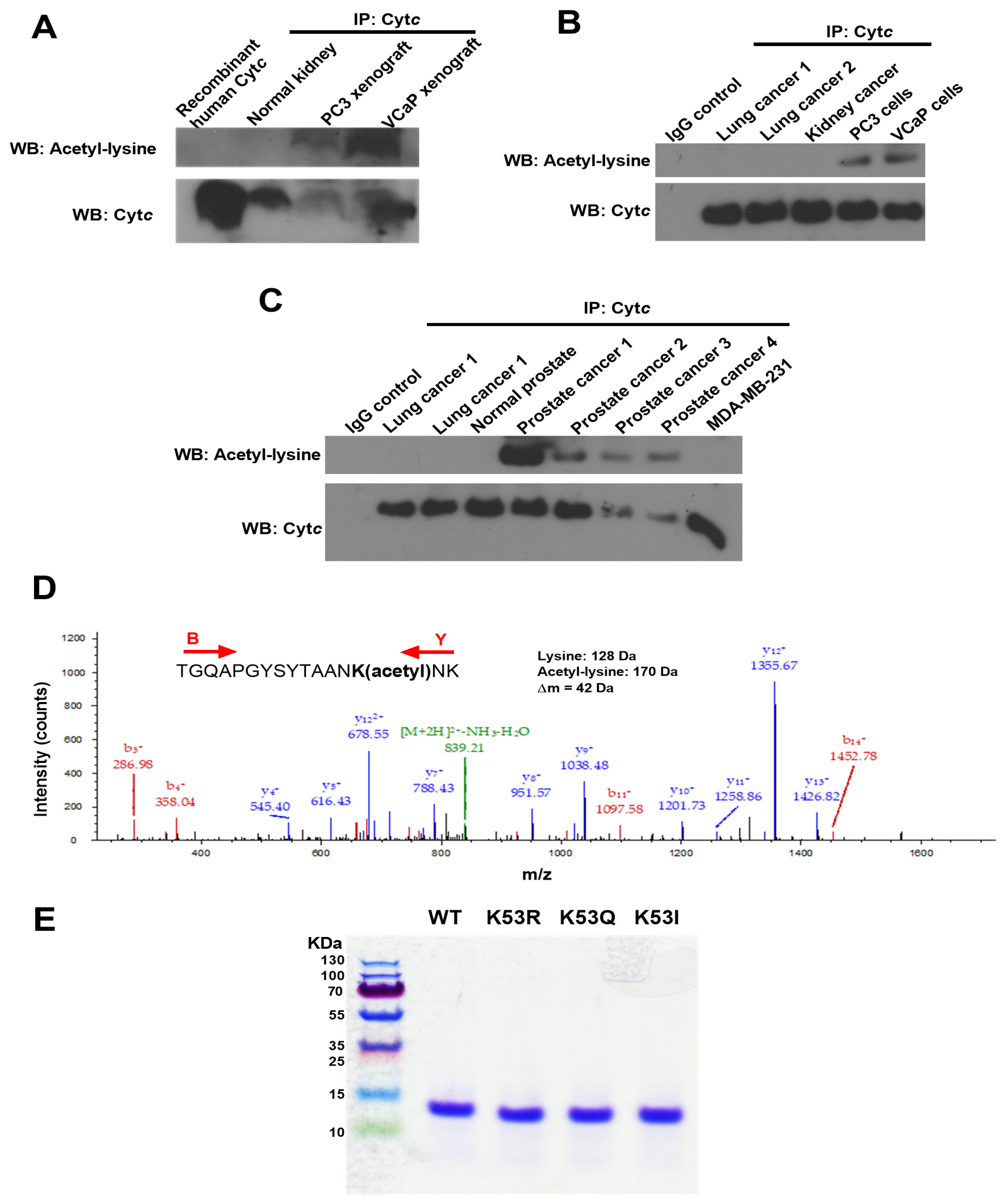
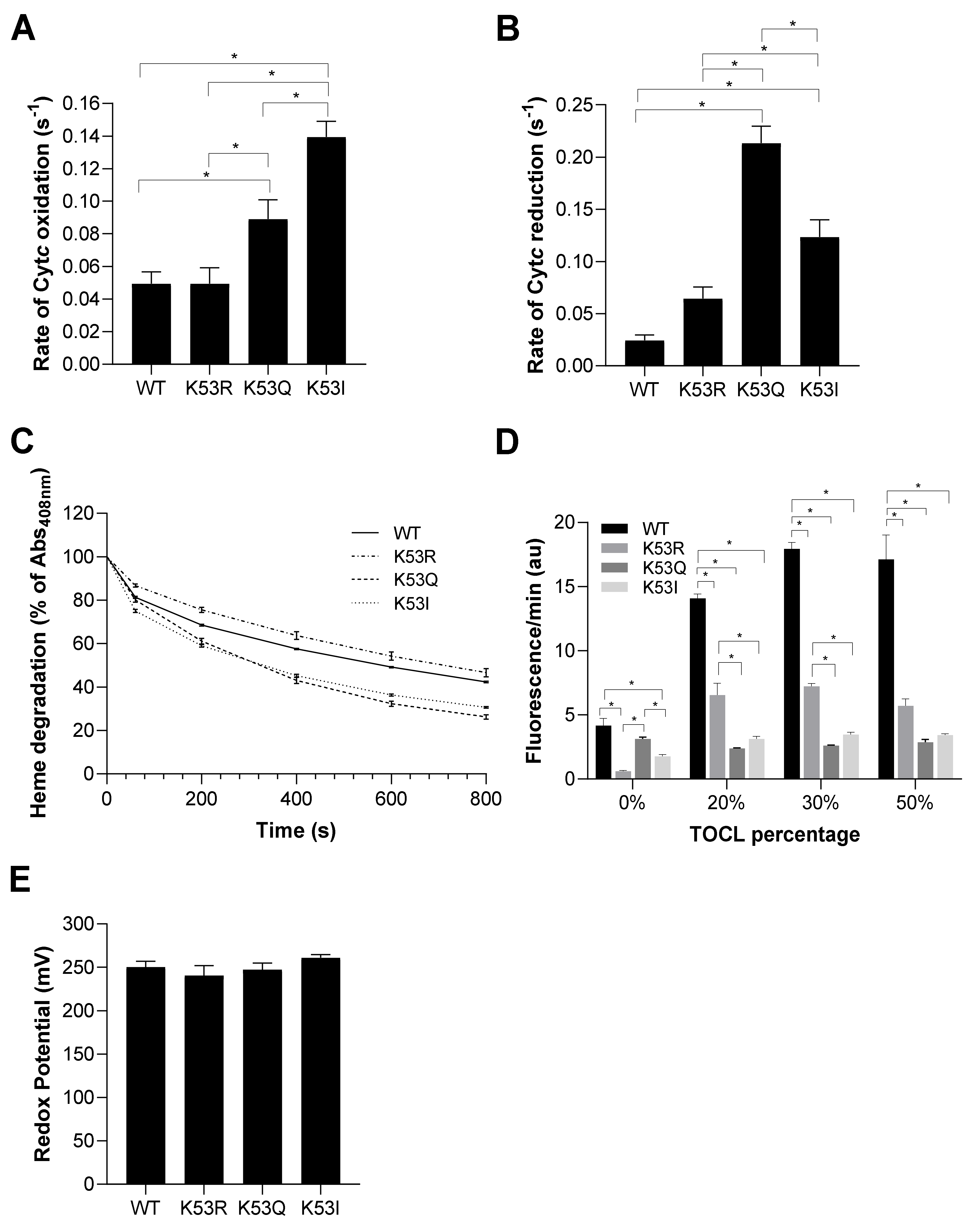
3.3. K53Q Acetylmimetic Cytc Results in Reduced Cytochrome c Oxidase and Caspase 3 Activity Compared to WT
3.4. Acetylmimetic Cytc Is a Superior ROS Scavenger
3.5. Acetylmimetic Cytc Is Less Stable at High H2O2 Concentrations and Shows Profoundly Reduced Cardiolipin Peroxidase Activity
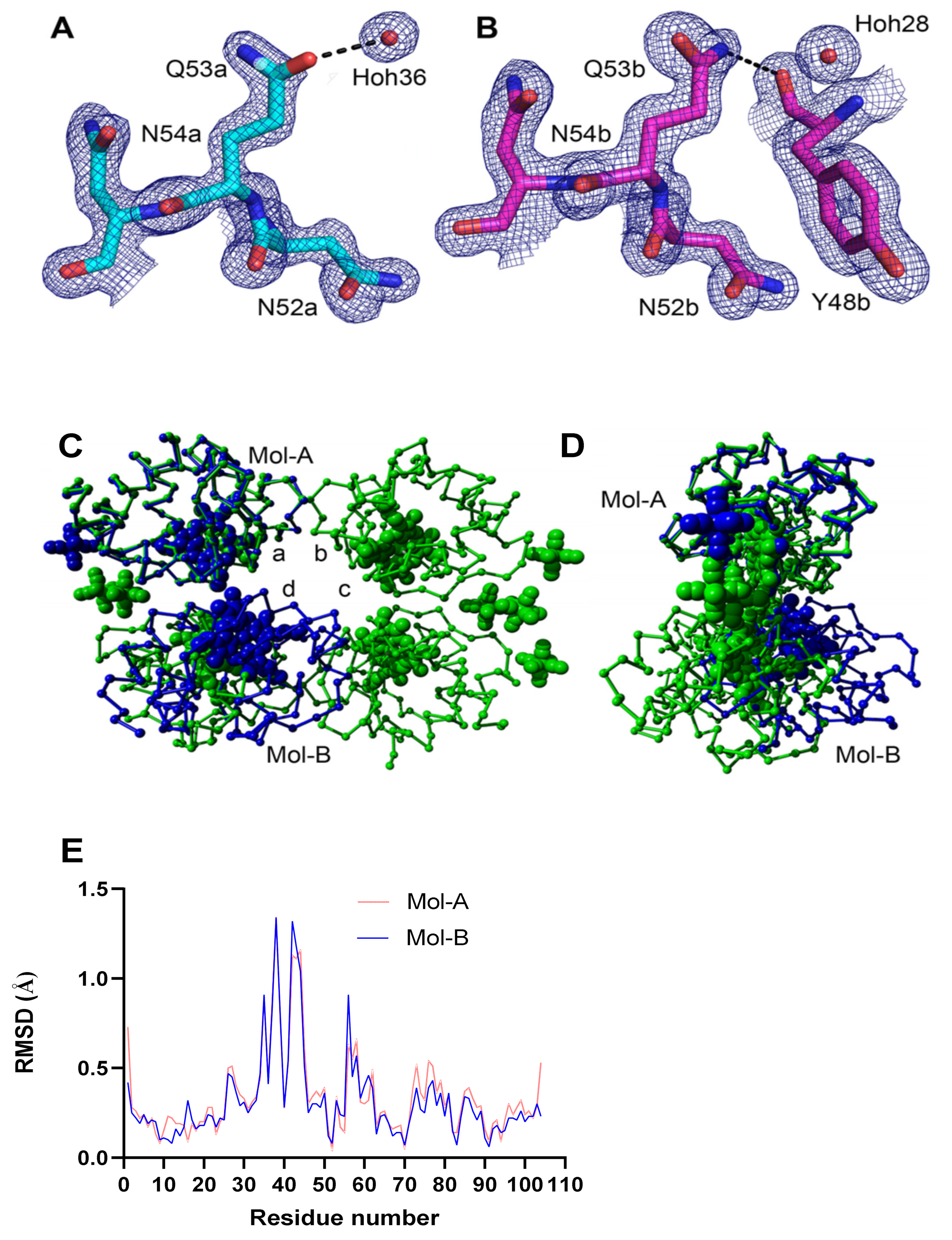
3.6. Crystal Structure of K53Q Cytc at a Resolution of 1.31 Å
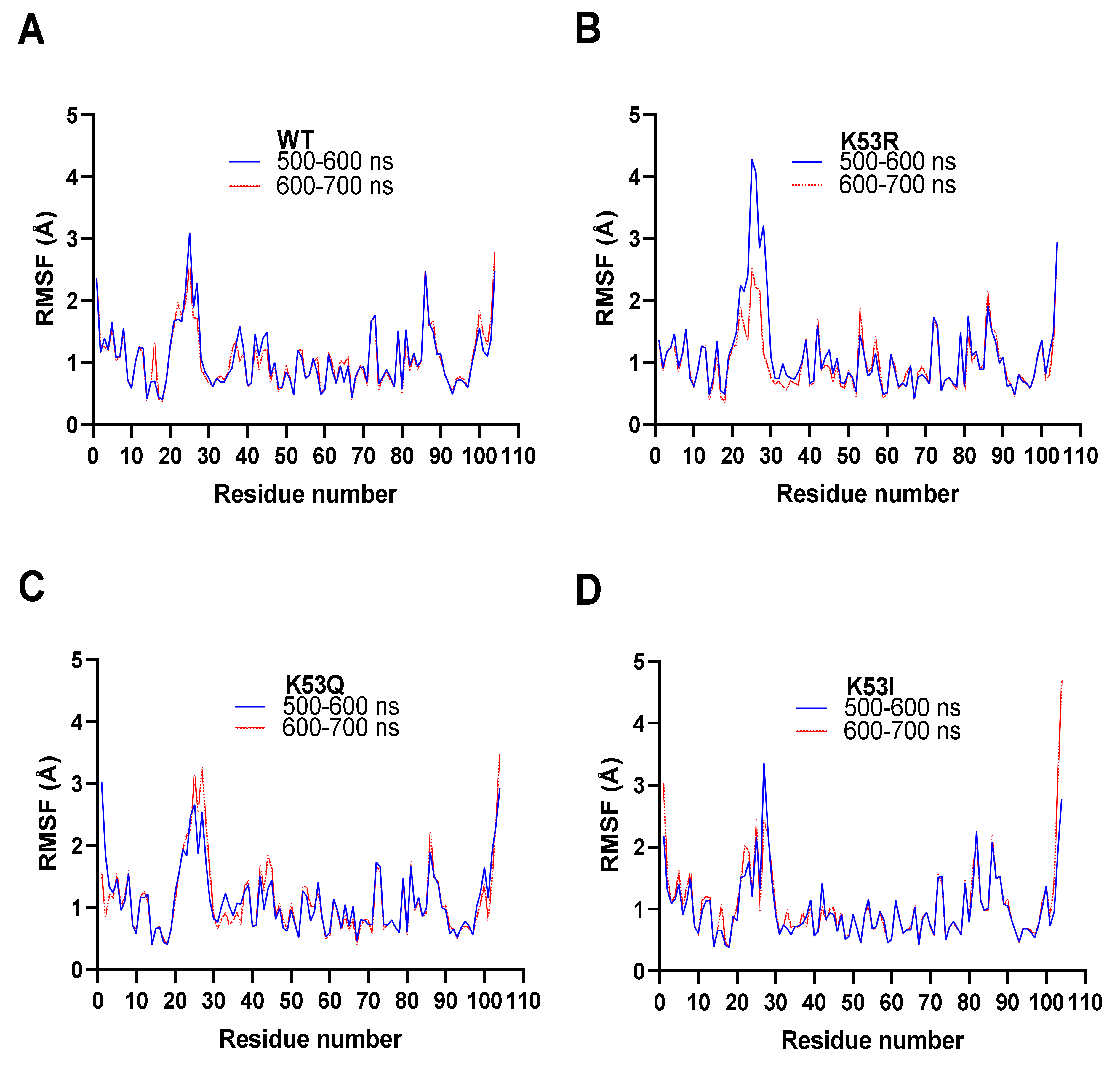
3.7. Molecular Dynamics Simulations
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| COX | cytochrome c oxidase |
| Cytc | cytochrome c |
| ETC | electron transport chain |
| OxPhos | oxidative phosphorylation |
| ROS | reactive oxygen species |
References
- Hüttemann, M.; Pecina, P.; Rainbolt, M.; Sanderson, T.H.; Kagan, V.E.; Samavati, L.; Doan, J.W.; Lee, I. The multiple functions of cytochrome c and their regulation in life and death decisions of the mammalian cell: From respiration to apoptosis. Mitochondrion 2011, 11, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Kalpage, H.A.; Bazylianska, V.; Recanati, M.A.; Fite, A.; Liu, J.; Wan, J.; Mantena, N.; Malek, M.H.; Podgorski, I.; Heath, E.I.; et al. Tissue-specific regulation of cytochrome c by post-translational modifications: Respiration, the mitochondrial membrane potential, ROS, and apoptosis. Faseb J. 2019, 33, 1540–1553. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Bayir, H.A.; Belikova, N.A.; Kapralov, O.; Tyurina, Y.Y.; Tyurin, V.A.; Jiang, J.; Stoyanovsky, D.A.; Wipf, P.; Kochanek, P.M.; et al. Cytochrome c/cardiolipin relations in mitochondria: A kiss of death. Free Radic. Biol. Med. 2009, 46, 1439–1453. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Tyurin, V.A.; Jiang, J.; Tyurina, Y.Y.; Ritow, V.B.; Amoscato, A.A.; Osipov, A.N.; Belikova, N.A.; Kapralov, A.A.; Kini, V.; et al. Cytochrome c acts as a cardiolipin oxygenase required for release of proapoptotic factors. Nat. Chem. Biol. 2005, 1, 223–232. [Google Scholar] [CrossRef]
- Vladimirov, Y.A.; Proskurnina, E.V.; Izmailov, D.Y.; Novikov, A.A.; Brusnichkin, A.V.; Osipov, A.N.; Kagan, V.E. Cardiolipin activates cytochrome c peroxidase activity since it facilitates H(2)O(2) access to heme. Biochemistry 2006, 71, 998–1005. [Google Scholar] [CrossRef] [PubMed]
