
Effects of Biochar on the Temporal Dynamics and Vertical Distribution of Iron and Phosphorus Under Soil Submergence



Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation and Characterization of Soil and Biochar
2.2. Soil Column Incubation Experiment
2.3. Chemical Analyses of Soil Pore Water
2.4. Sequential Extraction for Soil P Fractionation
2.5. Statistical Analysis
3. Results
3.1. Characteristics of Soil and Biochar
3.2. Temporal Dynamics of Soil Redox Potential at Different Depths
3.3. Temporal Dynamics of Pore-Water pH at Different Depths
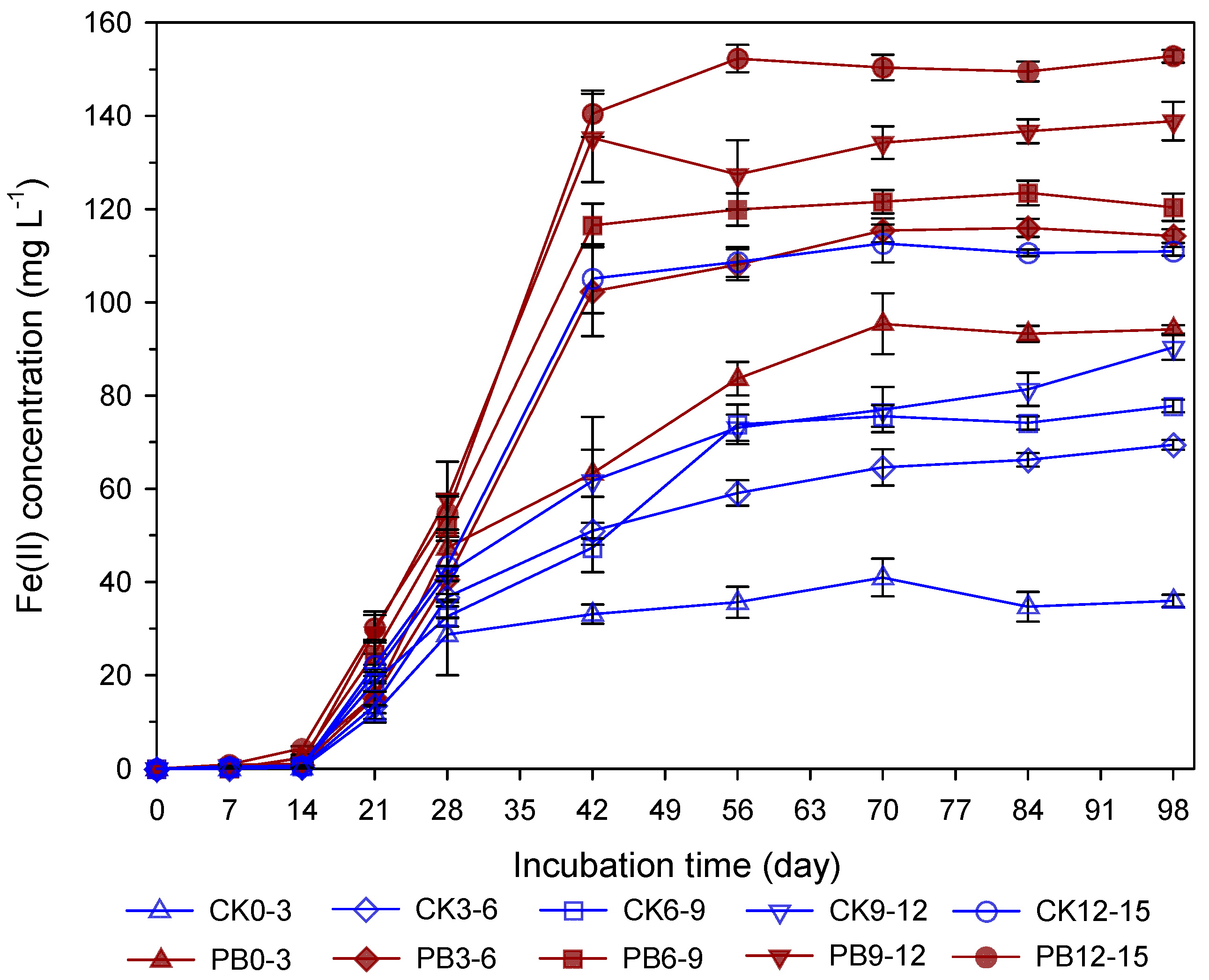
3.4. Temporal Dynamics of Pore-Water Fe(II) Concentration at Different Depths
3.5. Temporal Dynamics of Pore-Water Phosphate Concentration at Different Depths
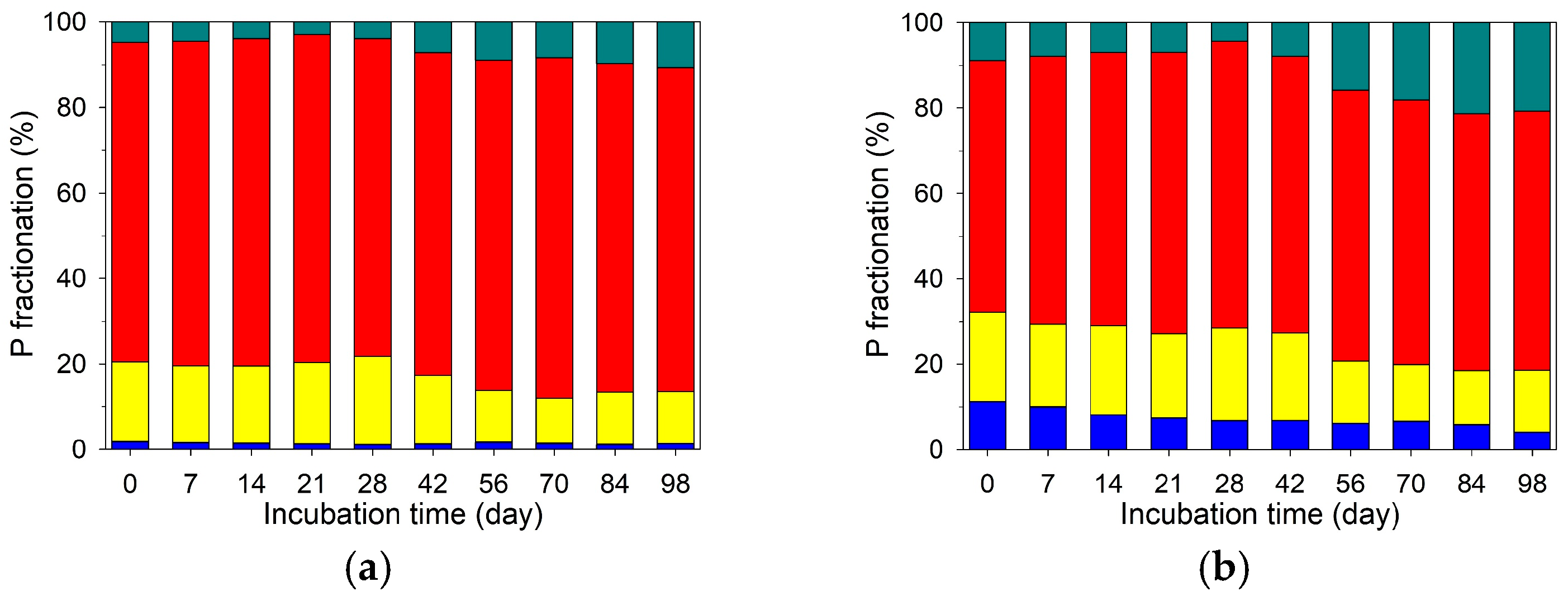
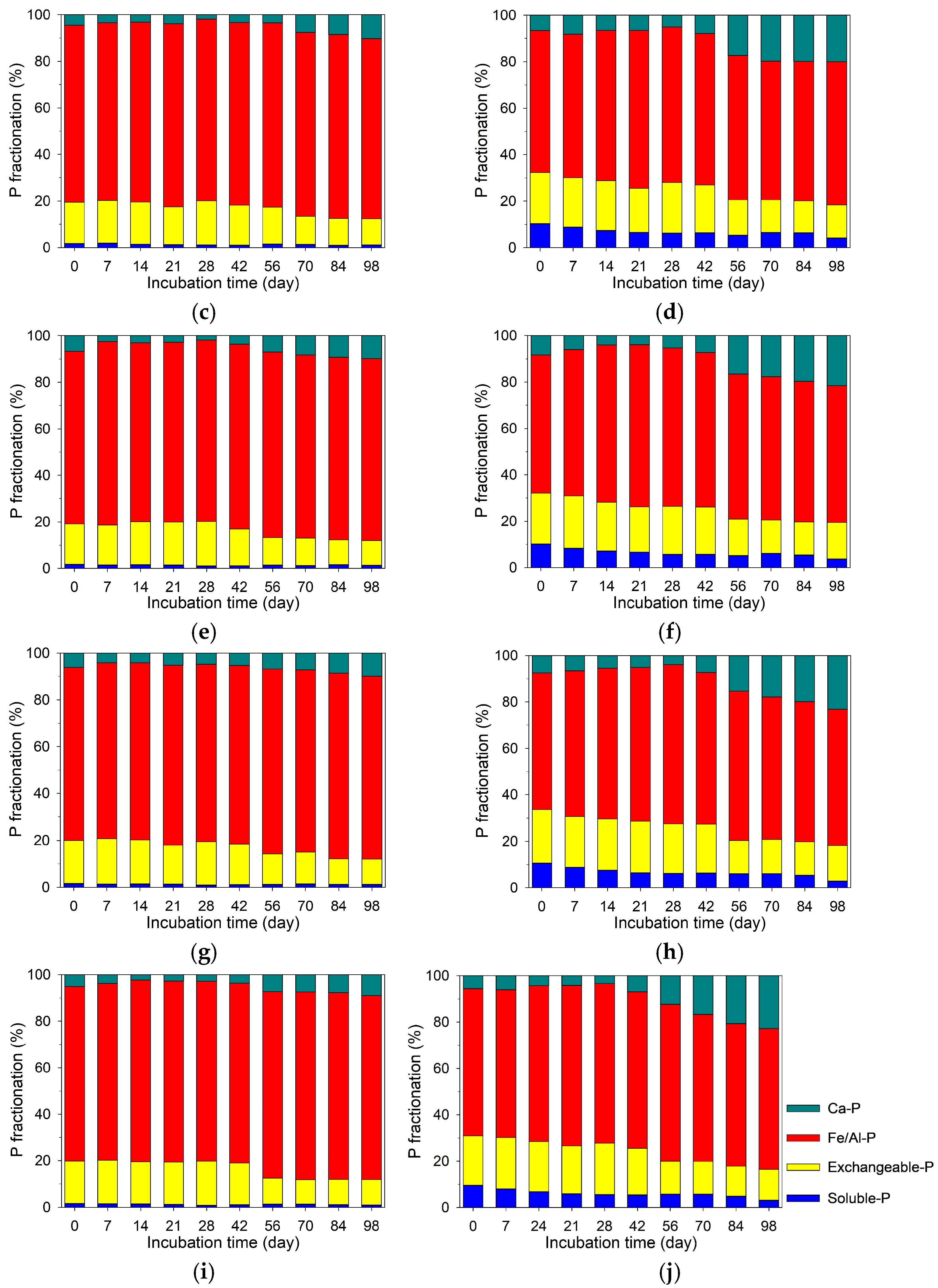
3.6. Temporal Variations in Soil P Fractionation at Different Depths
4. Discussion
4.1. Effects of Biochar on Temporal Dynamics of Soil pH and Eh at Different Depths During Soil Submergence
4.2. Effects of Biochar on Temporal Dynamics of Fe at Different Depths During Soil Submergence
4.3. Effects of Biochar on Temporal Dynamics of P at Different Depths During Soil Submergence
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luo, X.; Elrys, A.S.; Zhang, L.; Ibrahim, M.M.; Liu, Y.; Fu, S.; Yan, J.; Ye, Q.; Wen, D.; Hou, E. The global fate of inorganic phosphorus fertilizers added to terrestrial ecosystems. One Earth 2024, 7, 1402–1413. [Google Scholar] [CrossRef]
- Doydora, S.; Gatiboni, L.; Grieger, K.; Hesterberg, D.; Jones, J.L.; McLamore, E.S.; Peters, R.; Sozzani, R.; Van den Broeck, L.; Duckworth, O.W. Accessing legacy phosphorus in soils. Soil Syst. 2020, 4, 74. [Google Scholar] [CrossRef]
- Yuan, Z.; Jiang, S.; Sheng, H.; Liu, X.; Hua, H.; Liu, X.; Zhang, Y. Human perturbation of the global phosphorus cycle: Changes and consequences. Environ. Sci. Technol. 2018, 52, 2438–2450. [Google Scholar] [CrossRef] [PubMed]
