

Seed Dormancy Class and Germination Characteristics of Berberis amurensis var. latifolia Nakai, Native to Korea

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Morpho-Anatomical Characteristics of Seeds
2.3. Seed Disinfection and Setting
2.4. Water Imbibition Test
2.5. A Move-along Experiment
2.6. Effect of Cold Stratification on Germination
2.7. Effect of GA3 on Germination
2.8. Effect of Light Conditions on Seed Germination
2.9. Phenology of Embryo Growth, Germination, and Seedling Emergence under Natural Environmental Conditions
2.9.1. Embryo Growth
2.9.2. Germination
2.9.3. Seedling Emergence
2.10. Statistical Analyses
3. Results
3.1. Investigation of Morpho-Anatomical Characteristics of Seeds
3.2. Water Imbibition Test
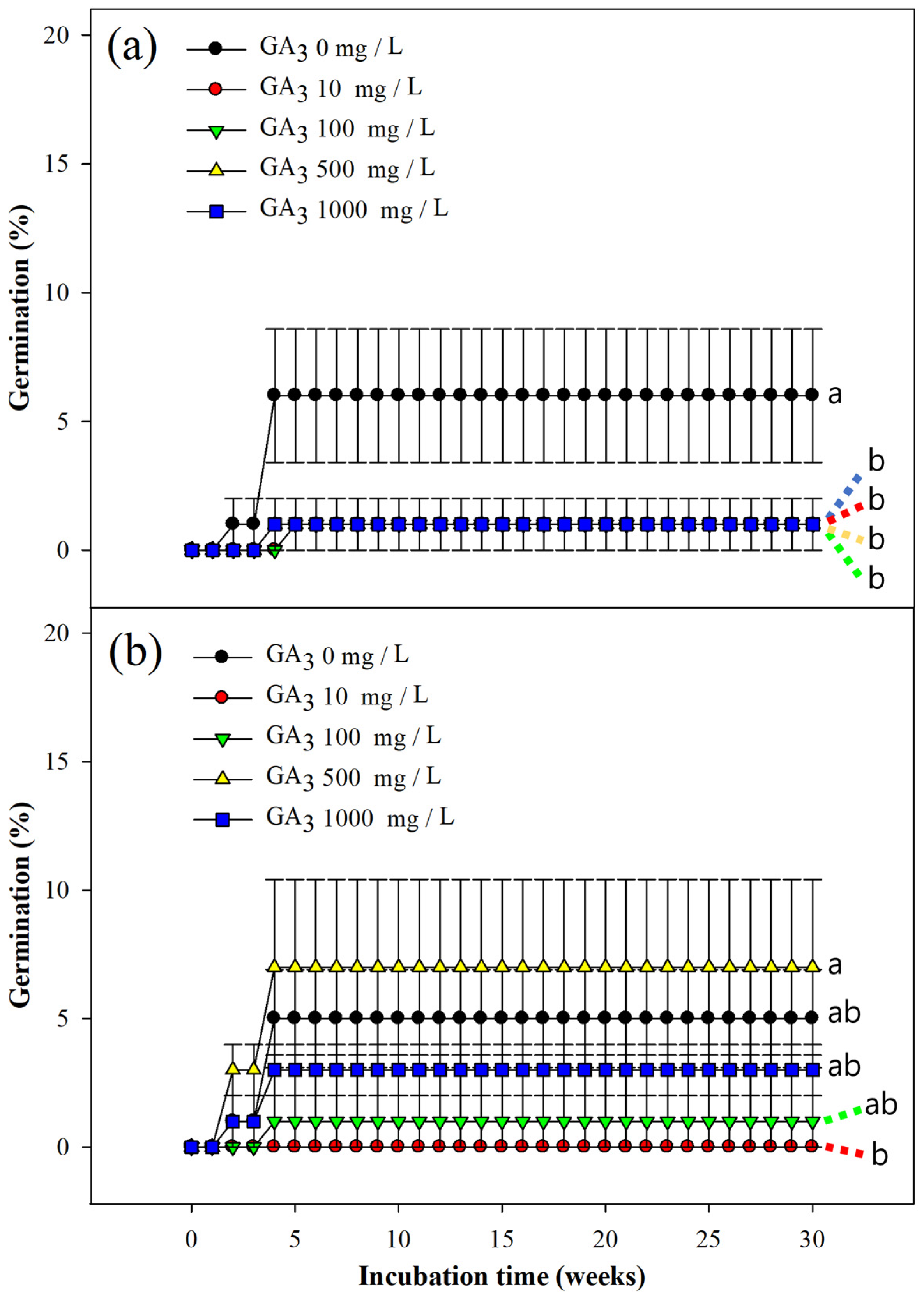
3.3. Effect of GA3 Treatment on Seed Germination
3.4. Effect of Cold Stratification Experiment on Seed Germination
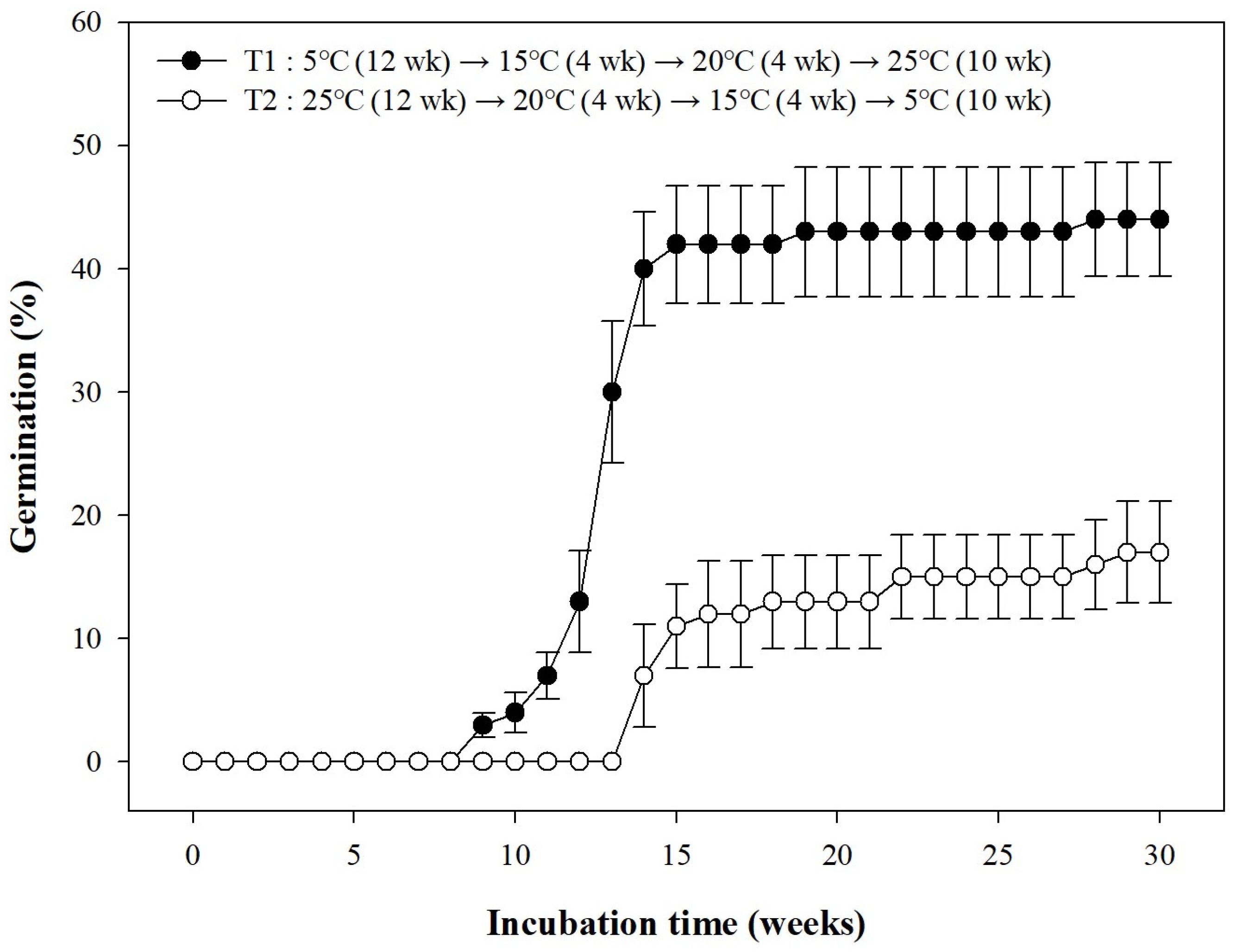
3.5. Seed Germination Based on Temperature Conditions: A Move-along Experiment
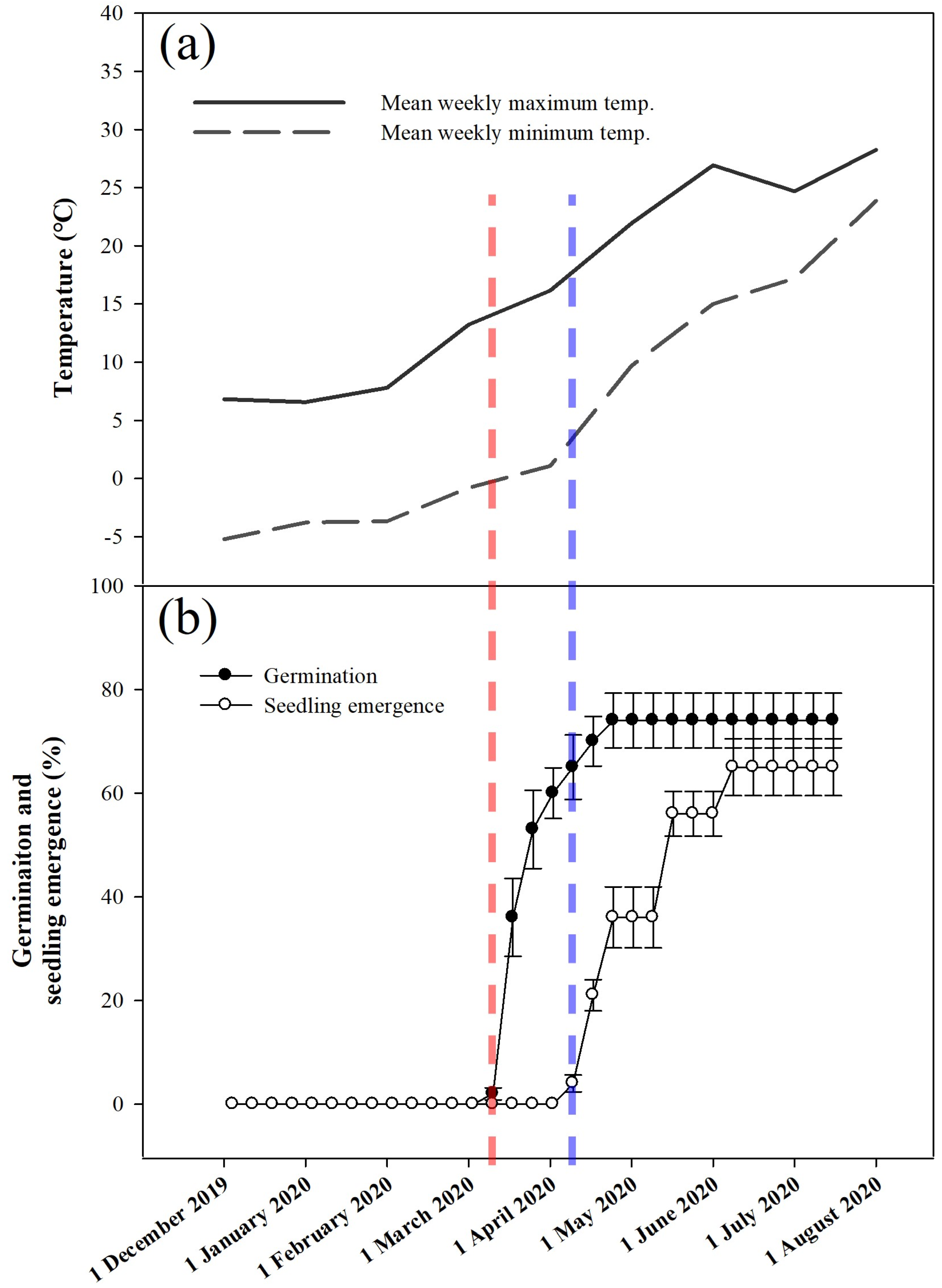
3.6. Phenology of Embryo Growth, Germination, and Seedling Emergence
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Willis, J.C. A Dictionary of the Flowering Plants & Ferns, 8th ed.; Cambridge University Press: Cambridge, UK, 1973. [Google Scholar]
- Kafi, M.; Balandary, A.; Rashed-Mohasel, M.H.; Koochaki, A.; Molafilabi, A. Berberis: Production and Processing; Zaban va Adab Press: Tehran, Iran, 2002; pp. 50–60. [Google Scholar]
- Rounsaville, T.J.; Ranney, T.G. Ploidy levels and genome sizes of Berberis L. and Mahonia Nutt. species, hybrids, and cultivars. HortScience 2010, 45, 1029–1033. [Google Scholar] [CrossRef]
