1. Introduction
Maize (
Zea mays L.) is a major crop for both human consumption and animal feed in Sub-Saharan Africa (SSA). Drought (DS) and heat stress (HTS) are two major limiting factors affecting maize productivity in the tropical lowlands, where erratic rains and increased temperatures are becoming of frequent occurrence [
1]. It is likely that high temperatures will occur more often and will last longer and extreme precipitations events will become more intense and frequent in many regions [
2]. High temperatures and changes in rainfall pattern can cause significant decline in maize yields under rainfed conditions in the tropical region with SSA being one of the worst affected areas [
3,
4]. An increase in temperature over 30 °C reduces grain yield by 1% under optimal rain-fed condition and by 1.7% under DS [
5] and up to 40% under combined drought and heat stress [
6].
With the current changes in climatic conditions, it is projected that by 2030 drought and higher temperatures may render 40% of the current maize growing areas in Africa unsuitable for varieties available today [
7]. Unless strong adaptation measures are taken, these changes are expected to reduce yields of maize and other food crops by 10% to 20%, causing marked decline in human welfare [
8,
9]. Adaptation to climate change may involve the use of crop varieties that are endowed with tolerance to higher temperatures and drought, and resistance to emerging pests and diseases [
9]. Approaches that improve the performance of maize genotypes under combined drought and heat stress (DSHTS) are therefore essential to sustain productivity and to avoid widespread famine in Africa.
Maize is more sensitive than other cereals to drought at flowering because anthers and silks are separated by about 1 m, and pollen and stigma are exposed to the environment [
10,
11]. The crop is particularly sensitive to DS one week before and two weeks after flowering [
12,
13], resulting in an average yield loss of 20% to 50% [
14]. On the other hand, HTS in maize is associated with temperatures rising above an optimum of 30 °C that adversely affect physiological processes [
15], resulting in increased respiration, reduced life cycle, light interception, photosynthesis, and increased pollen sterility [
16]. High temperature two weeks before flowering causes permanent tissue injury to developing leaves resulting in leaf firing [
15], accelerate the rate of leaf senescence [
17], and premature lodging. High temperature occurring at the on-set of flowering causes tassel blasting that reduces pollen production, pollen viability and pollination rate [
18,
19]. At grain-filling stage, high temperatures lead to slow and short grain-filling period, reduced kernel set, and grain weight [
18,
19,
20], resulting in 10% reduction in grain yield [
21].
Drought and heat stress occur simultaneously in farmers’ fields when the growing season is hot, and the distribution of rains is erratic. Heyne & Brunson [
22] showed that the combined effect of drought and heat stress in maize was much greater than the effect of each stress separately. The simultaneous occurrence of drought and heat stresses in farmers’ fields are becoming increasing common in the tropical lowlands, particularly in maize growing areas in West Africa and Sahelian regions [
1] and Eastern and Southern Africa [
4]. These regions are known to be vulnerable to climate changes due to high climate variability, high reliance on rainfed agriculture, and limited economic and institutional capacity to respond to climate change [
1]. Depending on the timing, duration and severity, yield losses in maize due to DSHTS can vary from 15% to 40% and they can exceed 70% [
6,
23]. Heiniger [
24] reported that DSTHTS occurring during pollination can cause up to 100% yield loss in maize. Tassel is more susceptible to heat than drought stress because of its position that exposes pollen to heat, whereas drought is more detrimental to silk than heat stress causing delay in silk emergency [
3,
25]. Sullivan & Blum [
26] observed that maize closes its stomata sooner than sorghum (
Sorghum bicolor L.) under DS thereby increasing the canopy temperature by more than 9 °C [
27]. Maize genotypes capable of keeping their stomata partially open may maintain enough transpirational cooling to avoid excessively high leaf temperature.