- Brownlie, W.J.; Sutton, M.A.; Reay, D.S.; Heal, K.V.; Hermann, L.; Kabbe, C.; Spears, B.M. Global actions for a sustainable phosphorus future. Nat. Food 2021, 2, 71–74. [Google Scholar] [CrossRef]
- Kögel-Knabner, I.; Amelung, W.; Cao, Z.; Fiedler, S.; Frenzel, P.; Jahn, R.; Kalbitz, K.; Kölbl, A.; Schloter, M. Biogeochemistry of paddy soils. Geoderma 2010, 157, 1–14. [Google Scholar] [CrossRef]
- Zhang, Z.; Furman, A. Soil redox dynamics under dynamic hydrologic regimes—A review. Sci. Total Environ. 2021, 763, 143026. [Google Scholar] [CrossRef]
- Kukkadapu, R.K.; Zachara, J.M.; Fredrickson, J.K.; Kennedy, D.W. Biotransformation of two-line silica-ferrihydrite by a dissimilatory Fe (III)-reducing bacterium: Formation of carbonate green rust in the presence of phosphate. Geochim. Cosmochim. Acta 2004, 68, 2799–2814. [Google Scholar] [CrossRef]
- Jia, R.; Qu, Z.; You, P.; Qu, D. Effect of biochar on photosynthetic microorganism growth and iron cycling in paddy soil under different phosphate levels. Sci. Total Environ. 2018, 612, 223–230. [Google Scholar] [CrossRef]
- Li, Y.C.; Yu, S.; Strong, J.; Wang, H.L. Are the biogeochemical cycles of carbon, nitrogen, sulfur, and phosphorus driven by the “Fe-III-Fe-II redox wheel” in dynamic redox environments? J. Soils Sediments 2012, 12, 683–693. [Google Scholar] [CrossRef]
- Wang, C.; Thielemann, L.; Dippold, M.A.; Guggenberger, G.; Kuzyakov, Y.; Banfield, C.C.; Ge, T.; Guenther, S.; Bork, P.; Horn, M.A.; et al. Can the reductive dissolution of ferric iron in paddy soils compensate phosphorus limitation of rice plants and microorganisms? Soil Biol. Biochem. 2022, 168, 108653. [Google Scholar] [CrossRef]
- Schulz, K.; Wisawapipat, W.; Barmettler, K.; Grigg, A.R.C.; Kubeneck, L.J.; Notini, L.; ThomasArrigo, L.K.; Kretzschmar, R. Iron Oxyhydroxide Transformation in a Flooded Rice Paddy Field and the Effect of Adsorbed Phosphate. Environ. Sci. Technol. 2024, 58, 10601–10610. [Google Scholar] [CrossRef] [PubMed]