- Hüttemann, M.; Doan, J.W.; Goustin, A.S.; Sinkler, C.; Mahapatra, G.; Shay, J.; Liu, J.; Elbaz, H.A.; Aras, S.; Grossman, L.I.; et al. Regulation of cytochrome c in respiration, apoptosis, neurodegeneration and cancer: The good, the bad and the ugly. In Cytochromes b and c; Thom, R., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2014; pp. 1–38. [Google Scholar]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Lee, I.; Salomon, A.R.; Yu, K.; Doan, J.W.; Grossman, L.I.; Hüttemann, M. New prospects for an old enzyme: Mammalian cytochrome c is tyrosine-phosphorylated in vivo. Biochemistry 2006, 45, 9121–9128. [Google Scholar] [CrossRef]
- Yu, H.; Lee, I.; Salomon, A.R.; Yu, K.; Hüttemann, M. Mammalian liver cytochrome c is tyrosine-48 phosphorylated in vivo, inhibiting mitochondrial respiration. Biochim. Biophys. Acta 2008, 1777, 1066–1071. [Google Scholar] [CrossRef]
- Mahapatra, G.; Varughese, A.; Ji, Q.; Lee, I.; Liu, J.; Vaishnav, A.; Sinkler, C.; Kapralov, A.A.; Moraes, C.T.; Sanderson, T.H.; et al. Phosphorylation of Cytochrome c Threonine 28 Regulates Electron Transport Chain Activity in Kidney: IMPLICATIONS FOR AMP KINASE. J. Biol. Chem. 2017, 292, 64–79. [Google Scholar] [CrossRef]
- Wan, J.; Kalpage, H.A.; Vaishnav, A.; Liu, J.; Lee, I.; Mahapatra, G.; Turner, A.A.; Zurek, M.P.; Ji, Q.; Moraes, C.T.; et al. Regulation of Respiration and Apoptosis by Cytochrome c Threonine 58 Phosphorylation. Sci. Rep. 2019, 9, 15815. [Google Scholar] [CrossRef]
- Kalpage, H.A.; Vaishnav, A.; Liu, J.; Varughese, A.; Wan, J.; Turner, A.A.; Ji, Q.; Zurek, M.P.; Kapralov, A.A.; Kagan, V.E.; et al. Serine-47 phosphorylation of cytochrome c in the mammalian brain regulates cytochrome c oxidase and caspase-3 activity. Faseb J. 2019, 33, 13503–13514. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhao, D.; Spring, D.J.; DePinho, R.A. Genetics and biology of prostate cancer. Genes Dev. 2018, 32, 1105–1140. [Google Scholar] [CrossRef]
- Peitzsch, C.; Gorodetska, I.; Klusa, D.; Shi, Q.; Alves, T.C.; Pantel, K.; Dubrovska, A. Metabolic regulation of prostate cancer heterogeneity and plasticity. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef]
- Chaudhary, A.K.; O’Malley, J.; Kumar, S.; Inigo, J.R.; Kumar, R.; Yadav, N.; Chandra, D. Mitochondrial dysfunction and prostate cancer racial disparities among American men. Front. Biosci. (Schol. Ed.) 2017, 9, 154–164. [Google Scholar] [CrossRef]
- Perkins, D.N.; Pappin, D.J.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Pecina, P.; Borisenko, G.G.; Belikova, N.A.; Tyurina, Y.Y.; Pecinova, A.; Lee, I.; Samhan-Arias, A.K.; Przyklenk, K.; Kagan, V.E.; Hüttemann, M. Phosphomimetic substitution of cytochrome c tyrosine 48 decreases respiration and binding to cardiolipin and abolishes ability to trigger downstream caspase activation. Biochemistry 2010, 49, 6705–6714. [Google Scholar] [CrossRef]