The breeding strategies in maize that have been adopted so far have focused on improvement of each stress separately. Preliminary results of hybrids evaluated under the Drought Tolerant Maize for Africa (DTMA) project revealed that tolerance to DS does not necessarily confer tolerance to HTS or DSHTS, which have implications to breeding for adaptation to climate change in maize production systems in SSA [
4]. Understanding the response of hybrids selected for drought tolerance (DT) to DSHTS may thus facilitate the identification of promising hybrids with tolerance to DSHTS fairly quickly for further testing, release and deployment. Parents of these hybrids can also be screened under DSHTS to identify source materials for breeding to further increase their tolerance to much higher levels. This study was, therefore, conducted to (i) assess the extent of variation in tolerance to DSHTS from among the existing best DT hybrids; (ii) examine the response patterns of the hybrids to DSHTS; (iii) identify traits that contributed to better performance under DSHTS; and (iv) select the best hybrids with tolerance to DSHTS stress.
2. Materials and Methods
2.1. Genetic Material and Experimental Design
The genetic material that was used in this study consisted of 29 promising DT three-way cross maize hybrids, 10 commercial hybrid checks, and a local variety commonly grown by farmers in the area. These hybrids have been evaluated in regional trials in partnership with the National Agricultural Research System (NARS) in West and Central Africa (WCA) for over three years (
Table 1).
2.2. Weather Conditions during the Trial
The trial under DSHTS was conducted during the dry season in 2013, 2014, and 2015. In each year, the lowest day and night temperatures were recorded in February at the time of planting and the highest in April. During evaluation of this trial, there were incidences of rains in the months of April and May, but the amounts were not sufficient to have significant effects on the intensity of drought stress under elevated temperature. In each year, the peak of high temperature occurred in April (
Figure 1), which coincided with the reproductive and grain-filling stages of the maize crop. During this period, the maximum day and night temperatures were 45 and 31 °C, respectively, which were detrimental to fertilization, seed setting, and grain-filling in maize. Minimum day and night temperatures of ≤27 °C and ≤20 °C, respectively, are considered to be optimal for the growth and development of maize in the tropical lowlands.
2.3. Experimental Design
The 40 hybrids including a local check was arranged in 8 × 5 alpha lattice design (incomplete design) with three replications evaluated for three years (2013–2015) under DSHTS and for two years (2014 and 2015) under DS and WW. Each replicate consisted of eight incomplete blocks with five plots in each block. In each year, the genetic materials, experimental design, and replications were the same. Each entry was planted in a two-row 5 m plot spaced 0.75 m apart with 0.50 m spacing between hills within a row. Three seeds were planted in a hill and thinned to two plants per hill three weeks after sowing to attain a population density of 53,000 plants per hectare (ha−1). A compound fertilizer was applied at the rates of 60 kg N, 60 kg P, and 60 kg K ha−1 at the time of planting. An additional 60 kg N ha−1 was applied as top dressing four weeks later. Gramoxone and atraxine were applied as pre-immergence herbicides at the rates of 1.5 L gramoxone and 2.5 L atraxine in 200 L of water ha−1. Subsequent manual weeding was done to keep the trials weed free.
2.4. Evaluation under Combined Drought and Heat Stress
The 40 maize hybrids were evaluated for tolerance to DSHTS at Kadawa (11°39′ N, 8°27′ E, elevation 500 m) in Kano State in Nigeria, where hot and dry weather occur between February and June. The soil type is Regosols, with mainly sandy to clay loam texture, pH of 5.9, organic carbon 4.3 g kg
−1, and residual nitrogen 0.24 g kg
−1 [
28]. In Kadawa, the temperature during the dry season ranges from 33 to 45 °C. This helped in conducting this trial under DSHTS, whereby water supply was carefully controlled through furrow irrigation. The flowering and grain filling periods were generally in April when rainfall incidences were negligible and relative humidity ranged from 15% to 46%, allowing for exposure of the hybrids to drought and elevated temperatures (
Figure 1).