- Gérard, F. Clay minerals, iron/aluminum oxides, and their contribution to phosphate sorption in soils—A myth revisited. Geoderma 2016, 262, 213–226. [Google Scholar] [CrossRef]
- Koch, M.; Kruse, J.; Eichler-Löbermann, B.; Zimmer, D.; Willbold, S.; Leinweber, P.; Siebers, N. Phosphorus stocks and speciation in soil profiles of a long-term fertilizer experiment: Evidence from sequential fractionation, P K-edge XANES, and 31P NMR spectroscopy. Geoderma 2018, 316, 115–126. [Google Scholar] [CrossRef]
- Willett, I. The reductive dissolution of phosphated ferrihydrite and strengite. Soil Res. 1985, 23, 237–244. [Google Scholar] [CrossRef]
- Scalenghe, R.; Edwards, A.C.; Ajmone Marsan, F.; Barberis, E. The effect of reducing conditions on the solubility of phosphorus in a diverse range of European agricultural soils. Eur. J. Soil Sci. 2002, 53, 439–447. [Google Scholar] [CrossRef]
- Martinengo, S.; Schiavon, M.; Santoro, V.; Said-Pullicino, D.; Romani, M.; Miniotti, E.F.; Celi, L.; Martin, M. Assessing phosphorus availability in paddy soils: The importance of integrating soil tests and plant responses. Biol. Fertil. Soils 2023, 59, 391–405. [Google Scholar] [CrossRef]
- Nanzyo, M.; Onodera, H.; Hasegawa, E.; Ito, K.; Kanno, H. Formation and dissolution of vivianite in paddy field soil. Soil Sci. Soc. Am. J. 2013, 77, 1452–1459. [Google Scholar] [CrossRef]
- Arai, Y.; Sparks, D.L. Phosphate Reaction Dynamics in Soils and Soil Components: A Multiscale Approach. Adv. Agron. 2007, 94, 135–179. [Google Scholar]
- Wisawapipat, W.; Charoensri, K.; Runglerttrakoolchai, J. Solid-phase speciation and solubility of phosphorus in an acid sulfate paddy soil during soil reduction and reoxidation as affected by oil palm ash and biochar. J. Agric. Food Chem. 2017, 65, 704–710. [Google Scholar] [CrossRef]
