Abstract
This review deals with the main mechanisms of action exerted by antagonistic bacteria, such as competition for space and nutrients, suppression via siderophores, hydrolytic enzymes, antibiosis, biofilm formation, and induction of plant resistance. These mechanisms inhibit phytopathogen growth that affects postharvest fruit since quality and safety parameters are influenced by the action of these microorganisms, which cause production losses in more than 50% of fruit tree species. The use of synthetic fungicide products has been the dominant control strategy for diseases caused by fungi. However, their excessive and inappropriate use in intensive agriculture has brought about problems that have led to environmental contamination, considerable residues in agricultural products, and phytopathogen resistance. Thus, there is a need to generate alternatives that are safe, ecological, and economically viable to face this problem. Phytopathogen inhibition in fruit utilizing antagonist microorganisms has been recognized as a type of biological control (BC), which could represent a viable and environmentally safe alternative to synthetic fungicides. Despite the ecological benefit that derives from the use of controllers and biological control agents (BCA) at a commercial level, their application and efficient use has been minimal at a global level.
1. Introduction
Nowadays, postharvest fruit loss caused by phytopathogen fungi is estimated to account for more than 50% of total agricultural fruit production [1]. In general, control of fungal phytopathogens is performed by applying synthetic fungicides. However, their use has led to problems for the environment and human and animal health [2]. Furthermore, the global trend of consumers in acquiring agricultural products that come from systems without synthetic product application keeps growing every day. Thus, a worldwide priority has been the search for alternatives to control postharvest phytopathogen fungi. In recent years, several strategies have been developed to biologically control phytopathogens, mainly based on the use of microbial antagonists, such as bacteria [3]. Biological control (BC) is an important management strategy for fungal diseases. It is principle based on utilizing live microorganisms to reduce and/or maintain the population of a phytopathogen below the levels which cause economic loss. This BC method is efficient in the short, medium, and long term, and does not represent a risk to the environment or cause harm to humans and animals [4]. Bacteria show diverse antagonistic mechanisms toward phytopathogen fungi, notably space and nutrient competition, hydrolytic enzymes, induction of resistance, volatile compound synthesis, and biofilms [5,6]. The use of bacteria as biocontrol agents (BCA) has been very important in the integrated management of cultivations and organic production, where their value as a postharvest control of fungal diseases stands out [7].
2. Biocontrol as a Method for Decreasing Phytopathogen Abundance
BC methods provide plant protection against fungal diseases, and currently represent a viable alternative for fruit protection against phytopathogens at the postharvest stage [8]. Implementing BC in cultivation management offers diverse benefits, such as a decrease of causal agents; cultivation protection; low cost; lack of contamination of soil, water, and plant; and lack of waste management problems [9,10]. The scope of BC shows some important limitations, such as temperature, humidity, its narrow range of activity, and so on. For these reasons, BC strategies in the field are subject to significant variability, which constitutes an important limitation to their practical implementation [11]. After more than three decades of BC research, it has been scaled up to allow greater integration in agricultural production systems [12].
2.1. Selection and Identification of a Biological Control Agent
Among the different types of microorganisms utilized in BC, bacterial communities show antagonistic effects toward phytopathogens, so they can be exploited as a form of BC (Table 1). A continuous interaction exists in the biological world between phytopathogens and their antagonists, in such a way that the latter inhibit disease presence and development. For the adequate use of this type of microorganism, it is important to understand the mechanisms of action involved in BC activity for the safe development of the application processes and as a basis for selecting the most efficient bacterial strains [13,14]. The selection of bacteria with potential for use as BCA starts from their isolation from soil samples or from any part of the plant. After that, they are identified by chemical and molecular biology methods; their antagonistic mechanisms against phytopathogens are characterized, and several tests are performed to determine whether they are non-toxic for the environment, plants, and human and animal health [15]. Bacteria should have particular characteristics to be considered as BCA, notably genetic stability, efficacy at low concentrations, and colonization capacity in diverse hosts (Table 2).

Table 1.
Examples of biological control of postharvest disease by bacterial species.
Table 2.
Ideal characteristics of a biological control agent [29].
2.2. Antagonistic Mechanisms of Bacteria Utilized as Biocontrol Agents
Phytopathogen inhibition by bacteria is generally performed with at least two antagonistic mechanisms, which make it more efficient to control diseases in postharvest fruit [5]. Several modes of anti-phytopathogen action of bacteria have been described [29,30], notably space and nutrient competition, parasitism, volatile compounds, and biofilms, among others (Figure 1). The main antagonistic mechanisms exerted by bacteria against phytopathogens are described below.
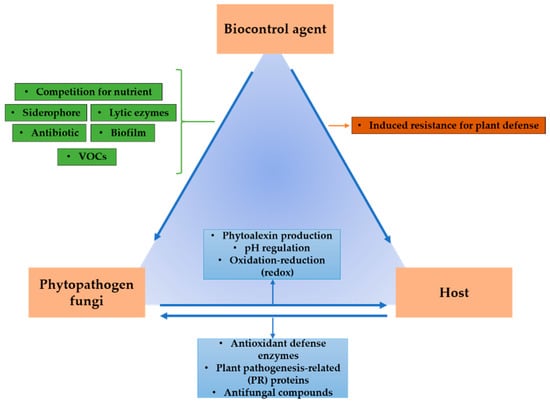
Figure 1.
Mechanisms of action of biocontrol agents.
2.2.1. Competition for Space and Nutrients
Competition for space and carbon sources is one of the main antagonistic mechanisms exerted by bacteria against phytopathogens [31]. The capacity of bacteria to colonize damaged fruit depends on the adaptation to assimilate the necessary carbon sources for survival and multiplication, limiting carbohydrate disposition for the phytopathogen fungus, reducing its spore germination percentage and thus its capacity to invade the host [32]. Different in vitro studies have demonstrated that antagonistic microorganisms limit phytopathogen fungi by different carbon sources, mainly saccharose, fructose, and glucose, among others [33]. Phytopathogen inhibition can be greater if the cellular dose of the antagonist increases; thus, there is a need to perform studies on the application of different bacterial doses on the host to determine the minimum concentration required to inhibit the phytopathogen [34]. Yu and Lee [35] demonstrated that the bacteria Pseudomonas putida inhibited spore germination of Penicillium digitatum due to nutrient availability of the host. Other bacteria have inhibited different phytopathogen fungi by carbon sources, such as Pseudomonas syringae, Pantoea agglomerans [24], Pseudomonas fluorescens [30] and Bacillus megaterium [16].
2.2.2. Siderophores
Iron is an essential element for microorganism growth [36]; Fe2+ oxidizes Fe3+ to form stable iron oxide complexes in the presence of oxygen and water. These molecules are sequestered by siderophores; they are produced by bacteria and are defined as small peptide molecules that contain lateral chains and functional groups that can provide a set of high-affinity to coordinate iron ions [37]. Siderophores are classified into four types: carboxylates, hydroxamates, catolate phenate, and pyridoxines [38]. When bacteria produce siderophores in a medium, they displace and inhibit phytopathogens in the host, limiting their spore germination and mycelial growth [39]. Diverse types of siderophores produced by antagonistic bacteria have been identified and reported; some of them are recognized and utilized by different microorganisms, while others are specific for each bacterial species (Table 3). Their production confers competitive advantages to the bacteria used as BCA by excluding other microorganisms (including phytopathogens) of the host [40,41]. Under competition conditions to acquire iron, siderophore production by antagonistic bacteria can determine a greater phytopathogen inhibition in the host [42].