Maize seeds were planted in mid-February for three consecutive years (2013 to 2015) and flowered in mid-April, the hottest period at Kadawa. The trial was initially grown under well-watered conditions prior to exposure to drought and heat stress. A gravity irrigation system was used to supply sufficient water through furrows to the crop every four days during the first 45 days after planting. Thereafter, irrigation was withdrawn in mid-April when mean day temperature ranged from 36 to 45 °C and night temperature varied from 27 to 30 °C, inducing the combined effects of drought and heat stress that coincided with flowering. Irrigation was resumed after 21 days and it was applied only once a week to allow grain filling of the crop until physiological maturity. Meteorological data were recorded during the trial period using an automated weather station at Bagauda in Kano State. As shown in
Table 2, the amount of precipitation received during the period when the trial was exposed to heat and drought stress each year was very small.
2.5. Evaluation under Drought Stress
The same set of hybrids were evaluated under controlled DS and well-watered (WW) conditions at Ikenne (6°52′ N, 3°43′ E, altitude 60 m) in Nigeria during the dry season. The experiment was conducted in two adjacent blocks separated by 50 m to prevent spill-over of irrigation water. Maize seeds were planted in mid-November in 2014 and 2015, so that flowering occurs in mid-January where incidences of rainfall were minimal. A sprinkler irrigation system was used to provide sufficient water every week during the first 35 days after planting. Thereafter, irrigation was withdrawn from DS block to induce drought stress at flowering and grain filling stages, allowing for the crop to mature on stored soil moisture. The WW block continued to receive irrigation once a week until physiological maturity.
2.6. Data Collection
Data were recorded on different traits in each plot. Days to anthesis and silking were recorded as the number of days from planting to when 50% of the plants in a plot were shedding pollen and had emerged silks, respectively. Data on tassel blast (TB) and barren plants (BP) were only recorded on DSHTS trial. TB was rated during flowering on a scale of 1 to 5, where 1 = few plants showing tassel blast and 5 = all plants showing severe tassel blast, whereas BP was rated on a scale of 1 to 5, where 1 = few plants without ears and 5 = more than 50% or all plants bearing no ears recorded in each plot two weeks after flowering. Anthesis to silkin interval (ASI) was computed as a difference between days to silking and anthesis. Plant and ear heights were recorded on five randomly selected plants, as distance from the ground to the base of the tassel and the upper ear, respectively. Leaf death score (leaf senescence) was rated on a scale of 1 to 10, where 1 = 10% dead leaf area and 10 = 100% dead leaf area recorded only in DS and DSHTS trials. Number of ears per plant (EPP) was computed as the proportion of the total number of ears harvested divided by the total number of plants at harvest. Plant aspect was visually scored on a scale of 1 to 5, where 1 = excellent overall phenotypic appeal and 5 = poor overall phenotypic appeal. Similarly, ear aspect was scored on a scale of 1 to 5, where 1 = clean, uniform, large, and well-filled ears and 5 = rotten, variable, small, and partially filled ears. All ears harvested from each plot were shelled and used to determine the percentage grain moisture and grain weight. Grain yield adjusted to 15% moisture was computed from the grain weight recorded form each plot.
2.7. Data Analysis
The analysis of data was performed using a mixed model procedure in SAS [
29] version 9.4, following the standard linear mixed model that was described by Vargas et al. [
30]. All effects except genotypes were considered random and Best Linear Unbiased Predictor (BLUP) was computed for all traits using the procedure of Robinson [
31]. Repeatability estimates were computed to determine the precision of measuring traits in the trials.