- Santoro, V.; Martin, M.; Persson, P.; Lerda, C.; Said-Pullicino, D.; Magnacca, G.; Celi, L. Inorganic and organic P retention by coprecipitation during ferrous iron oxidation. Geoderma 2019, 348, 168–180. [Google Scholar] [CrossRef]
- Heiberg, L.; Pedersen, T.V.; Jensen, H.S.; Kjaergaard, C.; Hansen, H.C.B. A Comparative study of phosphate sorption in lowland soils under oxic and anoxic conditions. J. Environ. Qual. 2010, 39, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Liesack, W.; Schnell, S.; Revsbech, P. Microbiology of flooded rice paddies. FEMS Microbiol. Rev. 2000, 24, 625–645. [Google Scholar] [CrossRef] [PubMed]
- Ratering, S.; Schnell, S. Localization of iron-reducing activity in paddy soil by profile studies. Biogeochemistry 2000, 48, 341–365. [Google Scholar] [CrossRef]
- Qi, Y.H.; Cheng, W.; Nan, X.Y.; Yang, F.; Li, J.; Li, D.C.; Lundstrom, C.C.; Yu, H.M.; Zhang, G.L.; Huang, F. Iron stable isotopes in bulk soil and sequential extracted fractions trace Fe redox cycling in paddy soils. J. Agric. Food Chem. 2020, 68, 8143–8150. [Google Scholar] [CrossRef]
- Xu, M.; Gao, P.; Yang, Z.; Su, L.; Yang, G.; Zhang, X.; Ma, J.; Peng, H.; Xiao, Y. Biochar impacts on phosphorus cycling in rice ecosystem. Chemosphere 2019, 225, 311–319. [Google Scholar] [CrossRef]
- Bagheri Novair, S.; Cheraghi, M.; Faramarzi, F.; Asgari Lajayer, B.; Senapathi, V.; Astatkie, T.; Price, G.W. Reviewing the role of biochar in paddy soils: An agricultural and environmental perspective. Ecotoxicol. Environ. Saf. 2023, 263, 115228. [Google Scholar] [CrossRef]
- Zhang, L.; Chang, L.; Liu, H.; de Jesús Puy Alquiza, M.; Li, Y. Biochar application to soils can regulate soil phosphorus availability: A review. Biochar 2025, 7, 13. [Google Scholar] [CrossRef]
- Shi, Y.; Yu, Y.; Chang, E.; Wang, R.; Hong, Z.; Cui, J.; Zhang, F.; Jiang, J.; Xu, R. Effect of biochar incorporation on phosphorus supplementation and availability in soil: A review. J. Soils Sediments 2023, 23, 672–686. [Google Scholar] [CrossRef]