Table 3.
Siderophore production by antagonistic bacteria.
2.2.3. Parasitism (Lithic Enzyme Production)
In BC, parasitism takes place when the antagonist feeds on the phytopathogen, producing partial or total lysis of its structures. Specifically, bacteria feed on the cell walls of fungi, mainly on chitin, glucans, and proteins, which constitute 20%, 50–60%, and 20–30% of the cell wall, respectively [51]. Chitin is a lineal and insoluble homopolymer formed by sub-units of N-Acetyl glucosamine linked by β-1,4, and acts as the medullar support of the cell wall [52]. β-1,3-glucan is considered as the main structural constituent of the cell wall, to which other components of the cell wall are covalently linked, providing mechanical strength and integrity. The majority of the cell wall proteins are glycoproteins with diverse modification with oligosaccharides. The proteins of the cell wall play an important role in maintenance of the cell form, synthesis and remodeling of the cell wall components, molecule absorption, and cell protection against external molecules. The decomposition of the fungal cell wall requires the participation of different enzymes, mainly β-1,3-glucanase, chitinase, and protease [5]. Glucan enzymes produced by bacteria can hydrolyze glucans by two possible mechanisms: (1) exo-β-1,3-glucanase, which hydrolyzes glucans by sequential segmentation of glucose residuals from the non-reducer extreme; and (2) endo-β-1,3-glucanase, which activates the links to aleatory sites along the polysaccharide chain, in which oligosaccharides and glucose are released in small amounts [51]. Chitinases hydrolyze chitin, the non-branched homopolymeric N-Acetyl glucosamine in a 1,4 link by two possible mechanisms: (1) exo-chitinase or N-Acetyl-b-glucosaminidase, which sequentially segments the NAG extreme residues; and (2) endo-chitinase, which activates links in aleatory sites along the polymer chain [53]. Proteases can be divided into four main groups according to their site of action, namely, serine proteinases, cysteine proteinases, aspartic proteinases, and metaloproteinases [54]. During the last decade, different research studies have been performed on hydrolytic enzymes that produce bacteria and yeasts [29]. The genera Bacillus and Pseudomonas are considered as some of the most efficient antagonists in phytopathogen control due to the direct action of chitinase [55]. Shivakumar et al. [56] performed partial purification, characterization, and kinetic studies of the chitinase enzyme of the bacteria Bacillus subtilis JN032305, in which the partially purified enzyme not only showed antifungal activity against Rhizoctonia solani and Colletotrichum gloeosporioides but also increased pepper seed germination when infected by the mentioned fungi. Mohammadi et al. [57] performed a study on potential bacterial isolation (Bacillus subtilis, Bacillus pumilus, Bacillus megaterium, and Agrobacterium radiobacter) for BC in citrus fruit against blue mold caused by Penicillium digitatum, in which bacterial extracellular enzymatic activities were determined, showing that all chitinase and glucanase enzymes were positive and protease enzyme activity was positive except for the bacteria Agrobacterium radiobacter.
2.2.4. Formation of Biofilms and Quorum Sensing
To colonize fruit surfaces successfully, antagonistic bacteria should have specific characteristics that facilitate their adherence, colonization, and multiplication. In the majority of cases, these characteristics are associated with the formation of biofilms, in which the microcolonies are within a protein-hydrated matrix produced by bacteria and nucleic and polysaccharide acids. The formation of this structure is measured by quorum sensing with regulators, such as tyrosol, farnesol, and phenethyl alcohol. The microcolonies maintain a form of communication through quorum sensing, employing diverse chemical signals to supervise their environment, altering the genetic expression and obtaining advantage over their competitors [58]. In BC, the formed biofilms act as barriers that stand between the host lesion surface and the phytopathogen. However, little is known about the main functions and mechanisms involved in the formation of biofilms. It has been suggested that the environmental signals that regulate the morphogenetic transformations help to select the bacteria more efficiently as BCA. In agricultural environments, there is a growing number of studies on the formation of biofilms [59,60]. The basis that links these models is that Bacillus subtilis functions as a cooperative community by population differentiation of specialized isogenic progenitors, as well as the production of macromolecules that form the biofilm matrix [61,62]. The matrix of this bacterium consists of proteins called TasA and TapA [63,64] besides a secreted polysaccharide of high molecular weight [65]. The assemblage of the mature biofilm also requires the presence of a coating protein called biofilm BsIA, previously known as YuaB [65,66,67]. Haggag and Timmusk [68] demonstrated that the bacteria Paenibacillus polymyxa colonizes plant roots, forming structures that look like biofilms and thus protecting the roots against diseases caused by phytopathogens. Another example is the highly mutant mucoid (forming an improved biofilm) of Pseudomonas fluorescens strain CHA0, which showed an improved capacity to colonize carrot roots [69]. Another study showed that a strain of B. subtilis (ATCC6051) can form biofilm-type structures in the roots of Arabidopsis plants and thereby protect them from infection by the bacteria Pseudomonas syringae [70].
2.2.5. Antibiosis
Antibiotic production is one of the mechanisms that has been classified as important after space and nutrient competition. The main antibiotics produced by antagonistic bacteria are iturin, a potent antifungal peptide produced by the bacteria Bacillus subtilis; pyrrolnitrin, produced by Pseudomonas cepacia; and trichothecene, produced by Myrothecium roridum [71]. Antibiosis can also be performed through the production of low-molecular-weight compounds by microorganisms, called volatile organic compounds (VOCs), which have a direct effect on phytopathogen growth. They are active in low concentrations and belong to several chemical groups, such as alcohols, aldehydes, ketones, esters, lactones, terpenes, and sulfur compounds. Due to their volatility, these compounds can travel great distances in structurally heterogeneous environments, as well as in solid, liquid, or gas compounds, which is a great advantage for BCA. VOCs have received limited attention compared to other antagonistic mechanisms. However, recent research studies have focused on these volatile metabolism products [72]. Fruit bio-fumigation using microorganisms that produce VOCs in closed chambers can be a viable alternative to control phytopathogens [73]. However, it is worth mentioning that VOCs emitted by BCA can only provide a limited contribution to phytopathogen control since they show fungistatic activity. Nevertheless, some phytopathogens can also introduce a wide array of VOCs. Thus, this approach should be assessed in-depth in future research studies [51]. Layton et al. [74] mentioned that the bio-controlling action of the genus Bacillus was measured by its biochemical profile since they are producers of multiple secondary metabolites that are biologically active. For example, iturin A and fengycin are produced by Bacillus subtilis, and Brevibacillus brevis produces gramicidin S (1–5) metabolites, which are capable of inhibiting the growth and development of diverse phytopathogens; therefore, both Bacillus subtilis and Brevibacillus brevis are recommended for BC. Velázquez-Becerra et al. [75] found that Arthrobacter agilis UMCV2 produced VOCs. This was confirmed by using gas chromatography–mass spectrometry analysis, which identified dimethyl hexadecylamine as the compound that inhibited Botrytis cinerea and Phytophthora cinnamomi, demonstrating an inhibitory activity 12 times higher than that caused by the fungicide Captan. Furthermore, Raza et al. [76] demonstrated that the bacteria Bacillus amyloliquefaciens SQR-9 produced VOCs capable of inhibiting the growth of Ralstonia solanacearum. The strain SQR-9 produced 22 organic volatile compounds; however, only nine showed 1–11% antibacterial activity against the disease.