- Ohwi, J. Flora of Japan; Smithsonian Institution: Washington, DC, USA, 1984. [Google Scholar]
- Jin, L.N.; Sung, C.C.; Hui, K. A morphometric study of Berberis amurensis and B. koreana. Bull. Seoul Natl. Univ. Arbor. 2004, 24, 83–93. [Google Scholar]
- Hyun, C.W.; Kim, Y.D. Morphological variation of Berberis amurensis complex. Korean J. Plant Taxon. 2008, 38, 93–109. [Google Scholar] [CrossRef]
- Majumder, P.; Sucharitas, S. 1,4-Bis-(2′-hydroxy-5′-methyl phenyl)-butan -1,4-dione- a biogenetically rare type of phenolic of Berberis coriaria. Phytochemistry 1978, 17, 1439–1440. [Google Scholar] [CrossRef]
- Masood, M.; Tiwar, K.P. 2,5-Bis-(2′-methoxy-5′- methyl phenyl)-furan, a rare type of compound from Berberis umbellata. Phytochemistry 1981, 20, 295–296. [Google Scholar] [CrossRef]
- Hussaini, F.A.; Shoeb, A. Isoquinoline derived alkaloids from Berberis chitria. Phytochemistry 1985, 24, 633. [Google Scholar] [CrossRef]
- Stadler, R.; Loeffler, S.; Cassels, B.K.; Zenk, M.H. Bisbenzyl isoquinoline biochemistry in Berberis stolonifera cell cultures. Phytochemistry 1988, 27, 2557–2565. [Google Scholar] [CrossRef]
- Saied, S.; Begum, S. Phytochemical studies of Berberis vulgaris. Chem. Nat. Compd. 2004, 40, 137–140. [Google Scholar] [CrossRef]
- Tomosaka, H.; Chin, Y.W.; Salim, A.A.; Keller, W.J.; Chai, H.; Kinghorn, A.D. Antioxidant and cytoprotective compounds from Berberis vulgaris (Barberry). Phytother. Res. 2008, 22, 979–981. [Google Scholar] [CrossRef]
- Cabezas, N.J.; Urzua, A.M.; Niemeyer, H.M. Translocation of isoquinoline alkaloids to the hemiparasite, from its host, Berberis montana. Biochem. Syst. Ecol. 2009, 37, 225–227. [Google Scholar] [CrossRef]