- Slee, E.A.; Harte, M.T.; Kluck, R.M.; Wolf, B.B.; Casiano, C.A.; Newmeyer, D.D.; Wang, H.G.; Reed, J.C.; Nicholson, D.W.; Alnemri, E.S.; et al. Ordering the cytochrome c-initiated caspase cascade: Hierarchical activation of caspases-2, -3, -6, -7, -8, and -10 in a caspase-9-dependent manner. J. Cell Biol. 1999, 144, 281–292. [Google Scholar] [CrossRef]
- Liu, Z.; Lin, H.; Ye, S.; Liu, Q.Y.; Meng, Z.; Zhang, C.M.; Xia, Y.; Margoliash, E.; Rao, Z.; Liu, X.J. Remarkably high activities of testicular cytochrome c in destroying reactive oxygen species and in triggering apoptosis. Proc. Natl. Acad. Sci. USA 2006, 103, 8965–8970. [Google Scholar] [CrossRef]
- Quick, K.L.; Hardt, J.I.; Dugan, L.L. Rapid microplate assay for superoxide scavenging efficiency. J. Neurosci. Methods 2000, 97, 139–144. [Google Scholar] [CrossRef]
- Fridovich, I. Quantitative aspects of the production of superoxide anion radical by milk xanthine oxidase. J. Biol. Chem. 1970, 245, 4053–4057. [Google Scholar] [CrossRef]
- Cammack, R. Redox States and Potentials. In Bioenergetics—A Practical Approach; Cooper, G., Ed.; IRL Press: Oxford, UK, 1995; pp. 93–95. [Google Scholar]
- Kabsch, W. Xds. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef]
- Vonrhein, C.; Flensburg, C.; Keller, P.; Sharff, A.; Smart, O.; Paciorek, W.; Womack, T.; Bricogne, G. Data processing and analysis with the autoPROC toolbox. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 293–302. [Google Scholar] [CrossRef]
- Krieger, E.; Vriend, G. New ways to boost molecular dynamics simulations. J. Comput. Chem. 2015, 36, 996–1007. [Google Scholar] [CrossRef]
- Hornak, V.; Abel, R.; Okur, A.; Strockbine, B.; Roitberg, A.; Simmerling, C. Comparison of multiple Amber force fields and development of improved protein backbone parameters. Proteins 2006, 65, 712–725. [Google Scholar] [CrossRef]
- Zaidi, S.; Hassan, M.I.; Islam, A.; Ahmad, F. The role of key residues in structure, function, and stability of cytochrome-c. Cell Mol. Life Sci. 2014, 71, 229–255. [Google Scholar] [CrossRef]
- Lu, J.; Tan, M.; Cai, Q. The Warburg effect in tumor progression: Mitochondrial oxidative metabolism as an anti-metastasis mechanism. Cancer Lett. 2015, 356, 156–164. [Google Scholar] [CrossRef]
- Lee, I.; Salomon, A.R.; Yu, K.; Samavati, L.; Pecina, P.; Pecinova, A.; Hüttemann, M. Isolation of regulatory-competent, phosphorylated cytochrome c oxidase. Methods Enzymol. 2009, 457, 193–210. [Google Scholar]