- Yang, L.; Wu, Y.; Wang, Y.; An, W.; Jin, J.; Sun, K.; Wang, X. Effects of biochar addition on the abundance, speciation, availability, and leaching loss of soil phosphorus. Sci. Total Environ. 2021, 758, 143657. [Google Scholar] [CrossRef]
- Chintala, R.; Mollinedo, J.; Schumacher, T.E.; Malo, D.D.; Julson, J.L. Effect of biochar on chemical properties of acidic soil. Arch. Agron. Soil Sci. 2014, 60, 393–404. [Google Scholar] [CrossRef]
- Haynes, R.J. Effects of liming on phosphate availability in acid soils. Plant Soil 1982, 68, 289–308. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.; Xiang, J.; Xiong, J.; Wang, Y.; Wang, Z.; Zhang, Y. Effect of rice-straw biochar application on the acquisition of rhizosphere phosphorus in acidified paddy soil. Agronomy 2022, 12, 1556. [Google Scholar] [CrossRef]
- Jackson, M.L.; Lim, C.H.; Zelazny, L.W. Oxides, hydroxides, and aluminosilicates. In Methods of Soil Analysis. Part 1. Physical and Mineralogical Methods, 2nd ed.; Klute, A., Ed.; Soil Science Society of America: Madison, WI, USA, 1986; pp. 101–150. [Google Scholar]
- Lovley, D.R.; Phillips, E.J.P. Requirement for a microbial consortium to completely oxidize glucose in Fe(III)-reducing Sediments. Appl. Environ. Microbiol. 1989, 55, 3234–3236. [Google Scholar] [CrossRef] [PubMed]
- Nagul, E.A.; McKelvie, I.D.; Worsfold, P.; Kolev, S.D. The molybdenum blue reaction for the determination of orthophosphate revisited: Opening the black box. Anal. Chim. Acta 2015, 890, 60–82. [Google Scholar] [CrossRef]
- Hedley, M.J.; Stewart, J.W.B.; Chauhan, B.S. Changes in inorganic and organic soil phosphorus fractions induced by cultivation practices and by laboratory incubations. Soil Sci. Soc. Am. J. 1982, 46, 970–976. [Google Scholar] [CrossRef]