- Papiya, M.M.; Saumya, D.; Sanjita, D.; Manas, K.D. Cytotoxic activity of methanolic extracts of Berberis aristata DC and Hemidesmus indicus R.Br. in MCF7 cell line. J. Curr. Pharm. Res. 2010, 1, 12–15. [Google Scholar]
- Mokhber-Dezfuli, N.; Saeidnia, S.; Gohari, A.R.; Kurepaz-Mahmoodabadi, M. Phytochemistry and pharmacology of Berberis species. Pharmacogn. Rev. 2014, 8, 8. [Google Scholar] [PubMed]
- Salehi, B.; Selamoglu, Z.; Sener, B.; Kilic, M.; Kumar Jugran, A.; de Tommasi, N.; Sinisgalli, C.; Milella, L.; Rajkovic, J.; Mo-rais-Braga, M.F.B.; et al. Berberis plants—Drifting from farm to food applications, phytotherapy, and phytopharmacology. Foods 2019, 8, 522. [Google Scholar] [CrossRef] [PubMed]
- Kolar, D.; Wimmerova, L.; Kadek, R. Acetylcholinesterase and butyrylcholinesterase inhibitory activities of Berberis vulgaris. Phytopharmacology 2010, 1, 7–11. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography and Evolution of Dormancy and Germination, 2nd ed.; Elsevier Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Lang, G.A. Dormancy: A new universal terminology. HortScience 1987, 22, 817–820. [Google Scholar] [CrossRef]
- Wang, J.H.; Baskin, C.C.; Chen, W.; Du, G.Z. Variation in seed germination between populations of five sub-alpine woody species from eastern Qinghai-Tibet Plateau following dry storage at low temperatures. Ecol. Res. 2010, 25, 195–203. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Bahuguna, V.K.; Rawart, M.M.S.; Joshi, S.R. Preliminary studies on seed germination behaviour of Berberis lycium Royle—An important shrub for reclamation of wastelands in the Himalaya. Indian For. 1988, 114, 181–183. [Google Scholar]