- Roberts, V.A.; Pique, M.E. Definition of the interaction domain for cytochrome c on cytochrome c oxidase. III. Prediction of the docked complex by a complete, systematic search. J. Biol. Chem. 1999, 274, 38051–38060. [Google Scholar] [CrossRef]
- Cheng, T.C.; Hong, C.; Akey, I.V.; Yuan, S.; Akey, C.W. A near atomic structure of the active human apoptosome. Elife 2016, 5. [Google Scholar] [CrossRef]
- Korshunov, S.S.; Krasnikov, B.F.; Pereverzev, M.O.; Skulachev, V.P. The antioxidant functions of cytochrome c. FEBS Lett. 1999, 462, 192–198. [Google Scholar] [CrossRef]
- Wang, Z.B.; Li, M.; Zhao, Y.; Xu, J.X. Cytochrome c is a hydrogen peroxide scavenger in mitochondria. Protein Pept. Lett. 2003, 10, 247–253. [Google Scholar] [CrossRef]
- Pereverzev, M.O.; Vygodina, T.V.; Konstantinov, A.A.; Skulachev, V.P. Cytochrome c, an ideal antioxidant. Biochem. Soc. Trans. 2003, 31, 1312–1315. [Google Scholar] [CrossRef]
- Nicholls, D.G.; Ferguson, S.J. Bioenergetics 2; Academic Press Limited: London, UK; San Diego, CA, USA, 1992. [Google Scholar]
- Hosp, F.; Lassowskat, I.; Santoro, V.; De Vleesschauwer, D.; Fliegner, D.; Redestig, H.; Mann, M.; Christian, S.; Hannah, M.A.; Finkemeier, I. Lysine acetylation in mitochondria: From inventory to function. Mitochondrion 2017, 33, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Baeza, J.; Smallegan, M.J.; Denu, J.M. Mechanisms and Dynamics of Protein Acetylation in Mitochondria. Trends Biochem. Sci. 2016, 41, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Casey, J.R.; Grinstein, S.; Orlowski, J. Sensors and regulators of intracellular pH. Nat. Rev. Mol. Cell Biol. 2010, 11, 50–61. [Google Scholar] [CrossRef]
- Wagner, G.R.; Payne, R.M. Widespread and enzyme-independent Nε-acetylation and Nε-succinylation of proteins in the chemical conditions of the mitochondrial matrix. J. Biol. Chem. 2013, 288, 29036–29045. [Google Scholar] [CrossRef] [PubMed]
- Konig, A.C.; Hartl, M.; Boersema, P.J.; Mann, M.; Finkemeier, I. The mitochondrial lysine acetylome of Arabidopsis. Mitochondrion 2014, 19 Pt B, 252–260. [Google Scholar] [CrossRef]
- Kalpage, H.A.; Wan, J.; Morse, P.T.; Lee, I.; Hüttemann, M. Brain-Specific Serine-47 Modification of Cytochrome c Regulates Cytochrome c Oxidase Activity Attenuating ROS Production and Cell Death: Implications for Ischemia/Reperfusion Injury and Akt Signaling. Cells 2020, 9, 1843. [Google Scholar] [CrossRef]
- Sanderson, T.H.; Mahapatra, G.; Pecina, P.; Ji, Q.; Yu, K.; Sinkler, C.; Varughese, A.; Kumar, R.; Bukowski, M.J.; Tousignant, R.N.; et al. Cytochrome c is tyrosine 97 phosphorylated by neuroprotective insulin treatment. PLoS ONE 2013, 8, e78627. [Google Scholar] [CrossRef]