2.2.6. Induction of Resistance
Induction of resistance is the capacity of an antagonistic bacterium to provoke defense response in the host through several chemical or biochemical reactions, including changes in tissue structure and protein production related to pathogenesis, expressed either locally or systemically [29]. Induction of resistance in fruit by bacteria, as BCA, has been considered a mode of action to control postharvest diseases [32,77]. Induction of defense response is given by the overproduction of different enzymes and metabolites, notably (1) proteins linked to pathogenicity (PR proteins), which include glucanases, chitinases, peroxidases, superoxide dismutase, catalase (which protects tissue against oxidative damage), protein inhibitors, or lipid-transport proteins; (2) compounds with antimicrobial activity as phytoalexines; and (3) papillae formed by callose and lignin deposited in the cell wall assuring their strengthening. Other patterns that trigger immunity are (1) the production of reactive oxygen species (ROS) with a signaling role and direct antimicrobial effect; and (2) stomatal closing. In the case of plants, the immune response is mediated by and dependent on phytohormones, as well as salicylic acid, jasmonic acid, ethylene, and abscisic acid and their interaction, allowing the activation of immune responses against specific organisms [78,79]. Although induction of resistance in the host tissue by the antagonistic bacterial treatment is correlated, direct evidence of the capacity of the substances induced to inhibit phytopathogen infection has not been established [51].
2.3. Commercializing Biological Products
Numerous microbial antagonists of postharvest phytopathogens have been identified in laboratory and field studies and exploited commercially [12]. To date, the development of products based on beneficial microorganisms is relevant to reducing chemical contaminants in the food supply [80]. Microorganisms can be isolated from environments such as soil and sea, including some parts of plants [81]. Developing a BCA is a complex, long, costly, and interactive process. Despite the great amount of research invested in studying BCAs, it is paradoxical that their commercial use has been very limited. One of the greatest constraints to products based on BC reaching a high level of commercial consumption is efficiency under field conditions. These conditions depend on environmental factors which are not possible to control, such as temperature, humidity, precipitation, and so on, abiotic aspects which synthetic fungicides have overcome and remained as an essential medium to control phytopathogens [2]. Before starting to develop a product based on antimicrobial agents, it is necessary to have a detailed knowledge of several factors of the fungal disease to be defended against, namely, the phytopathogen species, type of hosts it attacks, epidemiology of the disease, phytopathogen resistance, and environmental conditions under which the BCA will be used. Therefore, the antagonist should be selected successfully to avoid future difficulties. For a BCA product to be released into the market, it should pass different studies, and the production process should be scaled up to an experimental level to obtain sufficient quantities of the product to carry out its assessment in greenhouse, field, warehouse, or packing plant. If the product shows sufficient attributes, and its production, both technical and economic, is viable, its registration and commercialization follows [82]. The general objective of the majority of the research groups related to BC is to develop a product with a commercial purpose. However, despite all the efforts, only some BCs are commercially available for postharvest application, as has been described recently by Droby et al. [83].
There are currently several commercial products for BC of phytopathogens, which are considered first-generation biocontrol products for postharvest and whose active ingredients are antagonist yeast and bacteria that are available at commercial level in pre- and postharvest operations. Some of these products are Candida oleophila (Aspire, Ecogen, Langhorne, PA, USA) [84], Cryptococcus albidus (YieldPlus, Lallemand, Montreal, QC, Canada), Candida sake (Candifruit, Sipcam Ibérica, Valencia, Spain) [85], and Pseudomonas syringae (BioSave, JET Harvest, Longwood, FL, USA) [86]. Biosave Aspire and Candifruit were commercialized a few years ago; however, they were suspended due to commercial deficiencies related to marketing. Biosave still has a limited use in the USA market for application in fruit cultivation [87]. Bacillus subtilis (Avogreen, Pretoria University, Pretoria, South Africa) was introduced in South Africa to fight the spot caused by Cercospora sp., a disease in postharvest avocado; however, it did not reach commercial success due to inconsistent results [88]. Candida oleophila (Nexy, Leasafre, Lille, France) was developed in Belgium and was presented for statutory approval in 2005 for application in postharvest phytopathogen control in citrus and banana fruit [89]. Nexy received regulatory approval in all of the European Union in 2013 [90]. Aureobasidium pullulans (Boni-Protect, Bio-Ferm, Tulln, Austria) should be applied before preharvest to control phytopathogens in lesions developed in fruit during storage [91]. Pantovital (Pantoea agglomerans CPA-2) has been used to control pre- and postharvest diseases in fruit, such as citrus; it was formulated but never commercialized [9]. Amylo-X, made by Biogard CBC in Grassobbio, Italy, was based on Bacillus amyloliquefaciens and used to control fungal and bacterial diseases of several vegetables [92]. Metschnikowia fructicola (Shemer, Bayer, Leverkusen, Germany) was recorded in Israel to be applied pre- and postharvest in citrus fruit, grapes, peach, and strawberry, representing the most successful example of a postharvest biocontrol product. Shemer was acquired by Bayer CropScience (Monheim, Germany) and then sub-licensed to Koppert (Berkel en Rodenrijs, the Netherlands) [51]. Serenade was made by Bayer in Leverkusen, Germany; its active ingredient was the bacteria Bacillus subtilis, and it was used to control diseases in peach, strawberry and tomato cultivations [92]. However, none of these are used as an alternative to synthetic products [93].
2.4. Search for New Antagonists
In general, the majority of the antagonistic microorganisms are isolated from the surface of the fruit, plant (roots, leaves, flowers, and seeds), sea, or soil [81]. A study performed by Maida et al. [93] dealt with antagonistic interactions between isolated bacteria from three ecological niches (rhizospheric soil, roots, and stem-leaves) of Echinacea purpurea, whose antagonistic potential has also been exploited as medicine. Furthermore, Goudjal et al. [94] mentioned that wild plants adapt better to difficult soil conditions, especially to the attack of phytopathogens transmitted by themselves, which is why they are considered a potential source for isolating antagonistic endophyte microorganisms. Recent studies on antagonistic microorganisms have made evident their potential use in controlling phytopathogens, which affect important crops [95,96]. Endophyte microorganisms with bacteria that are antagonistic against phytopathogens are promising candidates for BC strategies [97]. Diverse studies have mentioned that the role of endophyte communities in seeds of different cultivations is poorly characterized [98,99,100]. Santos et al. [101] isolated endophyte bacteria in guarana seeds; the isolations showed growth inhibiting capacity of diverse phytopathogens. Antagonists have also been isolated from fruit, and these have shown potential for exploitation as BCA of postharvest diseases [14]. The marine environment is also a deposit of new bioactive compounds; thousands of unique compounds have been isolated from this environment [102,103]. In recent years, any bioactive compound and different samples from marine environments have been searched for. Many studies have shown that antagonistic bacterial communities associated with marine environments may have a beneficial role for plant protection [104]. However, little information has been found with respect to the study of microorganisms isolated from marine environments with the potential to control phytopathogens in postharvest, which could have agronomic potential compared to synthetic fungicides [105]. In China, bacteria isolated from marine environments have great potential for BC in peanut cultivation [16]. Thus, marine bacteria have gained attention as possible BCA against fungal postharvest diseases, mainly due to their high inhibiting capacity, fast colonization in fruit lesions, and simple nutritional requirements [106]. Bibi et al. [107] mentioned that compost could be considered as a source of microorganisms capable of producing secondary metabolites of agricultural interest for phytopathogen control. Other antagonists could be actinomycetes, which are characterized by the production of metabolites with antimicrobial activity. Marine actinomycetes are considered major producers of secondary metabolites that inhibit mycelial growth of phytopathogen fungi, a quality that allows them to be considered as candidates to be used in BD against phytopathogen bacteria [108,109].