- Rudolf, P.O.; Buckeye, A.L. Horse chestnut. Seeds of Woody Plants in the United States. Agric. Handb. 1974, 450, 195–199. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Mimicking the natural thermal environments experienced by seeds to break physiological dormancy to enhance seed testing and seedling production. Seed Sci. Technol. 2022, 50, 21–29. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, S.G.; Lee, H.; Na, C.S.; Lee, D.H. The Dormancy Types and Germination Characteristics of the Seeds of Berberis koreana Palibin, an Endemic Species of Korea. Horticulturae 2023, 9, 547. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M.; Meyer, S.E. Seed dormancy in the Colorado Plateau shrub Mahonia fremontii (Berberidaceae) and its ecological and evolutionary implications. Southwest. Nat. 1993, 38, 91–99. [Google Scholar] [CrossRef]
- Deb, C.R.; Sangtam, T.L.; Jamir, N.S. Seed biology of Berberis manipurana Ahrendt: A threatened natural dye yielding plant. Am. J. Plant Sci. 2017, 8, 1285–1295. [Google Scholar]
- Larsen, S.U.; Eriksen, E.N. Delayed release of primary dormancy and induction of secondary dormancy in seeds of woody taxa caused by temperature alternations. In Proceedings of the XXVI International Horticultural Congress: Nursery Crops; Development, Evaluation, Production and Use, Toronto, ON, Canada, 11 August 2002; Volume 630, pp. 91–100. [Google Scholar] [CrossRef]