- Guerra-Castellano, A.; Diaz-Moreno, I.; Velazquez-Campoy, A.; De la Rosa, M.A.; Diaz-Quintana, A. Structural and functional characterization of phosphomimetic mutants of cytochrome c at threonine 28 and serine 47. Biochim. Biophys. Acta 2016. [Google Scholar] [CrossRef]
- Guerra-Castellano, A.; Diaz-Quintana, A.; Perez-Mejias, G.; Elena-Real, C.A.; Gonzalez-Arzola, K.; Garcia-Maurino, S.M.; De la Rosa, M.A.; Diaz-Moreno, I. Oxidative stress is tightly regulated by cytochrome c phosphorylation and respirasome factors in mitochondria. Proc. Natl. Acad. Sci. USA 2018, 115, 7955–7960. [Google Scholar] [CrossRef]
- Moreno-Beltran, B.; Guerra-Castellano, A.; Diaz-Quintana, A.; Del Conte, R.; Garcia-Maurino, S.M.; Diaz-Moreno, S.; Gonzalez-Arzola, K.; Santos-Ocana, C.; Velazquez-Campoy, A.; De la Rosa, M.A.; et al. Structural basis of mitochondrial dysfunction in response to cytochrome c phosphorylation at tyrosine 48. Proc. Natl. Acad. Sci. USA 2017, 114, E3041–E3050. [Google Scholar] [CrossRef]
- Garcia-Heredia, J.M.; Diaz-Quintana, A.; Salzano, M.; Orzaez, M.; Perez-Paya, E.; Teixeira, M.; De la Rosa, M.A.; Diaz-Moreno, I. Tyrosine phosphorylation turns alkaline transition into a biologically relevant process and makes human cytochrome c behave as an anti-apoptotic switch. J. Biol. Inorg. Chem. 2011, 16, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.S.; Thompson, C.B. Metabolic reprogramming: A cancer hallmark even warburg did not anticipate. Cancer Cell 2012, 21, 297–308. [Google Scholar] [CrossRef]
- Ngo, D.C.; Ververis, K.; Tortorella, S.M.; Karagiannis, T.C. Introduction to the molecular basis of cancer metabolism and the Warburg effect. Mol. Biol Rep. 2015, 42, 819–823. [Google Scholar] [CrossRef]
- Iwasaki, W.; Tachiwana, H.; Kawaguchi, K.; Shibata, T.; Kagawa, W.; Kurumizaka, H. Comprehensive structural analysis of mutant nucleosomes containing lysine to glutamine (KQ) substitutions in the H3 and H4 histone-fold domains. Biochemistry 2011, 50, 7822–7832. [Google Scholar] [CrossRef]
- Kalpage, H.A.; Wan, J.; Morse, P.T.; Zurek, M.P.; Turner, A.A.; Khobeir, A.; Yazdi, N.; Hakim, L.; Liu, J.; Vaishnav, A.; et al. Cytochrome c phosphorylation: Control of mitochondrial electron transport chain flux and apoptosis. Int. J. Biochem. Cell Biol. 2020, 121, 105704. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Z.B.; Xu, J.X. Effect of cytochrome c on the generation and elimination of O2*- and H2O2 in mitochondria. J. Biol. Chem. 2003, 278, 2356–2360. [Google Scholar] [CrossRef] [PubMed]