2.5. Application of Biocontrol Agents in Preharvest Stages
Several antagonistic microorganisms have been recommended to be used in fields before harvest with the purpose of protecting postharvest cultivation [7,29]. Lopes et al. [110] assessed the efficacy of different antagonistic microorganisms in controlling Colletotrichum acutatum with applications on preharvest citrus fruit and found promising results. Silva and De Costa [111] assessed the potential of Burkholderia spinosa with preharvest applications for the BC of phytopathogens in banana (Musa spp.). Camañas et al. [112] assessed the efficiency of different formulations of the antagonistic bacteria Pantoea agglomerans with preharvest application to control postharvest diseases in citrus fruit; the results showed an efficient protection of orange against Penicillium digitatum during storage. Luo et al. [78] showed that the preharvest application of antagonistic microorganisms decreased the presence of diseases caused by Penicillium digitatum and Penicillium italicum significantly in orange cultivation. The combination of antagonists with other antimicrobial compounds could be an efficient method to improve the yield of BC. Combination with salts, such as bicarbonates, and natural compounds, such as chitosan, have improved the yield of BCA [113].
2.6. Application of Biocontrol Agents Postharvest
Microbial antagonists are applied directly to fruit by pulverization or immersion in solution. Suppressing phytopathogen agents during storage was shown to be more successful in several studies of BC compared with previous application at harvest [6]. Antagonistic bacteria isolated from natural habitats produce several metabolites with antifungal and antibacterial capabilities [114]. In recent years, several strains of the genera Bacillus, Burkholderia, Enterobacter, and Pseudomonas have been studied and used effectively to treat diseases caused by phytopathogens. Suppressing fungal growth has been achieved by using antagonistic bacterial species such as Bacillus subtilis, Bacillus thuringiensis, Burkholderia sp., Enterobacter cloacae, Pantoea agglomerans and Serratia plymuthica [21]. Nevertheless, due to the high cost and complex process of the sanitary registry, as well as the limited market and strong demand for phytosanitary products, the number of BCAs registered is lower than the large number of research studies that have been performed on BC [93].
3. Conclusions
The residue of synthetic products on fruit has been and will continue to be one of the main worries of regulatory agencies and consumers. The growing concern about the hazards involved relating to human health and environmental contamination has led to a demand for the development of alternatives to control postharvest diseases. The use of BCA as an alternative to synthetic products has been a focus of research in the last 30 years by many researchers and several commercial enterprises worldwide. This review has provided a general description on modes of action that antagonistic bacteria exert, which are considered viable alternatives to synthetic fungicides, as well as the success of some of them under laboratory conditions, leading to the production of biological products based on antagonistic bacteria for postharvest application. More research should be developed with respect to the mode of action of antagonists to better understand the phytopathogen–antagonist–host interactions. In recent years, there has been an advance in the use of molecular techniques that contribute to improving the knowledge of antagonistic mechanisms of BCA. Nonetheless, the search for new antagonists should be ongoing to widen the use of BCA postharvest.
Author Contributions
S.C.-H., R.G.C.-C., and L.G.H.-M. carried out the initial analysis of the literature and wrote the first draft of the paper; G.R.-E., J.J.R.-P., and C.R.C.-C. added references and improved the analysis; all authors co-wrote the final text. R.G.C.-C. and L.G.H.-M. are the main thesis advisors of S.C.-H., a Master of Science student.
Funding
All research was funded by the grant project SEP-CONACYT 181972, the grant project Problemas Nacionales 2015-01-352 CONACYT (National Council for Science and Technology) and the CONACYT grant awarded to S. Carmona-Hernandez.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, H.; Mahunu, G.K.; Castoria, R.; Apaliya, M.T.; Yang, Q. Argumentation of biocontrol agents with physical methods against postharvest diseases of fruits and vegetables. Trends Food Sci. Technol. 2017, 69, 36–45. [Google Scholar] [CrossRef]
- Nunes, C.A. Biological control of postharvest diseases of fruit. Eur. J. Plant Pathol. 2012, 133, 181–196. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty years of postharvest biocontrol Research: Is it time for a new paradigm? Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- Cuthbert, R.N.; Dick, J.T.; Callaghan, A.; Dickey, J.W. Biological control agent selection under environmental change using functional responses, abundances and fecundities; the relative control potential (RCP) metric. Biol. Control 2018, 121, 50–57. [Google Scholar] [CrossRef]
- Safdarpour, F.; Khodakaramian, G. Assessment of antagonistic and plant growth promoting activities of tomato endophytic bacteria in challenging with Verticillium dahliae under in-vitro and in-vivo conditions. Biol. J. Microorg. 2019, 7, 77–90. [Google Scholar]
- Dukare, A.S.; Sangeeta, P.; Eyarki, N.V.; Ram, K.G.; Rajbir, S.; Kalyani, S.; Rajesh, K.V. Exploitation of microbial antagonists for the control of postharvest diseases of fruits: A review. Crit. Rev. Food Sci. Nutr. 2018. [Google Scholar] [CrossRef] [PubMed]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting rhizobacteria (PGPR): their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Ghazanfar, M.U.; Hussan, M.; Hamid, M.I.; Ansari, S.U. Utilization of biological control agents for the management of postharvest pathogens of tomato. Pak. J. Bot. 2016, 48, 2093–2100. [Google Scholar]
- Torres, M.J.; Brandan, C.P.; Petroselli, G.; Erra-Balsells, R.; Audisio, M.C. Antagonistic effects of Bacillus subtilis subsp., subtilis and B. amyloliquefaciens against Macrophomina phaseolina: SEM study of fungal changes and UV-MALDI-TOF MS analysis of their bioactive compounds. Microbiol. Res. 2016, 182, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Mcdonald, B.A. Using dynamic diversity to achieve durable diseases resistance in agricultural ecosystems. Trop. Plant Pathol. 2014, 39, 191–196. [Google Scholar] [CrossRef]