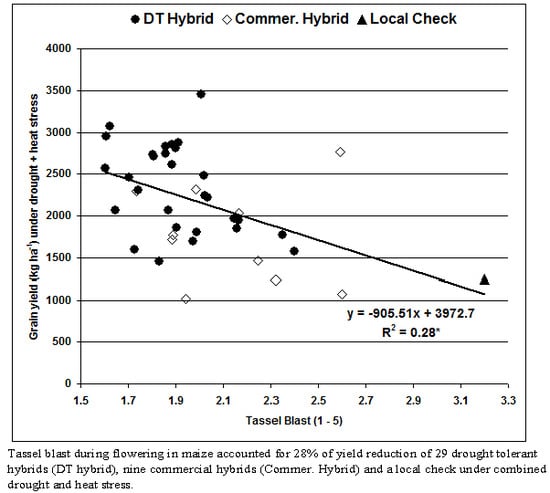
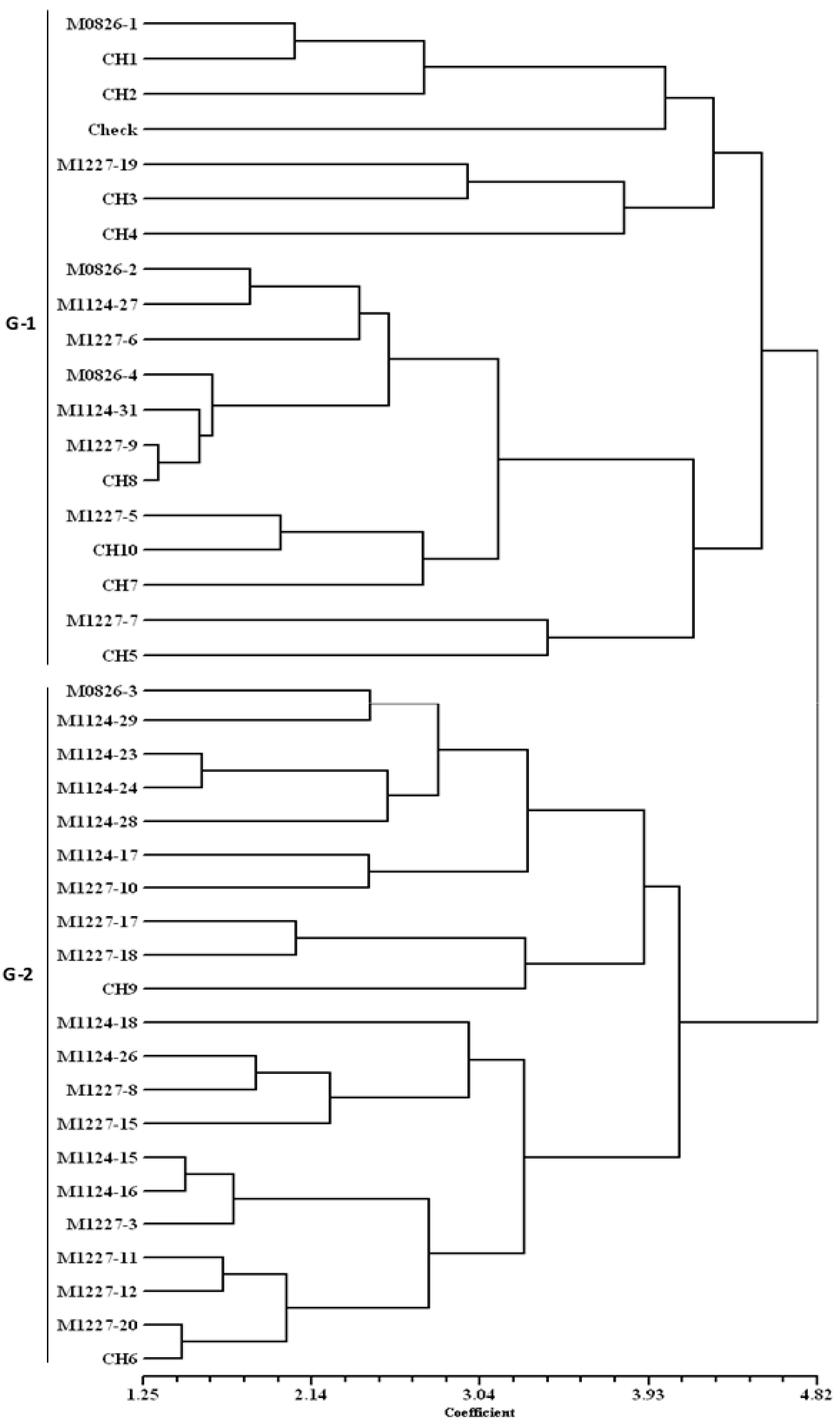
Cluster analysis using Euclidean distance with standardized values of agronomic data was computed to classify the DT hybrids into groups. A dendrogram was constructed using the Unweighted Pair Group Method with Arithmetic means (UPGMA) and the resulting tree was used to determine the association between the DT hybrids under DS and DSHTS. Phenotypic coefficients of correlations for grain yield with secondary traits were computed to further determine the secondary traits contributing to grain yield. Grain yields and TB scores were plotted on a graph to show the trend of yields of the hybrids under DSHTS conditions.