- Miriyala, S.; Spasojevic, I.; Tovmasyan, A.; Salvemini, D.; Vujaskovic, Z.; St Clair, D.; Batinic-Haberle, I. Manganese superoxide dismutase, MnSOD and its mimics. Biochim. Biophys. Acta 2012, 1822, 794–814. [Google Scholar] [CrossRef]
- Kagan, V.E.; Tyurina, Y.Y.; Bayir, H.; Chu, C.T.; Kapralov, A.A.; Vlasova, I.I.; Belikova, N.A.; Tyurin, V.A.; Amoscato, A.; Epperly, M.; et al. The “pro-apoptotic genies” get out of mitochondria: Oxidative lipidomics and redox activity of cytochrome c/cardiolipin complexes. Chem. Biol. Interact. 2006, 163, 15–28. [Google Scholar] [CrossRef]
- Eidelman, E.; Twum-Ampofo, J.; Ansari, J.; Siddiqui, M.M. The Metabolic Phenotype of Prostate Cancer. Front. Oncol. 2017, 7, 131. [Google Scholar] [CrossRef]
- Schöpf, B.; Weissensteiner, H.; Schäfer, G.; Fazzini, F.; Charoentong, P.; Naschberger, A.; Rupp, B.; Fendt, L.; Bukur, V.; Giese, I.; et al. OXPHOS remodeling in high-grade prostate cancer involves mtDNA mutations and increased succinate oxidation. Nat. Commun. 2020, 11, 1487. [Google Scholar] [CrossRef]
- Pecinová, A.; Alán, L.; Brázdová, A.; Vrbacký, M.; Pecina, P.; Drahota, Z.; Houštěk, J.; Mráček, T. Role of Mitochondrial Glycerol-3-Phosphate Dehydrogenase in Metabolic Adaptations of Prostate Cancer. Cells 2020, 9, 1764. [Google Scholar] [CrossRef] [PubMed]
- Diedrich, J.D.; Rajagurubandara, E.; Herroon, M.K.; Mahapatra, G.; Hüttemann, M.; Podgorski, I. Bone marrow adipocytes promote the warburg phenotype in metastatic prostate tumors via HIF-1alpha activation. Oncotarget 2016, 7, 64854–64877. [Google Scholar] [CrossRef] [PubMed]
- Vayalil, P.K.; Landar, A. Mitochondrial oncobioenergetic index: A potential biomarker to predict progression from indolent to aggressive prostate cancer. Oncotarget 2015, 6, 43065–43080. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, T.; Verdone, J.E.; Huang, J.; Kahlert, U.D.; Hernandez, J.R.; Torga, G.; Zarif, J.C.; Epstein, T.; Gatenby, R.; McCartney, A.; et al. Glycolysis is the primary bioenergetic pathway for cell motility and cytoskeletal remodeling in human prostate and breast cancer cells. Oncotarget 2015, 6, 130–143. [Google Scholar] [CrossRef]
- Kumar, R.; Bhat, T.A.; Walsh, E.M.; Chaudhary, A.K.; O’Malley, J.; Rhim, J.S.; Wang, J.; Morrison, C.D.; Attwood, K.; Bshara, W.; et al. Cytochrome c Deficiency Confers Apoptosome and Mitochondrial Dysfunction in African-American Men with Prostate Cancer. Cancer Res. 2019, 79, 1353–1368. [Google Scholar] [CrossRef]