- Marin, A.; Atarés, L.; Chialt, A. Improving function of biocontrol agents incorporated in antifungal fruit coatings: A review. Biocontrol Sci. Technol. 2017, 27, 1220–1241. [Google Scholar] [CrossRef]
- Gotor-Vila, A.; Teixidó, N.; Casals, C.; Torres, R.; De Cal, A.; Guijarro, B.; Usall, J. Biological control of brown rot in Stone fruit using Bacillus amyloliquefaciens CPA-8 under field conditions. Crop Prot. 2017, 102, 72–80. [Google Scholar] [CrossRef]
- Nikolić, I.; Berić, T.; Dimkić, I.; Popović, T.; Lozo, J.; Fira, D.; Stanković, S. Biological control of Pseudomonas syringae pv. aptata on sugar beet with Bacillus pumilus SS-10.7 and Bacillus amyloliquefaciens (SS-12.6 and SS-38.4) strains. J. Appl. Microbiol. 2019, 126, 165–176. [Google Scholar]
- Paz, I.C.P.; Santin, R.D.C.M.; Guimarães, A.M.; da Rosa, O.P.P.; Quecine, M.C.; e Silva, M.D.C.P.; Azevedo, J.L.; Matsumura, A.T.S. Biocontrol of Botrytis cinerea and Calonectria gracilis by eucalypts growth promoters Bacillus spp. Microb. Pathog. 2018, 121, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Utilization of antagonistic yeast to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Y.; Fu, X.; Li, Y.; Wang, Q. Isolation and characterization of Bacillus amyloliquefaciens PG12 for the biological control of apple ring rot. Postharvest Biol. Technol. 2016, 115, 113–121. [Google Scholar] [CrossRef]
- Aiello, D.; Restuccia, C.; Stefani, E.; Vitale, A.; Cirvilleri, G. Postharvest biocontrol ability of Pseudomonas synxantha against Monilinia fructicola and Monilinia fructigena on stone fruit. Postharvest Biol. Technol. 2019, 149, 83–89. [Google Scholar] [CrossRef]
- Wallace, R.L.; Hirkala, D.L.; Nelson, L.M. Mechanisms of action of three isolates of Pseudomonas fluorescens active against postharvest grey mold decay of apple during commercial storage. Biol. Control 2018, 117, 13–20. [Google Scholar] [CrossRef]
- Kong, Q.; Shan, S.; Liu, Q.; Wang, Q.; Yu, F. Biocontrol of Aspergillus flavus on peanut kernels by use of a strain of Marine Bacillus megaterium. Int. J. Food Microbiol. 2010, 139, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Montiel, L.G.; Zulueta-Rodriguez, R.; Angulo, C.; Rueda-Puente, E.O.; Quiñonez-Aguilar, E.E.; Galicia, R. Marine yeasts and bacteria as biological control agents against anthracnose on mango. J. Phytopathol. 2017, 165, 833–840. [Google Scholar] [CrossRef]
- Fu, G.; Huang, S.; Ye, Y.; Wu, Y.; Cen, Z.; Lin, S. Characterization of a bacterial biocontrol strain B106 and its efficacy in controlling banana leaf spot and post-harvest anthracnose diseases. Biol. Control 2010, 55, 1–10. [Google Scholar] [CrossRef]
- Khleekorn, S.; Wongrueng, S. Evaluation of antagonistic bacteria inhibitory to Colletotrichum musae on banana. J. Agric. Technol. 2014, 10, 383–390. [Google Scholar]
- Calvo, H.; Marco, P.; Blanco, D.; Oria, R.; Venturini, M.E. Potential of new strain Bacillus amyloliquefaciens BUZ-14 as a biocontrol agent of postharvest fruit diseases. Food Microbiol. 2017, 63, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Balaraju, K.; Jeon, Y. Effects of rhizobacteria Paenibacillus polymyxa APEC136 and Bacillus subtilis APEC170 on biocontrol of postharvest pathogens of apple fruits. J. Zhejiang Univ-Sci. B 2016, 17, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, S.; Vitale, A.; Polizzi, G.; Scala, F.; Cirvilleri, G. Enhanced control of postharvest citrus fruit decay by means of the combined use of compatible biocontrol agents. Biol. Control 2015, 84, 19–27. [Google Scholar] [CrossRef]
- Kasfi, K.; Taheri, P.; Jafarpour, B.; Tarighi, S. Identification of epiphytic yeast and bacteria with potential for biocontrol of gray mold diseases on table grapes caused by Botrytis cinerea. Span. J. Agric. Res. 2018, 16, 1–16. [Google Scholar] [CrossRef]
- Bull, C.T.; Stack, J.P.; Smilanick, J.L. Pseudomonas syringae Strains ESC-10 and ESC-11 survive in wounds on citrus and control green and blue molds of citrus. Biol. Control 1997, 8, 81–88. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Z.; Zhu, P.; Liu, Y.; Zhang, Z.; Mastuda, Y.; Toyoda, H.; Xu, L. Postharvest biological control of melon pathogens using Bacillus subtilis EXWB1. J. Plant Pathol. 2010, 92, 645–652. [Google Scholar]
- Bahadou, S.A.; Ouijja, A.; Karfach, A.; Tahiri, A.; Lahlali, R. New potential bacterial antagonists for the biocontrol of fire blight disease (Erwinia amylovora) in Morocco. Microb. Pathog. 2018, 117, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Thokchom, E.; Thakuria, D.; Kalita, M.C.; Sharma, C.K.; Talukdar, N.C. Root colonization by host-specific rhizobacteria alters indigenous root endophyte and rhizosphere soil bacterial communities and promotes the growth of mandarin orange. Eur. J. Soil Biol. 2017, 79, 48–56. [Google Scholar] [CrossRef]
- Di Francesco, A.; Mari, M. Biological control of postharvest diseases by microbial antagonists: How many mechanisms of action? Eur. J. Plant Pathol. 2016, 145, 711–717. [Google Scholar] [CrossRef]
- Hernandez-Montiel, L.G.; Gutierrez-Perez, E.D.; Murillo-Amador, B.; Vero, S.; Chiquito-Contreras, R.G.; Rincon-Enriquez, G. Mechanisms employed by Debaryomyces hansenii in biological control of anthracnose disease on papaya fruit. Postharvest Biol. Technol. 2018, 139, 31–37. [Google Scholar] [CrossRef]
- Adrees, H.; Haider, M.S.; Anjum, T.; Akram, W. Inducing systemic resistance in cotton plants against charcoal root rot pathogen using indigenous rhizospheric bacterial strains and chemical elicitors. Crop Prot. 2019, 115, 75–83. [Google Scholar] [CrossRef]
- Asari, S.; Matzén, S.; Petersen, M.A.; Bejai, S.; Meijer, J. Multiple effects of Bacillus amyloliquefaciens volatile compounds: Plant growth promotion and growth inhibition of phytopathogens. FEMS Microbiol. Ecol. 2016, 92, fiw070. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.M.; Lee, Y.H. Genes involved in nutrient competition by Pseudomonas putida JBC17 to suppress green mold in postharvest satsuma mandarin. J. Basic Microbiol. 2015, 55, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Terpilowska, S.; Siwicki, A.K. Cell cycle and transmembrane mitochondrial potential analysis after treatment with chromium (iii), iron (iii), molybdenum (iii) or nickel (ii) and their mixtures. Toxicol. Res. 2019. [Google Scholar] [CrossRef]
- Golonka, R.; San Yeoh, B.; Vijay-Kumar, M. The iron tug-of-war between bacterial siderophores and innate immunity. J. Innate Immun. 2019. [Google Scholar] [CrossRef] [PubMed]