- Lee, S.Y.; Rhie, Y.H.; Kim, K.S. Non-deep simple morphophysiological dormancy in seeds of Thalictrum rochebrunianum, an endemic perennial herb in the Korean Peninsula. Hortic. Environ. Biotechnol. 2015, 56, 366–375. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. When breaking seed dormancy is a problem try a move-along experiment. Nativ. Plants J. 2003, 4, 17–21. [Google Scholar] [CrossRef]
- Piotto, B.; De Noi, A. Seed propagation of Mediterranean trees and shrubs [Manuali e linee guida]. Seed Sci. Res. 2004, 14, 91–92. [Google Scholar]
- Arena, M.E.; Martinez-Pastur, G. Seed propagation in Berberis buxifolia Lam. Phyton 1994, 56, 59–63. [Google Scholar]
- Figueroa, J.A.; Armesto, J.J.; Hernandez, J.F. Estrategias de germinacion y latencia de semillas en especies del bosque templado de Chiloe, Chile. Rev. Chil. Hist. Nat. 1996, 69, 243–251. [Google Scholar]
- Figueroa, J.A.; Armesto, J.J. Community-wide germination strategies in a temperate rainforest of southern Chile: Ecological and evolutionary correlates. Aust. J. Bot. 2001, 49, 411–425. [Google Scholar] [CrossRef]
- Thakur, A.; Thakur, P.S.; Mehta, R. Studies on germination, viability and vigour in Indian barberry (Berberis aristata DC.)—An endangered medicinal plant species of western Himalayas. Indian For. 2006, 132, 485–492. [Google Scholar]
- Wang, J.H.; Du, G.Z.; Cui, X.L.; Zheng, X.F.; Qi, W. Germination characteristics of 61 common woody species from the eastern Qinghai-Tibet Plateau of China and their life history correlates. J. Plant Ecol. 2009, 33, 171–179. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Collection Date | Altitude (m) | Geographical Coordinates |
|---|---|---|---|
| 10-12, Namyang 1-gil, Seo-myeon, Ulleung-gun, Gyeongsangbuk-do, Republic of Korea | 29 October 2019 | 3 | 37°28′00.46″ N 130°50′22.67″ E |