- Bolan, N.; Sarmah, A.K.; Bordoloi, S.; Bolan, S.; Padhye, L.P.; Van Zwieten, L.; Sooriyakumar, P.; Khan, B.A.; Ahmad, M.; Solaiman, Z.M.; et al. Soil acidification and the liming potential of biochar. Environ. Pollut. 2023, 317, 120632. [Google Scholar] [CrossRef]
- Joseph, S.; Husson, O.; Graber, E.R.; Van Zwieten, L.; Taherymoosavi, S.; Thomas, T.; Nielsen, S.; Ye, J.; Pan, G.; Chia, C.; et al. The electrochemical properties of biochars and how they affect soil redox properties and processes. Agronomy 2015, 5, 322–340. [Google Scholar] [CrossRef]
- Graber, E.; Tsechansky, L.; Lew, B.; Cohen, E. Reducing capacity of water extracts of biochars and their solubilization of soil Mn and Fe. Eur. J. Soil Sci. 2014, 65, 162–172. [Google Scholar] [CrossRef]
- Jia, R.; Li, L.; Qu, D.; Mi, N. Enhanced iron(III) reduction following amendment of paddy soils with biochar and glucose modified biochar. Environ. Sci. Pollut. Res. 2018, 25, 91–103. [Google Scholar] [CrossRef]
- Jia, R.; Fan, F.; Li, L.; Qu, D. Temporal response of bacterial community associated Fe(III) reduction to initial pH shift of paddy soils. Agronomy 2022, 12, 1304. [Google Scholar] [CrossRef]
- Jia, R.; Li, L.-N.; Qu, D. pH shift-mediated dehydrogenation and hydrogen production are responsible for microbial iron(III) reduction in submerged paddy soils. J. Soils Sediments 2015, 15, 1178–1190. [Google Scholar] [CrossRef]
- Munch, J.C.; Hillebrand, T.; Ottow, J.C.G. Transformations in the Feo/Fed ratio of pedogenic iron oxides affected by iron-reducing bacteria. Can. J. Soil Sci. 1978, 58, 475–486. [Google Scholar] [CrossRef]
- Brennan, E.W.; Lindsay, W.L. Reduction and oxidation effect on the solubility and transformation of iron oxides. Soil Sci. Soc. Am. J. 1998, 62, 930–937. [Google Scholar] [CrossRef]
- Chen, A.; Arai, Y. A review of the reactivity of phosphatase controlled by clays and clay minerals: Implications for understanding phosphorus mineralization in soils. Clay. Clay Miner. 2023, 71, 119–142. [Google Scholar] [CrossRef]