- Carroll, C.S.; Moore, M.M. Ironing out siderophore biosynthesis: A review of non-ribosomal peptide synthetase (NRPS)-independent siderophore synthetases. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 356–381. [Google Scholar] [CrossRef] [PubMed]
- Cordova-Albores, L.C.; Zapotitla, E.S.; Ríos, M.Y.; Barrera-Necha, L.L.; Hernández-López, M.; Bautista-Banos, S. Microscopic study of the morphology and metabolic activity of Fusarium oxysporum f. sp. gladioli treated with Jatropha curcas oil and derivatives. J. Microsc. Ultrastruct. 2016, 4, 28–35. [Google Scholar] [PubMed]
- Zeng, J.; Xu, T.; Cao, L.; Tong, C.; Zhang, X.; Luo, D.; Han, S.; Pang, P.; Fu, W.; Yan, J.; et al. The role of iron competition in the antagonistic action of the rice endophyte Streptomyces sporocinereus OsiSh-2 against the pathogen Magnaporthe oryzae. Microb. Ecol. 2018, 76, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Drehe, I.; Simonetti, E.; Ruiz, J.A. Contribution of the siderophores pyoverdine and enantio-pyochelin to fitness in soil of Pseudomonas protegens Pf-5. Curr. Microbiol. 2018, 75, 1560–1565. [Google Scholar] [CrossRef] [PubMed]
- Andreolli, M.; Zapparoli, G.; Angelini, E.; Lucchetta, G.; Lampis, S.; Vallini, G. Pseudomonas protegens MP12: A plant growth-promoting endophytic bacterium with broad-spectrum antifungal activity against grapevine phytopathogens. Microbiol. Res. 2019, 219, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, D.; Hemantaranjan, A.; Dwivedi, P. Plant growth promoting rhizobacteria and their roles as fungal biocontrol agents: An overview. J. Plant Sci. Res. 2018, 34, 127–136. [Google Scholar] [CrossRef]
- De Los Santos-Villalobos, S.; Barrera-Galicia, G.C.; Miranda-Salcedo, M.A.; Peña-Cabriales, J.J. Burkholderia cepacia XXVI siderophore with biocontrol capacity against Colletotrichum gloeosporioides. World J. Microbiol. Biotechnol. 2013, 28, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Kesaulya, H.; Hasinun, J.V.; Tuhumury, G.N.C. Potential of Bacillus spp. produces siderophores in suppressing the wilt diseases of banana plants. In Proceedings of the International Symposium on Food and Agro-Biodiversity, Semarang, Indonesia, 26–27 September 2017. [Google Scholar]
- Sasirekha, B.; Srividya, S. Siderophore production by Pseudomonas aeruginosa PF6, a biocontrol strain for Rhizoctonia solani and Colletotrichum gloeosporioides causing diseases in chilli. Agric. Nat. Resour. 2016, 50, 250–256. [Google Scholar]
- Yu, X.; Ai, C.; Xin, L.; Zhou, G. The siderophore-producing bacterium, Bacillus subtilis CAS15, has a biocontrol effect on Fusarium wilt and promotes the growth of pepper. Eur. J. Soil Biol. 2011, 47, 138–145. [Google Scholar] [CrossRef]
- Santoyo, G.; Valencia-Cantero, E.; Orozco-Mosqueda, M.C.; Peña-Cabriles, J.J.; Farias-Rodríguez, R. Role of siderophores in antagonic activity of Pseudomonas fluorescens ZUM80 against plant fungi. Terra Latinoam. 2010, 10, 53–60. [Google Scholar]
- Sanz, F.M.I.; Benuzzi, D.; Calvente, V.; Calvo, J.; Sansone, G.; Cerutti, S.; Raba, J. The use of siderophores for improving the control of postharvest in stored fruits and vegetables. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Méndez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2013; pp. 1385–1394. [Google Scholar]
- Acebedo, G.Y.; Hernández-Rodríguez, A.; Rives, R.N.; Velázquez del Valle, M.G.; Hernández-Lauzardo, A.N. Perspectivas del uso de bacterias rizosféricas en el control de Pyricularia grisea (Cooke Sacc.) en el cultivo del arroz (Oryza sativa L). Rev. Colomb. Biotecnol. 2011, 13, 16–22. [Google Scholar]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruits: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Foods Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Seidl, V. Chitinase of filamentous fungi a large group old the diverse proteins with multiple physiological functions. Fungal Biol. Rev. 2008, 22, 36–42. [Google Scholar] [CrossRef]
- Stoykov, Y.M.; Paulov, A.I.; Krastanov, A.I. Chitinase biotechnology: Production, purification and application. Eng. Life Sci. 2015, 15, 30–38. [Google Scholar] [CrossRef]
- Barret, A.J.; Rawlings, N.D.; Woesnner, J.F. The Handbook of Proteolytic Enzymes, 2nd ed.; Academic Press: London, UK, 2003. [Google Scholar]
- Yu, T.; Wang, L.; Yin, Y.; Wang, Y.; Zheng, X. Effect of chitin on the antagonistic activity of Cryotococcus laurentii against Penicillium expansum in pear fruit. Int. J. Food Microbiol. 2008, 122, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Shivakumar, S.; Karmali, A.N.; Ruhimbana, C. Partial purification, characterization, and kinetic studies of a low-molecular-weight, alkali-tolerant chitinase enzyme Bacillus subtilis JN032305, a potential biocontrol strain. Prep. Biochem. Biotechnol. 2014, 44, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, P.; Tozlu, E.; Kotan, R.; Kotan, M.S. Potential of some bacteria for biological control of postharvest citrus green mold caused by Penicillum digitatum. Plant Prot. Sci. 2017, 53, 1–10. [Google Scholar]
- Chi, M.; Li, G.; Liu, Y.; Liu, G.; Li, G.; Li, M.; Zhang, X.; Sun, Z.; Sui, Y.; Liu, J. Increase in antioxidant enzyme activity, stress tolerance and biocontrol efficacy of Pichia kudriavzevii with the transition from a yeast-like to biofilm morphology. Biol. Control 2015, 90, 113–119. [Google Scholar] [CrossRef]
- Che, Y.; Yan, F.; Chai, Y.; Lu, H.; Kolter, R.; Losick, R.; Guo, J.H. Biocontrol of tomato wilt disease by Bacillus subtilis isolates from natural environments depends on conserved genes mediating biofilm formation. Environ. Microbiol. 2013, 15, 848–864. [Google Scholar]
- Beauregard, P.B.; Chai, Y.; Vlamakis, H.; Losick, R.; Kolter, R. Bacillus subtilis biofilm induction by plant polysaccharides. Proc. Natl. Acad. Sci. USA 2013, 110, E1621–E1630. [Google Scholar] [CrossRef] [PubMed]
- Vlamakis, H.; Aguilar, C.; Losick, R.; Kolter, R. Control of cell fate the formation of an architecturally complex bacterial community. Genes Dev. 2008, 22, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, A.; Meheter, A.; Prescitt, A.; Kiley, T.B.; Stanley-Wall, N.R. YuaB functions synergistically with the exopolysaccharide and TasA amyloid fibers to allow biofilm formation by Bacillus subtilis. J. Bacteriol. 2011, 193, 4821–4831. [Google Scholar] [CrossRef] [PubMed]