4. Discussion
When considering the importance of identifying maize genotypes with high levels of tolerance to combined drought and heat stress for climate change adaptation, we screened promising drought tolerant hybrids under DS and DSHTS. The significant differences found among DT hybrids for grain yield and several other agronomic traits recorded under both DS and DSHTS, indicate the presence of genotypic difference among promising DT hybrids. The significant interactions of hybrids with environments for grain yield and other agronomic traits suggest the influence of DS and DSHTS on the expression of these traits. The greatest influence of both DS and DSHTS occurred in mid-April each year, during the three weeks around flowering and grain-filling stages. These two stages are the most critical period determining yield potential in maize.
The extent of yield loss is dependent on severity of drought and heat stress, field environment and the maize genotypes under study. In this study, drought stress reduced grain yield by 58%, while DSHTS reduced grain yield by 77%, agreeing with previous findings that demonstrated more yield loss from combined effects of drought and heat stress than drought stress alone [
6,
22,
23]. Combined drought and heat stress adversely affected grain yield and days to flowering of most hybrids more than drought, possibly due to its adverse effects on pollen production, or ovule fertility, leading to premature embryo abortion and reduced grain weight [
32]. Our results corroborate earlier findings [
20,
32,
33] that the effect of DSHTS was higher in maize at the reproductive stage than at the vegetative stage.
Previous studies in maize [
4,
34] and wheat (
Triticum aestivum L.) [
35] showed that tolerance to DSHTS is genetically distinct from tolerance to individual drought and heat stresses. However, our results indicated that breeding for DS, to some extent, has spill-over effect under DSHTS, with 50% of the best DT hybrids selected under DS showing appreciable grain yields under DSHTS. Some DT hybrids that produced high grain yields under both DS and DSHTS were found in the present study, implying that in a short term, breeders should screen available DT varieties under DSHTS to identify those with high levels of tolerance to combined drought and heat stress quickly for further testing and release, as well as for use as donors of DSHTS.
The observed significant correlations between the same traits measured under DS, DSHTS as well as WW conditions suggest the presence of common genetic factors controlling expression of these traits under the three growing conditions. However, the observed weak correlations between grain yield under DS and DSHTS indicate the presence of independent genetic factors controlling yield performance under these conditions, which agrees with previous studies [
4,
34]. The significant association of grain yield under DS with yield under WW observed in our study is consistent with the findings of other researchers [
4,
36]. This result, however, contradicts the findings of Trachsel et al. [
34] who reported very weak correlation (
r = 0.075) between grain yield under optimum and that under stressed conditions.
The strong correlation between grain yield and TB and BP demonstrated that these traits together with other secondary traits (ASI, EPP, ear and plant aspect cores, days to anthesis, and silking) should be included in an index to select promising genotypes under DSHTS. Secondary traits that are strongly correlated with grain yield suggest that they function together in determining the plants’ response to combined stress [
37]. Although we found weak (0.13) repeatability for TB relative to grain yield under DSHTS in our study, this trait has the potential to improve the selection efficiency for grain yield under DSHTS. Other researchers [
38] also found weak heritability for ASI relative to grain yield, but suggested its use for selection under DS [
39,
40,
41,
42].