- Barrow, N.J. The effects of pH on phosphate uptake from the soil. Plant Soil 2017, 410, 401–410. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Composition | Soil | Biochar |
|---|---|---|
| Pseudo-total content (g kg−1) | ||
| P | 0.49 ± 0.01 | 3.41 ± 0.16 |
| Al | 67.1 ± 1.8 | 0.76 ± 0.03 |
| Fe | 40.9 ± 1.0 | 0.44 ± 0.04 |
| Ca | 0.87 ± 0.02 | 9.86 ± 0.60 |
| Mg | 2.23 ± 0.05 | 3.72 ± 0.16 |
| Free oxide 1 (g kg−1) | ||
| Fed | 30.65 ± 0.95 | 0.32 ± 0.15 |
| Ald | 5.87 ± 0.17 | 0.17 ± 0.01 |
| Amorphous oxide 2 (g kg−1) | ||
| Feo | 3.16 ± 0.33 | 0.27 ± 0.02 |
| Alo | 1.46 ± 0.01 | 0.08 ± 0.01 |
| Material | Element (%) | ||||
|---|---|---|---|---|---|
| C | H | O | N | S | |
| Rice straw | 39.4 | 5.8 | 39.1 | 0.9 | 0.1 |
| Rice–straw biochar | 56.6 | 1.9 | 5.8 | 1.2 | 0.3 |
| P Fraction (mg kg−1) | Soil | Biochar |
|---|---|---|
| Water-P (soluble P) | 4.4 ± 0.6 | 1233 ± 12 |
| NaHCO3-P (exchangeable P) | 43.8 ± 1.2 | 1054 ± 24 |
| NaOH-P (Fe/Al-bound P) | 183.4 ± 2.5 | 22.8 ± 1.2 |
| HCl-P (Ca-bound P) | 12.3 ± 3.2 | 694.6 ± 6.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, Y.-R.; Wang, S.-L. Effects of Biochar on the Temporal Dynamics and Vertical Distribution of Iron and Phosphorus Under Soil Submergence. Agronomy 2025, 15, 1394. https://doi.org/10.3390/agronomy15061394
Lai Y-R, Wang S-L. Effects of Biochar on the Temporal Dynamics and Vertical Distribution of Iron and Phosphorus Under Soil Submergence. Agronomy. 2025; 15(6):1394. https://doi.org/10.3390/agronomy15061394
Chicago/Turabian StyleLai, Ying-Ren, and Shan-Li Wang. 2025. "Effects of Biochar on the Temporal Dynamics and Vertical Distribution of Iron and Phosphorus Under Soil Submergence" Agronomy 15, no. 6: 1394. https://doi.org/10.3390/agronomy15061394
APA StyleLai, Y.-R., & Wang, S.-L. (2025). Effects of Biochar on the Temporal Dynamics and Vertical Distribution of Iron and Phosphorus Under Soil Submergence. Agronomy, 15(6), 1394. https://doi.org/10.3390/agronomy15061394