- Branda, S.S.; Chu, F.; Kearns, D.B.; Losick, R.; Kolter, R. A major protein component of the Bacillus subtilis biofilm matrix. Mol. Microbiol. 2006, 59, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Romero, D.; Vlamakis, H.; Losick, R.; Kolter, R. An accessory protein required for anchoring and assembly of amyloid fibers in B. subtilis. Mol. Microbiol. 2011, 80, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Branda, S.S.; Gonzales-Pastor, J.E.; Ben-Yehuda, S.; Losick, R.; Kolter, R. Fruiting body formation by Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2001, 98, 11621–11626. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Iwano, M. BsIA (YuaB) forms a hydrophobic layer on the surface on the Bacillus subtilis biofilms. Mol. Microbiol. 2012, 85, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Kaufrnan, G.; Liu, W.; Williams, D.M.; Choo, Y.; Gopinadhan, M.; Samudrala, N.; Sarfati, R.; Yan, E.C.Y.; Regan, L.; Osuji, C.O. Flat drops, elastic sheets, and microcapsules by interfacial assembly of a bacterial biofilm protein, BsIA. Langmuir 2017, 33, 13590–13597. [Google Scholar]
- Haggag, W.M.; Timmusk, S. Colonization of peanut roots by biofilm-forming Paenibacillus polymyxa initiates biocontrol against crown rot diseases. J. Appl. Microbiol. 2008, 104, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Bianciotto, V.; Andreotti, S.; Balstrini, R.; Bonfante, P.; Perotto, S. Mucoide mutants of the biocontrol strain Pseudomonas fluorescens CHA0 show increased ability in biofilm formation on mycorrhizal and nomycorrhizal carrot roots. Mol. Plant Microbe Interact. 2001, 14, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Fall, R.; Vivanco, J.M. Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomona syringae is facilitated by biofilm formation and surfaction production. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.; Solsana, C.; Viñas, I.; Usall, J.; Plaza, P.; Teixidó, N. Optimization of packaging and storage conditions of a freeze dried Pantoea agglomerans formulation for controlling postharvest diseases in fruit. J. Appl. Microbiol. 2014, 117, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Batista, F.M.; Duarte, M.M.H.; Tremocoldi, A.R.; Pascholati, S.F. Potential of antimicrobial volatile organic compounds to control Sclerotinis sclerotiorum in vean seeds. Pesq. Agropec. Bras. 2011, 4, 137–142. [Google Scholar]
- Guevara-Avendaño, E.; Bejarano-Bolívar, A.A.; Kiel-Martínez, A.L.; Ramírez-Vázquez, M.; Méndez-Bravo, A.; von Wobeser, E.A.; Sanchez-Rangel, D.; Guerrero-Analco, J.A.; Eskalen, A.; Reverchon, F. Avocado rhizobacteria emit volatile organic compounds with antifungal activity against Fusarium solani, Fusarium sp. associated with Kuroshio shot hole borer, and Colletotrichum gloeosporioides. Microbiol. Res. 2019, 219, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Layton, C.; Maldonado, E.; Monroy, L.; Corrales, L.C.; Sánchez, L.C. Bacillus spp.; perspectiva de su efecto biocontrolador mediante antibiosis en cultivos afectados por fitopatógenos. Cienc. Bioméd. 2011. [Google Scholar] [CrossRef]
- Velázquez-Becerra, C.; Macías-Rodríguez, L.I.; López-Bucio, J.; Flores-Cortez, I.; Santoyo, G.; Hernández-Soberano, C.; Valencia-Cantero, E. The rhizobacterium Arthrobacter agilis produces dimethylhexadecylamine, a compound that inhibits growth of phytopathogenic fungi in vitro. Protoplasma 2013, 250, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Raza, W.; Ling, N.; Yang, L.; Huang, Q.; Shen, Q. Response of tomato wilt pathogen Ralstonia solanacearum to the volatile organic compounds produced by biocontrol strain Bacillus amyloliquefaciens SQR-9. Sci. Rep. 2015, 6, 24856. [Google Scholar] [CrossRef] [PubMed]
- Jamalizadeh, M.; Etebarian, H.R.; Aminian, H.; Alizadeh, A. A review of mechanisms of action of biological control organisms against post-harvest fruit spoilage. Bull. OEPP/EPPO 2011, 41, 65–71. [Google Scholar] [CrossRef]
- Hacquard, S.; Spaepen, S.; Garrido-Oter, R.; Schulze-Lefert, P. Interplay between innate immunity and the plant microbiota. Annu. Rev. Phytopathol. 2017, 55, 565–589. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Nolan, T.M.; Song, G.; Liu, S.; Xie, Z.; Chen, J.; Schnable, P.S.; Walley, J.W.; Yin, Y. FERONIA receptor kinase contributes to plant immunity by suppressing jasmonic acid signaling in Arabidopsis thaliana. Curr. Biol. 2018, 28, 3316–3324. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Xia, W.; Cao, P.; Xiao, Z.A.; Zhang, Y.; Liu, M.; Zhan, C.; Wang, N. Integrated transcriptome analysis reveals plant hormones jasmonic acid and salicylic acid coordinate growth and defense responses upon fungal infection in poplar. Biomolecules 2019, 9, E12. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, B.; Marques, A.; Ramos, C.; Neng, R.N.; Nogueira, M.F.J.; Saravia, A.J.; Nunes, L.M. Chemical compositions and antibacterial and antioxidant properties of commercial essential oils. Ind. Crop. Prod. 2013, 43, 587–595. [Google Scholar] [CrossRef]
- Holert, J.; Cardenas, E.; Bergstrand, L.H.; Zaikova, E.; Hahn, A.S.; Hallam, S.J.; Mohn, W.W. Metagenomes Reveal global distribution of bacterial steroid catabolism in natural, engineered, and host environments. mBio 2018, 9, e0234517. [Google Scholar] [CrossRef] [PubMed]
- Droby, S.; Wisniewski, M.; Teixidó, N.; Spadaro, D.; Jijakli, M.H. The science, development, and commercialization of postharvest biocontrol products. Postharvest Biol. Technol. 2016, 122, 22–29. [Google Scholar] [CrossRef]
- Blachinsky, D.; Antonov, J.; Bercovitz, A.; El-ad, B.; Feldman, K.; Husid, A.; Lazare, M.; Marcov, N.; Shamai, I.; Droby, S.; et al. Commercial applications of shemer for the control of pre- and post-harvest diseases. IOBC/WPRS Bull. 2007, 30, 75–78. [Google Scholar]
- Teixidó, N.; Torres, R.; Viñas, I.; Abadias, M.; Usall, J. Biological control of postharvest diseases in fruit and vegetables. In Protective Cultures, Antimicrobial Metabolites and Bacteriophages for Food and Beverage Biopreservation; Woodhead Publishing: Cambridge, UK, 2011; pp. 364–402. [Google Scholar]
- Janisiewicz, W.J.; Korsten, L. Biological control of postharvest diseases of fruits. Ann. Rev. Phytopathol. 2002, 40, 411–441. [Google Scholar] [CrossRef] [PubMed]
- Janisiewicz, W.J.; Peterson, D.J. Susceptibility of the stem pull area of mechanically harvest apples to blue mold decay and its control with a biocontrol agent. Plant Dis. 2004, 88, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Demoz, B.T.; Korsten, L. Bacillus subtilis attachment, colonization, and survival on avocado flowers and its mode of action on stem-end rot pathogens. Biol. Control 2006, 37, 68–74. [Google Scholar] [CrossRef]
- Lahlali, R.; Raffaele, B.; Jijakli, M.H. UV protectants for Candida oleophila (strain O), a biocontrol agent of postharvest fruit diseases. Plant Pathol. 2011, 60, 288–295. [Google Scholar] [CrossRef]