| Weeks at Treatment Temperatures | |||||||
|---|---|---|---|---|---|---|---|
| 4 Weeks | 4 Weeks | 4 Weeks | 4 Weeks | 4 Weeks | 4 Weeks | ||
| Move along | T1 | 5 °C (winter) | 5 °C (winter) | 5 °C (winter) | 15 °C (early spring) | 20 °C (late spring) | 25 °C (summer) |
| T2 | 25 °C (summer) | 25 °C (summer) | 25 °C (summer) | 20 °C (early autumn) | 15 °C (early autumn) | 5 °C (winter) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-H.; Lee, D.-H.; Park, J.-Y.; Kim, H.-M.; Kim, J.-H.; Kim, H.-J.; Che, S.-H.; Na, C.-S.; Lee, D.-H. Seed Dormancy Class and Germination Characteristics of Berberis amurensis var. latifolia Nakai, Native to Korea. Agronomy 2024, 14, 956. https://doi.org/10.3390/agronomy14050956
Kim D-H, Lee D-H, Park J-Y, Kim H-M, Kim J-H, Kim H-J, Che S-H, Na C-S, Lee D-H. Seed Dormancy Class and Germination Characteristics of Berberis amurensis var. latifolia Nakai, Native to Korea. Agronomy. 2024; 14(5):956. https://doi.org/10.3390/agronomy14050956
Chicago/Turabian StyleKim, Do-Hyun, Da-Hyun Lee, Ji-Yoon Park, Hyeon-Min Kim, Jun-Hyeok Kim, Hoi-Jin Kim, Sang-Hoon Che, Chae-Sun Na, and Do-Hyung Lee. 2024. "Seed Dormancy Class and Germination Characteristics of Berberis amurensis var. latifolia Nakai, Native to Korea" Agronomy 14, no. 5: 956. https://doi.org/10.3390/agronomy14050956
APA StyleKim, D.-H., Lee, D.-H., Park, J.-Y., Kim, H.-M., Kim, J.-H., Kim, H.-J., Che, S.-H., Na, C.-S., & Lee, D.-H. (2024). Seed Dormancy Class and Germination Characteristics of Berberis amurensis var. latifolia Nakai, Native to Korea. Agronomy, 14(5), 956. https://doi.org/10.3390/agronomy14050956