- Kuhns, K.J.; Zhang, G.; Wang, Z.; Liu, W. ARD1/NAA10 acetylation in prostate cancer. Exp. Mol. Med. 2018, 50, 1–8. [Google Scholar] [CrossRef]
- Xie, X.; Xu, Z.; Wang, C.; Fang, C.; Zhao, J.; Xu, L.; Qian, X.; Dai, J.; Sun, F.; Xu, D.; et al. Tip60 is associated with resistance to X-ray irradiation in prostate cancer. FEBS Open Bio. 2018, 8, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Liu, X.; Zhu, S.; Hu, X.; Niu, H.; Zhang, X.; Zhu, D.; Nesa, E.U.; Tian, K.; Yuan, H. Hyper-acetylation contributes to the sensitivity of chemo-resistant prostate cancer cells to histone deacetylase inhibitor Trichostatin A. J. Cell Mol. Med. 2018, 22, 1909–1922. [Google Scholar] [CrossRef] [PubMed]
- Richa, S.; Dey, P.; Park, C.; Yang, J.; Son, J.Y.; Park, J.H.; Lee, S.H.; Ahn, M.Y.; Kim, I.S.; Moon, H.R.; et al. A New Histone Deacetylase Inhibitor, MHY4381, Induces Apoptosis via Generation of Reactive Oxygen Species in Human Prostate Cancer Cells. Biomol. Ther. 2020, 28, 184–194. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structure | Cytc Mouse | Cytc (K53Q) Mouse | Cytc (Y46F) Human |
|---|---|---|---|
| PDB code | 5C0Z | 7LJX | 3ZOO |
| Data code | 141,118 × 21a | 161,109 × 06a | |
| CRYSTALLIZATION | |||
| Iron | Oxidized | Oxidized | Oxidized |
| Protein | 15 mg/mL Cytc (WT) + 5 mM K3Fe(CN)6 in water | 17mg/mL Cytc (K53Q) + 5 mM K3Fe(CN)6 in water | 12.5 mg/mL oxidized Cytc in 22.5% (w/v) PEG-1000, 50 mM |
| KH2PO4, pH 7.0 | |||
| Well | 25% PEG 4K, 8% isopropanol, 0.1 M Na Acetate, pH 6.5 | 30% PEG 2K MME, 0.1M Na MES, 0.1 M Na Acetate, pH 6.5 | 26–31% (w/v) PEG 1000, 40 mM, KH2PO4. pH 7.0 |
| Drop | 1:1 Protein:Well | 1:1 Protein:Well | n/a |
| Cryoprotectant | 30% PEG 4K, 8% isopropanol, 0.1 M Na Acetate, 20% Ethylene glycol, 10 min soak with 5 mM K3Fe(CN)6, final pH 6.5 | 35% PEG 2K MME, 0.1M Na MES, 0.1 M Na Acetate, pH 6.5, 20% Ethylene glycol | 26–31% (w/v) PEG 1000, 40 mM, KH2PO4, 15% Glycerol, pH 7.0 |
| CRYSTAL DATA | |||
| Space group: | P1 | P2(1)2(1)2 | P1 |
| Unit cell: a | 34.401 | 52.837 | 36.367 |
| b | 52.471 | 96.767 | 53.952 |
| c | 61.647 | 38.372 | 58.95 |
| Alpha | 110.04 | 90 | 76.55 |
| Beta | 92.77 | 90 | 88.73 |
| Gamma | 92.02 | 90 | 71.86 |
| Chains per A.U. | 4 | 2 | 4 |
| Matthews Coeff | 2.24 | 2.1 | 2.3 |
| Solvent % | 45.12 | 41.4 | 46.7 |
| X-RAY DATA | |||
| Resolution-high (Å) | 1.12 | 1.31 | 1.35 |
| Resolution-low (Å) | 49.22 | 48.38 | 30.59 |
| Beamline | APS 21-ID-F | APS 21-ID-D | DIAMOND 103 (UK) |
| Wavelength | 0.97872 | 1.07822 | 0.9762 |