- Massart, S.; Jijakli, M.H. Pichia anomala and Candida oleophila in biocontrol of postharvest diseases of fruits: 20 years of fundamental and practical Research. In Post-Harvest Pathology, Plant Pathology in the 21st Century; Prusky, D., Gullino, M.L., Eds.; Springer International Publishing: Basel, Switzerland, 2014; pp. 111–122. [Google Scholar]
- Lima, G.; Sanzani, S.M.; Curtis, F.; Ippolito, A. Biological control of postharvest diseases. In Advances in Postharvest Fruit and Vegetables Technology; Wills, R.B.H., Golding, J., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 65–81. [Google Scholar]
- Usall, T.; Torres, R.; Teixidó, N. Biological control of postharvest diseases on fruit a suitable alternative? Curr. Opin. Food Sci. 2016, 11, 51–55. [Google Scholar] [CrossRef]
- Maida, I.; Chiellini, C.; Mengoni, A.; Bosi, E.; Firenzuoli, F.; Fondi, M.; Fani, R. Antagonistic interactions between endophytic cultivable bacterial communities isolated from the medicinal plant Echinacea purpurea. Environ. Microbiol. 2016, 18, 2357–2365. [Google Scholar] [CrossRef] [PubMed]
- Goudjal, Y.; Toumatia, O.; Yekkour, A.; Sabaou, N.; Mathieu, F.; Zitouni, A. Biocontrol of Rhizoctonia solani damping-off and promoting of tomato plant growth by endophytic actinomycete isolated from native plants of Algerian Sahara. Microbiol. Res. 2014, 169, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Chenniappan, C.; Narayanasamy, M.; Daniel, G.M.; Ramaraj, G.B.; Ponnusamy, P.; Sekar, J.; Ramalingam, P.V. Biocontrol efficiency of native plant growth promoting rhizobacteria against rhizome rot disease of turmeric. Biol. Control 2019, 129, 55–64. [Google Scholar] [CrossRef]
- Larran, S.; Simón, M.R.; Moreno, M.V.; Santamarina, S.M.P.; Perelló, A. Endophytes from wheat as biocontrol agents against tan spot disease. Biol. Control 2016, 92, 17–23. [Google Scholar] [CrossRef]
- Comby, M.; Gacoin, M.; Robineau, M.; Rabenoelina, F.; Ptas, S.; Dupont, J.; Prozi, C.; Baillieul, F. Screening of wheat endophytes as biological control agents against Fusarium head blight using two different in vitro test. Microbiol. Res. 2017, 202, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda, M.C.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Hu, G.; Yu, J.; Zhu, X.; Lin, Y.; Chen, S.; Yuan, J. Statistical Research on the bioactivity of new marine natural products discovered during the 28 years from 1985 to 2012. Mar. Drugs 2015, 13, 202–221. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.M.C.; Polonio, J.C.; Quecine, M.C.; Almeida, T.T.; Bogas, A.C.; Pamphile, J.A.; Pereira, J.O.; Asrolfi-Fiho, S.; Azevedo, J.L. Endophytic cultivable bacteria community obtained from the Paullinia cupana seed in Amazonas and Bahia regions and its antagonistic effects against Colletotrichum gloeosporioides. Microb. Pathog. 2016, 98, 16–22. [Google Scholar]
- Sinimol, S.; Sarika, A.R.; Nair, A.J. Diversity and antagonistic potential of marine microbes collected from south-west coast of India. 3 Biotech. 2016, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Bibi, F.; Nasser, M.I.; Hassan, A.M.; Yasir, M.; Al-Ghamdi, A.A.K.; Azhar, E.I. Diversity and antagonistic potential of bacteria isolated from marine grass Halodule uninervis. 3 Biotech. 2018, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Montiel, L.G.; Larralde-Corona, C.P.; Vero, S.; Lopez-Aburto, M.G.; Ochoa, J.L.; Ascencio-Valle, F. Characterization of yeast Debaryomyces hansenii for biological control of blue mold decay of Mexican lemon. CyTA J. Food. 2010, 8, 49–56. [Google Scholar] [CrossRef]
- Aloui, H.; Liccardello, F.; Khawldia, K.; Hamdi, M.; Restuccia, C. Physical properties and antifungal activity of bioactive films containing Wickerhamomyces anomalus killer yeast and their application for preservation of oranges and control of postharvest green mold caused by Penicillium digitatum. Int. J. Food Microbiol. 2015, 200, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Garcia, T.; Murillo-Amador, B.; Nieto-Garibay, A.; Chiquito-Contreras, R.; Rincon-Enriquez, G.; Hernandez-Montiel, L.G. Effect of ulvan on the biocontrol activity of Debaryomyces hansenii and Stenotrophomonas rhizophila against fruit rot of Cucumis melo L. Agronomy 2018, 8, 273. [Google Scholar] [CrossRef]
- Bibi, F.; Strobel, G.A.; Naseer, M.I.; Yasir, M.; Khalaf Al-Ghamdi, A.A.; Azhar, E.I. Halophytes-associated endophytic and rhizospheric bacteria: Diversity, antagonism and metabolite production. Biocontrol Sci. Technol. 2018, 28, 192–213. [Google Scholar] [CrossRef]
- Cong, Z.; Huang, X.; Liu, Y.; Liu, Y.; Wang, P.; Liao, S.; Yang, B.; Zhou, X.; Huang, D.; Wang, J. Cytotoxic anthracycline and antibacterial tirandamycin analogues from a marine-derived Streptomyces sp. SCSIO 41399. J. Antibiot. 2019, 72, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Gomes, E.D.B.; Dias, L.R.L.; Rita de Cassia, M. Actinomycetes bioactive compounds: Biological control of fungi and phytopathogenic insect. Afr. J. Biotechnol. 2018, 17, 552–559. [Google Scholar]
- Lopes, M.R.; Nadjara, K.M.; Pompeo, F.L.; Silva, A.C.; Kupper, K.C. Saccharomyces cerevisiae: A novel and efficient biological control agent for Colletotrichum acutatum during pre-harvest. Microbiol. Res. 2015, 175, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.Y.; De Costa, D.M. Potential of pre-harvest application of Burkholderia spinosa for biological control of epiphytic and pathogenic microorganisms on the phyllosphere of banana (Musa spp.). Trop. Agric. Res. 2014, 25, 443–454. [Google Scholar] [CrossRef]
- Camañas, T.P.; Viñas, I.; Usall, J.; Torres, R.; Anguera, M.; Teixidó, N. Control of postharvest diseases on citrus fruit by preharvest applications of biocontrol agent Pantoea agglomerans CPA-2 Part II. Effectiveness of different cell formulations. Postharvest Biol. Technol. 2008, 49, 96–106. [Google Scholar] [CrossRef]
- Qin, X.; Xiao, H.; Xue, C.; Yu, Z.; Yang, R.; Cai, Z.; Si, L. Biocontrol of gray mold in grapes with the yeast Hanseniaspora uvarum alone and in combination with salicylic acid or sodium bicarbonate. Postharvest Biol. Technol. 2015, 100, 160–167. [Google Scholar] [CrossRef]
- Han, J.; Chen, D.; Huang, J.; Li, X.; Zhou, W.W.; Gao, W.; Jia, Y. Antifungal activity and biocontrol potential of Paenibacillus polymyxa HT16 against white rot pathogen (Coniella diplodiella Speq.) in table grapes. Biocontrol Sci. Technol. 2015, 25, 1120–1132. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).