Knowledge of the relationships among DT hybrids developed from diverse genetic backgrounds can help to identify a set of specific DT hybrids that have good levels of tolerance to DS and DSHTS. Grain yield and morphological (adaptive secondary) traits were used to estimate these relationships. Under DSHTS, group G-2 clustered DT hybrids tolerant to DSHTS, including two commercial hybrids (CH6 and CH9), which were competitive with the best DT hybrid, suggesting that these hybrids had common adaptive secondary traits that were better expressed under DSHTS as predicted by dendrogram. The majority of DT hybrids and commercial hybrids grouped in G-1 showed high levels of susceptibility to DSHTS with low grain yields. However, seven of the susceptible hybrids (M1124-27, M1227-5, M1227-6, M1227-7, M1227-9, M0826-1, CH5), including a released hybrid and commercial hybrid were grouped (G-1) with best DT hybrids under DS, suggesting that some DT hybrids that are susceptible to DSHTS perform better under DS condition.
Although we observed few DT hybrids with desirable traits grouped together in the dendrogram, most desirable agronomic traits were not found in a single DT hybrid, suggesting the need for multiple traits pyramiding by introgressing the desired traits into the genetic background of adapted DT hybrids. Secondary traits, like EPP, TB, ear and plant aspect scores, days to anthesis, and silking that were found to be strongly correlated with grain yield under stressed conditions can be combined with other traits, such as canopy temperature and leaf firing for field screening. Alam et al. [
43] suggested that traits that are indicative of reproductive success under HTS, including ASI, TB, tassel sterility, pollen viability, and stigma receptivity and other morpho-physiological traits, including leaf firing, senescence, and chlorophyll content need to be used along with grain yield for selecting stress tolerant germplasm. Most of these traits are simple and rapid to measure in large number of progenies and are used for selecting for tolerance to DSHTS.
We observed that some productive DT hybrids were tolerant to DSHTS as well as under WW conditions. However, some DT hybrids together with a commercial hybrid CH7 ranked among the worst yielders under DSHTS, with grain yields well below the trial mean, suggesting that not all DT hybrids are tolerant to DSHTS as found in other studies [
4,
34,
35]. We identified three DT hybrids, M1227-17, M0826-3, and M1124-18, among the top 10 hybrids with consistently high grain yields under both DS and DSHTS. These hybrids combined high grain yields with least TB and BP and increased EPP under stress conditions. M0826-3 is a released DT hybrid in Nigeria that also has attributes of resistance to maize streak virus and tolerance to low soil nitrogen [
44].
Although we have not designed an experiment specifically for heat stress, the level of tolerance observed in some DT hybrids under DSHTS may be attributed to the effects of the relatively high day temperatures (32–37 °C) when the hybrids used in this study were selected under DS at Ikenne during the dry season. This agrees with Sehgal et al. [
37] who observed combined tolerance to drought and heat stress in lentil (
Lens culinaris Medic.) genotypes evaluated under DS, HTS, and DSHTS. Limited information is available on performance of DT maize hybrids under DSHTS, which need further investigation to understand the relationship between tolerance to DS and DSHTS and the genetic and biological mechanisms that control tolerance to DSHTS in maize. Comprehensive study by the use of a combination of conventional and molecular breeding tools is needed to understand the mode of inheritance of adaptive secondary traits that confer tolerance to DSHTS to enhance breeding scheme for DSHTS. Genotypes showing tolerance to DSHTS will be used as a source for the development of new inbred lines. In addition, stress tolerant maize germplasm introduced from diverse sources should be screened to identify source materials for developing new multiple-stress tolerant inbred lines with higher levels of tolerance to DSHTS.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}