| Reflections | 127840 | 48257 | 81707 |
| Completeness | 87.4 (44.7) | 98.9 (86.8) | 94.7 (80.7) |
| Average I/sigma | 14.4 (2.0) | 12.2 (2.1) | 8.70 (n/a) |
| Redundancy | 3.9 (3.7) | 11.1 (6.6) | n/a |
| Resolution-high (Å) | 1.12 | 1.31 | 1.35 |
| Resolution-low (Å) | 49.22 | 48.38 | 30.59 |
| Rmerge | 0.050 (0.548) | 0.120 (0.821) | 0.07 |
| REFINEMENT | |||
| Rfactor | 0.132 (0.239) | 0.138 (0.291) | 0.138 |
| Rfree | 0.159 (0.240) | 0.170 (0.316) | 0.179 |
| Avg B-factor (Å2) | 15.97 | 21.32 | 17.703 |
| Protein atoms per A.U. | 3228 | 1614 | 3981 |
| Water molecules | 537 | 194 | 417 |
| Bond RMSD | 0.017 | 0.016 | 0.017 |
| Angle RMSD | 1.903 | 2.276 | 1.99 |
| FC6 OCCUPANCY | |||
| 202a/A | 0.28 | 0.5 | |
| 202b/A | 0.37 | ||
| 202/B | 0.93 | ||
| 203/B | – | – | |
| 203a/B | 0.3 | ||
| 203b/B | 0.32 | ||
| 202/C | 0.9 | ||
| HEME LINKS: | |||
| Average (Å) | |||
| C14 SG—HEM CAB | 1.90 (0.04) | 1.77 (0.01) | 1.95 (0.02) |
| C17 SG—HEM CAC | 1.98 (0.04) | 1.86 (0.00) | 2.13 (0.07) |
| H18 NE2—FE2 (Å) | 2.02 (0.01) | 1.99 (0.01) | 2.02 (0.03) |
| M80 SD—FE2 (Å) | 2.30 (0.01) | 2.31 (0.01) | 2.28 (0.02) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazylianska, V.; Kalpage, H.A.; Wan, J.; Vaishnav, A.; Mahapatra, G.; Turner, A.A.; Chowdhury, D.D.; Kim, K.; Morse, P.T.; Lee, I.; et al. Lysine 53 Acetylation of Cytochrome c in Prostate Cancer: Warburg Metabolism and Evasion of Apoptosis. Cells 2021, 10, 802. https://doi.org/10.3390/cells10040802
Bazylianska V, Kalpage HA, Wan J, Vaishnav A, Mahapatra G, Turner AA, Chowdhury DD, Kim K, Morse PT, Lee I, et al. Lysine 53 Acetylation of Cytochrome c in Prostate Cancer: Warburg Metabolism and Evasion of Apoptosis. Cells. 2021; 10(4):802. https://doi.org/10.3390/cells10040802
Chicago/Turabian StyleBazylianska, Viktoriia, Hasini A. Kalpage, Junmei Wan, Asmita Vaishnav, Gargi Mahapatra, Alice A. Turner, Dipanwita Dutta Chowdhury, Katherine Kim, Paul T. Morse, Icksoo Lee, and et al. 2021. "Lysine 53 Acetylation of Cytochrome c in Prostate Cancer: Warburg Metabolism and Evasion of Apoptosis" Cells 10, no. 4: 802. https://doi.org/10.3390/cells10040802
APA StyleBazylianska, V., Kalpage, H. A., Wan, J., Vaishnav, A., Mahapatra, G., Turner, A. A., Chowdhury, D. D., Kim, K., Morse, P. T., Lee, I., Brunzelle, J. S., Polin, L., Subedi, P., Heath, E. I., Podgorski, I., Marcus, K., Edwards, B. F. P., & Hüttemann, M. (2021). Lysine 53 Acetylation of Cytochrome c in Prostate Cancer: Warburg Metabolism and Evasion of Apoptosis. Cells, 10(4), 802. https://doi.org/10.3390/cells10040802